

Serendipitous Discovery of a Competitive Inhibitor of FraB, a Salmonella Deglycase and Drug Target

 , , and
, , and 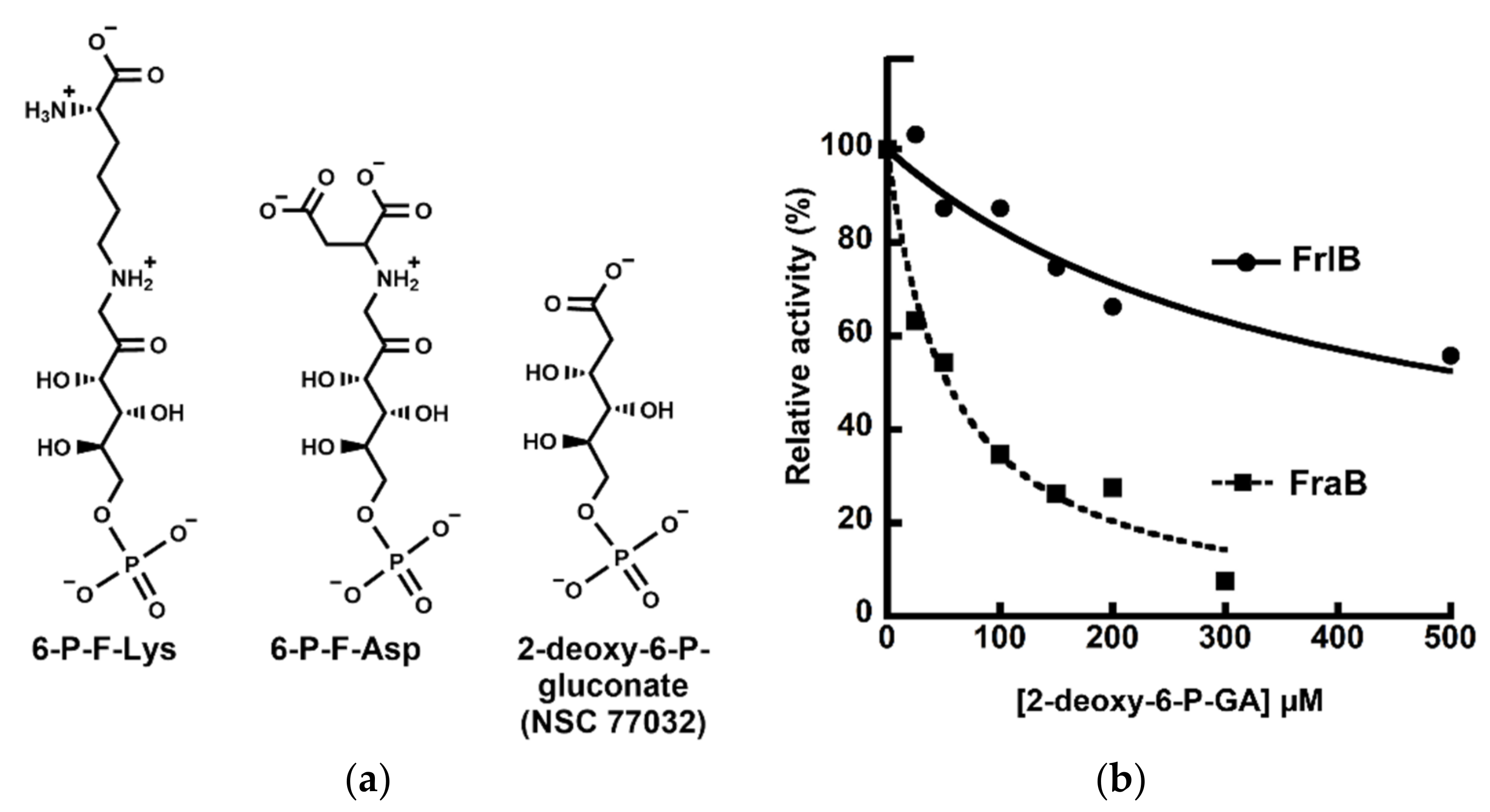{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Tanimoto Search of the NCI Database for Small Molecules Based on Structural Similarity
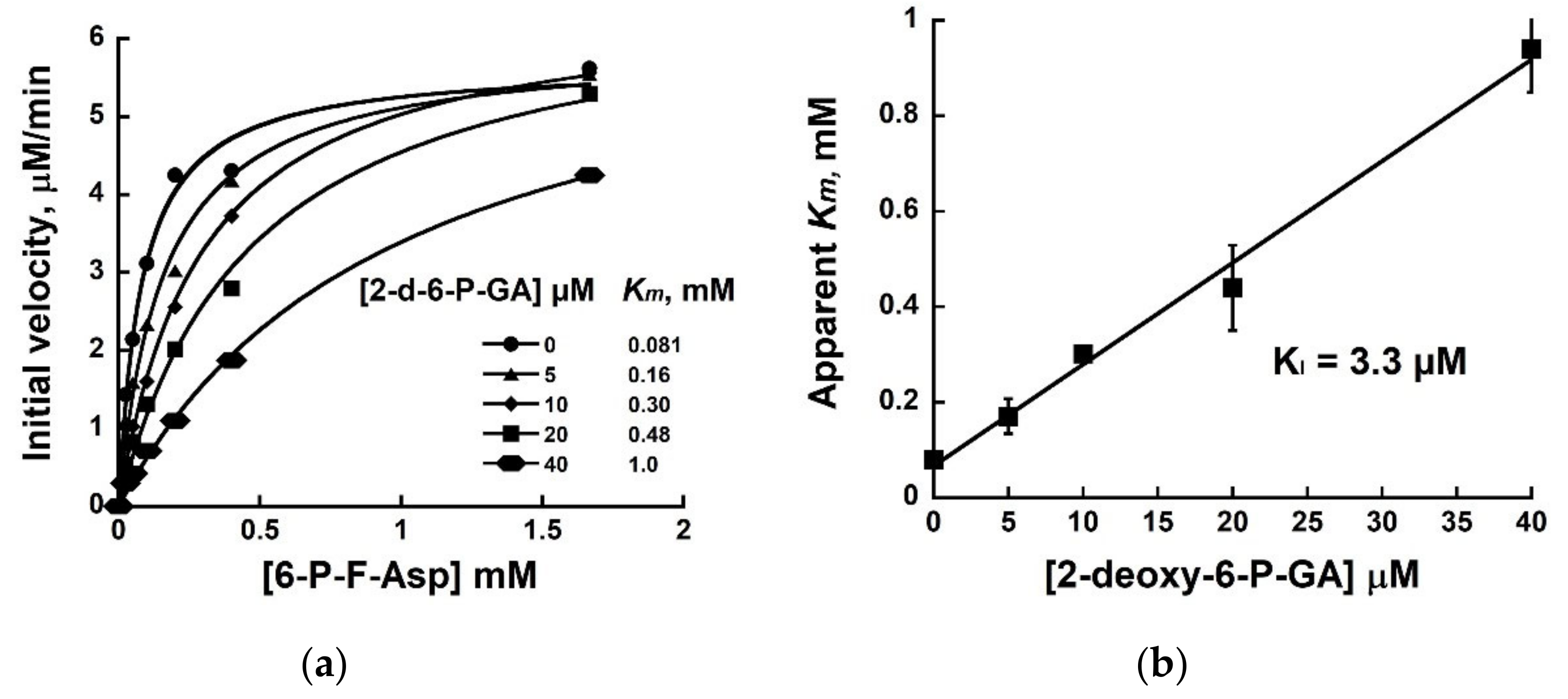
2.2. Evaluation of 2-deoxy-6-P-GA as an Inhibitor of FraB and FrlB
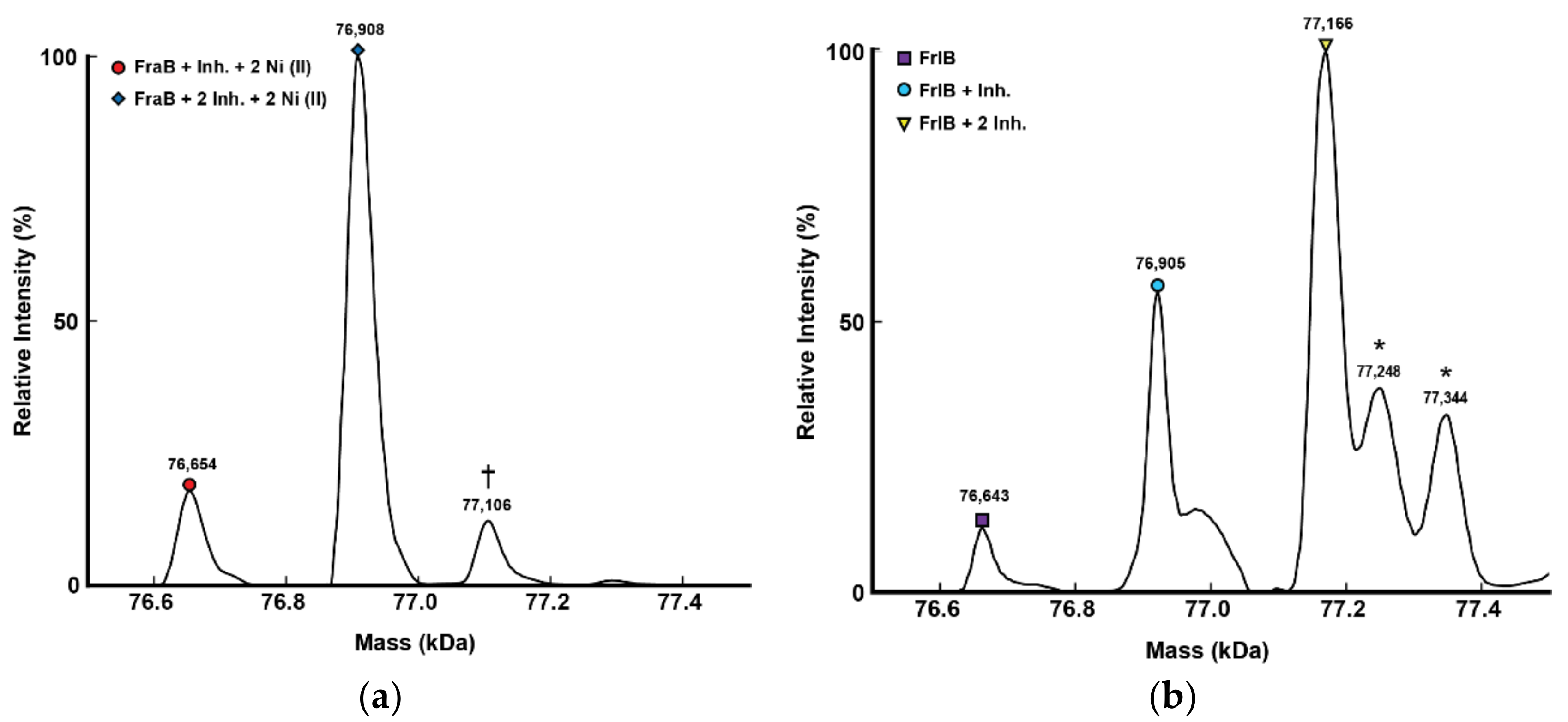
2.3. Mass Spectrometry-Based Confirmation of FraB/FrlB–2-deoxy-6-P-GA Complexes
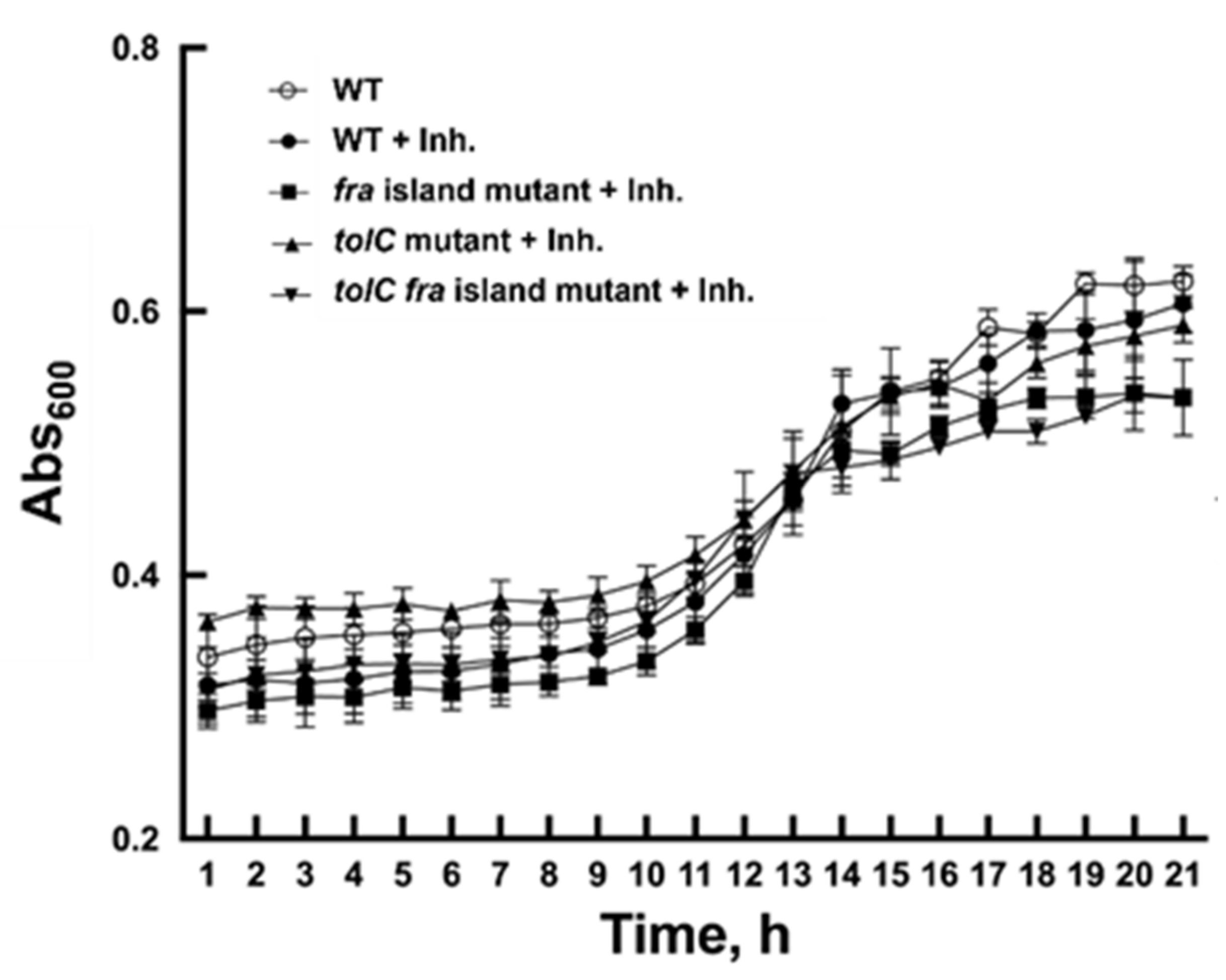
2.4. Cell-Based Assays with Salmonella Wild-Type, ∆fra, and ∆tolC Strains
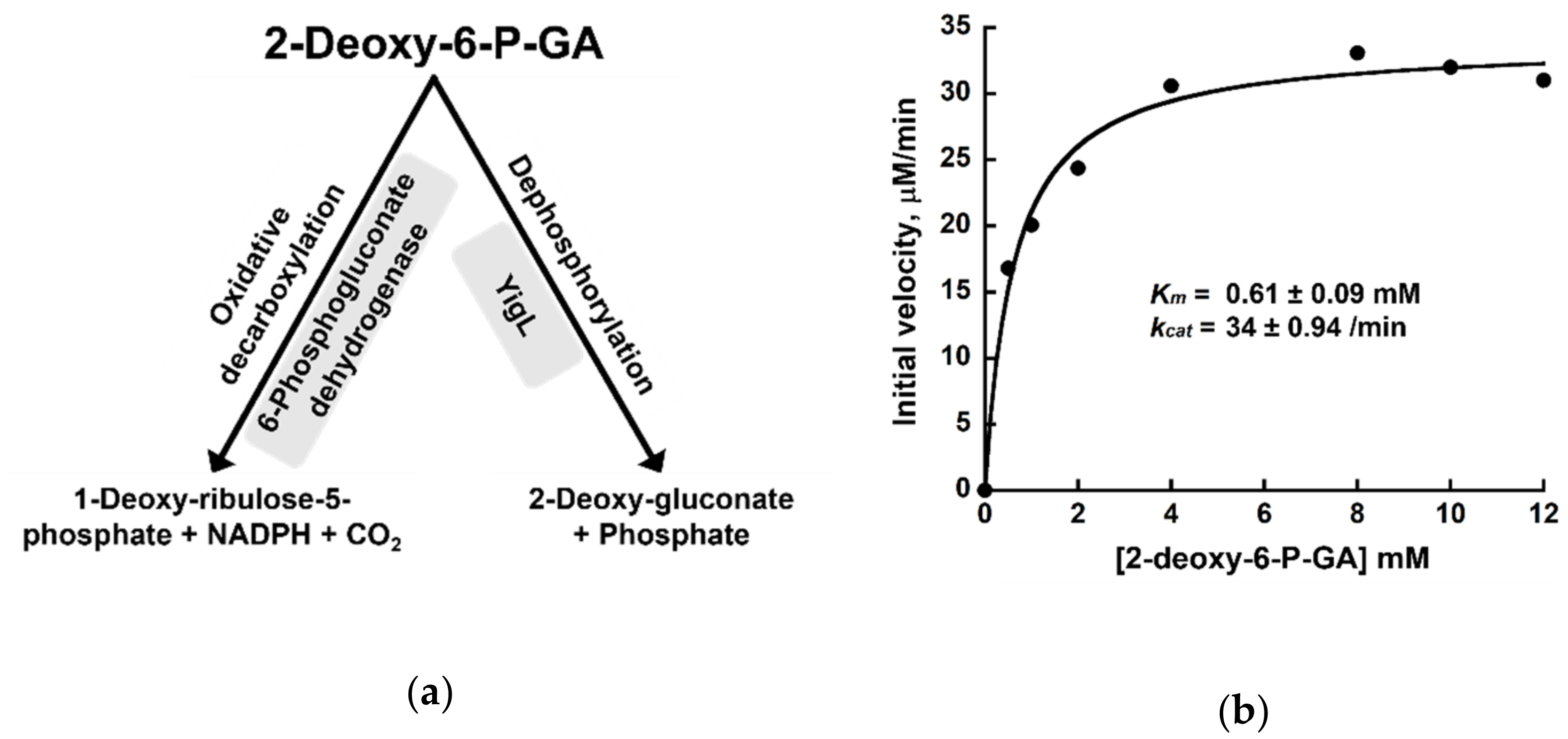
2.5. Dephosphorylation of 2-deoxy-6-P-GA by YigL
3. Discussion
4. Materials and Methods
4.1. Tanimoto Search for Structurally Similar Compounds
4.2. NMR Studies
4.3. Protein Overexpression and Purification
4.4. Inhibition Assays
4.5. Mass Spectrometry (MS) Analyses
4.6. Live-Cell Assays
4.7. Dephosphorylation of 2-deoxy-6-P-GA by YigL
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, Z.; Zheng, J. Occurrence, characteristics, and applications of fructosyl amine oxidases (amadoriases). Appl. Microbiol. Biotechnol. 2010, 86, 1613–1619. [Google Scholar] [CrossRef] [PubMed]
- Wiame, E.; Delpierre, G.; Collard, F.; Van Schaftingen, E. Identification of a pathway for the utilization of the Amadori product fructoselysine in Escherichia coli. J. Biol. Chem. 2002, 277, 42523–42529. [Google Scholar] [CrossRef] [PubMed]
- Wiame, E.; Duquenne, A.; Delpierre, G.; Van Schaftingen, E. Identification of enzymes acting on alpha-glycated amino acids in Bacillus subtilis. FEBS Lett. 2004, 577, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Newsom, D.L.; González, J.F.; Sabag-Daigle, A.; Stahl, C.; Steidley, B.; Dubena, J.; Dyszel, J.L.; Smith, J.N.; Dieye, Y.; et al. Fructose-asparagine is a primary nutrient during growth of Salmonella in the inflamed intestine. PLoS Pathog. 2014, 10, e1004209. [Google Scholar] [CrossRef]
- Sabag-Daigle, A.; Blunk, H.M.; Sengupta, A.; Wu, J.; Bogard, A.J.; Ali, M.M.; Stahl, C.; Wysocki, V.H.; Gopalan, V.; Behrman, E.J.; et al. A metabolic intermediate of the fructose-asparagine utilization pathway inhibits growth of a Salmonella fraB mutant. Sci. Rep. 2016, 6, 28117. [Google Scholar] [CrossRef]
- Sengupta, A.; Wu, J.; Seffernick, J.T.; Sabag-Daigle, A.; Thomsen, N.; Chen, T.H.; Capua, A.D.; Bell, C.E.; Ahmer, B.M.M.; Lindert, S.; et al. Integrated use of biochemical, native mass spectrometry, computational, and genome-editing methods to elucidate the mechanism of a Salmonella deglycase. J. Mol. Biol. 2019, 431, 4497–4513. [Google Scholar] [CrossRef]
- Biswas, P.K.; Behrman, E.J.; Gopalan, V. Characterization of a Salmonella sugar kinase essential for the utilization of fructose-asparagine. Biochem. Cell Biol. 2017, 95, 304–309. [Google Scholar] [CrossRef]
- Sabag-Daigle, A.; Boulanger, E.; Thirugnanasambantham, P.; Law, J.; Bogard, A.; Behrman, E.J.; Gopalan, V.; Ahmer, B. Identification of small molecule inhibitors of the Salmonella FraB deglycase using a live-cell assay. PLoS ONE 2022. under review. [Google Scholar]
- Miller, K.A.; Phillips, R.S.; Kilgore, P.B.; Smith, G.L.; Hoover, T.R. A mannose family phosphotransferase system permease and associated enzymes are required for utilization of fructoselysine and glucoselysine in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2015, 197, 2831–2839. [Google Scholar] [CrossRef]
- Kovvali, S.; Di Capua, A.; Wysocki, V.H.; Bell, C.E.; Gopalan, V. Elucidating the catalytic mechanism of a bacterial deglycase essential for utilization of fructose-lysine, an Amadori product. FASEB J 2022, 36. [Google Scholar] [CrossRef]
- National Cancer Institute. Downloadable Structure of NCI Open Database Compounds. Available online: https://cactus.nci.nih.gov/download/nci/ (accessed on 12 December 2020).
- Pezzotti, F.; Therisod, M. Enzymatic synthesis of aldonic acids. Carbohydr. Res. 2006, 341, 2290–2292. [Google Scholar] [CrossRef]
- Hanau, S.; Dallocchio, F.; Rippa, M. NADPH activates a decarboxylation reaction catalysed by lamb liver 6-phosphogluconate dehydrogenase. Biochim. Biophys. Acta 1992, 1122, 273–277. [Google Scholar] [CrossRef]
- Rippa, M.; Giovannini, P.P.; Barrett, M.P.; Dallocchio, F.; Hanau, S. 6-Phosphogluconate dehydrogenase: The mechanism of action investigated by a comparison of the enzyme from different species. Biochim. Biophys. Acta 1998, 1429, 83–92. [Google Scholar] [CrossRef]
- Allen, K.N.; Dunaway-Mariano, D. Markers of fitness in a successful enzyme superfamily. Curr. Opin. Struct. Biol. 2009, 19, 658–665. [Google Scholar] [CrossRef]
- Kuznetsova, E.; Proudfoot, M.; Gonzalez, C.F.; Brown, G.; Omelchenko, M.V.; Borozan, I.; Carmel, L.; Wolf, Y.I.; Mori, H.; Savchenko, A.V.; et al. Genome-wide analysis of substrate specificities of the Escherichia coli haloacid dehalogenase-like phosphatase family. J. Biol. Chem. 2006, 281, 36149–36161. [Google Scholar] [CrossRef]
- Koonin, E.V.; Tatusov, R.L. Computer analysis of bacterial haloacid dehalogenases defines a large superfamily of hydrolases with diverse specificity. Application of an iterative approach to database search. J. Mol. Biol. 1994, 244, 125–132. [Google Scholar] [CrossRef]
- Papenfort, K.; Sun, Y.; Miyakoshi, M.; Vanderpool, C.K.; Vogel, J. Small RNA-mediated activation of sugar phosphatase mRNA regulates glucose homeostasis. Cell 2013, 153, 426–437. [Google Scholar] [CrossRef]
- Cao, Y.; Peng, Q.; Li, S.; Deng, Z.; Gao, J. The intriguing biology and chemistry of fosfomycin: The only marketed phosphonate antibiotic. RSC Adv. 2019, 9, 42204–42218. [Google Scholar] [CrossRef]
- RDKit. RDKit: Open-Source Cheminformatics Software. Available online: http://www.rdkit.org (accessed on 12 December 2020).
- Cereto-Massagué, A.; Ojeda, M.J.; Valls, C.; Mulero, M.; Garcia-Vallvé, S.; Pujadas, G. Molecular fingerprint similarity search in virtual screening. Methods 2015, 71, 58–63. [Google Scholar] [CrossRef]
- Bajusz, D.; Rácz, A.; Héberger, K. Why is Tanimoto index an appropriate choice for fingerprint-based similarity calculations? J. Cheminform. 2015, 7, 20. [Google Scholar] [CrossRef]
- Bauer, S.; Biely, P. Metabolism of 2-deoxy-D-glucose by baker’s yeast. II. Formation of 2-deoxy-D-gluconic acid. Collect. Czechoslov. Chem. Commun. 1968, 33, 1165–1173. [Google Scholar] [CrossRef]
- Rippa, M.; Signorini, M.; Dallocchio, F. A multiple role for the coenzyme in the mechanism of action of 6-phosphogluconate dehydrogenase. The oxidative decarboxylation of 2-deoxy-6-phosphogluconate. J. Biol. Chem. 1973, 248, 4920–4925. [Google Scholar] [CrossRef]
- Sung, S.S.; Silverstein, S.C. Role of 2-deoxy-D-glucose in the inhibition of phagocytosis by mouse peritoneal macrophage. Biochim. Biophys. Acta 1985, 845, 204–215. [Google Scholar] [CrossRef]
- Marty, M.T.; Baldwin, A.J.; Marklund, E.G.; Hochberg, G.K.; Benesch, J.L.; Robinson, C.V. Bayesian deconvolution of mass and ion mobility spectra: From binary interactions to polydisperse ensembles. Anal. Chem. 2015, 87, 4370–4376. [Google Scholar] [CrossRef]
- Porwollik, S.; Santiviago, C.A.; Cheng, P.; Long, F.; Desai, P.; Fredlund, J.; Srikumar, S.; Silva, C.A.; Chu, W.; Chen, X.; et al. Defined single-gene and multi-gene deletion mutant collections in Salmonella enterica sv Typhimurium. PLoS ONE 2014, 9, e99820. [Google Scholar] [CrossRef]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1972. [Google Scholar]
- Sabag-Daigle, A.; Blunk, H.M.; Gonzalez, J.F.; Steidley, B.L.; Boyaka, P.N.; Ahmer, B.M.M. Use of attenuated but metabolically competent Salmonella as a probiotic to prevent or treat Salmonella infection. Infect. Immun. 2016, 84, 2131–2140. [Google Scholar] [CrossRef]
- Van Veldhoven, P.P.; Mannaerts, G.P. Inorganic and organic phosphate measurements in the nanomolar range. Anal. Biochem. 1987, 161, 45–48. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thirugnanasambantham, P.; Kovvali, S.; Cool, A.; Gao, Y.; Sabag-Daigle, A.; Boulanger, E.F.; Mitton-Fry, M.; Capua, A.D.; Behrman, E.J.; Wysocki, V.H.; et al. Serendipitous Discovery of a Competitive Inhibitor of FraB, a Salmonella Deglycase and Drug Target. Pathogens 2022, 11, 1102. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11101102
Thirugnanasambantham P, Kovvali S, Cool A, Gao Y, Sabag-Daigle A, Boulanger EF, Mitton-Fry M, Capua AD, Behrman EJ, Wysocki VH, et al. Serendipitous Discovery of a Competitive Inhibitor of FraB, a Salmonella Deglycase and Drug Target. Pathogens. 2022; 11(10):1102. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11101102
Chicago/Turabian StyleThirugnanasambantham, Pankajavalli, Sravya Kovvali, Austin Cool, Yuan Gao, Anice Sabag-Daigle, Erin F. Boulanger, Mark Mitton-Fry, Angela Di Capua, Edward J. Behrman, Vicki H. Wysocki, and et al. 2022. "Serendipitous Discovery of a Competitive Inhibitor of FraB, a Salmonella Deglycase and Drug Target" Pathogens 11, no. 10: 1102. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11101102