
Distribution and Molecular Identification of Culex pipiens and Culex tritaeniorhynchus as Potential Vectors of Rift Valley Fever Virus in Jazan, Saudi Arabia

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Method
2.1. Study Area
2.2. Collection of Adult Mosquito Samples
2.3. Relative Abundance
2.4. Pattern of Occurrence
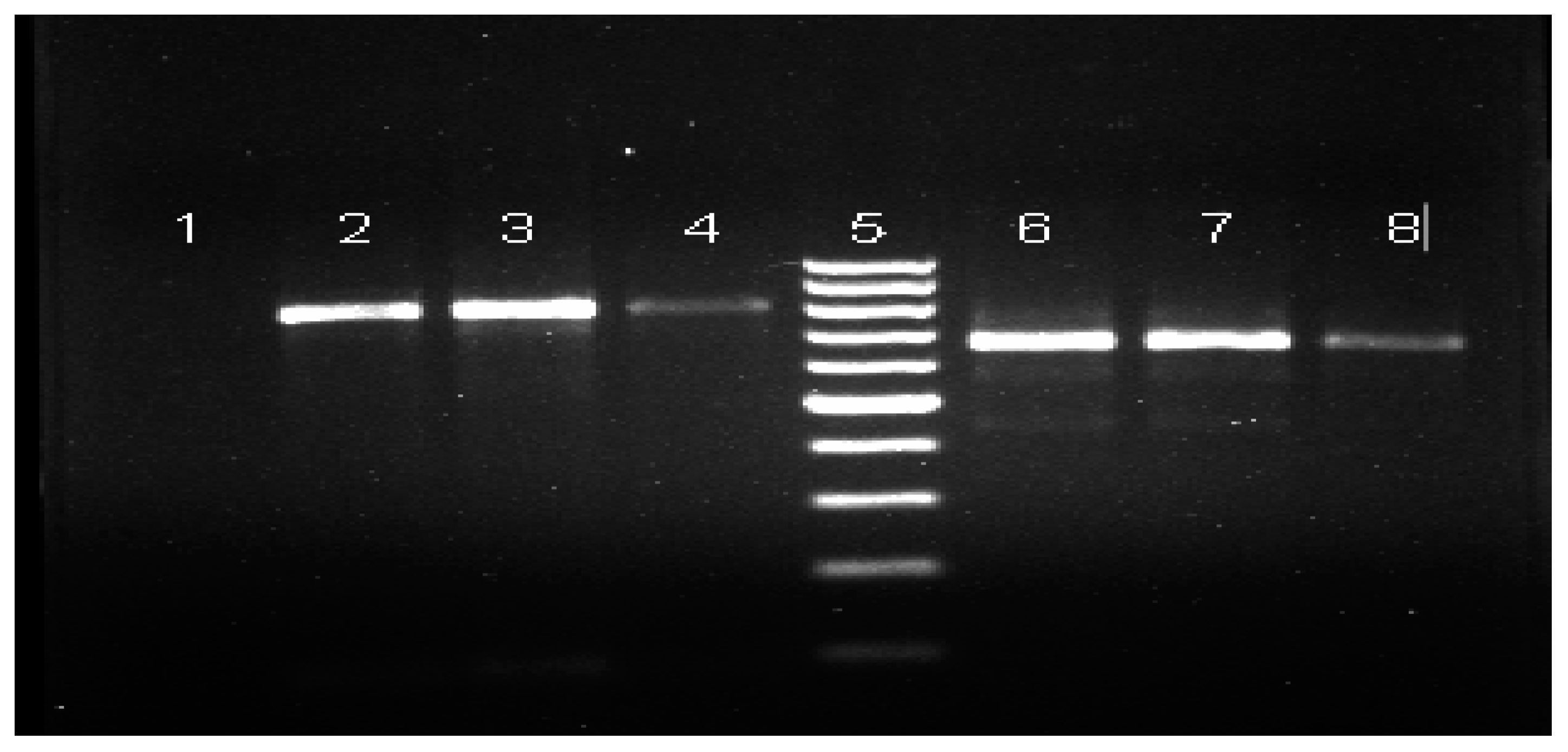
2.5. Molecular Confirmation of Mosquito Identification
2.5.1. DNA Extraction
2.5.2. Amplification of Mosquito COI Fragment
2.6. Sequencing and Data Analysis
2.7. Detection of RVFV in Mosquitoes
2.7.1. RNA Isolation
2.7.2. Reverse Transcriptase Polymerase Chain Reaction
2.7.3. Nested-PCR
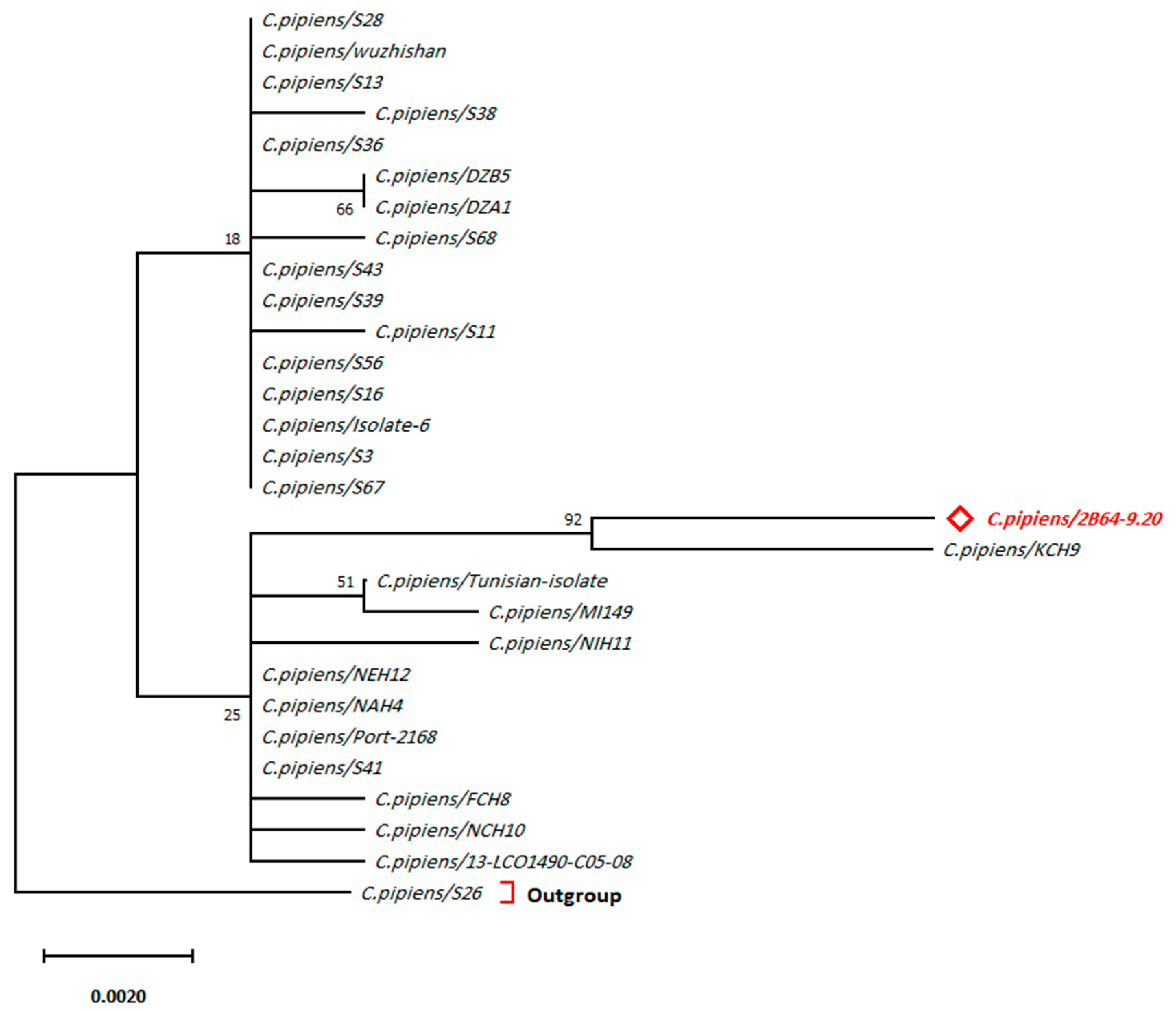
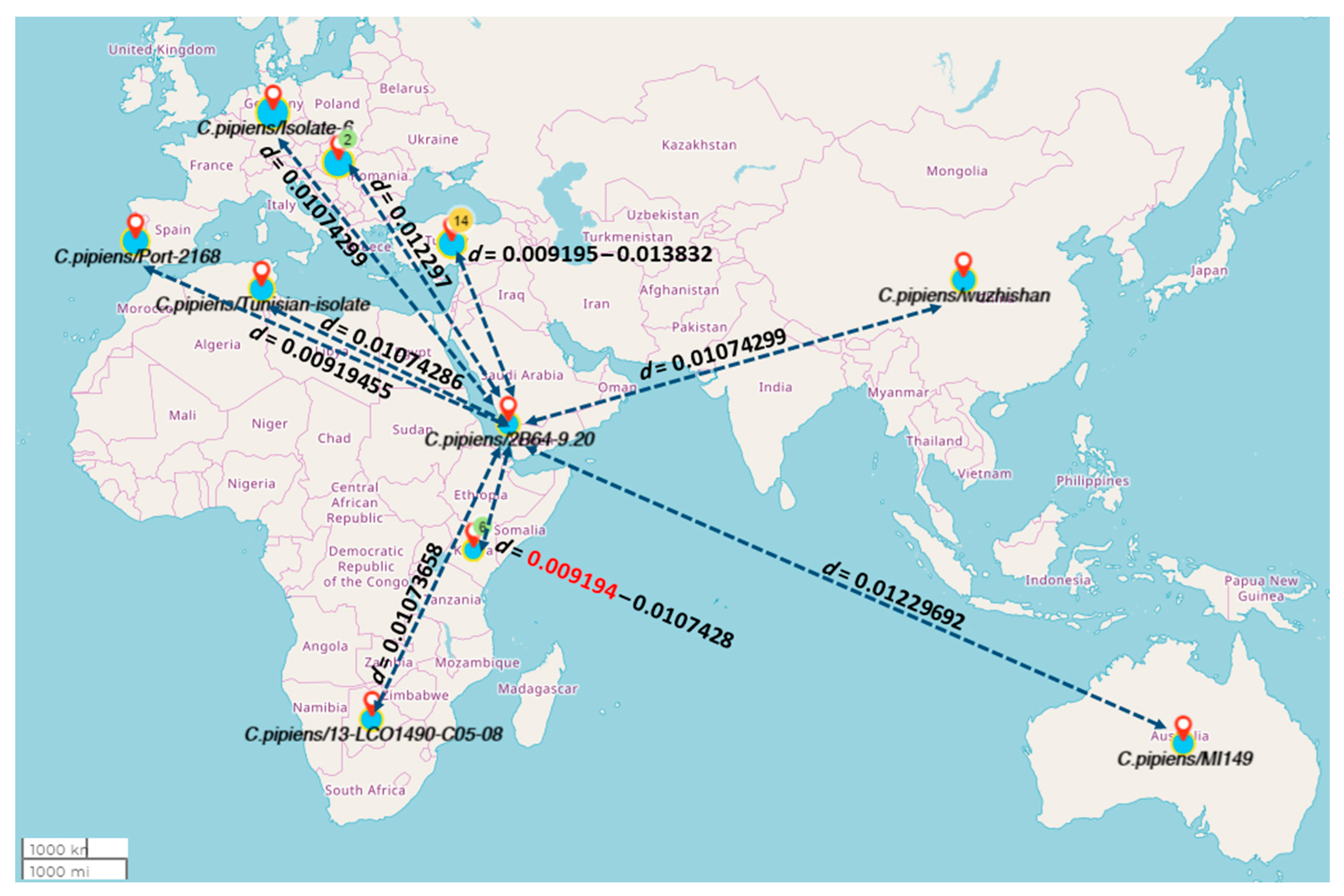
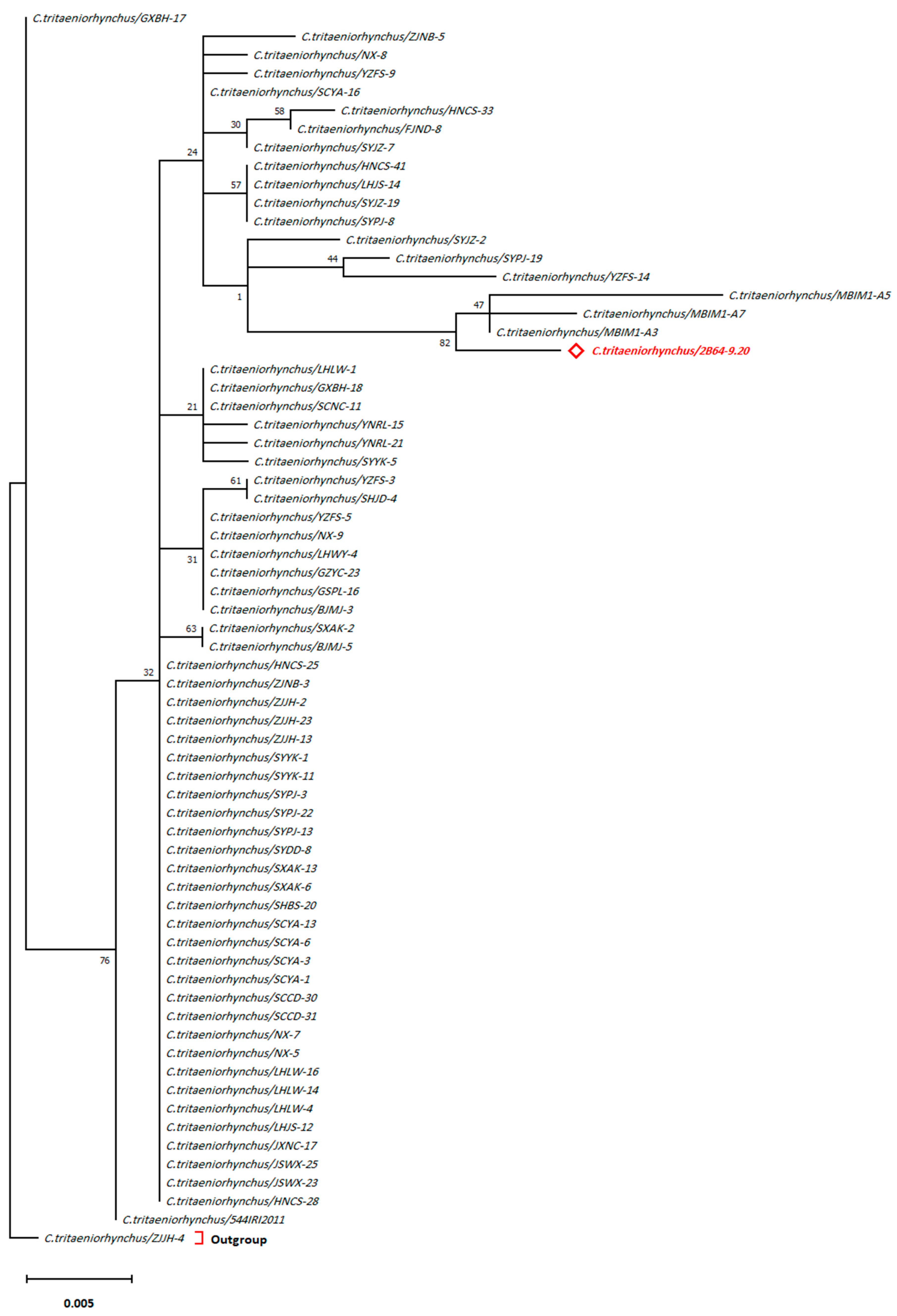
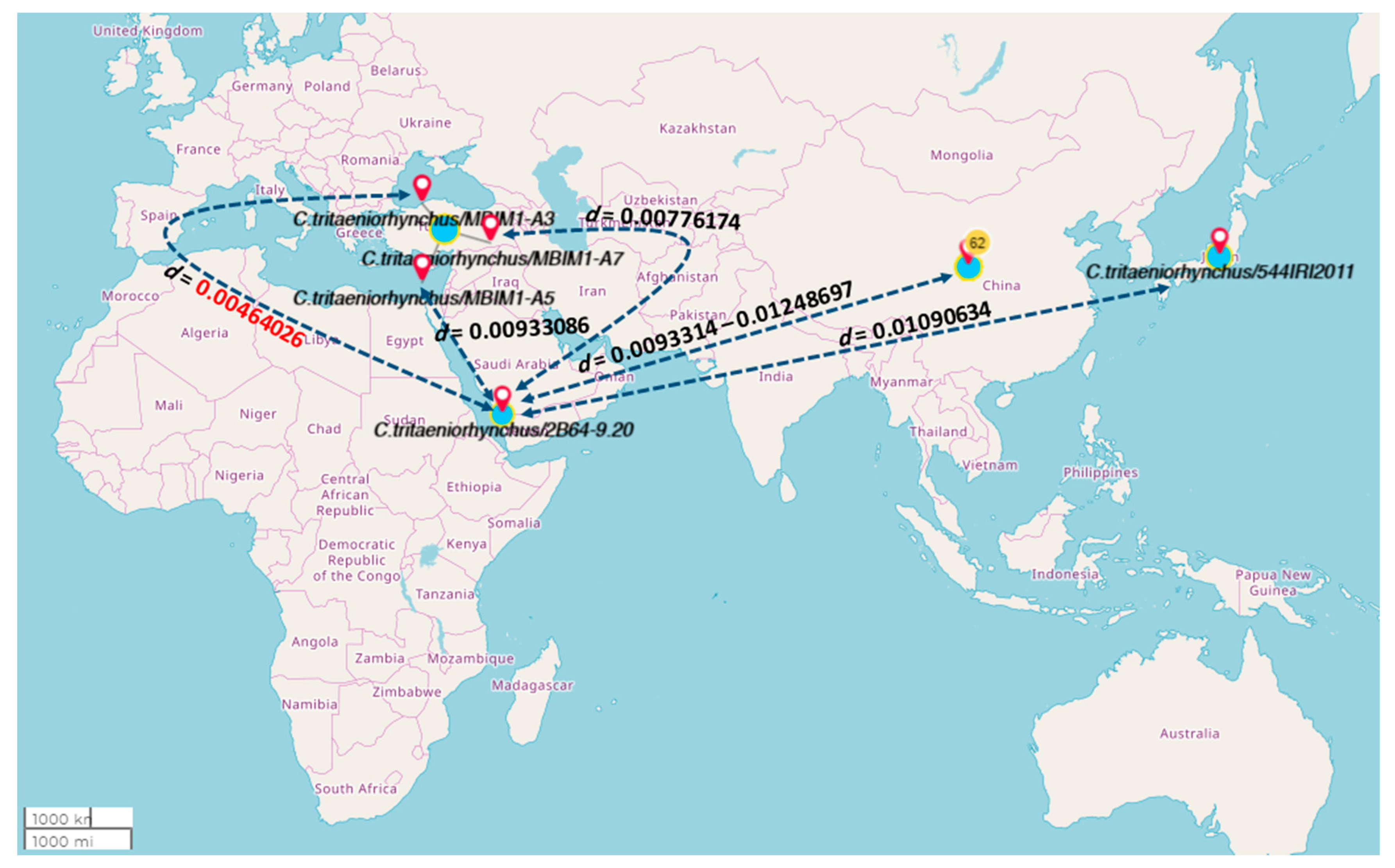
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pepin, M.; Bouloy, M.; Bird, B.H.; Kemp, A.; Paweska, J. Rift Valley Fever Virus (Bunyaviridae: Phlebovirus): An Update on Pathogenesis, Molecular Epidemiology, Vectors, Diagnostics and Prevention. Vet. Res. 2010, 41, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumley, S.; Horton, D.L.; Hernandez-Triana, L.L.; Johnson, N.; Fooks, A.R.; Hewson, R. Rift Valley Fever Virus: Strategies for Maintenance, Survival and Vertical Transmission in Mosquitoes. J. Gen. Virol. 2017, 98, 875–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogrefe, N.; Reindl, S.; Günther, S.; Rosenthal, M. Structure of a Functional Cap-Binding Domain in Rift Valley Fever Virus L Protein. PLoS Pathog. 2019, 15, e1007829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linthicum, K.J.; Britch, S.C.; Anyamba, A. Rift Valley Fever: An Emerging Mosquito-Borne Disease. Annu. Rev. Entomol. 2016, 61, 395–415. [Google Scholar] [CrossRef]
- Daubney, R.; Hudson, J. Enzootic Hepatitis or Rift Valley Fever. An Un-Described Virus Disease of Sheep, Cattle and Man from East Africa. J. Pathol. Bacteriol. 1931, 34, 545–579. [Google Scholar] [CrossRef]
- Al-Afaleq, A.I.; Hussein, M.F. The Status of Rift Valley Fever in Animals in Saudi Arabia: A Mini Review. Vector-Borne Zoonotic Dis. 2011, 11, 1513–1520. [Google Scholar] [CrossRef]
- Lichoti, J.K.; Kihara, A.; Oriko, A.A.; Okutoyi, L.A.; Wauna, J.O.; Tchouassi, D.P.; Tigoi, C.C.; Kemp, S.; Sang, R.; Mbabu, R.M. Detection of Rift Valley Fever Virus Interepidemic Activity in Some Hotspot Areas of Kenya by Sentinel Animal Surveillance, 2009–2012. Vet. Med. Int. 2014, 2014, 379010. [Google Scholar] [CrossRef] [Green Version]
- Manore, C.; Beechler, B. Inter-epidemic and Between-season Persistence of Rift Valley Fever: Vertical Transmission or Cryptic Cycling? Transbound. Emerg. Dis. 2015, 62, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.; Braack, L.; Guarido, M.; Venter, M.; Gouveia Almeida, A.P. Mosquito Community Composition and Abundance at Contrasting Sites in Northern South Africa, 2014–2017. J. Vector Ecol. 2020, 45, 104–117. [Google Scholar] [CrossRef]
- Rodrigue Simonet, P.N.; Alexandre Michel, N.-N.; Abel, W.; Albert, E.; Martin Hermann, G.; Franziska, S. Diversity and Abundance of Potential Vectors of Rift Valley Fever Virus in the North Region of Cameroon. Insects 2020, 11, 814. [Google Scholar] [CrossRef]
- Mohammed, B.; Yayo, A.; Ajanusi, O.; Lawal, I. Relative Abundance and Molecular Identification of Culex Pipiens Complex (Diptera: Culicidae), in Kura Local Government Area, North-Western Nigeria. Parasite Epidemiol. Control 2021, 14, e00213. [Google Scholar] [CrossRef]
- Nicholas, D.E.; Jacobsen, K.H.; Waters, N.M. Risk Factors Associated with Human Rift Valley Fever Infection: Systematic Review and Meta-analysis. Trop. Med. Int. Health 2014, 19, 1420–1429. [Google Scholar] [CrossRef]
- Alhaj, M.S. An Overview of Sixteen Years Control Program against Rift Valley Fever in Saudi Arabia: A Review Study. Insights Biomed. Res. 2019, 3, 75–80. [Google Scholar]
- Elfadil, A.; Hasab-Allah, K.; Dafa-Allah, O. Factors Associated with Rift Valley Fever in South-West Saudi Arabia. Rev. Sci. Tech. Off. Int. Epizoot. 2006, 25, 1137–1146. [Google Scholar] [CrossRef]
- Alhaj, M.S.; Elmanea, A.A.; Shazali, L.A.; Yousif, M.Q. Surveillance Study on Rift Valley Fever in Jazan Region, Saudi Arabia. Int. J. Adv. Sci. Tech. Res. 2015, 5, 46–57. [Google Scholar]
- Alhaj, M.; Aljidhean, G.; Almanae, A.A.; Abdalla, M.; Muhanna, H. Abundance and Spatial Distribution of Culex and Aedes Mosquitoes as Potential Vectors of Rift Valley Fever in Jazan Region. Int. J. Mosq. Res. 2017, 4, 42–47. [Google Scholar]
- Al-Sheik, A.A. Larval Habitat, Ecology, Seasonal Abundance and Vectorial Role in Malaria Transmission of Anopheles Arabiensis in Jazan Region of Saudi Arabia. J. Egypt Soc. Parasitol. 2011, 41, 615–634. [Google Scholar]
- Jupp, P.G. Mosquitoes of Southern Africa: Culicinae and Toxorhynchitinae; Ekogilde Pubs.: Hartebeespoort, South Africa, 1996. [Google Scholar]
- Aïssaoui, L.; Boudjelida, H. Diversity and Distribution of Culicinae Fauna in Tebessa District (North-East of Algeria). Int. J. Mosq. Res. 2017, 4, 07–12. [Google Scholar]
- Rydzanicz, K.; Lonc, E. Species Composition and Seasonal Dynamics of Mosquito Larvae in the Wrocław, Poland Area. J. Vector Ecol. J. Soc. Vector Ecol. 2003, 28, 255–266. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the Number of Nucleotide Substitutions When There Are Strong Transition-Transversion and G+ C-Content Biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Sall, A.; Macondo, E.; Sène, O.; Diagne, M.; Sylla, R.; Mondo, M.; Girault, L.; Marrama, L.; Spiegel, A.; Diallo, M. Use of Reverse Transcriptase PCR in Early Diagnosis of Rift Valley Fever. Clin. Vaccine Immunol. 2002, 9, 713–715. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.R.; Godsey, M.S.; Crabtree, M.B.; Savage, H.M.; Al-Mazrao, Y.; Al-Jeffri, M.H.; Abdoon, A.-M.M.; Al-Seghayer, S.M.; Al-Shahrani, A.M.; Ksiazek, T.G. Isolation and Genetic Characterization of Rift Valley Fever Virus from Aedes Vexans Arabiensis, Kingdom of Saudi Arabia. Emerg. Infect. Dis. 2002, 8, 1492. [Google Scholar] [CrossRef]
- Jupp, P.; Kemp, A.; Grobbelaar, A.; Leman, P.; Burt, F.; Alahmed, A.; Mujalli, D.A.; Khamees, M.A.; Swanepoel, R. The 2000 Epidemic of Rift Valley Fever in Saudi Arabia: Mosquito Vector Studies. Med. Vet. Entomol. 2002, 16, 245–252. [Google Scholar] [CrossRef]
- Meegan, J.M.; Khalil, G.M.; Hoogstraal, H.; Adham, F.K. Experimental Transmission and Field Isolation Studies Implicating Culex Pipiens as a Vector of Rift Valley Fever Virus in Egypt. Am. J. Trop. Med. Hyg. 1980, 29, 1405–1410. [Google Scholar] [CrossRef] [PubMed]
- Vloet, R.P.; Vogels, C.B.; Koenraadt, C.J.; Pijlman, G.P.; Eiden, M.; Gonzales, J.L.; van Keulen, L.J.; Wichgers Schreur, P.J.; Kortekaas, J. Transmission of Rift Valley Fever Virus from European-Breed Lambs to Culex Pipiens Mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0006145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pachka, H.; Annelise, T.; Alan, K.; Power, T.; Patrick, K.; Véronique, C.; Janusz, P.; Ferran, J. Rift Valley Fever Vector Diversity and Impact of Meteorological and Environmental Factors on Culex Pipiens Dynamics in the Okavango Delta, Botswana. Parasit. Vectors 2016, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ogoma, S.B.; Lweitoijera, D.W.; Ngonyani, H.; Furer, B.; Russell, T.L.; Mukabana, W.R.; Killeen, G.F.; Moore, S.J. Screening Mosquito House Entry Points as a Potential Method for Integrated Control of Endophagic Filariasis, Arbovirus and Malaria Vectors. PLoS Negl. Trop. Dis. 2010, 4, e773. [Google Scholar] [CrossRef] [PubMed]
- Amraoui, F.; Krida, G.; Bouattour, A.; Rhim, A.; Daaboub, J.; Harrat, Z.; Boubidi, S.-C.; Tijane, M.; Sarih, M.; Failloux, A.-B. Culex Pipiens, an Experimental Efficient Vector of West Nile and Rift Valley Fever Viruses in the Maghreb Region. PLoS ONE 2012, 7, e36757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naeem, M.; Alahmed, A.; Kheir, S.; Sallam, M. Spatial Distribution Modeling of Stegomyia Aegypti and Culex Tritaeniorhynchus (Diptera: Culicidae) in Al-Bahah Province, Kingdom of Saudi Arabia. Trop. Biomed. 2016, 33, 295–310. [Google Scholar] [PubMed]
- Sallam, M.F.; Al Ahmed, A.M.; Abdel-Dayem, M.S.; Abdullah, M.A. Ecological Niche Modeling and Land Cover Risk Areas for Rift Valley Fever Vector, Culex Tritaeniorhynchus Giles in Jazan, Saudi Arabia. PLoS ONE 2013, 8, e65786. [Google Scholar] [CrossRef] [PubMed]
- Khater, E.; Sowilem, M.; Sallam, M.; Alahmed, A. Ecology and Habitat Characterization of Mosquitoes in Saudi Arabia. Trop. Biomed. 2013, 30, 409–427. [Google Scholar] [PubMed]
- Alten, B.; Bellini, R.; Caglar, S.; Simsek, F.; Kaynas, S. Species Composition and Seasonal Dynamics of Mosquitoes in the Belek Region of Turkey. J. Vector Ecol. J. Soc. Vector Ecol. 2000, 25, 146–154. [Google Scholar]
- Failloux, A.-B.; Bouattour, A.; Faraj, C.; Gunay, F.; Haddad, N.; Harrat, Z.; Jancheska, E.; Kanani, K.; Kenawy, M.A.; Kota, M. Surveillance of Arthropod-Borne Viruses and Their Vectors in the Mediterranean and Black Sea Regions within the MediLabSecure Network. Curr. Trop. Med. Rep. 2017, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musa, A.A.; Muturi, M.W.; Musyoki, A.M.; Ouso, D.O.; Oundo, J.W.; Makhulu, E.E.; Wambua, L.; Villinger, J.; Jeneby, M.M. Arboviruses and Blood Meal Sources in Zoophilic Mosquitoes at Human-Wildlife Interfaces in Kenya. Vector-Borne Zoonotic Dis. 2020, 20, 444–453. [Google Scholar] [CrossRef]
- Gunay, F.; Alten, B.; Simsek, F.; Aldemir, A.; Linton, Y.-M. Barcoding Turkish Culex Mosquitoes to Facilitate Arbovirus Vector Incrimination Studies Reveals Hidden Diversity and New Potential Vectors. Acta Trop. 2015, 143, 112–120. [Google Scholar] [CrossRef]
- Lõhmus, M.; Lindström, A.; Björklund, M. How Often Do They Meet? Genetic Similarity between European Populations of a Potential Disease Vector Culex Pipiens. Infect. Ecol. Epidemiol. 2012, 2, 12001. [Google Scholar]
- Mweya, C.N.; Kimera, S.I.; Kija, J.B.; Mboera, L.E. Predicting Distribution of Aedes Aegypti and Culex Pipiens Complex, Potential Vectors of Rift Valley Fever Virus in Relation to Disease Epidemics in East Africa. Infect. Ecol. Epidemiol. 2013, 3, 21748. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Amplicon Size (bp) |
|---|---|---|
| NSca | 5′ CCTTAACCTCTAATCAAC 3′ | 810 |
| NSng | 5′ TATCATGGATTACTTTCC 3′ | |
| NS2g | 5′ GATTTGCAGAGTGGTCGTC 3′ | 667 |
| NS3a | 5′ ATGCTGGGAAGTGATGAGCG 3′ |
| District | C. pipiens | C. tritaeniorhynchus | ||||
|---|---|---|---|---|---|---|
| Number of Mosquitoes | RA %/Species * | RA %/Total Mosquitoes ** | Number of Mosquitoes | RA %/Species * | RA %/Total Mosquitoes ** | |
| Al-Darb | 298 | 100 | 58.33 | 0 | 0 | 0.00 |
| Al-Reath | 103 | 100 | 35.71 | 0 | 0 | 0.00 |
| Al-Aridah | 173 | 100 | 47.06 | 0 | 0 | 0.00 |
| Abuareesh | 252 | 92.30 | 44.44 | 21 | 7.70 | 3.70 |
| Al-Ahad | 390 | 93.75 | 53.57 | 26 | 6.25 | 3.21 |
| Samttah | 193 | 100 | 55.56 | 0 | 0 | 0.00 |
| Sabyah | 322 | 93.33 | 50 | 23 | 6.66 | 3.57 |
| Damad | 163 | 100 | 50 | 0 | 0 | 0.00 |
| Beash | 187 | 91.66 | 44 | 17 | 8.33 | 4.00 |
| Districts | Al-Darb | Al-Reath | Al-Aridah | Abuareesh | Al-Ahad | Samttah | Sabyah | Damad | Beash |
|---|---|---|---|---|---|---|---|---|---|
| No of samples | 14 | 5 | 8 | 13 | 16 | 10 | 15 | 7 | 12 |
| RVF positive | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eifan, S.; Hanif, A.; Nour, I.; Alqahtani, S.; Eisa, Z.M.; Dafalla, O.; Kohl, A. Distribution and Molecular Identification of Culex pipiens and Culex tritaeniorhynchus as Potential Vectors of Rift Valley Fever Virus in Jazan, Saudi Arabia. Pathogens 2021, 10, 1334. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10101334
Eifan S, Hanif A, Nour I, Alqahtani S, Eisa ZM, Dafalla O, Kohl A. Distribution and Molecular Identification of Culex pipiens and Culex tritaeniorhynchus as Potential Vectors of Rift Valley Fever Virus in Jazan, Saudi Arabia. Pathogens. 2021; 10(10):1334. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10101334
Chicago/Turabian StyleEifan, Saleh, Atif Hanif, Islam Nour, Sultan Alqahtani, Zaki M. Eisa, Ommer Dafalla, and Alain Kohl. 2021. "Distribution and Molecular Identification of Culex pipiens and Culex tritaeniorhynchus as Potential Vectors of Rift Valley Fever Virus in Jazan, Saudi Arabia" Pathogens 10, no. 10: 1334. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10101334