
Degradation of Host Proteins and Apoptosis Induced by Foot-and-Mouth Disease Virus 3C Protease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Plasmids
2.3. Antibodies and Reagents
2.4. Western Blotting
2.5. Site-Direct Mutagenesis
2.6. Caspase Inhibitors Treatment
2.7. Cytotoxicity Test
2.8. Quantitative Reverse-Transcription-PCR(RT-qPCR) Analysis
2.9. Flow Cytometric Analysis of Apoptosis
2.10. Caspases Activity Assay
2.11. Statistical Analysis
3. Results
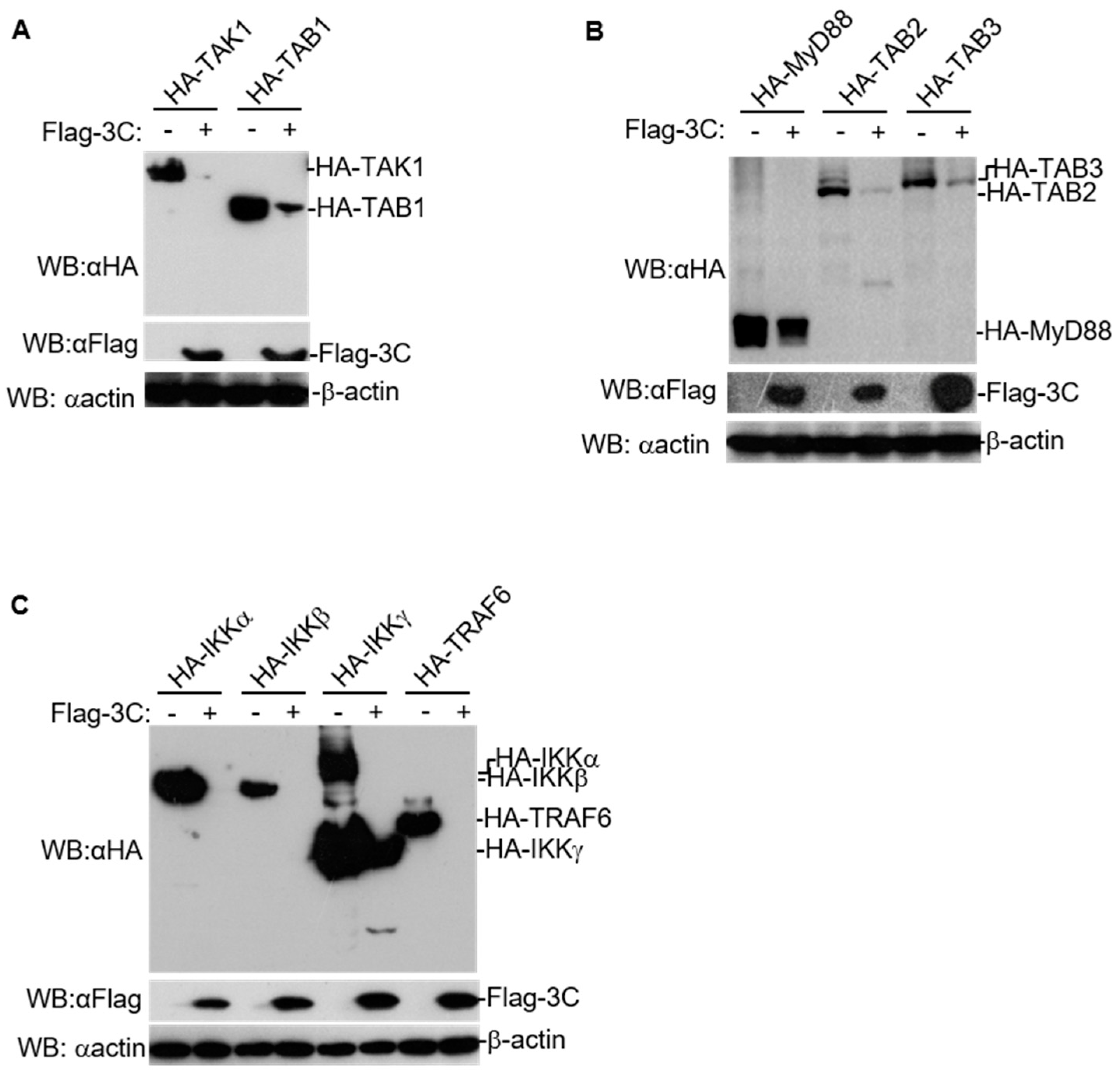
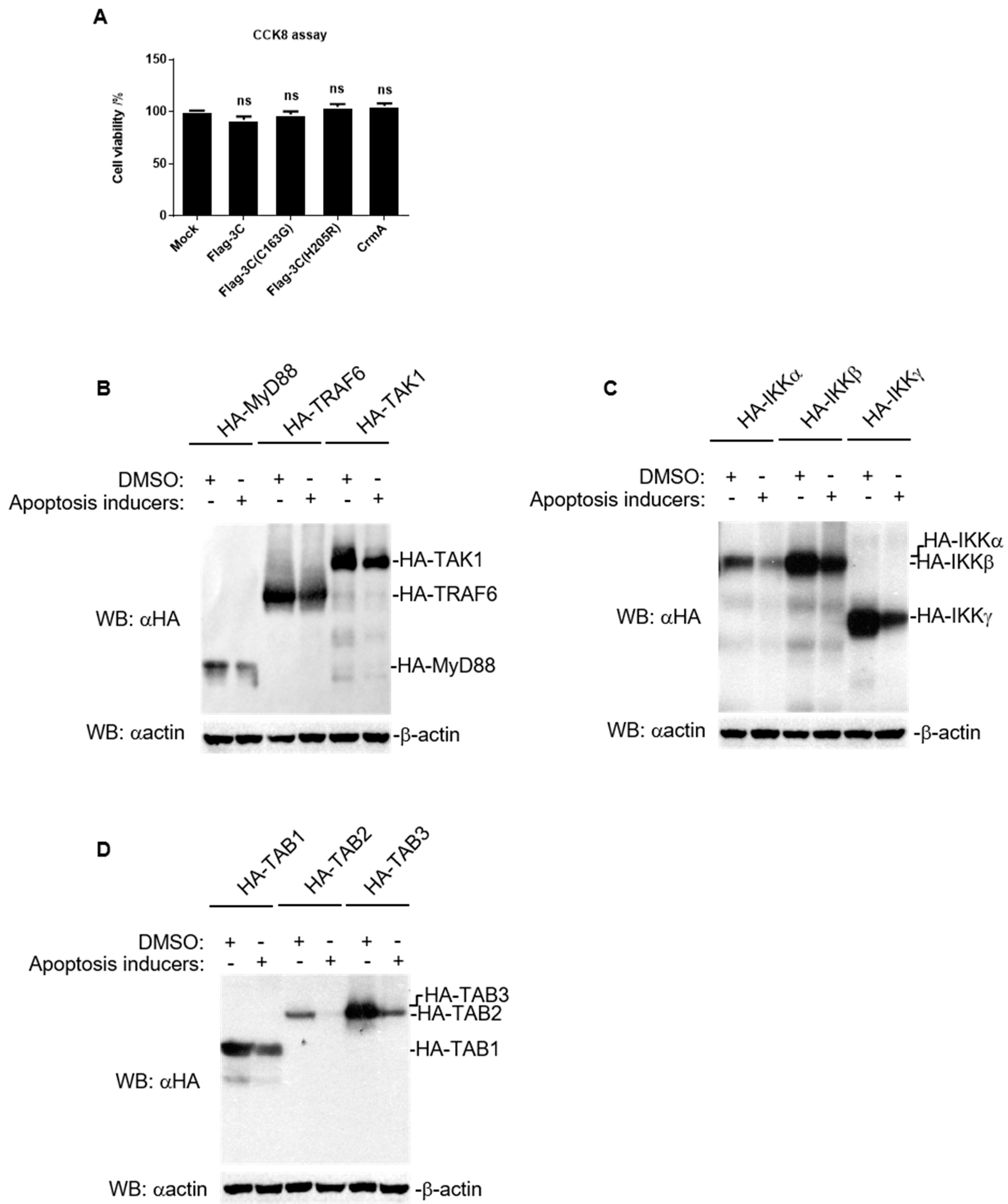
3.1. FMDV 3C Degrades Various Molecules in NF-κB Signaling
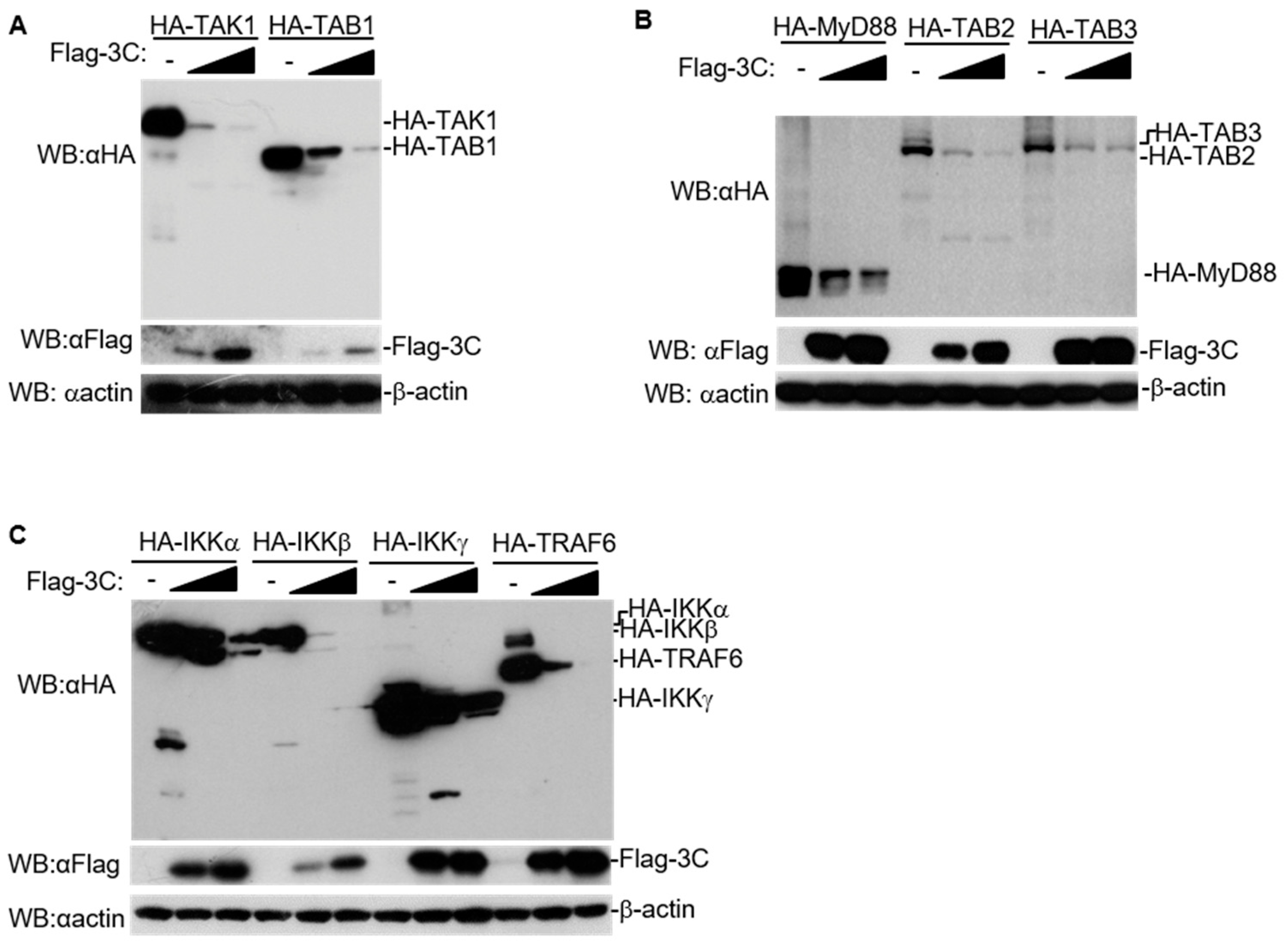
3.2. FMDV 3C Degrades Host Proteins Involved in NF-κB Signaling in a Dose-Dependent Manner
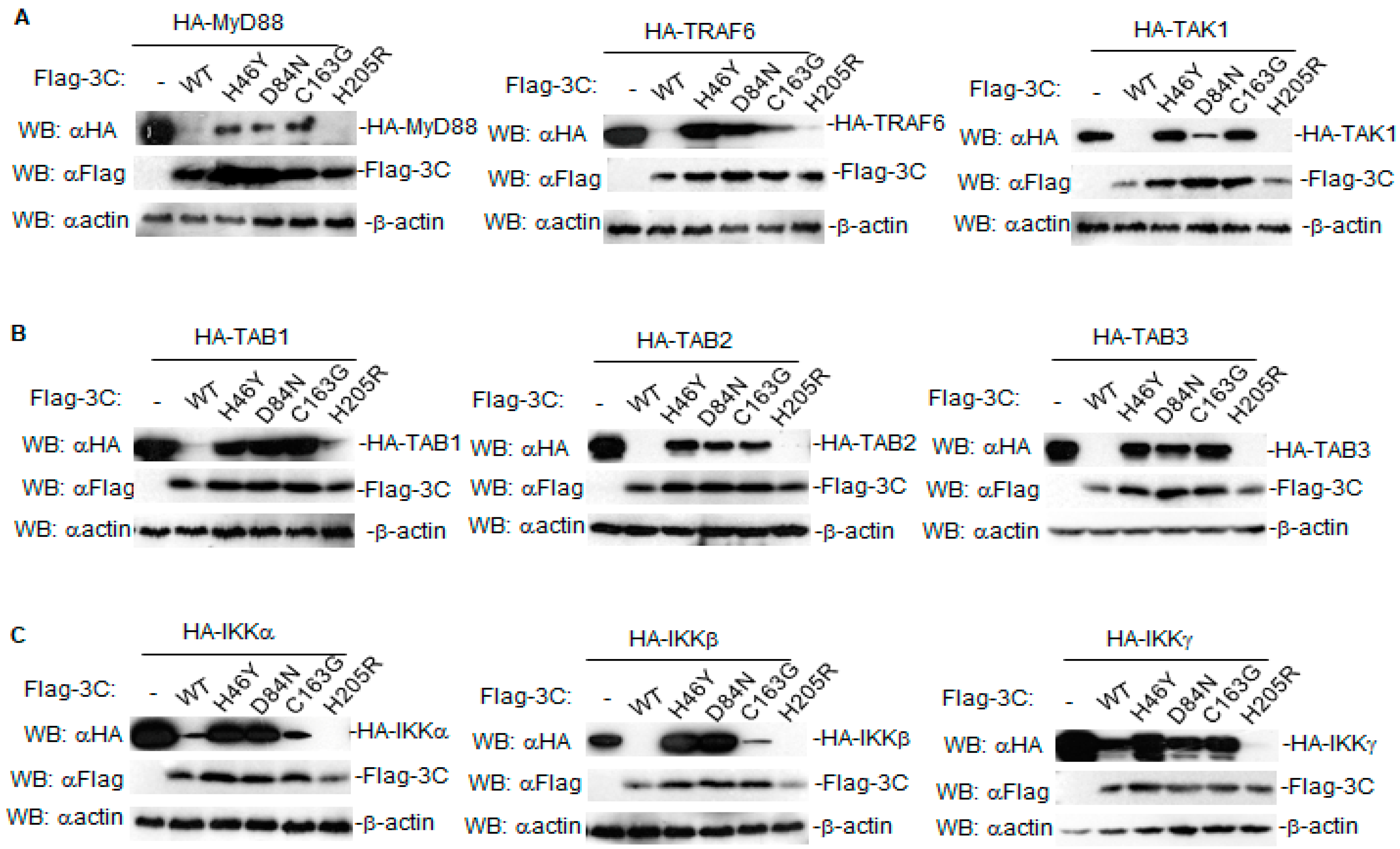
3.3. Proteolytic Activity Is Involved in the Degradation Induced by FMDV 3C
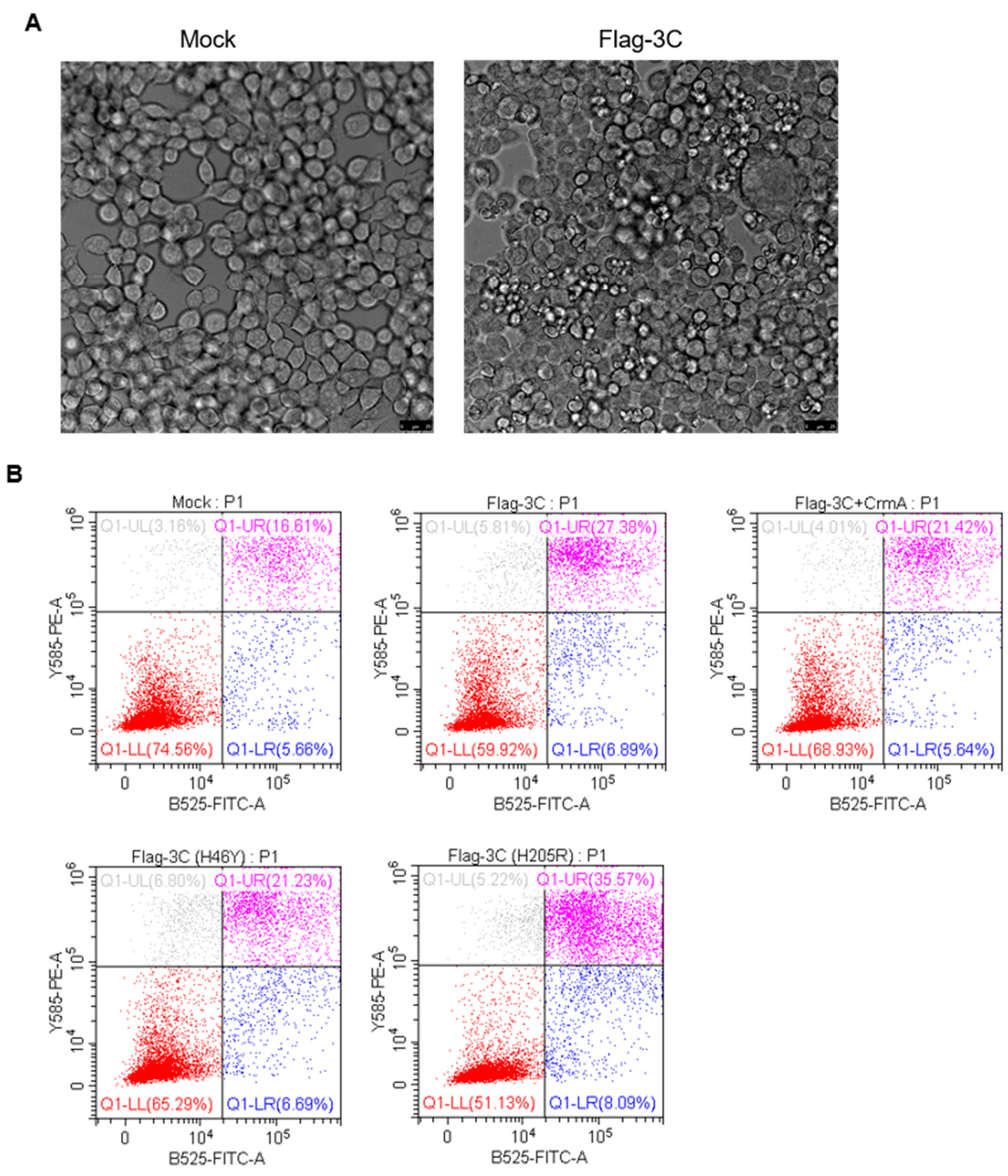
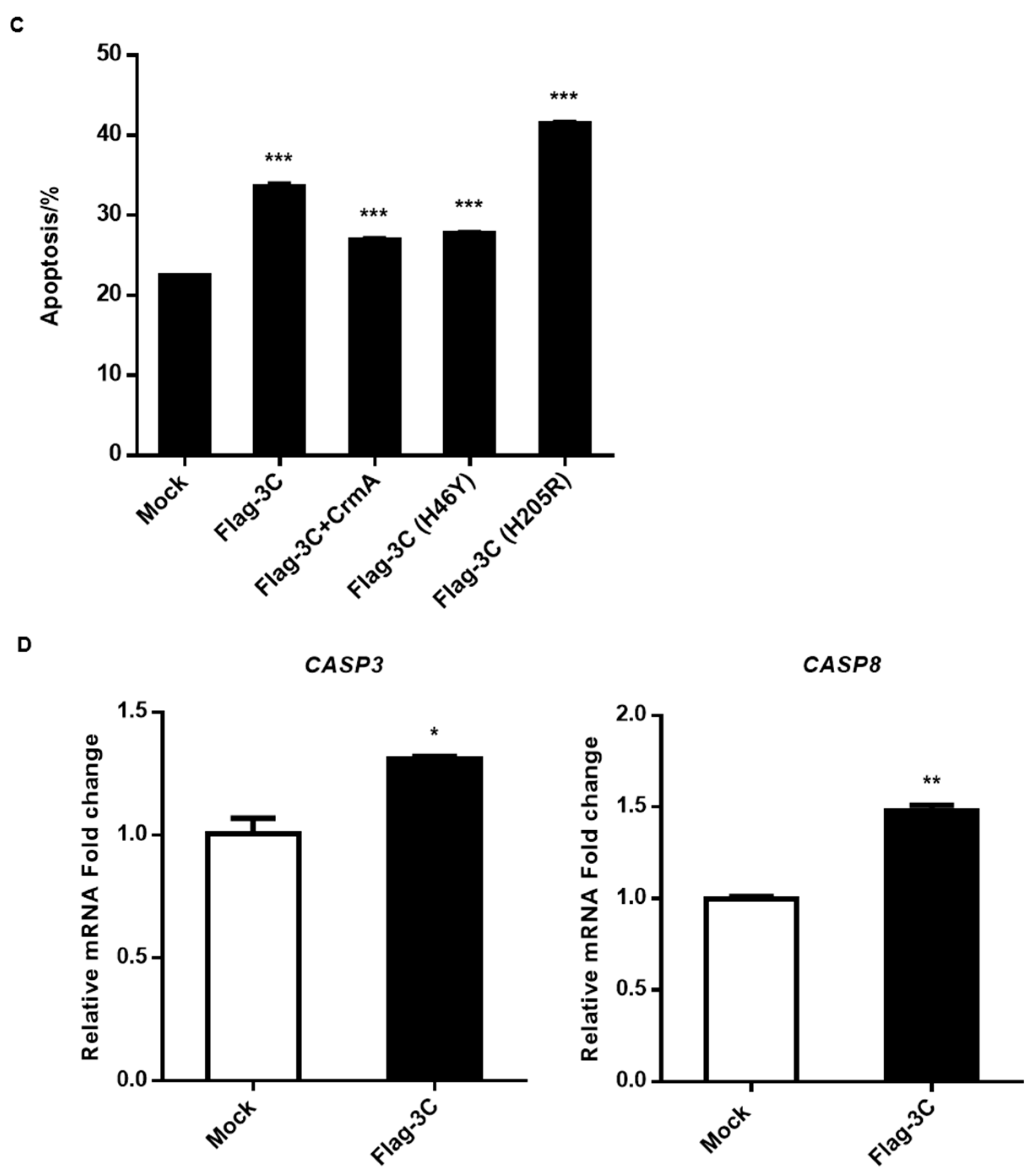
3.4. Expression of 3C Induces Apoptosis
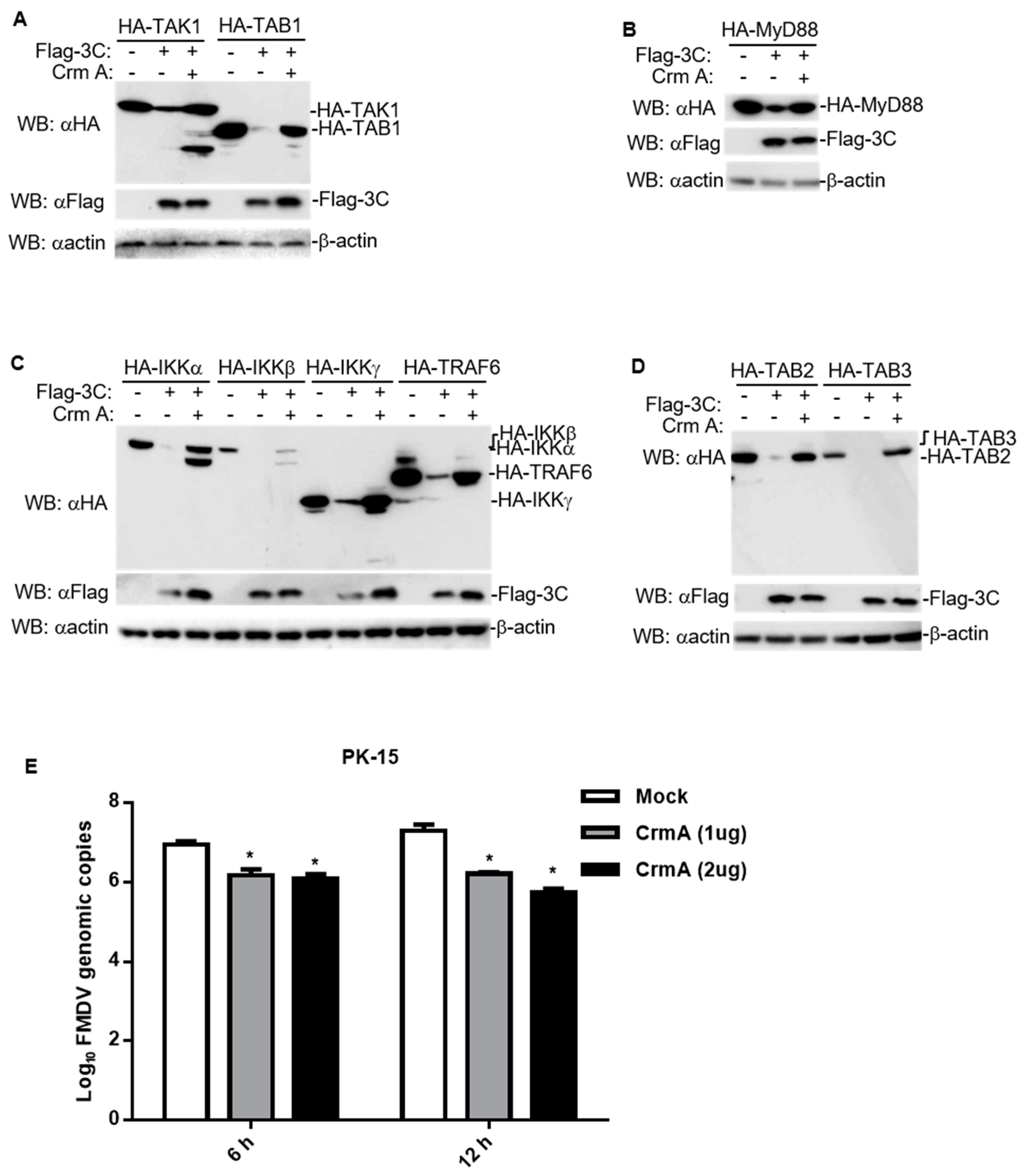
3.5. CrmA Abolished the Ability of 3C to Degrade Host Proteins Involved in NF-κB Signaling
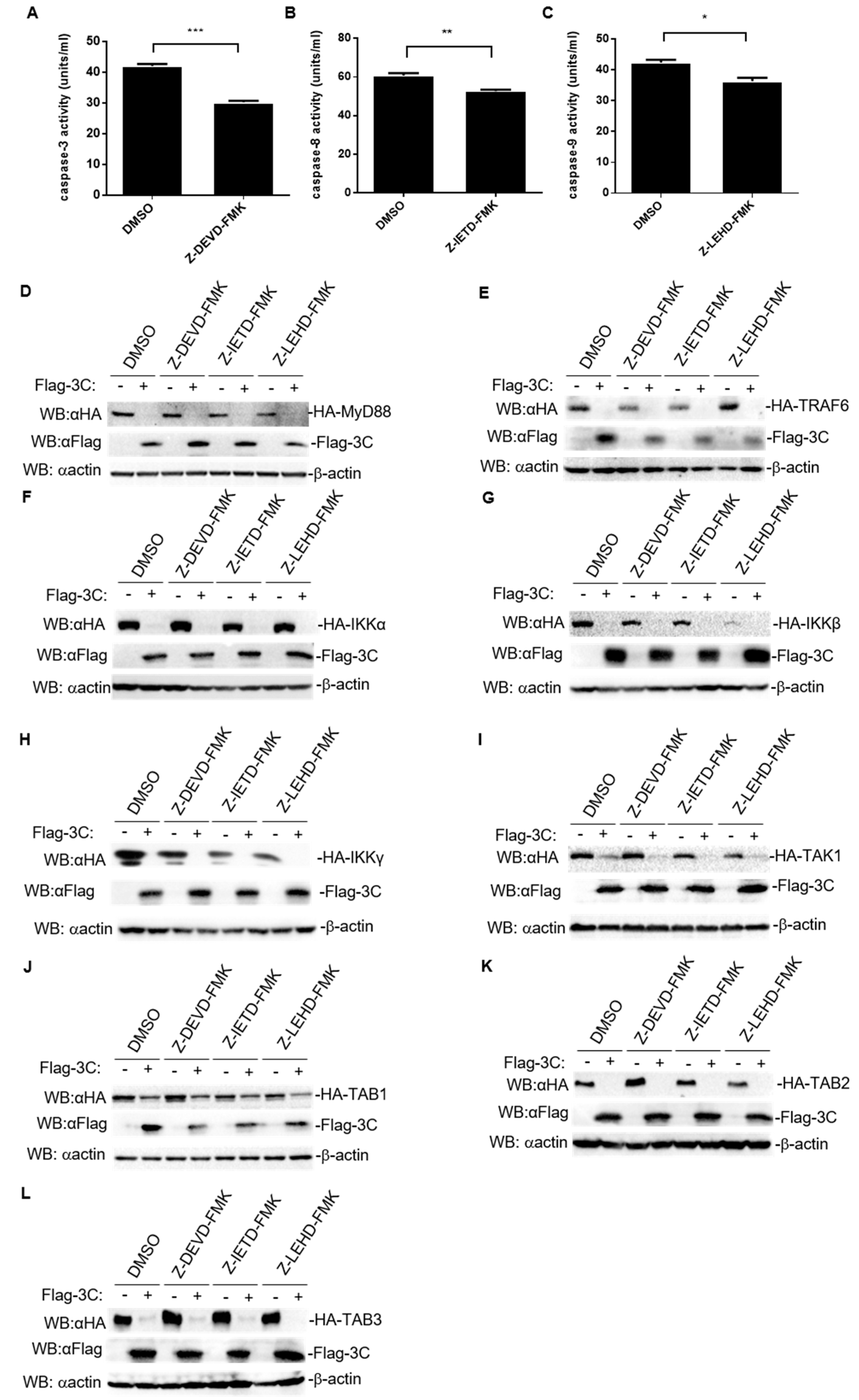
3.6. Degradation of Host Proteins Involved in NF-κB Signaling Induced by FMDV 3C Is Independent of the Activity of Caspase3, Caspase8, and Caspase9
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alexandersen, S.; Mowat, N. Foot-and-mouth disease: Host range and pathogenesis. Curr. Top. Microbiol. Immunol. 2005, 288, 9–42. [Google Scholar]
- Belsham, G.J. Distinctive features of foot-and-mouth disease virus, a member of the picornavirus family; aspects of virus protein synthesis, protein processing and structure. Prog. Biophys. Mol. Biol. 1993, 60, 241–260. [Google Scholar] [CrossRef]
- Sun, D.; Chen, S.; Cheng, A.; Wang, M. Roles of the Picornaviral 3C Proteinase in the Viral Life Cycle and Host Cells. Viruses 2016, 8, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, J.; Peng, J.; Yang, W.; Zhu, G.; Ren, J.; Li, D.; Zheng, H. Picornavirus 3C—A protease ensuring virus replication and subverting host responses. J. Cell Sci. 2021, 134, jcs.253237. [Google Scholar] [CrossRef] [PubMed]
- Falk, M.M.; Grigera, P.R.; E Bergmann, I.; Zibert, A.; Multhaup, G.; Beck, E. Foot-and-mouth disease virus protease 3C induces specific proteolytic cleavage of host cell histone H3. J. Virol. 1990, 64, 748–756. [Google Scholar] [CrossRef] [Green Version]
- Belsham, G.J.; McInerney, G.M.; Ross-Smith, N. Foot-and-mouth disease virus 3C protease induces cleavage of translation initiation factors eIF4A and eIF4G within infected cells. J. Virol. 2000, 74, 272–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Ross-Smith, N.; Proud, C.; Belsham, G.J. Cleavage of translation initiation factor 4AI (eIF4AI) but not eIF4AII by foot-and-mouth disease virus 3C protease: Identification of the eIF4AI cleavage site. FEBS Lett. 2001, 507, 1–5. [Google Scholar] [CrossRef]
- Lawrence, P.; Schafer, E.A.; Rieder, E. The nuclear protein Sam68 is cleaved by the FMDV 3C protease redistributing Sam68 to the cytoplasm during FMDV infection of host cells. Virology 2012, 425, 40–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Pan, T.; Wang, D.; Fang, L.; Ma, J.; Zhu, X.; Shi, Y.; Zhang, K.; Zheng, H.; Chen, H.; et al. Foot-and-Mouth Disease Virus Counteracts on Internal Ribosome Entry Site Suppression by G3BP1 and Inhibits G3BP1-Mediated Stress Granule Assembly via Post-Translational Mechanisms. Front. Immunol. 2018, 9, 1142. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Yang, D.; Sun, C.; Wang, H.; Zhao, B.; Zhou, G.; Yu, L. hnRNP K is a novel ITAF that negatively regulates foot-and-mouth disease virus translation and replication and is antagonized by viral 3C protease. J. Virol. 2020, 38, 39. [Google Scholar]
- Wang, D.; Fang, L.; Li, K.; Zhong, H.; Fan, J.; Ouyang, C.; Zhang, H.; Duan, E.; Luo, R.; Zhang, Z.; et al. Foot-and-mouth disease virus 3C protease cleaves NEMO to impair innate immune signaling. J. Virol. 2012, 86, 9311–9322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Mogensen, M.M.; Powell, P.P.; Curry, S.; Wileman, T. Foot-and-mouth disease virus 3C protease induces fragmentation of the Golgi compartment and blocks intra-Golgi transport. J. Virol. 2013, 87, 11721–11729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Bi, J.; Liu, J.; Liu, X.; Wu, X.; Jiang, P.; Yoo, D.; Zhang, Y.; Wu, J.; Wan, R.; et al. 3Cpro of foot-and-mouth disease virus antagonizes the interferon signaling pathway by blocking STAT1/STAT2 nuclear translocation. J. Virol. 2014, 88, 4908–4920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-H.; Lau, K.S.; Kuo, R.-L.; Horng, J.-T. dsRNA Binding Domain of PKR Is Proteolytically Released by Enterovirus A71 to Facilitate Viral Replication. Front. Cell. Infect. Microbiol. 2017, 7, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Han, S.; Yan, D.; Gao, Y.; Wei, Y.; Liu, X.; Liao, Y.; Guo, H.; Sun, S. Foot-and-mouth disease virus infection suppresses autophagy and NF-small ka, CyrillicB antiviral responses via degradation of ATG5-ATG12 by 3C(pro). Cell Death Dis. 2017, 8, e2561. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Croft, S.N.; Walker, E.J.; Ghildyal, R. Picornaviruses and Apoptosis: Subversion of Cell Death. mBio 2017, 8, e01009-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; He, J.; Guo, J.; Jin, Y.; Yang, F.; Lv, L.; Liu, X. Genetic characterization of a new pandemic Southeast Asia topotype strain of serotype O foot-and-mouth disease virus isolated in China during 2010. Virus Genes 2012, 44, 80–88. [Google Scholar] [CrossRef]
- Zhu, Z.; Shi, Z.; Yan, W.; Wei, J.; Shao, N.; Deng, X.; Wang, S.; Li, B.; Tong, G.; Ma, Z. Nonstructural protein 1 of influenza A virus interacts with human guanylate-binding protein 1 to antagonize antiviral activity. PLoS ONE 2013, 8, e55920. [Google Scholar]
- Grubman, M.J.; Zellner, M.; Bablanian, G.; Mason, P.W.; Piccone, M.E. Identification of the active-site residues of the 3C proteinase of foot-and-mouth disease virus. Virology 1995, 213, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Moniwa, M.; Clavijo, A.; Li, M.; Collignon, B.; Kitching, R.P. Performance of a foot-and-mouth disease virus reverse transcription-polymerase chain reaction with amplification controls between three real-time instruments. J. Veter-Diagn. Investig. 2007, 19, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Barco, A.; Feduchi, E.; Carrasco, L. Poliovirus protease 3C(pro) kills cells by apoptosis. Virology 2000, 266, 352–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.-L.; Hsu, T.-A.; Chen, T.-C.; Chang, S.-C.; Lee, J.-C.; Chen, C.-C.; Stollar, V.; Shih, S.-R. The 3C protease activity of enterovirus 71 induces human neural cell apoptosis. Virology 2002, 293, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.H.V.; Maggioli, M.F.; Otta, J.; Joshi, L.R.; Lawson, S.; Diel, D.G. Senecavirus A 3C Protease Mediates Host Cell Apoptosis Late in Infection. Front. Immunol. 2019, 10, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Li, X.; Wu, M.; Qin, L.; Chen, H.; Qian, P. Seneca Valley Virus 2C and 3C(pro) Induce Apoptosis via Mitochondrion-Mediated Intrinsic Pathway. Front. Microbiol. 2019, 10, 1202. [Google Scholar] [CrossRef]
- Chau, D.H.; Yuan, J.; Zhang, H.; Cheung, P.; Lim, T.; Liu, Z.; Sall, A.; Yang, D. Coxsackievirus B3 proteases 2A and 3C induce apoptotic cell death through mitochondrial injury and cleavage of eIF4GI but not DAP5/p97/NAT1. Apoptosis 2007, 12, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Cohen, G.M. Caspases: The executioners of apoptosis. Biochem. J. 1997, 326 Pt 1, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhu, Z.; Du, X.; Cao, W.; Yang, F.; Zhang, X.; Feng, H.; Li, D.; Zhang, K.; Liu, X.; et al. Foot-and-mouth disease virus induces lysosomal degradation of host protein kinase PKR by 3C proteinase to facilitate virus replication. Virology 2017, 509, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Su, J.; Wang, H.; Chang, J.; Wang, S.; Zheng, W.; Cai, Y.; Wei, W.; Gordy, J.; Markham, R.; et al. Disruption of MDA5-Mediated Innate Immune Responses by the 3C Proteins of Coxsackievirus A16, Coxsackievirus A6, and Enterovirus D68. J. Virol. 2017, 91, e00546-17. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.; Han, N.; Xiao, X.; Jin, Q.; He, B.; Wang, J. Enterovirus 71 3C inhibits cytokine expression through cleavage of the TAK1/TAB1/TAB2/TAB3 complex. J. Virol. 2014, 88, 9830–9841. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Fang, L.; Wei, D.; Zhang, H.; Luo, R.; Chen, H.; Li, K.; Xiao, S. Hepatitis A virus 3C protease cleaves NEMO to impair induction of beta interferon. J. Virol. 2014, 88, 10252–10258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croft, S.; Walker, E.J.; Ghildyal, R. Human Rhinovirus 3C protease cleaves RIPK1, concurrent with caspase 8 activation. Sci. Rep. 2018, 8, 1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekert, P.G.; Silke, J.; Vaux, D.L. Inhibition of apoptosis and clonogenic survival of cells expressing crmA variants: Optimal caspase substrates are not necessarily optimal inhibitors. EMBO J. 1999, 18, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevich, R.M.; Regula, K.M.; Kirshenbaum, L.A. Serpin protein CrmA suppresses hypoxia-mediated apoptosis of ventricular myocytes. Circulation 2001, 103, 1984–1991. [Google Scholar] [CrossRef] [Green Version]
- Komiyama, T.; A Ray, C.; Pickup, D.J.; Howard, A.D.; A Thornberry, N.; Peterson, E.P.; Salvesen, G. Inhibition of interleukin-1 beta converting enzyme by the cowpox virus serpin CrmA. An example of cross-class inhibition. J. Biol. Chem. 1994, 269, 19331–19337. [Google Scholar] [CrossRef]
- Bloomer, D.T.; Kitevska-Ilioski, T.; Pantaki-Eimany, D.; Ji, Y.; Miles, M.A.; Heras, B.; Hawkins, C.J. CrmA orthologs from diverse poxviruses potently inhibit caspases-1 and -8, yet cleavage site mutagenesis frequently produces caspase-1-specific variants. Biochem. J. 2019, 476, 1335–1357. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, J.; Peng, J.; Ren, J.; Zhu, G.; Ru, Y.; Tian, H.; Li, D.; Zheng, H. Degradation of Host Proteins and Apoptosis Induced by Foot-and-Mouth Disease Virus 3C Protease. Pathogens 2021, 10, 1566. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121566
Yi J, Peng J, Ren J, Zhu G, Ru Y, Tian H, Li D, Zheng H. Degradation of Host Proteins and Apoptosis Induced by Foot-and-Mouth Disease Virus 3C Protease. Pathogens. 2021; 10(12):1566. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121566
Chicago/Turabian StyleYi, Jiamin, Jiangling Peng, Jingjing Ren, Guoqiang Zhu, Yi Ru, Hong Tian, Dan Li, and Haixue Zheng. 2021. "Degradation of Host Proteins and Apoptosis Induced by Foot-and-Mouth Disease Virus 3C Protease" Pathogens 10, no. 12: 1566. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121566