
Microbial Genetics and Clonal Dissemination of Salmonella enterica Serotype Javiana Isolated from Human Populations in Arkansas, USA
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Epidemiology and Demographics
2.2. Antimicrobial Susceptibility Testing
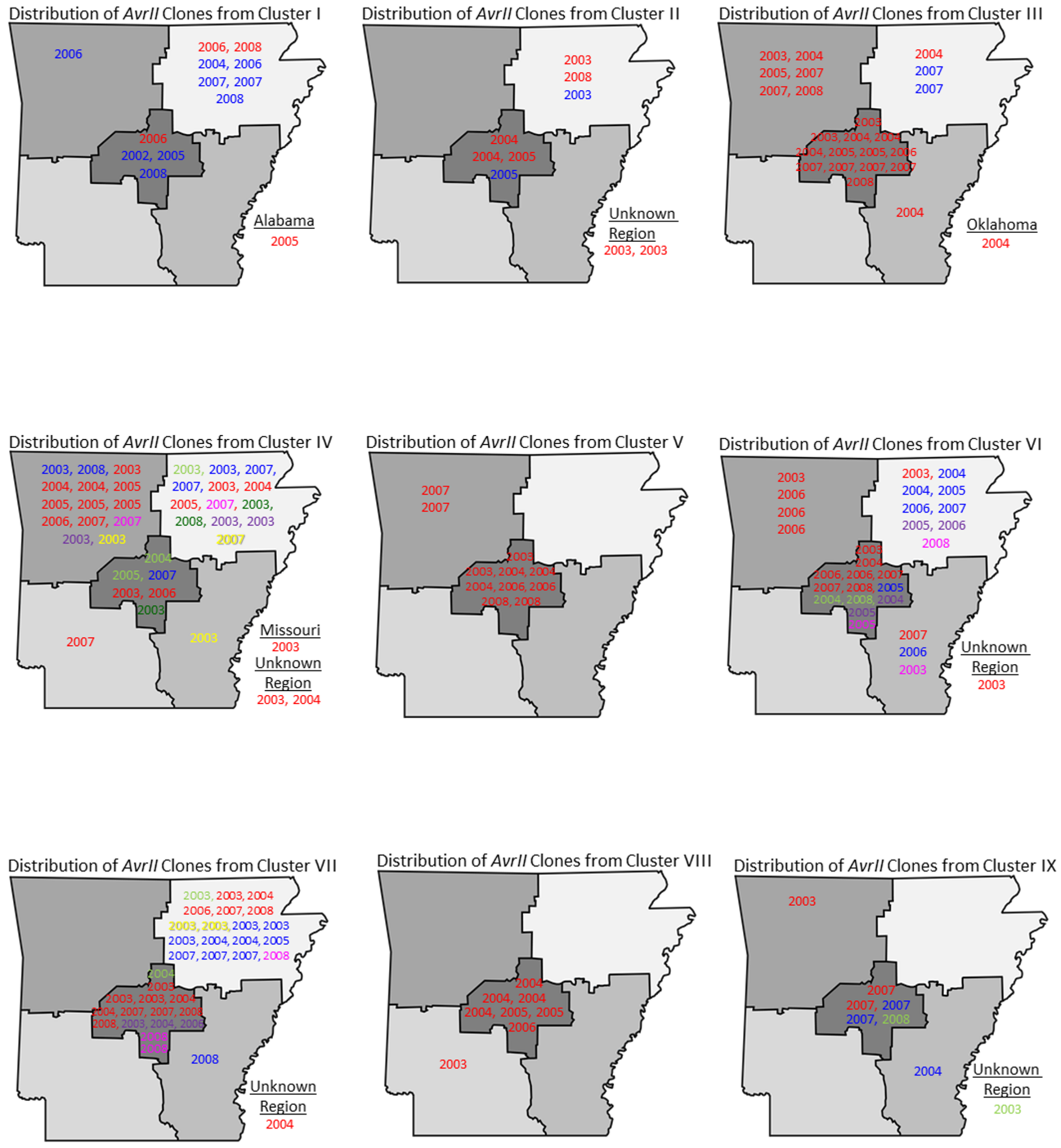
2.3. Distribution of Salmonella Javiana Genotypes
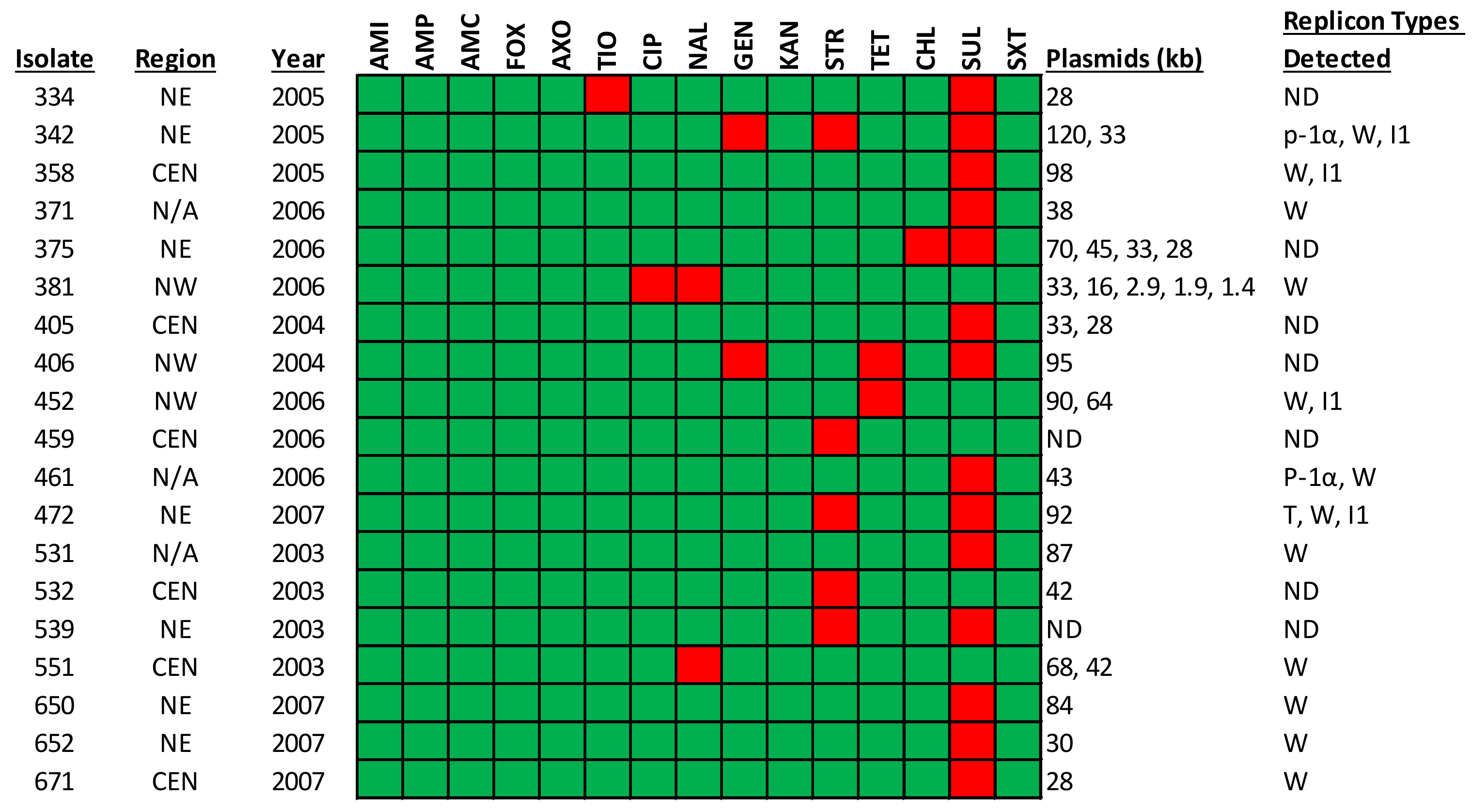
2.4. Plasmid Profiles
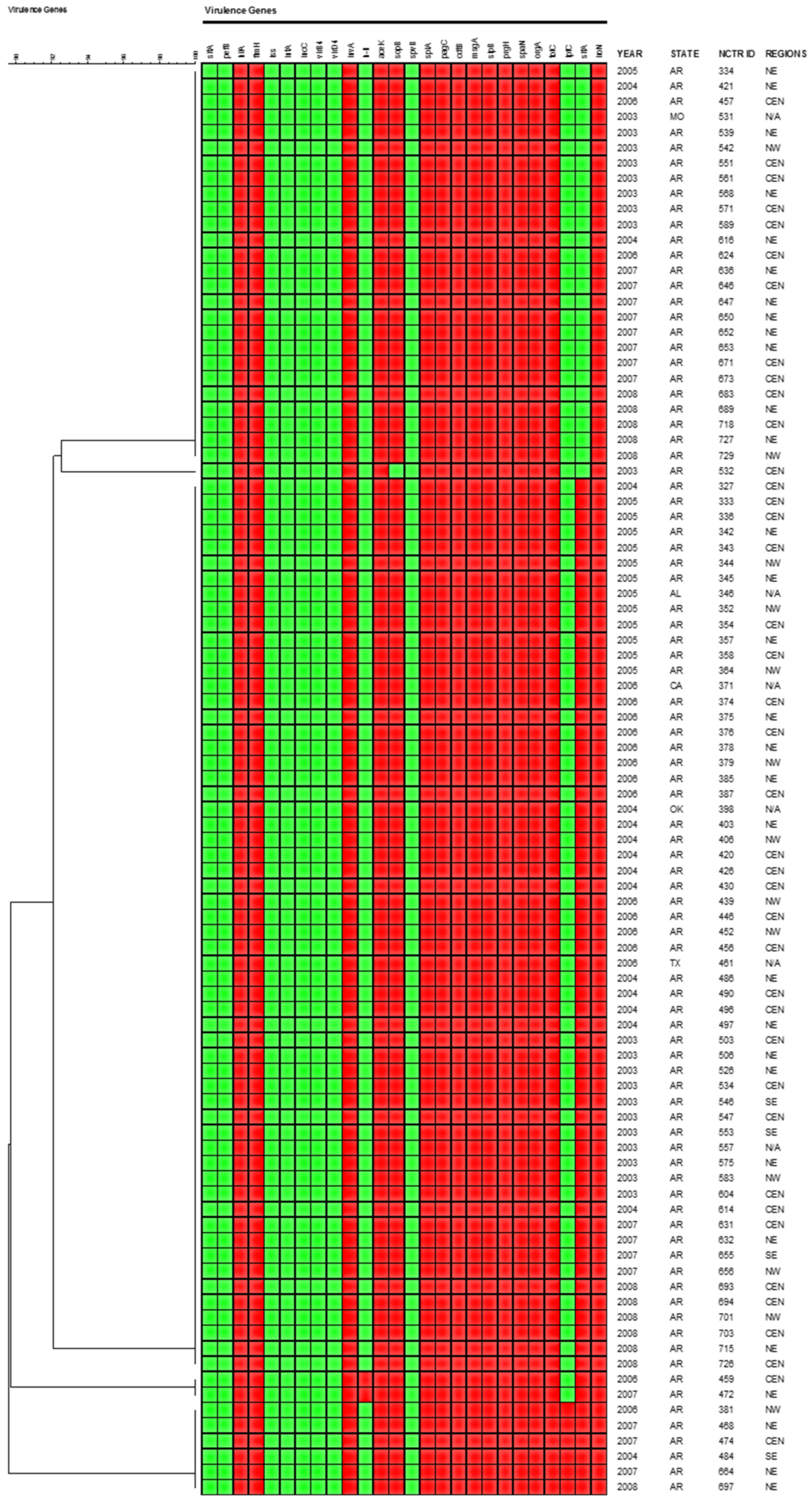
2.5. Virulence Gene Profiles
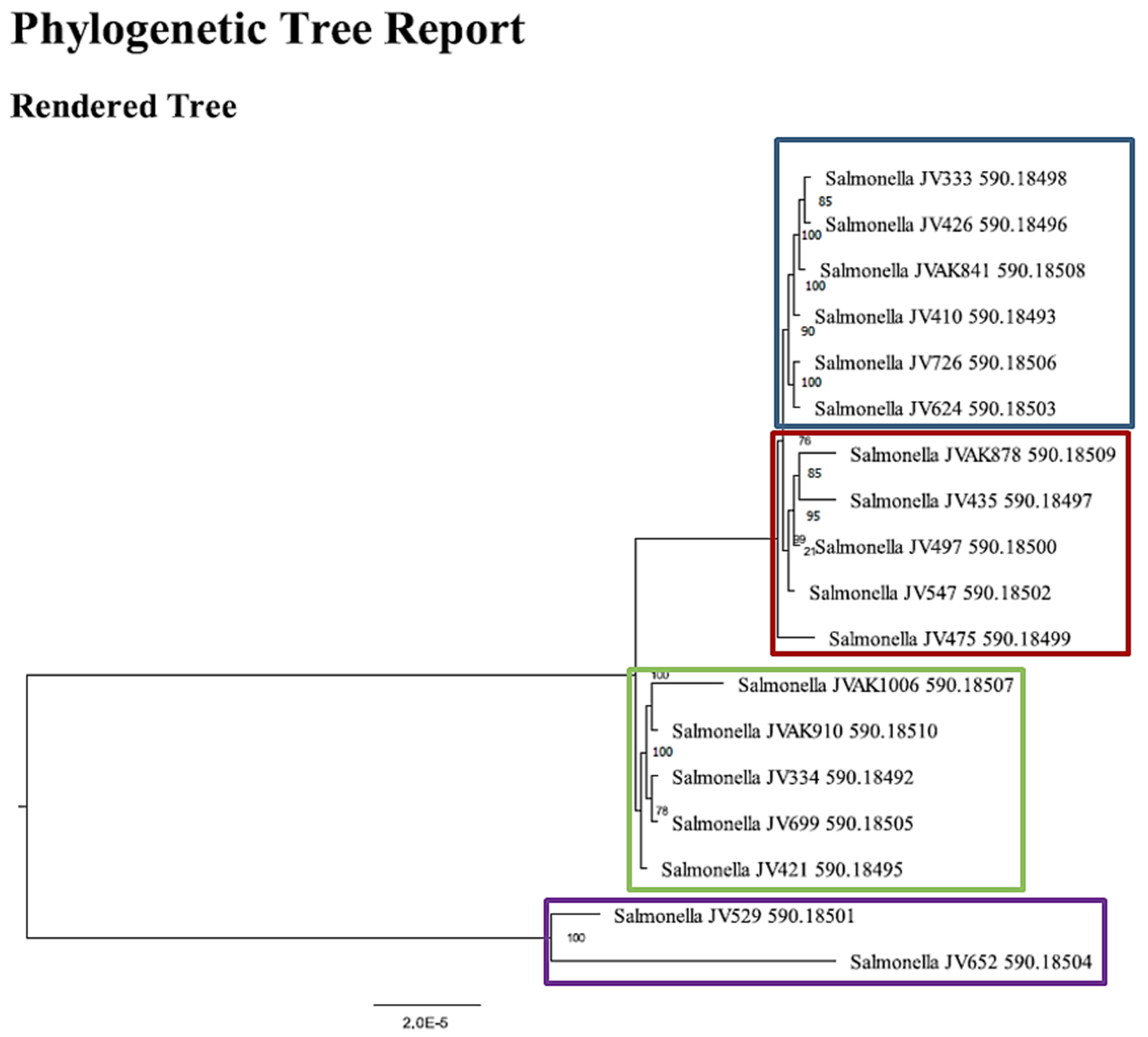
2.6. Whole Genome Sequence (WGS) Analysis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Pulsed-Field Gel Electrophoresis (PFGE)
4.3. Antimicrobial Susceptibility Testing (AST)
4.4. Plasmid Analysis and Replicon Typing
4.5. Detection of Virulence Genes
4.6. Whole Genome Sequencing (WGS)
4.7. Bioinformatics and Phylogenetic Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Centers for Disease Control and Prevention (CDC). National Salmonella Surveillance Annual Summary, 2022; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2022. [Google Scholar]
- Foley, S.L.; Nayak, R.; IHanning, B.; Johnson, T.J.; Han, J.; Ricke, S.C. Population dynamics of Salmonella enterica serotypes in commercial egg and poultry production. Appl. Environ. Microbiol. 2011, 77, 4273–4279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boore, A.L.; Hoekstra, R.M.; Iwamoto, M.; Fields, P.I.; Bishop, R.D.; Swerdlow, D.L. Salmonella enterica infections in the United States and assessment of coefficients of variation: A novel approach to identify epidemiologic characteristics of individual serotypes, 1996–2011. PLoS ONE 2015, 10, e0145416. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). National Enteric Disease Surveillance: Salmonella Annual Report, 2015; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2017. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). National Salmonella Surveillance Annual Summary, 2009; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2012. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Preliminary FoodNet Data on the incidence of infection with pathogens transmitted commonly through food—10 States, 2008. MMWR Morb. Mortal. Wkly. Rep. 2009, 58, 333–337. [Google Scholar]
- Lynch, M.F.; Tauxe, R.; Hedberg, C. The growing burden of foodborne outbreaks due to contaminated fresh produce: Risks and opportunities. Epidemiol. Infect. 2009, 137, 307–315. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). National Enteric Disease Surveillance: Salmonella Annual Report, 2019; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2020. [Google Scholar]
- Li, B.; Vellidis, G.; Liu, H.; Jay-Russell, M.; Zhao, S.; Hu, Z.; Wright, A.; Elkins, C.A. Diversity and antimicrobial resistance of Salmonella enterica isolates from surface water in Southeastern United States. Appl. Environ. Microbiol. 2014, 80, 6355–6365. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention (CDC). National Salmonella Surveillance Annual Summary, 2004; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2007. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Preliminary FoodNet data on the incidence of infection with pathogens transmitted commonly through food—10 sites, United States, 2004. MMWR Morb. Mortal. Wkly. Rep. 2005, 54, 352–356. [Google Scholar]
- Srikantiah, P.; Lay, J.C.; Hand, S.; Crump, J.A.; Campbell, J.; Van Duyne, M.S.; Bishop, R.; Middendor, R.; Currier, M.; Mead, P.S.; et al. Salmonella enterica serotype Javiana infections associated with amphibian contact, Mississippi, 2001. Epidemiol. Infect. 2004, 132, 273–281. [Google Scholar] [CrossRef]
- Marder Mph, E.P.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; Jervis, R.; Lathrop, S.; Muse, A.; Ryan, P.; Smith, K.; et al. Preliminary Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2006–2017. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 324–328. [Google Scholar] [CrossRef]
- Sandt, C.H.; Krouse, D.A.; Cook, C.R.; Hackman, A.L.; Chmielecki, W.A.; Warren, N.G. The key role of pulsed-field gel electrophoresis in investigation of a large multiserotype and multistate food-borne outbreak of Salmonella infections centered in Pennsylvania. J. Clin. Microbiol. 2006, 44, 3208–3212. [Google Scholar] [CrossRef] [Green Version]
- Srikantiah, P.; Bodager, D.; Toth, B.; Kass-Hout, T.; Hammond, R.; Stenzel, S.; Hoekstra, R.M.; Adams, J.; Van Duyne, S.; Mead, P.S. Web-based investigation of multistate salmonellosis outbreak. Emerg. Infect. Dis. 2005, 11, 610–612. [Google Scholar] [CrossRef] [Green Version]
- Hedberg, C.W.; Korlath, J.A.; D’Aoust, J.Y.; White, K.E.; Schell, W.L.; Miller, M.R.; Cameron, D.N.; MacDonald, K.L.; Osterholm, M.T. A multistate outbreak of Salmonella Javiana and Salmonella Oranienburg infections due to consumption of contaminated cheese. JAMA 1992, 268, 3203–3207. [Google Scholar] [CrossRef]
- Lee, R.; Peppe, J.; George, H. Pulsed-field gel electrophoresis of genomic digests demonstrates linkages among food, food handlers, and patrons in a food-borne Salmonella Javiana outbreak in Massachusetts. J. Clin. Microbiol. 1998, 36, 284–285. [Google Scholar] [CrossRef] [Green Version]
- Elward, A.; Grim, A.; Schroeder, P.; Kieffer, P.; Sellenriek, P.; Ferrett, R.; Adams, H.C.; Phillips, V.; Bartow, R.; Mays, D.; et al. Outbreak of Salmonella Javiana infection at a children’s hospital. Infect. Control Hosp. Epidemiol. 2006, 27, 586–592. [Google Scholar] [CrossRef]
- Jackson, B.R.; Griffin, P.M.; Cole, D.; Walsh, K.A.; Chai, S.J. Outbreak-associated Salmonella enterica serotypes and food commodities, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 1239–1244. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention (CDC). An Atlas of Salmonella in the United States, 1968–2011; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2013. [Google Scholar]
- Clarkson, L.S.; Tobin-D’Angelo, M.; Shuler, C.; Hanna, S.; Benson, J.; Voetsch, A.C. Sporadic Salmonella enterica serotype Javiana infections in Georgia and Tennessee: A hypothesis-generating study. Epidemiol. Infect. 2010, 138, 340–346. [Google Scholar] [CrossRef] [Green Version]
- Ackman, D.M.; Drabkin, P.; Birkhead, G.; Cieslak, P. Reptile-associated salmonellosis in New York State. Pediatr. Infect. Dis. J. 1995, 14, 955–959. [Google Scholar] [CrossRef]
- Mermin, J.; Hoar, B.; Angulo, F.J. Iguanas and Salmonella marina infection in children: A reflection of the increasing incidence of reptile-associated salmonellosis in the United States. Pediatrics 1997, 99, 399–402. [Google Scholar] [CrossRef]
- Lehmacher, A.; Bockemühl, J.; Aleksic, S. Nationwide outbreak of human salmonellosis in Germany due to contaminated paprika and paprika-powdered potato chips. Epidemiol. Infect. 1995, 115, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Scallan, E.; Mahon, B.E.; Hoekstra, R.M.; Griffin, P.M. Estimates of illnesses, hospitalizations and deaths caused by major bacterial enteric pathogens in young children in the United States. Pediatr. Infect. Dis. J. 2013, 32, 217–221. [Google Scholar] [CrossRef]
- Bell, R.L.; Zheng, J.; Burrows, E.; Allard, S.; Wang, C.Y.; Keys, C.E.; Melka, D.C.; Strain, E.; Luo, Y.; Allard, M.W.; et al. Ecological prevalence, genetic diversity, and epidemiological aspects of Salmonella isolated from tomato agricultural regions of the Virginia Eastern Shore. Front. Microbiol. 2015, 6, 415. [Google Scholar] [CrossRef]
- Mezal, E.H.; Stefanova, R.; Khan, A.A. Isolation and molecular characterization of Salmonella enterica serovar Javiana from food, environmental and clinical samples. Int. J. Food Microbiol. 2013, 164, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Micallef, S.A.; Rosenberg Goldstein, R.E.; George, A.; Kleinfelter, L.; Boyer, M.S.; McLaughlin, C.R.; Estrin, A.; Ewing, L.; Jean-Gilles Beaubrun, J.; Hanes, D.E.; et al. Occurrence and antibiotic resistance of multiple Salmonella serotypes recovered from water, sediment and soil on mid-Atlantic tomato farms. Environ. Res. 2012, 114, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lynne, A.M.; David, D.E.; Tang, H.; Xu, J.; Nayak, R.; Kaldhone, P.; Logue, C.M.; Foley, S.L. DNA sequence analysis of plasmids from multidrug resistant Salmonella enterica serotype Heidelberg isolates. PLoS ONE 2012, 7, e51160. [Google Scholar] [CrossRef] [PubMed]
- Kaldhone, P.R.; Han, J.; Deck, J.; Khajanchi, B.; Nayak, R.; Foley, S.L.; Ricke, S.C. Evaluation of the genetics and functionality of Incompatibility 1 (IncI1) plasmids from Salmonella enterica. Foodborne Pathog. Dis. 2018, 15, 168–176. [Google Scholar] [CrossRef]
- Nayak, R.; Stewart-King, T. Molecular epidemiological analysis and microbial source tracking of Salmonella enterica serovars in a preharvest turkey production environment. Foodborne Pathog. Dis. 2008, 5, 115–126. [Google Scholar] [CrossRef]
- Williams, K.; Gokulan, K.; Shelman, D.; Akiyama, T.; Khan, A.; Khare, S. Cytotoxic mechanism of cytolethal distending toxin in nontyphoidal Salmonella Serovar (Salmonella Javiana) during macrophage infection. DNA Cell Biol. 2015, 34, 113–124. [Google Scholar] [CrossRef]
- Miller, R.A.; Betteken, M.I.; Guo, X.; Altier, C.; Duhamel, G.E.; Wiedmann, M. The typhoid toxin produced by the nontyphoidal Salmonella enterica serotype Javiana is required for induction of a DNA damage response in vitro and systemic spread in vivo. mBio 2018, 9, e00467-18. [Google Scholar] [CrossRef] [Green Version]
- Hayward, M.R.; AbuOun, M.; La Ragione, R.M.; Tchórzewska, M.A.; Cooley, W.A.; Everest, D.J.; Petrovska, L.; Jansen, V.A.; Woodward, M.J. SPI-23 of S. Derby: Role in adherence and invasion of porcine tissues. PLoS ONE 2014, 9, e107857. [Google Scholar] [CrossRef] [Green Version]
- Egan, F.; Barret, M.; O’Gara, F. The SPI-1-like Type III secretion system: More roles than you think. Front. Plant Sci. 2014, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Jennings, E.; Thurston, T.L.M.; Holden, D.W. Salmonella SPI-2 Type III Secretion System Effectors: Molecular Mechanisms and Physiological Consequences. Cell Host Microbe 2017, 22, 217–231. [Google Scholar] [CrossRef]
- Rychlik, I.; Karasova, D.; Sebkova, A.; Volf, J.; Sisak, F.; Havlickova, H.; Kummer, V.; Imre, A.; Szmolka, A.; Nagy, B. Virulence potential of five major pathogenicity islands (SPI-1 to SPI-5) of Salmonella enterica serovar Enteritidis for chickens. BMC Microbiol. 2009, 9, 268. [Google Scholar] [CrossRef] [Green Version]
- Dorsey, C.W.; Laarakker, M.C.; Humphries, A.D.; Weening, E.H.; Bäumler, A.J. Salmonella enterica serotype Typhimurium MisL is an intestinal colonization factor that binds fibronectin. Mol. Microbiol. 2005, 57, 196–211. [Google Scholar] [CrossRef] [PubMed]
- Velásquez, J.C.; Hidalgo, A.A.; Villagra, N.; Santiviago, C.A.; Mora, G.C.; Fuentes, J.A. SPI-9 of Salmonella enterica serovar Typhi is constituted by an operon positively regulated by RpoS and contributes to adherence to epithelial cells in culture. Microbiology 2016, 162, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Adkins, J.N.; Coleman, J.R.; Schepmoes, A.A.; Dohnkova, A.; Mottaz, H.M.; Norbeck, A.D.; Purvine, S.O.; Manes, N.P.; Smallwood, H.S.; et al. Proteomic Analysis of Salmonella enterica Serovar Typhimurium Isolated from RAW 264.7 Macrophages. J. Biol. Chem. 2006, 281, 29131–29140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Feng, L.; Yang, B.; Zhang, W.; Wang, P.; Jiang, X.; Wang, L. Signal transduction pathway mediated by the novel regulator LoiA for low oxygen tension induced Salmonella Typhimurium invasion. PLoS Pathog. 2017, 13, e1006429. [Google Scholar] [CrossRef] [Green Version]
- Aljahdali, N.H.; Khajanchi, B.K.; Weston, K.; Deck, J.; Cox, J.; Singh, R.; Gilbert, J.; Sanad, Y.M.; Han, J.; Nayak, R.; et al. Genotypic and phenotypic characterization of incompatibility group FIB positive Salmonella enterica serovar typhimurium isolates from food animal sources. Genes 2020, 11, 1307. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, N.; Nolan, V.G.; Dunn, J.R.; Banerjee, P. Sources of human infection by Salmonella enterica serotype Javiana: A systematic review. PLoS ONE 2019, 14, e0222108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk and Diffusion Susceptibility Testing for Bacteria Isolated from Animals; Approved Standard-Second Edition (M31-A2); National Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2002. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; Twelfth Informational Supplement (M100-S12); National Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2002. [Google Scholar]
- Johnson, T.J.; Nolan, L.K. Plasmid replicon typing. Methods Mol. Biol. 2009, 551, 27–35. [Google Scholar]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Courtney, S.; Mossoba, M.E.; Hammack, T.S.; Keys, C.; Al-Khaldi, S.F. Using PCR amplification to increase the confidence level of Salmonella typhimurium DNA microarray chip hybridization. Mol. Cell. Probes 2006, 20, 163–171. [Google Scholar] [CrossRef]
- Dezfulian, H.; Batisson, I.; Fairbrother, J.M.; Lau, P.C.; Nassar, A.; Szatmari, G.; Harel, J. Presence and characterization of extraintestinal pathogenic Escherichia coli virulence genes in F165-positive E. coli strains isolated from diseased calves and pigs. J. Clin. Microbiol. 2003, 41, 1375–1385. [Google Scholar] [CrossRef] [Green Version]
- Foley, S.L.; White, D.G.; McDermott, P.F.; Walker, R.D.; Rhodes, B.S.; Simjee, S.; Zhao, S. Comparison of Subtyping Methods for Salmonella enterica serovar Typhimurium of Food Animal Origins. J. Clin. Microbiol. 2006, 44, 3569–3577. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Gokulan, K.; Barnette, D.; Khare, S.; Rooney, A.W.; Deck, J.; Nayak, R.; Stefanova, R.; Hart, M.E.; Foley, S.L. Evaluation of virulence and antimicrobial resistance in Salmonella enterica serovar Enteritidis isolates from humans and chicken- and egg-associated sources. Foodborne Pathog. Dis. 2013, 10, 1008–1015. [Google Scholar] [CrossRef]
- Nayak, R.; Stewart, T.; Wang, R.F.; Lin, J.; Cerniglia, C.E.; Kenney, P.B. Genetic diversity and virulence gene determinants of antibiotic-resistant Salmonella isolated from preharvest turkey production sources. Int. J. Food Microbiol. 2004, 91, 51–62. [Google Scholar] [CrossRef]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Vet. Res. 2005, 36, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Skyberg, J.A.; Logue, C.M.; Nolan, L.K. Virulence genotyping of Salmonella spp. with multiplex PCR. Avian Dis. 2006, 50, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Zou, W.; Al-Khaldi, S.F.; Branham, W.S.; Han, T.; Fuscoe, J.C.; Han, J.; Foley, S.L.; Cerniglia, C.E.; Nayak, R. Microarray analysis of virulence genes in Salmonella from poultry production environments and an outbreak investigation. J. Infect. Dev. Ctries. 2011, 5, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Wattam, A.R.; Abraham, D.; Dalay, O.; Disz, T.L.; Driscoll, T.; Gabbard, J.L.; Gillespie, J.J.; Gough, R.; Hix, D.; Kenyon, R.; et al. Patric, the bacterial bioinformatics database and analysis resource. Nucleic Acids Res. 2014, 42, D581–D591. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| STATE | Isolates | Percentage | AR REGION | Isolates | Percentage |
|---|---|---|---|---|---|
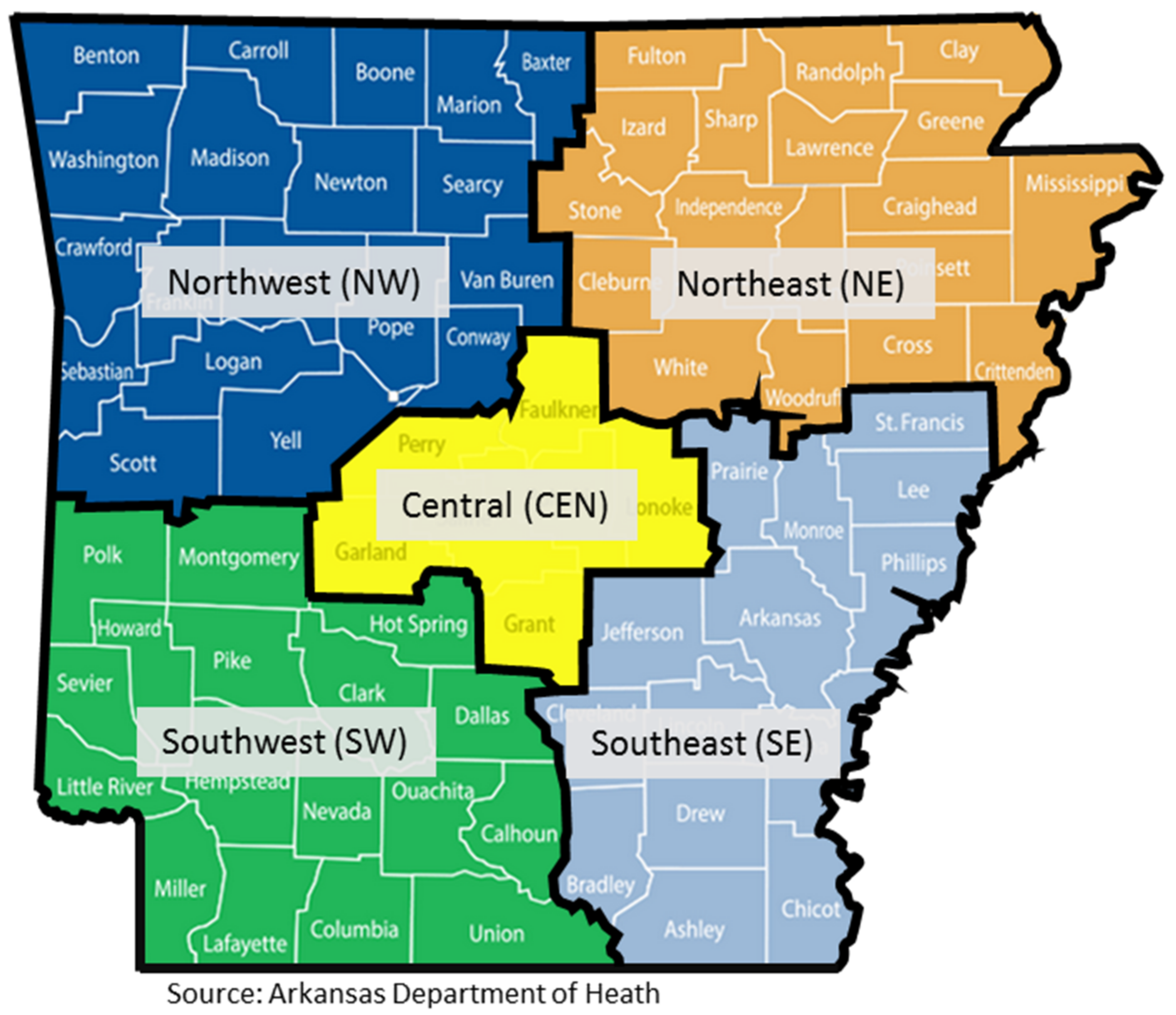
| Arkansas | 394 | 96.3 | Central | 168 | 42.6 |
| Alabama | 1 | 0.2 | Northeast | 134 | 34.0 |
| California | 2 | 0.5 | Northwest | 57 | 14.5 |
| Missouri | 2 | 0.5 | Southeast | 14 | 3.6 |
| Oklahoma | 4 | 1.0 | Southwest | 13 | 3.3 |
| Texas | 1 | 0.2 | Unknown | 8 | 2.0 |
| Unknown | 5 | 1.2 | Total | 394 | 100 |
| Total | 409 | 100 |
| SEX | Isolates | Percentage | AGE | Isolates | Percentage | YEAR | Isolates | Percentage |
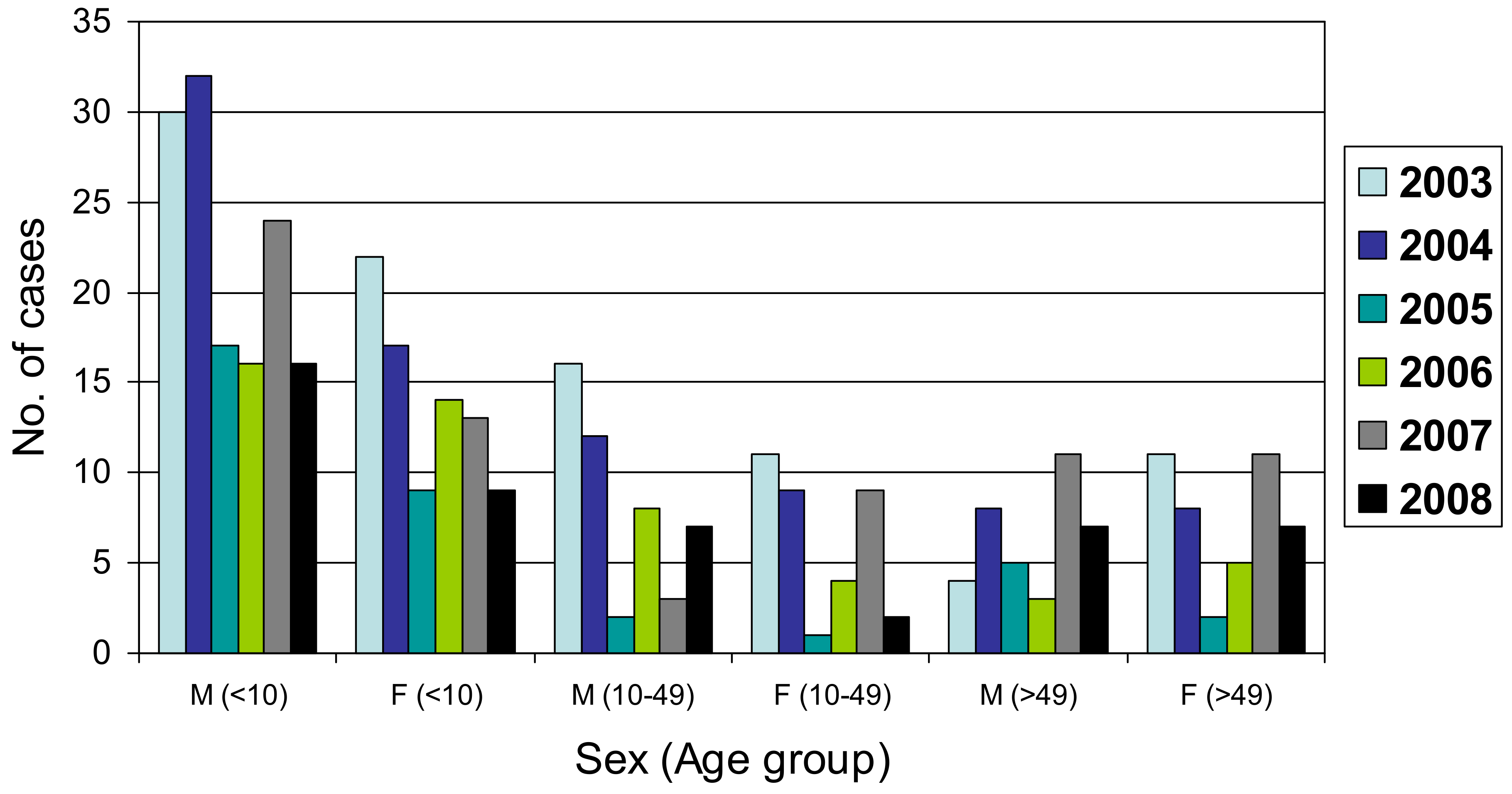
|---|---|---|---|---|---|---|---|---|
| Female | 177 | 43.3 | <1 to 10 | 221 | 54.0 | 2003 | 101 | 24.7 |
| Male | 231 | 56.5 | 11 to 20 | 29 | 7.1 | 2004 | 96 | 23.5 |
| Unknown | 1 | 0.2 | 21 to 30 | 15 | 3.7 | 2005 | 42 | 10.3 |
| Total | 409 | 100 | 31 to 40 | 20 | 4.9 | 2006 | 49 | 12.0 |
| 41 to 50 | 18 | 4.4 | 2007 | 71 | 17.4 | |||
| 51 to 60 | 28 | 6.8 | 2008 | 50 | 12.2 | |||
| 61 to 70 | 22 | 5.4 | Total | 409 | 100 | |||
| 71 to 80 | 23 | 5.6 | ||||||
| 81 to 90 | 6 | 1.5 | ||||||
| Unknown | 27 | 6.6 | ||||||
| Total | 409 | 100 |
| Antimicrobial Resistance Profile | Number of Isolates | % Prevalence |
|---|---|---|
| Pan-susceptible | 34 | 8 |
| SUL only | 364 | 89 |
| SUL, STR, GEN | 1 | <1 |
| SUL, TET, GEN | 1 | <1 |
| SUL, CHL | 1 | <1 |
| SUL, STR | 2 | <1 |
| SUL, TIO | 1 | <1 |
| NAL, CIP | 1 | <1 |
| TET | 1 | <1 |
| STR | 2 | <1 |
| NAL | 1 | <1 |
| TOTAL | 409 | 100 |
| Isolates | Location | Virulence Genes | SPI Genes | Resistance Genes | Phenotype |
|---|---|---|---|---|---|
| JV-333 | AR, CEN | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJ | SPI-2, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| JV-334 | AR, NE | sopAD2, fimYD, pagO, sitA, iroB, pipBACD, sspA, rfaJI | SPI-2, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV410 | MI | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJ | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV421 | AR, NE | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJI | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV426 | AR, CEN | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJ | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV435 | GA | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaI | SPI-1, SPI-2, SPI-3, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV475 | USA | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJI | SPI-1, SPI-2, SPI-3, SPI-4, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV497 | AR, NE | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV529 | AR, NE | sopAD2, fimYD, pagO, sitA, iroB, pipBACD, sspA rfaI, | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV547 | AR, CEN | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJI | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV624 | AR, CEN | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJ | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV652 | AR, NE | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJI | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV699 | USA | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaI | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV726 | AR, CEN | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJ | SPI-1, SPI-2, SPI-3, SPI-4, SPI-5, SPI-9, SPI-13, SPI-14, C63PI | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV841 | AR, NE | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJ | N/A | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV878 | AR, NE | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJI | N/A | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV910 | AR, NE | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJI | N/A | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin | ||||
| JV1006 | AR, NE | sopAD2, fimYD, pagOD, sitA, iroB, pipBACD, sspA, rfaJI | N/A | aac(6’)-Iaa | Aminoglycoside resistance |
| parC p.T57S | Nalidixic acid, Ciprofloxacin |
| Isolate | Source | Isolation Location | Year | GenBank Accession |
|---|---|---|---|---|
| 333 | Clinical | AR | 2005 | JAHWXF000000000 |
| 334 | Clinical | AR | 2005 | JAHWXE000000000 |
| 410 | Clinical | MI | 1999 | JAHWXD000000000 |
| 421 | Clinical | AR | 2004 | JAHWXC000000000 |
| 426 | Clinical | AR | 2004 | JAHWXB000000000 |
| 435 | Clinical | GA | 1999 | JAHWXA000000000 |
| 475 | Clinical | USA | 1999 | JAHWWZ000000000 |
| 497 | Clinical | AR | 2004 | JAHWWY000000000 |
| 529 | Clinical | AR | 2003 | JAHWWX000000000 |
| 547 | Clinical | AR | 2003 | JAHWWW000000000 |
| 624 | Clinical | AR | 2006 | JAHWWV000000000 |
| 652 | Clinical | AR | 2007 | JAHWWU000000000 |
| 699 | Clinical | USA | 1999 | JAHWWT000000000 |
| 726 | Clinical | AR | 2008 | JAHWWS000000000 |
| 841 | Clinical | AR | 2011 | JAHWWR000000000 |
| 878 | Clinical | AR | 2011 | JAHWWQ000000000 |
| 910 | Clinical | AR | 2011 | JAHWWP000000000 |
| 1006 | Clinical | AR | 2011 | JAHWWO000000000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanad, Y.M.; Aljahdali, N.H.; Khajanchi, B.K.; Nayak, R.; Khan, A.; Foley, S.L. Microbial Genetics and Clonal Dissemination of Salmonella enterica Serotype Javiana Isolated from Human Populations in Arkansas, USA. Pathogens 2022, 11, 1192. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11101192
Sanad YM, Aljahdali NH, Khajanchi BK, Nayak R, Khan A, Foley SL. Microbial Genetics and Clonal Dissemination of Salmonella enterica Serotype Javiana Isolated from Human Populations in Arkansas, USA. Pathogens. 2022; 11(10):1192. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11101192
Chicago/Turabian StyleSanad, Yasser M., Nesreen H. Aljahdali, Bijay K. Khajanchi, Rajesh Nayak, Ashraf Khan, and Steven L. Foley. 2022. "Microbial Genetics and Clonal Dissemination of Salmonella enterica Serotype Javiana Isolated from Human Populations in Arkansas, USA" Pathogens 11, no. 10: 1192. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11101192