
Cryopreservation of Plasmodium Sporozoites

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Parasites
2.2. Determining Sporozoite Integrity and Viability
2.3. In Vitro and In Vivo P. berghei and P. yoelii Sporozoite Infections
2.4. Flow Cytometry
2.5. Luminescence Signal Imaging
2.6. Primary Human Hepatocyte Culture for P. falciparum and P. vivax Infection
2.7. P. falciparum and P. vivax Sporozoite Infection and Quantification
2.8. Radiation Attenuated Sporozoite (RAS) Vaccination
2.9. Statistical Analysis
3. Results
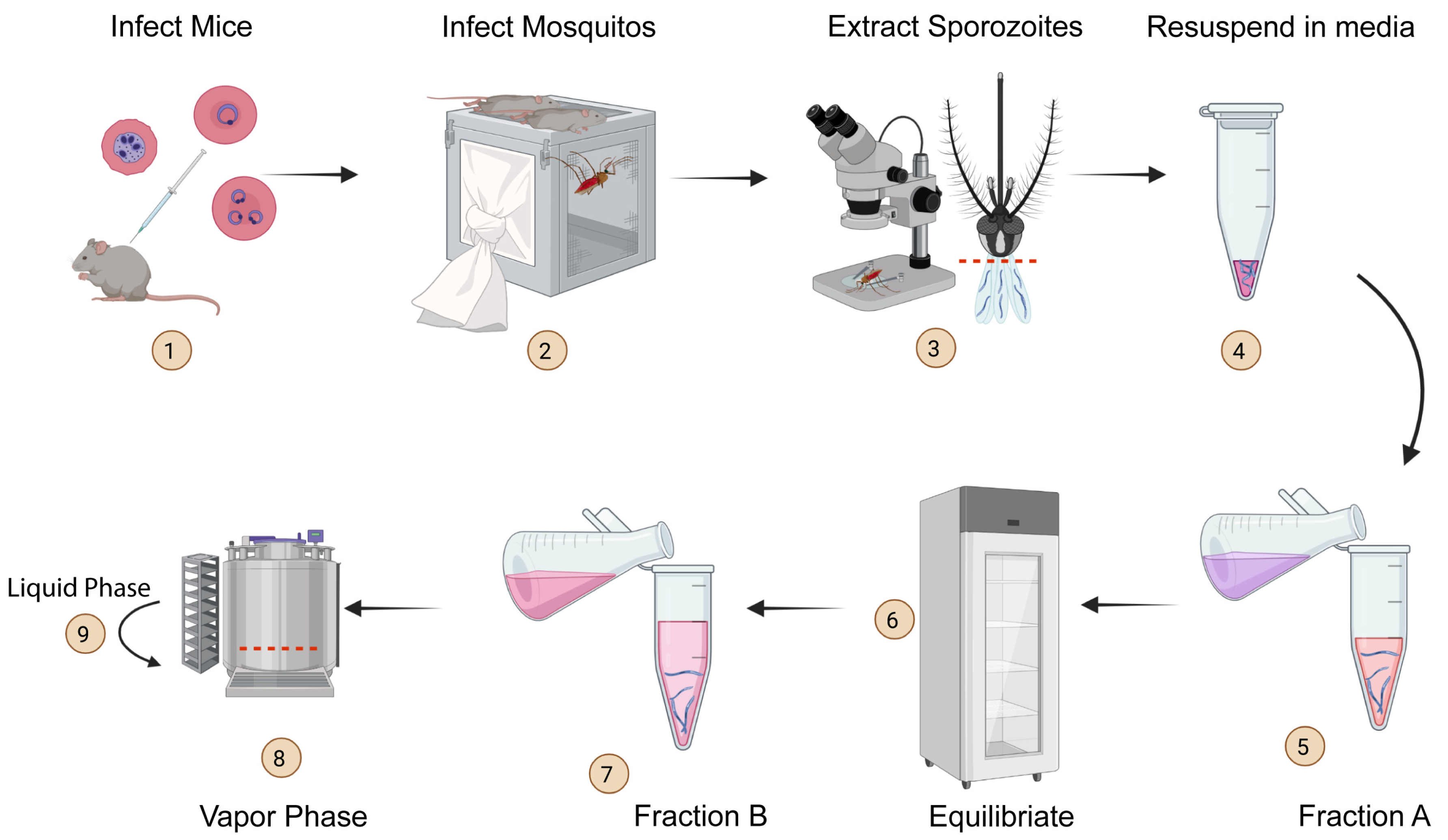
3.1. Cryopreservation of Plasmodium Sporozoites
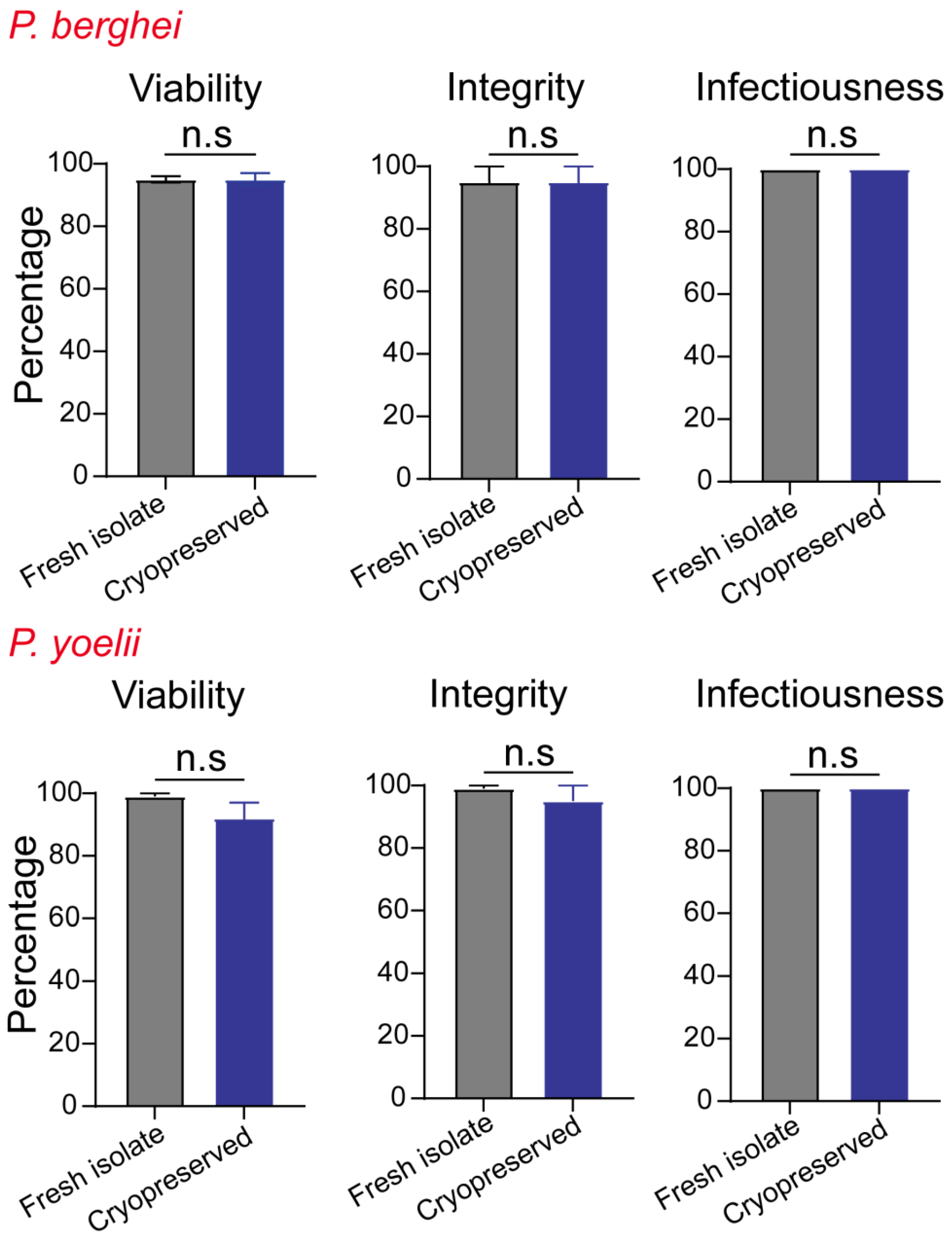
3.2. The Metrics of Successful Cryopreservation and Recovery
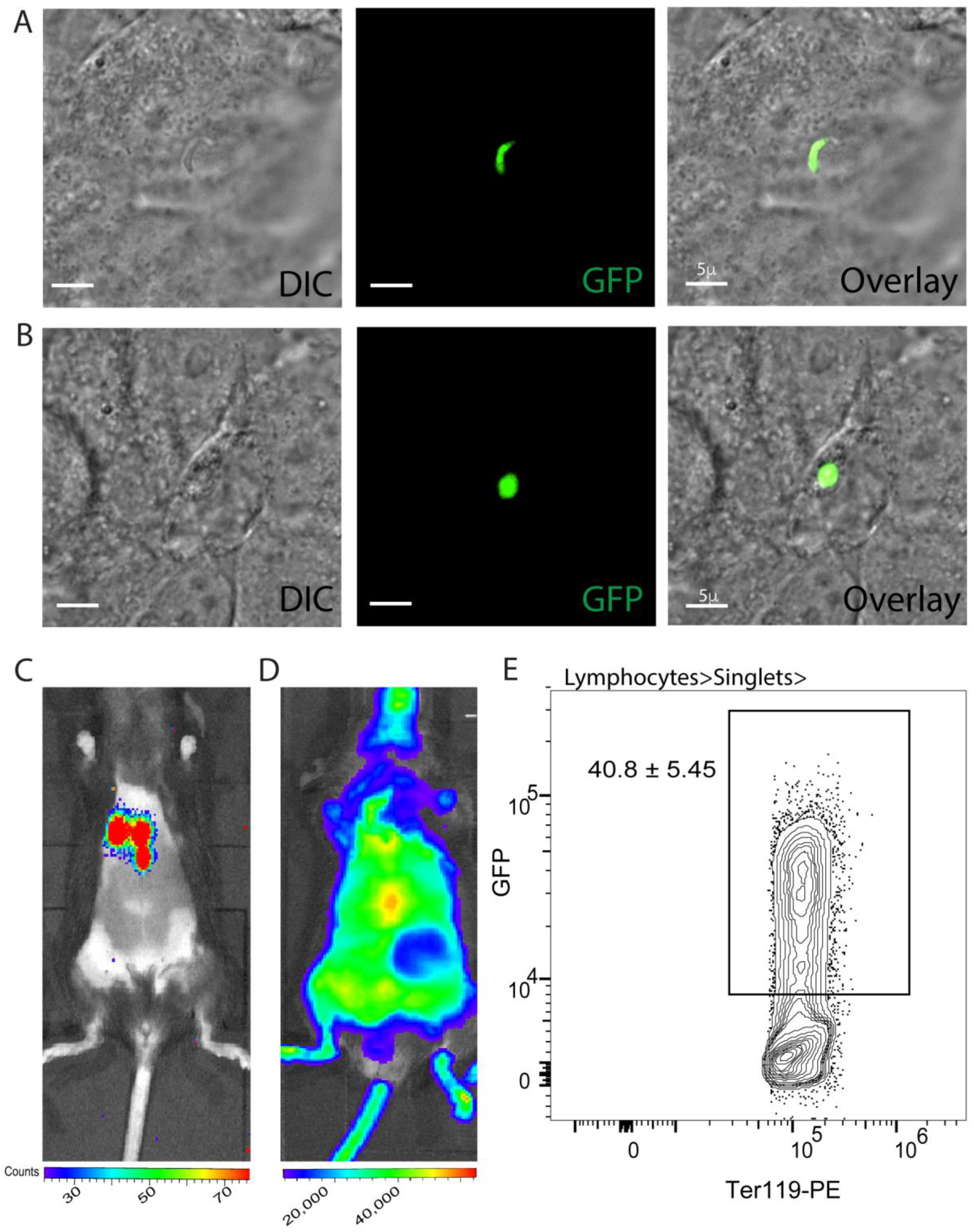
3.3. Effective Cryopreservation and Recovery of Plasmodium Sporozoites
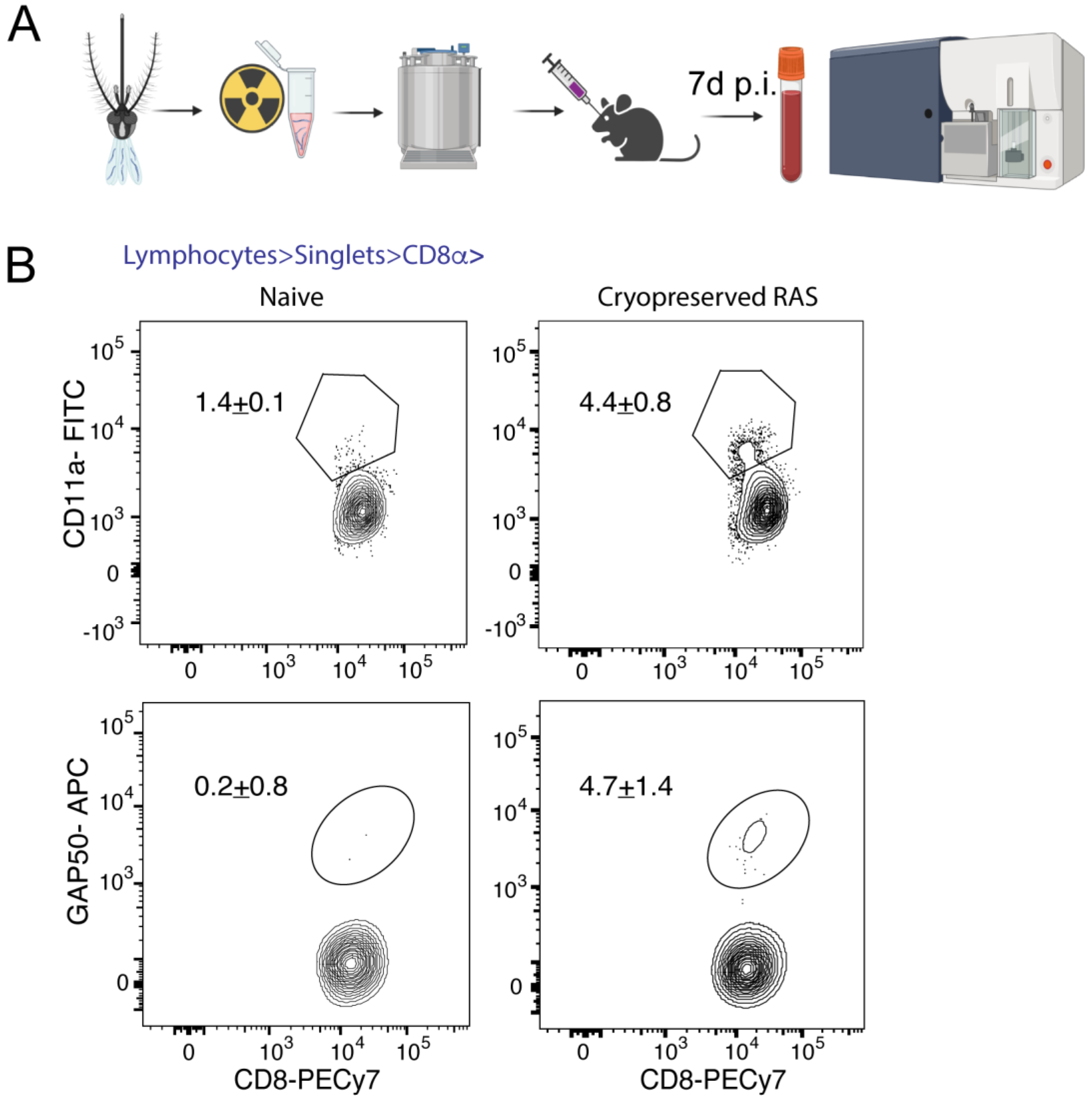
3.4. Immunization with Cryopreserved Radiation Attenuated Sporozoites
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021.
- Djimde, A.A.; Makanga, M.; Kuhen, K.; Hamed, K. The emerging threat of artemisinin resistance in malaria: Focus on artemether-lumefantrine. Expert. Rev. Anti. Infect. Ther. 2015, 13, 1031–1045. [Google Scholar] [CrossRef] [PubMed]
- Marques-da-Silva, C.; Peissig, K.; Kurup, S.P. Pre-Erythrocytic Vaccines against Malaria. Vaccines 2020, 8, 400. [Google Scholar] [CrossRef] [PubMed]
- RTS,S Clinical Trials Partnership. Efficacy and safety of RTS,S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: Final results of a phase 3, individually randomised, controlled trial. Lancet 2015, 386, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and Disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Franky, J.; Cuy-Chaparro, L.; Camargo, A.; Reyes, C.; Gomez, M.; Salamanca, D.R.; Patarroyo, M.A.; Patarroyo, M.E. Plasmodium falciparum pre-erythrocytic stage vaccine development. Malar. J. 2020, 19, 56. [Google Scholar] [CrossRef] [Green Version]
- Duffy, P.E.; Patrick Gorres, J. Malaria vaccines since 2000: Progress, priorities, products. NPJ Vaccines 2020, 5, 48. [Google Scholar] [CrossRef]
- Nussenzweig, R.S.; Vanderberg, J.; Most, H.; Orton, C. Protective immunity produced by the injection of x-irradiated sporozoites of plasmodium berghei. Nature 1967, 216, 160–162. [Google Scholar] [CrossRef]
- Clyde, D.F. Immunization of man against falciparum and vivax malaria by use of attenuated sporozoites. Am. J. Trop. Med. Hyg. 1975, 24, 397–401. [Google Scholar] [CrossRef]
- Kramer, L.D.; Vanderberg, J.P. Intramuscular immunization of mice with irradiated Plasmodium berghei sporozoites. Enhancement of protection with albumin. Am. J. Trop. Med. Hyg. 1975, 24, 913–916. [Google Scholar] [CrossRef]
- Hill, A.V. Pre-erythrocytic malaria vaccines: Towards greater efficacy. Nat. Rev. Immunol. 2006, 6, 21–32. [Google Scholar] [CrossRef]
- malERA Consultative Group on Vaccines. A research agenda for malaria eradication: Vaccines. PLoS Med. 2011, 8, e1000398. [Google Scholar] [CrossRef]
- Hoffman, S.L.; Vekemans, J.; Richie, T.L.; Duffy, P.E. The march toward malaria vaccines. Vaccine 2015, 33 (Suppl. S4), D13–D23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itsara, L.S.; Zhou, Y.; Do, J.; Grieser, A.M.; Vaughan, A.M.; Ghosh, A.K. The Development of Whole Sporozoite Vaccines for Plasmodium falciparum Malaria. Front. Immunol. 2018, 9, 2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meneghel, J.; Kilbride, P.; Morris, G.J. Cryopreservation as a Key Element in the Successful Delivery of Cell-Based Therapies-A Review. Front. Med. 2020, 7, 592242. [Google Scholar] [CrossRef]
- Whaley, D.; Damyar, K.; Witek, R.P.; Mendoza, A.; Alexander, M.; Lakey, J.R. Cryopreservation: An Overview of Principles and Cell-Specific Considerations. Cell Transplant. 2021, 30, 963689721999617. [Google Scholar] [CrossRef]
- Pegg, D.E. The history and principles of cryopreservation. Semin. Reprod. Med. 2002, 20, 5–13. [Google Scholar] [CrossRef]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; Di Nardo, M.; Adiga, S.K.; Talevi, R. Mitochondrial Dysfunction and Oxidative Stress Caused by Cryopreservation in Reproductive Cells. Antioxidants 2021, 10, 337. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Marin-Mogollon, C.; Imai, T.; Mendes, A.M.; van der Laak, R.; Sturm, A.; Geurten, F.J.A.; Miyazaki, S.; Chevalley-Maurel, S.; Ramesar, J.; et al. Generation of a Genetically Modified Chimeric Plasmodium falciparum Parasite Expressing Plasmodium vivax Circumsporozoite Protein for Malaria Vaccine Development. Front. Cell Infect. Microbiol. 2020, 10, 591046. [Google Scholar] [CrossRef]
- Jeffery, G.M.; Rendtorff, R.C. Preservation of viable human malaria sporozoites by low-temperature freezing. Exp. Parasitol. 1955, 4, 445–454. [Google Scholar] [CrossRef]
- Weathersby, A.B.; McCall, J.W. Survival of sporozoites of Plasmodium gallinaceum Brumpt for 767 days in liquid nitrogen (-197 C). J. Parasitol. 1967, 53, 638–640. [Google Scholar] [CrossRef]
- Leef, J.L.; Strome, C.P.; Beaudoin, R.L. Low-temperature preservation of sporozoites of Plasmodium berghei. Bull. World Health Organ. 1979, 57 (Suppl. S1), 87–91. [Google Scholar] [PubMed]
- Orjih, A.U.; Nussenzweig, R.S. Immunization against rodent malaria with cryopreserved irradiated sporozoites of Plasmodium berghei. Am. J. Trop. Med. Hyg. 1980, 29, 343–347. [Google Scholar] [CrossRef]
- Singh, N.; Barnes, S.J.; Jenwithisuk, R.; Sattabongkot, J.; Adams, J.H. A simple and efficient method for cryopreservation and recovery of viable Plasmodium vivax and P. falciparum sporozoites. Parasitol. Int. 2016, 65, 552–557. [Google Scholar] [CrossRef] [Green Version]
- Chakravarty, S.; Shears, M.J.; James, E.R.; Rai, U.; Kc, N.; Conteh, S.; Lambert, L.E.; Duffy, P.E.; Murphy, S.C.; Hoffman, S.L. Efficient infection of non-human primates with purified, cryopreserved Plasmodium knowlesi sporozoites. Malar. J. 2022, 21, 247. [Google Scholar] [CrossRef] [PubMed]
- Ruben, A.; Awe, A.; Hoffman, S.L.; Sim, B.K.L.; Billingsley, P.F.; Manoj, A.; Chakravarty, S.; Gunasekera, A.; Li, T.; Stafford, R. 157 Cryopreservation of Plasmodium falciparum sporozoites and Sanaria® PfSPZ vaccine. Cryobiology 2013, 67, 442. [Google Scholar] [CrossRef]
- Marques-da-Silva, C.; Peissig, K.; Walker, M.P.; Shiau, J.; Bowers, C.; Kyle, D.E.; Vijay, R.; Lindner, S.E.; Kurup, S.P. Direct type I interferon signaling in hepatocytes controls malaria. Cell Rep. 2022, 40, 111098. [Google Scholar] [CrossRef]
- Pathak, A.K.; Shiau, J.C.; Franke-Fayard, B.; Shollenberger, L.M.; Harn, D.A.; Kyle, D.E.; Murdock, C.C. Streamlining sporozoite isolation from mosquitoes by leveraging the dynamics of migration to the salivary glands. Malar. J. 2022, 21, 264. [Google Scholar] [CrossRef]
- Maher, S.P.; Vantaux, A.; Cooper, C.A.; Chasen, N.M.; Cheng, W.T.; Joyner, C.J.; Manetsch, R.; Witkowski, B.; Kyle, D. A Phenotypic Screen for the Liver Stages of Plasmodium vivax. Bio Protoc. 2021, 11, e4253. [Google Scholar] [CrossRef]
- Kurup, S.P.; Anthony, S.M.; Hancox, L.S.; Vijay, R.; Pewe, L.L.; Moioffer, S.J.; Sompallae, R.; Janse, C.J.; Khan, S.M.; Harty, J.T. Monocyte-Derived CD11c(+) Cells Acquire Plasmodium from Hepatocytes to Prime CD8 T Cell Immunity to Liver-Stage Malaria. Cell Host Microbe 2019, 25, 565–577.e6. [Google Scholar] [CrossRef]
- Crowley, L.C.; Scott, A.P.; Marfell, B.J.; Boughaba, J.A.; Chojnowski, G.; Waterhouse, N.J. Measuring Cell Death by Propidium Iodide Uptake and Flow Cytometry. Cold Spring Harbor Protoc. 2016, 2016. [Google Scholar] [CrossRef]
- Kurup, S.P.; Obeng-Adjei, N.; Anthony, S.M.; Traore, B.; Doumbo, O.K.; Butler, N.S.; Crompton, P.D.; Harty, J.T. Regulatory T cells impede acute and long-term immunity to blood-stage malaria through CTLA-4. Nat. Med. 2017, 23, 1220–1225. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.L.; Murray, S.; Vaughan, A.M.; Harupa, A.; Sack, B.; Baldwin, M.; Crispe, I.N.; Kappe, S.H. Quantitative bioluminescent imaging of pre-erythrocytic malaria parasite infection using luciferase-expressing Plasmodium yoelii. PLoS ONE 2013, 8, e60820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, A.; Maher, S.P.; Conway, A.J.; Ubalee, R.; Chaumeau, V.; Andolina, C.; Kaba, S.A.; Vantaux, A.; Bakowski, M.A.; Thomson-Luque, R.; et al. A comprehensive model for assessment of liver stage therapies targeting Plasmodium vivax and Plasmodium falciparum. Nat. Commun. 2018, 9, 1837. [Google Scholar] [CrossRef] [Green Version]
- Maher, S.P.; Vantaux, A.; Chaumeau, V.; Chua, A.C.Y.; Cooper, C.A.; Andolina, C.; Peneau, J.; Rouillier, M.; Rizopoulos, Z.; Phal, S.; et al. Probing the distinct chemosensitivity of Plasmodium vivax liver stage parasites and demonstration of 8-aminoquinoline radical cure activity in vitro. Sci. Rep. 2021, 11, 19905. [Google Scholar] [CrossRef] [PubMed]
- Schafer, C.; Dambrauskas, N.; Steel, R.W.; Carbonetti, S.; Chuenchob, V.; Flannery, E.L.; Vigdorovich, V.; Oliver, B.G.; Roobsoong, W.; Maher, S.P.; et al. A recombinant antibody against Plasmodium vivax UIS4 for distinguishing replicating from dormant liver stages. Malar. J. 2018, 17, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, N.W.; Butler, N.S.; Badovinac, V.P.; Harty, J.T. Extreme CD8 T cell requirements for anti-malarial liver-stage immunity following immunization with radiation attenuated sporozoites. PLoS Pathog. 2010, 6, e1000998. [Google Scholar] [CrossRef] [Green Version]
- Tasdemir, U.; Buyukleblebici, S.; Tuncer, P.B.; Coskun, E.; Ozgurtas, T.; Aydin, F.N.; Buyukleblebici, O.; Gurcan, I.S. Effects of various cryoprotectants on bull sperm quality, DNA integrity and oxidative stress parameters. Cryobiology 2013, 66, 38–42. [Google Scholar] [CrossRef]
- Mazur, P. The role of intracellular freezing in the death of cells cooled at supraoptimal rates. Cryobiology 1977, 14, 251–272. [Google Scholar] [CrossRef]
- Lefebvre, M.N.; Surette, F.A.; Anthony, S.M.; Vijay, R.; Jensen, I.J.; Pewe, L.L.; Hancox, L.S.; Van Braeckel-Budimir, N.; van de Wall, S.; Urban, S.L.; et al. Expeditious recruitment of circulating memory CD8 T cells to the liver facilitates control of malaria. Cell Rep. 2021, 37, 109956. [Google Scholar] [CrossRef]
- Kurup, S.P.; Butler, N.S.; Harty, J.T. T cell-mediated immunity to malaria. Nat. Rev. Immunol. 2019, 19, 457–471. [Google Scholar] [CrossRef]
- Fernandez-Ruiz, D.; Ng, W.Y.; Holz, L.E.; Ma, J.Z.; Zaid, A.; Wong, Y.C.; Lau, L.S.; Mollard, V.; Cozijnsen, A.; Collins, N.; et al. Liver-Resident Memory CD8(+) T Cells Form a Front-Line Defense against Malaria Liver-Stage Infection. Immunity 2016, 45, 889–902. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Perez, G.P.; Legarda, A.; Munoz, J.; Sim, B.K.; Ballester, M.R.; Dobano, C.; Moncunill, G.; Campo, J.J.; Cistero, P.; Jimenez, A.; et al. Controlled human malaria infection by intramuscular and direct venous inoculation of cryopreserved Plasmodium falciparum sporozoites in malaria-naive volunteers: Effect of injection volume and dose on infectivity rates. Malar. J. 2015, 14, 306. [Google Scholar] [CrossRef] [Green Version]
- Flores-Garcia, Y.; Herrera, S.M.; Jhun, H.; Perez-Ramos, D.W.; King, C.R.; Locke, E.; Raghunandan, R.; Zavala, F. Optimization of an in vivo model to study immunity to Plasmodium falciparum pre-erythrocytic stages. Malar. J. 2019, 18, 426. [Google Scholar] [CrossRef] [Green Version]
- Prinz, H.; Sattler, J.M.; Roth, A.; Ripp, J.; Adams, J.H.; Frischknecht, F. Immunization efficacy of cryopreserved genetically attenuated Plasmodium berghei sporozoites. Parasitol. Res. 2018, 117, 2487–2497. [Google Scholar] [CrossRef]
- Patrapuvich, R.; Lerdpanyangam, K.; Jenwithisuk, R.; Rungin, S.; Boonhok, R.; Duangmanee, A.; Yimamnuaychok, N.; Sattabongkot, J. Viability and Infectivity of Cryopreserved Plasmodium Vivax Sporozoites. Southeast Asian J. Trop Med. Public Health 2016, 47, 171–181. [Google Scholar]
- Mutetwa, S.; James, E. Cryopreservation of Plasmodium chabaudi I: Protection by glycerol and dimethyl sulfoxide during cooling and by glucose following thawing. Cryobiology 1984, 21, 329–339. [Google Scholar] [CrossRef]
- Hollingdale, M.R.; Leland, P.; Sigler, C.I.; Leef, J.L. In vitro infectivity of cryopreserved Plasmodium berghei sporozoites to cultured cells. Trans. R. Soc. Trop. Med. Hyg. 1985, 79, 206–208. [Google Scholar] [CrossRef]
- Roestenberg, M.; Bijker, E.M.; Sim, B.K.L.; Billingsley, P.F.; James, E.R.; Bastiaens, G.J.H.; Teirlinck, A.C.; Scholzen, A.; Teelen, K.; Arens, T.; et al. Controlled human malaria infections by intradermal injection of cryopreserved Plasmodium falciparum sporozoites. Am. J. Trop. Med. Hyg. 2013, 88, 5–13. [Google Scholar] [CrossRef]
- Shekalaghe, S.; Rutaihwa, M.; Billingsley, P.F.; Chemba, M.; Daubenberger, C.A.; James, E.R.; Mpina, M.; Ali Juma, O.; Schindler, T.; Huber, E.; et al. Controlled human malaria infection of Tanzanians by intradermal injection of aseptic, purified, cryopreserved Plasmodium falciparum sporozoites. Am. J. Trop. Med. Hyg. 2014, 91, 471–480. [Google Scholar] [CrossRef]
- Udomsangpetch, R.; Kaneko, O.; Chotivanich, K.; Sattabongkot, J. Cultivation of Plasmodium vivax. Trends Parasitol. 2008, 24, 85–88. [Google Scholar] [CrossRef]
- Gunalan, K.; Rowley, E.H.; Miller, L.H. A Way forward for Culturing Plasmodium vivax. Trends Parasitol. 2020, 36, 512–519. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Fraction A (pH 7 with NaOH or HCl) | Fraction B (pH 7 with NaOH or HCl) | Purpose |
|---|---|---|---|
| Citric Acid | 1.34 g | 1.34 g | Preservative, antioxidant, metabolite |
| Glucose | 1 g | 1 g | Sugar, secondary LMW cryoprotectant |
| Glycerol | none | 14 mL | Primary LMW cryoprotectant |
| Sporozoite media * | 20 mL | 20 mL | Antibiotics, eukaryotic cell protective proteins |
| Sterile Water | To make 100 mL | To make 100 mL | Solvent |
| Strain | Integrity | Viability | Infectiousness |
|---|---|---|---|
| P. falciparum | 95 + 1.22 | 84.3 ± 8.0 | 100 ± 0.0 |
| P. vivax | 66.35 ± 0.12 | 44.17 ± 7.6 | 100 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowers, C.; Hancox, L.; Peissig, K.; Shiau, J.C.; Vantaux, A.; Witkowski, B.; Phal, S.; Maher, S.P.; Harty, J.T.; Kyle, D.E.; et al. Cryopreservation of Plasmodium Sporozoites. Pathogens 2022, 11, 1487. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121487
Bowers C, Hancox L, Peissig K, Shiau JC, Vantaux A, Witkowski B, Phal S, Maher SP, Harty JT, Kyle DE, et al. Cryopreservation of Plasmodium Sporozoites. Pathogens. 2022; 11(12):1487. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121487
Chicago/Turabian StyleBowers, Carson, Lisa Hancox, Kristen Peissig, Justine C. Shiau, Amélie Vantaux, Benoit Witkowski, Sivchheng Phal, Steven P. Maher, John T. Harty, Dennis E. Kyle, and et al. 2022. "Cryopreservation of Plasmodium Sporozoites" Pathogens 11, no. 12: 1487. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121487