


Impact of Infection with Flavobacterium psychrophilum and Antimicrobial Treatment on the Intestinal Microbiota of Rainbow Trout

Abstract
:1. Introduction
2. Material and Methods
2.1. Fish and Rearing Conditions
2.2. Preparation of Medicated Feed
2.3. Preparation of Bacterial Inoculum
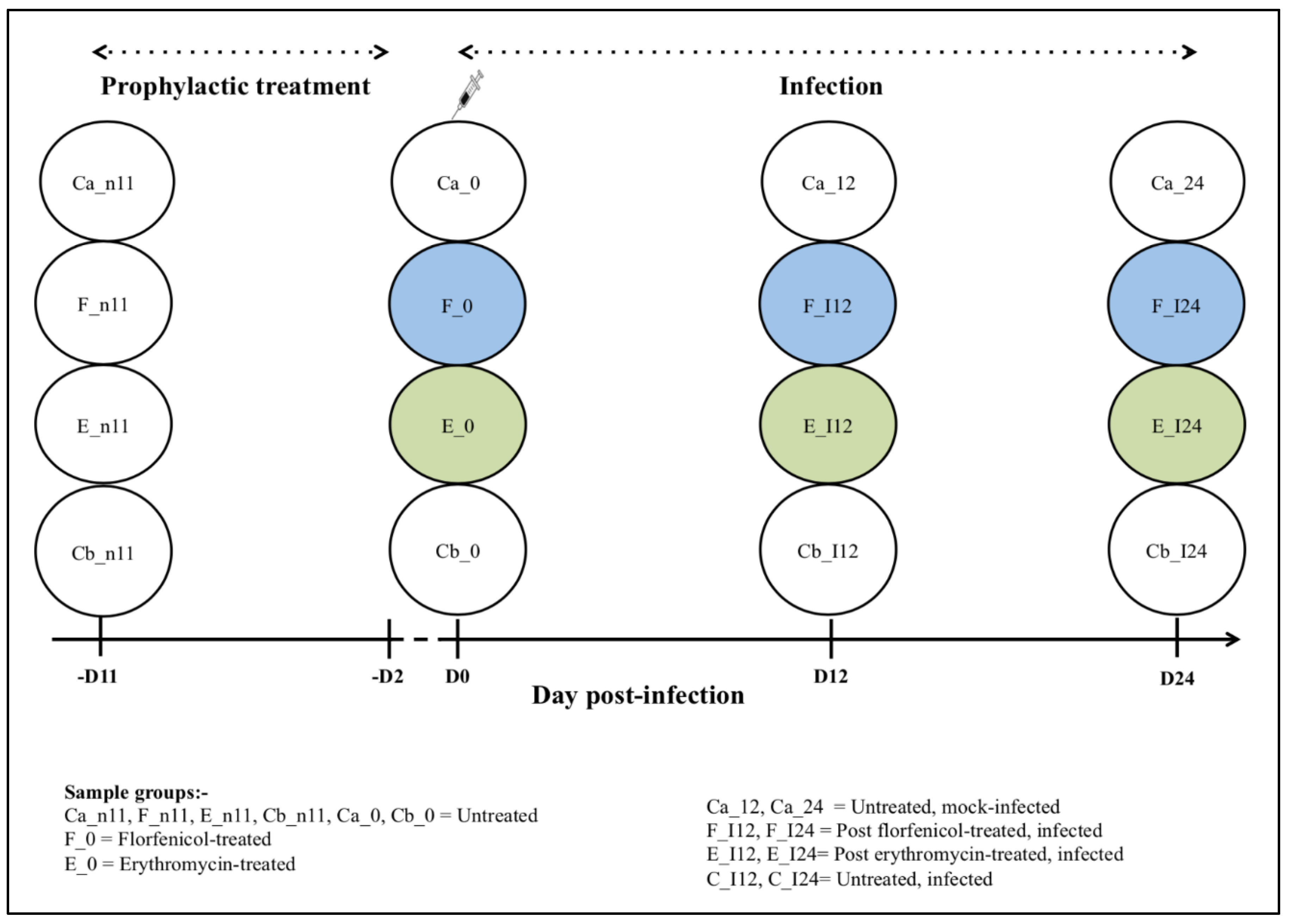
2.4. Experimental Design
2.5. Prophylactic Oral Antibiotic Treatment
2.6. Flavobacterium Psychrophilum Infection
2.7. Sample Collection
2.7.1. Intestinal Contents
2.7.2. Spleen
2.8. Microbiota Analysis
2.8.1. Intestinal Contents DNA Extraction
2.8.2. 16S rRNA Gene PCR Amplification and Purification
2.8.3. Index PCR, Purification, and Sequencing
2.9. Bioinformatic Analysis
Statistical Analysis of Microbiota Data
2.10. Enumeration of Splenic Bacterial Load
Splenic DNA Extraction and qPCR
2.11. Statistical Analysis of Survival and Spleen Data
3. Results
3.1. Diversity of Rainbow Trout Intestinal Microbiota
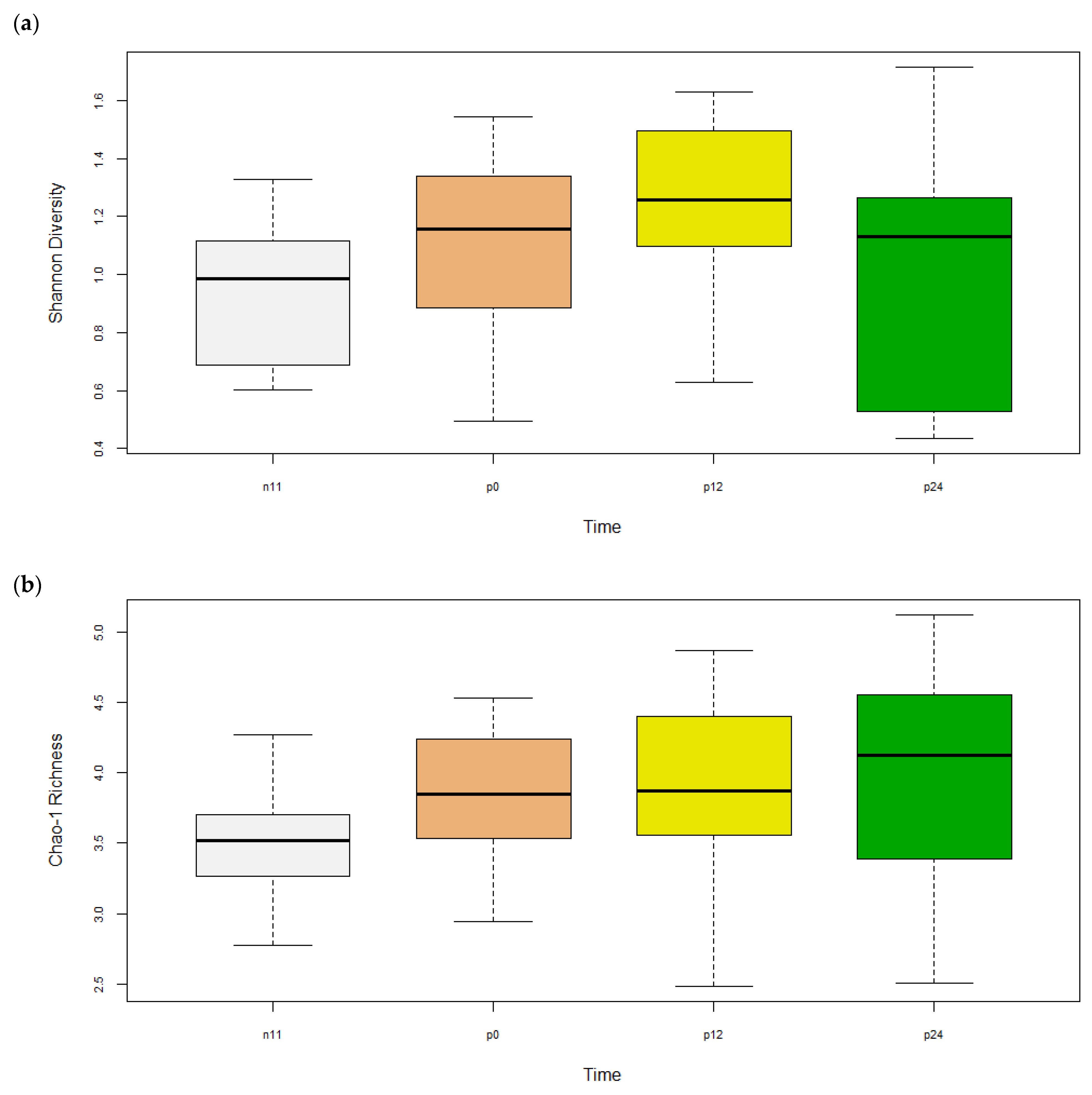
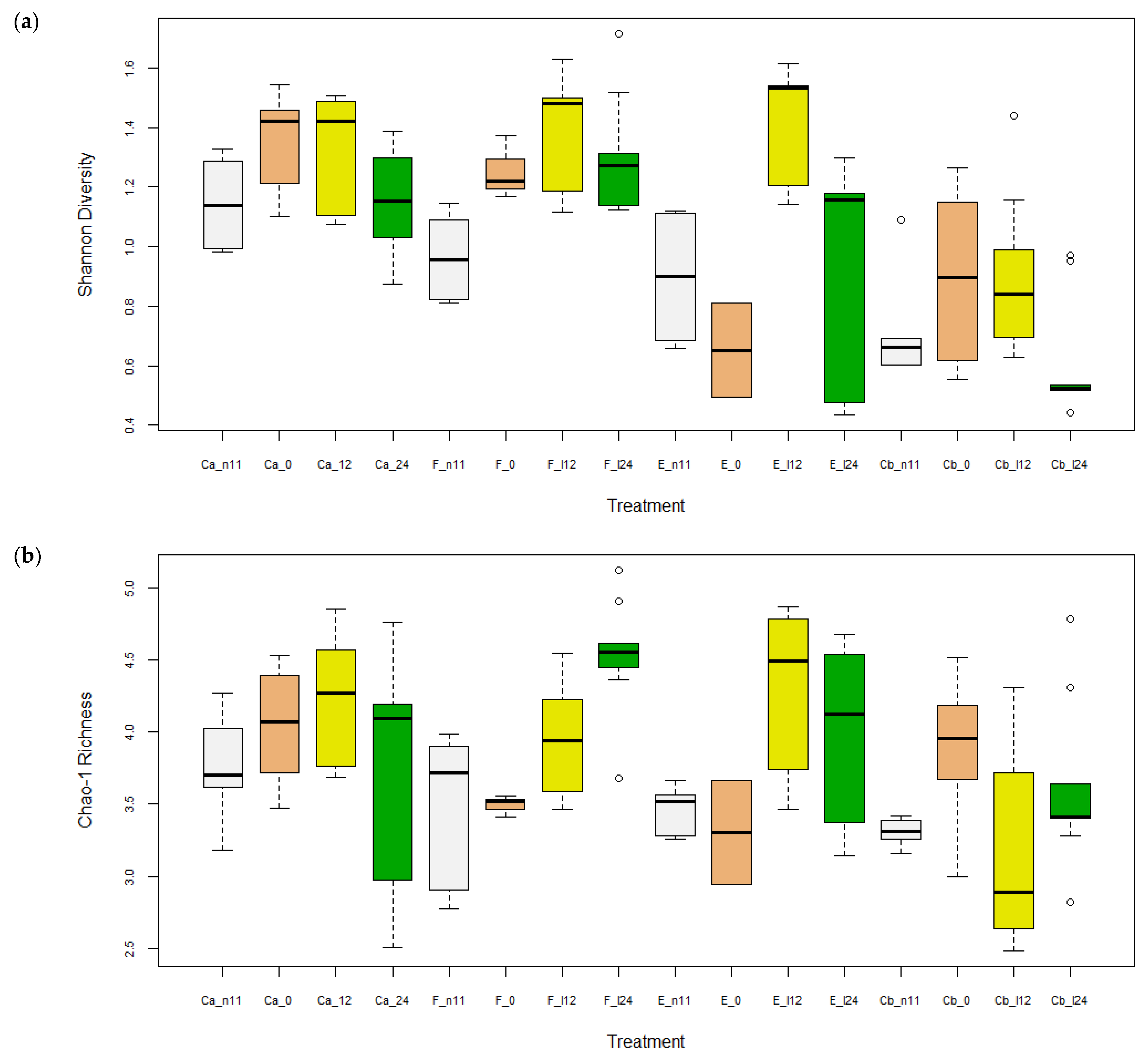
3.1.1. Alpha Diversity
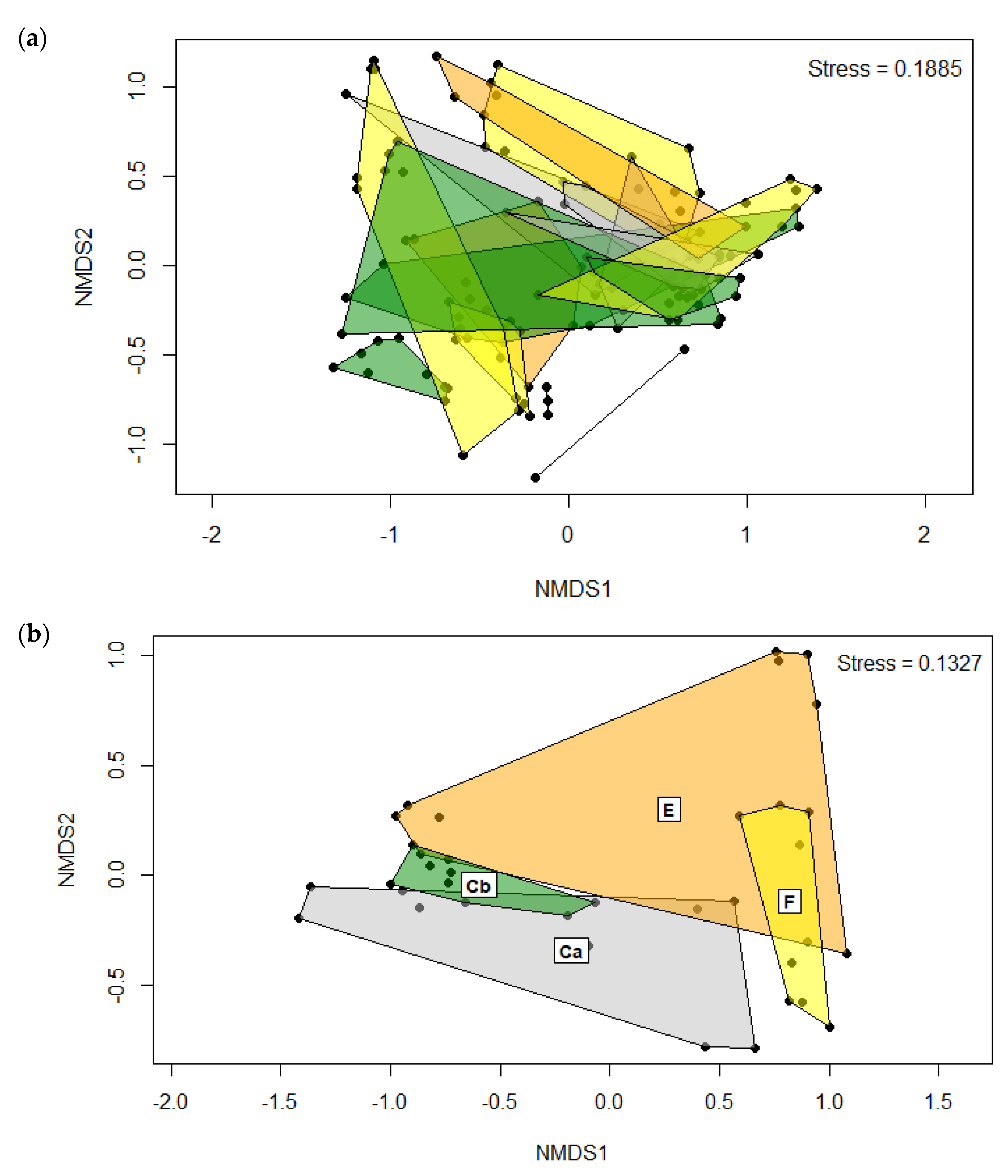
3.1.2. Beta Diversity
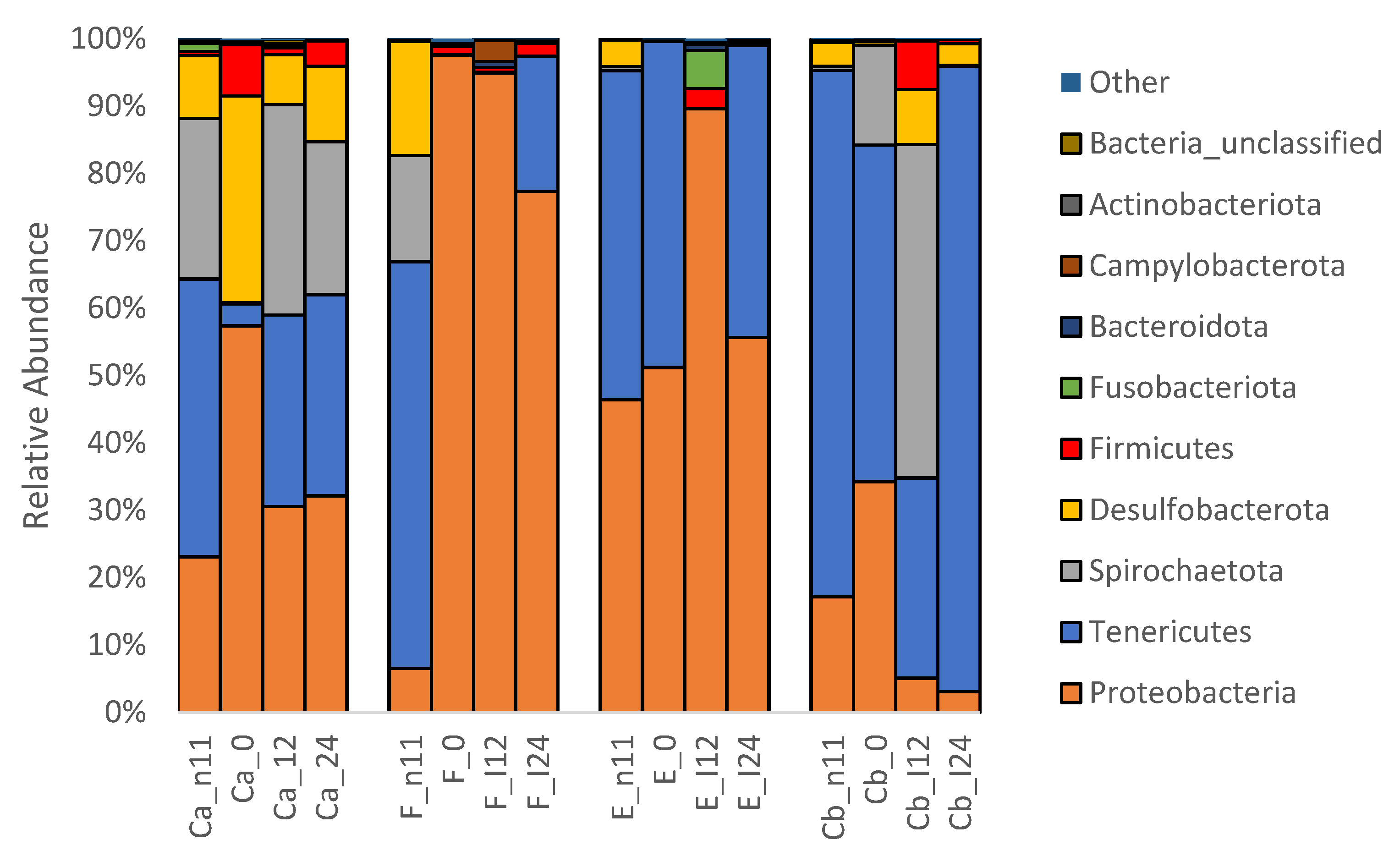
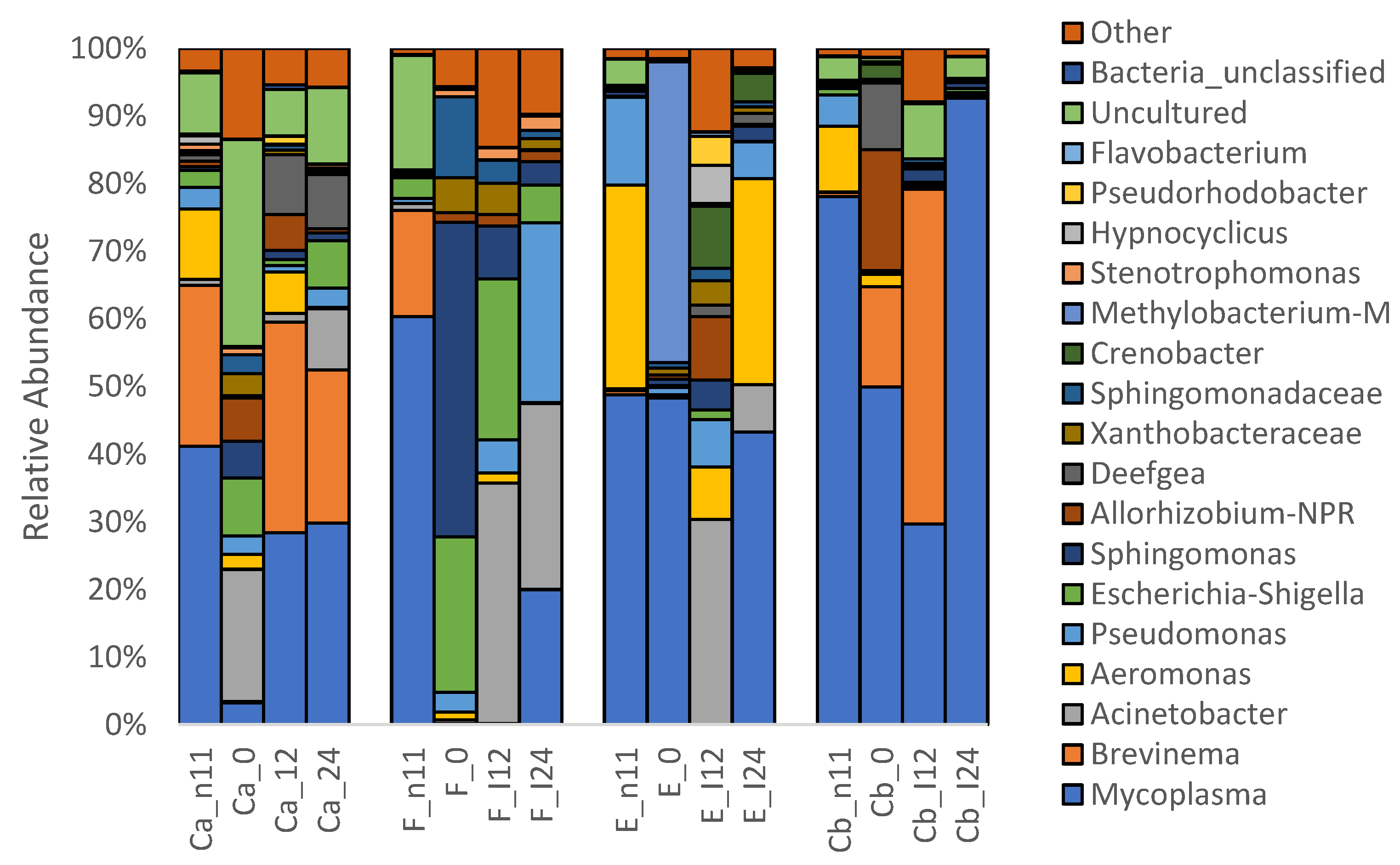
3.1.3. Microbiota before and after Oral Antibiotic Treatment (-D11 and D0 p.i.)
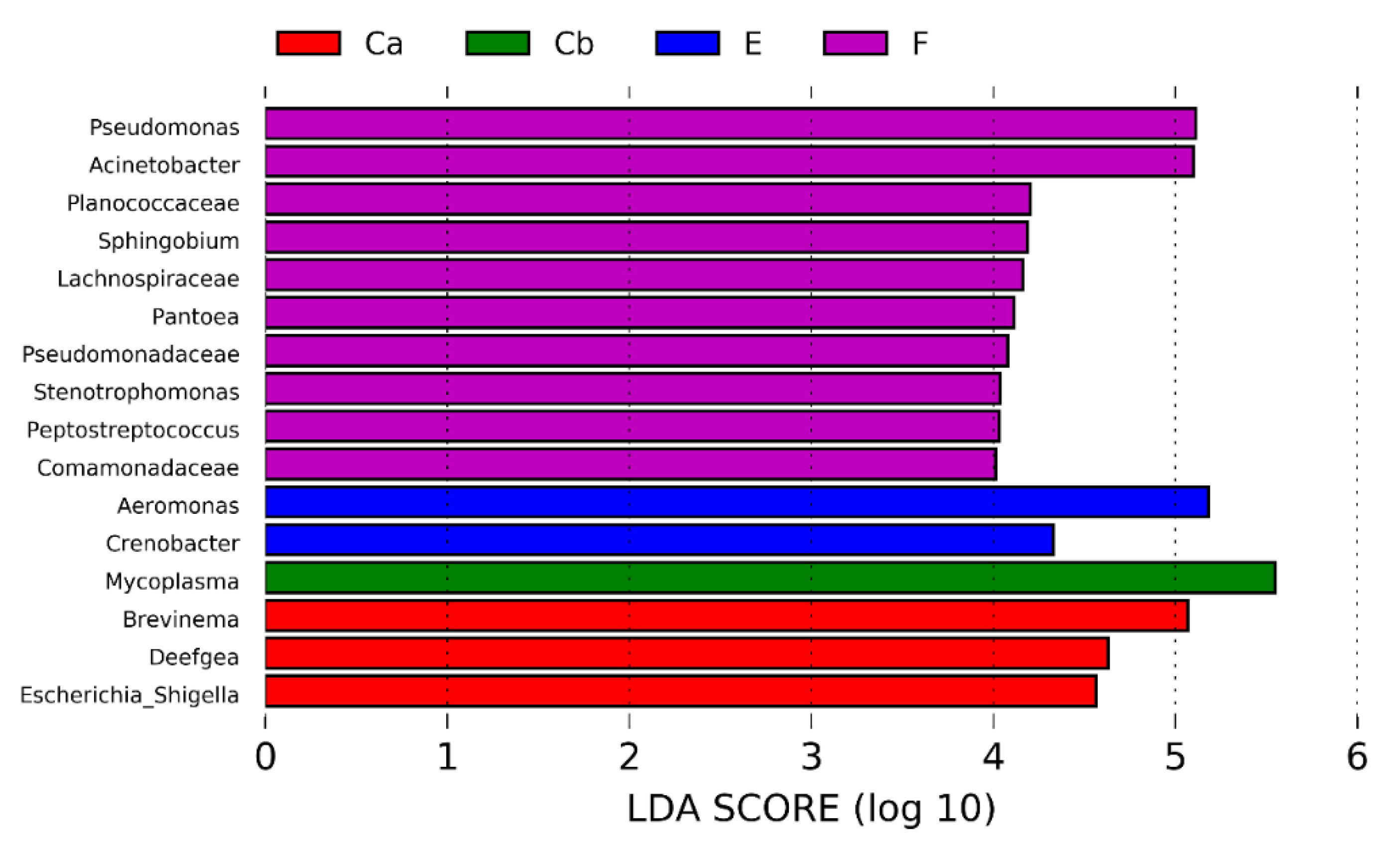
3.1.4. Microbiota after F. psychrophilum Infection (D12 & D24 p.i.)
3.2. Flavobacterium Psychrophilum Infection
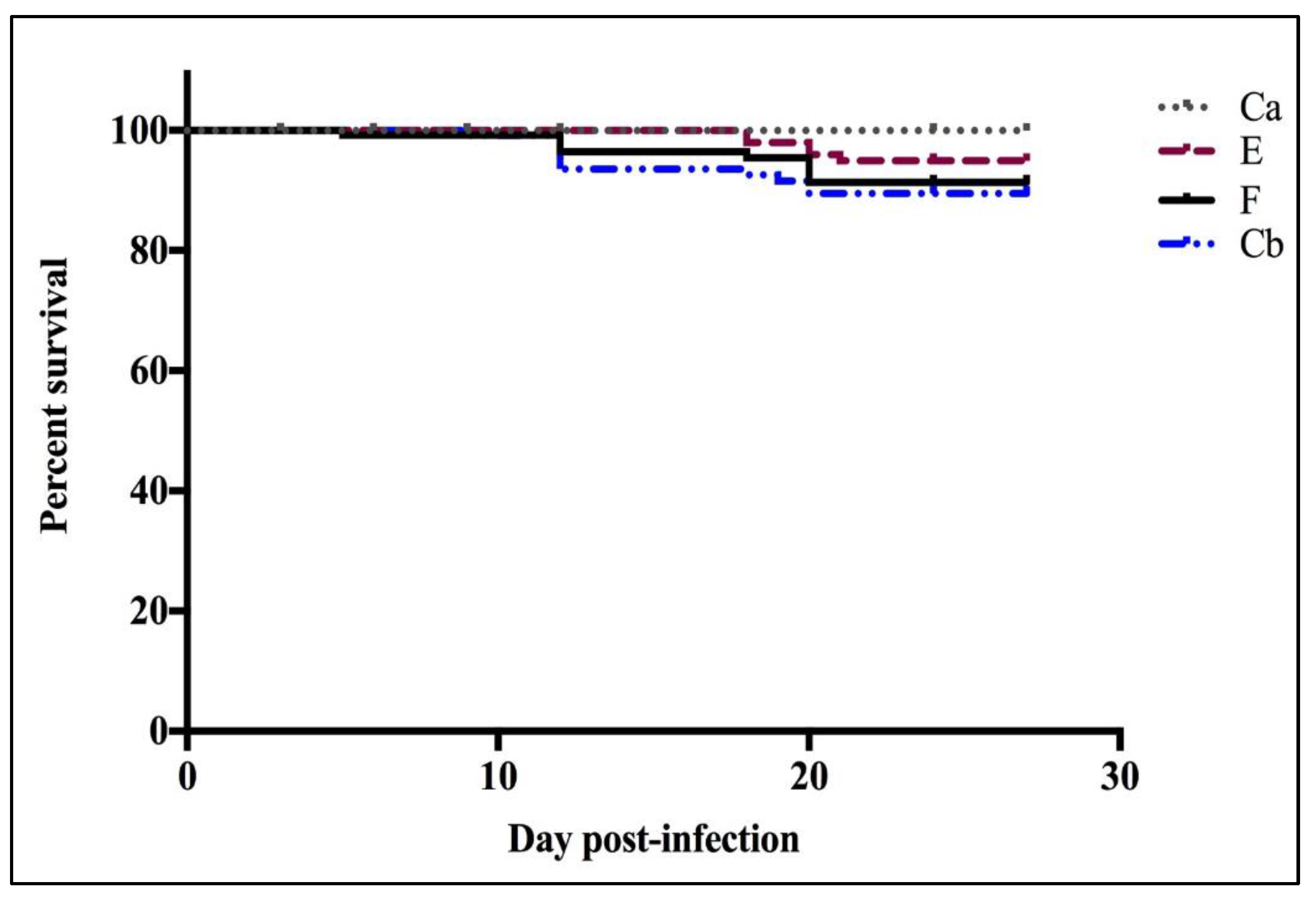
3.2.1. Survival Curves
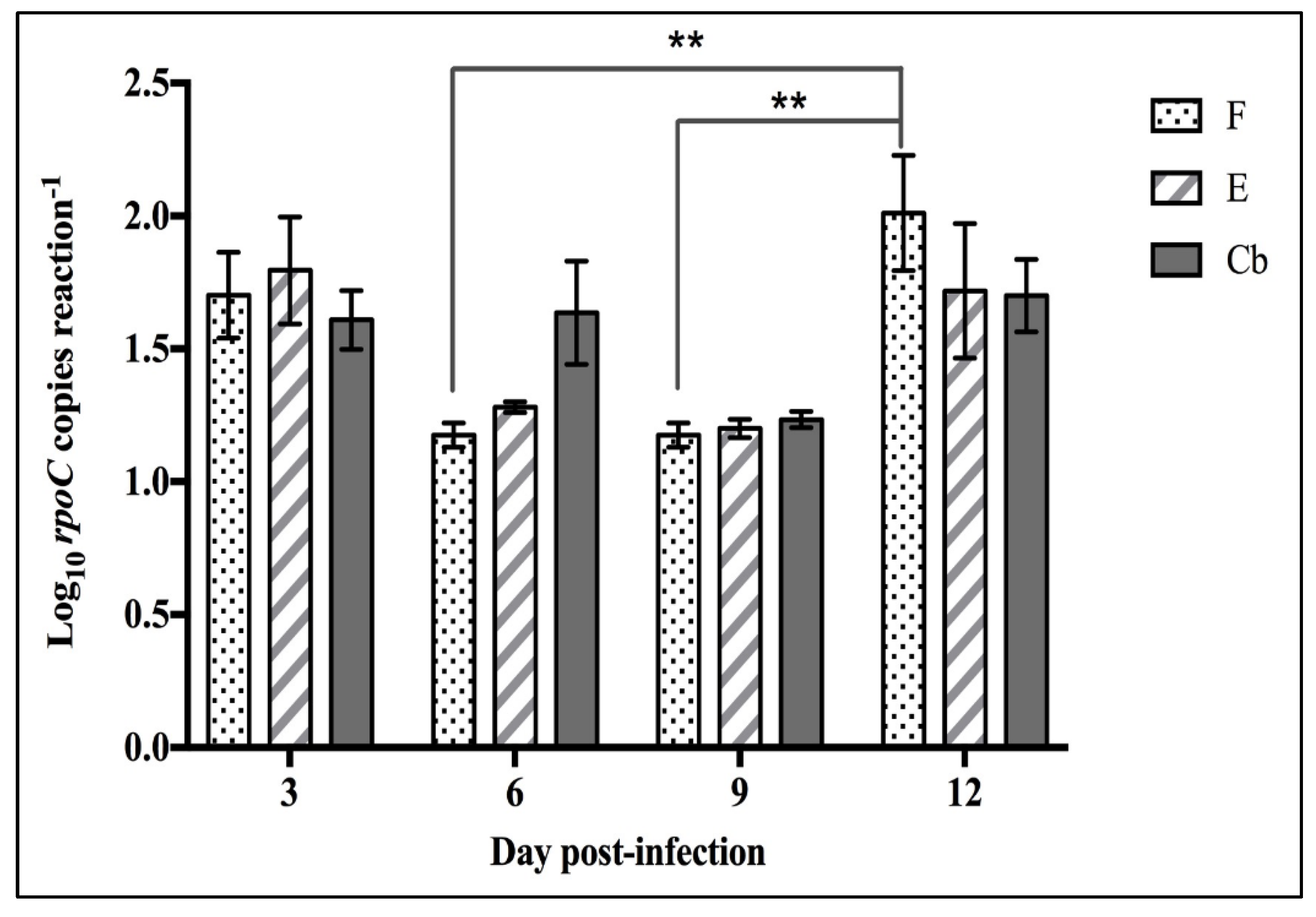
3.2.2. Splenic Bacterial Load
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cahill, M.M. Bacterial flora of fishes: A review. Microb. Ecol. 1990, 19, 21–41. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Strøm, E.; Tabachek, J.A. Intestinal microflora of salmonids: A review. Aquacult. Res. 1995, 26, 773–789. [Google Scholar] [CrossRef]
- Zhou, Z.; Yao, B.; Romero, J.; Waines, P.; Ringø, E.; Emery, M.; Liles, M.R.; Merrifield, D.L. Methodological approaches used to assess fish gastrointestinal communities. In Aquaculture Nutrition: Gut Health, Probiotics and Prebiotics, Merrifield, D.L., Ringø, E., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2014; pp. 101–127. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2020—Sustainability in Action; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2020; pp. 1–244. [Google Scholar]
- Huyben, D.; Chiasson, M.; Lumsden, J.S.; Pham, P.H.; Chowdhury, M.A.K. Dietary microencapsulated blend of organic acids and plant essential oils affects intestinal morphology and microbiome of rainbow trout (Oncorhynchus mykiss). Microorganisms 2021, 9, 2063. [Google Scholar] [CrossRef] [PubMed]
- Huyben, D.; Nyman, A.; Vidaković, A.; Passoth, V.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Effects of dietary inclusion of the yeasts Saccharomyces cerevisiae and Wickerhamomyces anomalus on gut microbiota of rainbow trout. Aquaculture 2017, 473, 528–537. [Google Scholar] [CrossRef]
- Huyben, D.; Sun, L.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Dietary live yeast and increased water temperature influence the gut microbiota of rainbow trout. J. Appl. Microbiol. 2018, 124, 1377–1392. [Google Scholar] [CrossRef]
- Lyons, P.P.; Turnbull, J.F.; Dawson, K.A.; Crumlish, M. Exploring the microbial diversity of the distal intestinal lumen and mucosa of farmed rainbow trout Oncorhynchus mykiss (Walbaum) using next generation sequencing (NGS). Aquacult. Res. 2017, 48, 77–91. [Google Scholar] [CrossRef]
- Ingerslev, H.C.; von Gersdorff Jørgensen, L.; Lenz Strube, M.; Larsen, N.; Dalsgaard, I.; Boye, M.; Madsen, L. The development of the gut microbiota in rainbow trout (Oncorhynchus mykiss) is affected by first feeding and diet type. Aquaculture 2014, 424–425, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Cipriano, R.C.; Holt, R.A. Flavobacterium psychrophilum, Cause of Bacterial Cold-Water Disease and Rainbow Trout Fry Syndrome; US Department of the Interior, US Geological Survey, National Fish Health: Kearneysville, WV, USA, 2005. [Google Scholar]
- Holt, R.A. Cytophaga psychrophila, the Causative Agent of Bacterial Cold-Water Disease in Salmonid Fish; Oregon State University: Corvallis, OR, USA, 1987. [Google Scholar]
- Francino, M. Antibiotics and the human gut microbiome: Dysbioses and accumulation of resistances. Front. Microbiol. 2016, 6, 1543. [Google Scholar] [CrossRef] [Green Version]
- Parshukov, A.N.; Fokina, N.N.; Sukhovskaya, I.V.; Kantserova, N.P.; Lysenko, L.A. Infection and antibiotic treatment have prolonged effects on gut microbiota, muscle and hepatic fatty acids in rainbow trout (Oncorhynchus mykiss). J. Appl. Microbiol. 2022, 133, 1709–1724. [Google Scholar] [CrossRef]
- Payne, C.J.; Turnbull, J.F.; MacKenzie, S.; Crumlish, M. The effect of oxytetracycline treatment on the gut microbiome community dynamics in rainbow trout (Oncorhynchus mykiss) over time. Aquaculture 2022, 560, 738559. [Google Scholar] [CrossRef]
- Roy Choudhury, A.; Park, J.-Y.; Kim, D.Y.; Choi, J.; Acharya, S.; Park, J.-H. Exposure to Oxy-Tetracycline Changes Gut Bacterial Community Composition in Rainbow Trout: A Preliminary Study. Animals 2021, 11, 3404. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pascual, D.; Pérez-Cobas, A.E.; Rigaudeau, D.; Rochat, T.; Bernardet, J.-F.; Skiba-Cassy, S.; Marchand, Y.; Duchaud, E.; Ghigo, J.-M. Sustainable plant-based diets promote rainbow trout gut microbiota richness and do not alter resistance to bacterial infection. Anim. Microbiome 2021, 3, 47. [Google Scholar] [CrossRef] [PubMed]
- Donati, V.L.; Madsen, L.; Middelboe, M.; Strube, M.L.; Dalsgaard, I. The Gut Microbiota of Healthy and Flavobacterium psychrophilum-Infected Rainbow Trout Fry Is Shaped by Antibiotics and Phage Therapies. Front. Microbiol. 2022, 13, 771296. [Google Scholar] [CrossRef] [PubMed]
- Jarau, M.; MacInnes, J.I.; Lumsden, J.S. Erythromycin and florfenicol treatment of rainbow trout Oncorhynchus mykiss (Walbaum) experimentally infected with Flavobacterium psychrophilum. J. Fish Dis. 2019, 42, 325–334. [Google Scholar] [CrossRef]
- Orieux, N.; Bourdineaud, J.P.; Douet, D.G.; Daniel, P.; Le Henaff, M. Quantification of Flavobacterium psychrophilum in rainbow trout, Oncorhynchus mykiss (Walbaum), tissues by qPCR. J. Fish Dis. 2011, 34, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Jarau, M.; Di Natale, A.; Huber, P.E.; MacInnes, J.I.; Lumsden, J.S. Virulence of Flavobacterium psychrophilum isolates in rainbow trout Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2018, 41, 1505–1514. [Google Scholar] [CrossRef]
- Maturin, L. Aerobic Plate Count. In Bacteriological Analytical Manual Online. 2001. Available online: https://cir.nii.ac.jp/all?q=http://www.cfsan.fda.gov/%E3%80%9Cebam/bam-3.html (accessed on 12 October 2022).
- Strepparava, N.; Wahli, T.; Segner, H.; Petrini, O. Detection and quantification of Flavobacterium psychrophilum in water and fish tissue samples by quantitative real time PCR. BMC Microbiol. 2014, 14, 105. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, 1–11. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R-Core-Team. R: A Language and Environment for Statistical Computing, 3.2.2; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Vegan: Community ecology package. R package Version 2.5-1. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 12 October 2022).
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Lyons, P.P.; Turnbull, J.F.; Dawson, K.A.; Crumlish, M. Phylogenetic and functional characterization of the distal intestinal microbiome of rainbow trout Oncorhynchus mykiss from both farm and aquarium settings. J. Appl. Microbiol. 2017, 122, 347–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huyben, D.; Vidaković, A.; Werner Hallgren, S.; Langeland, M. High-throughput sequencing of gut microbiota in rainbow trout (Oncorhynchus mykiss) fed larval and pre-pupae stages of black soldier fly (Hermetia illucens). Aquaculture 2019, 500, 485–491. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Ascione, C.; Gini, E.; Ceccotti, C.; Gasco, L. Rainbow trout (Oncorhynchus mykiss) gut microbiota is modulated by insect meal from Hermetia illucens prepupae in the diet. Rev. Fish Biol. Fish. 2019, 29, 465–486. [Google Scholar] [CrossRef]
- Huyben, D.; Roehe, B.K.; Bekaert, M.; Ruyter, B.; Glencross, B. Dietary lipid:protein ratio and n-3 long-chain polyunsaturated fatty acids alters the gut microbiome of Atlantic salmon under hypoxic and normoxic conditions. Front. Microbiol. 2020, 11, 589898. [Google Scholar] [CrossRef]
- Zarkasi, K.Z.; Abell, G.C.; Taylor, R.S.; Neuman, C.; Hatje, E.; Tamplin, M.L.; Katouli, M.; Bowman, J.P. Pyrosequencing based characterization of gastrointestinal bacteria of Atlantic salmon (Salmo salar L.) within a commercial mariculture system. J. Appl. Microbiol. 2014, 117, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Llewellyn, M.S.; McGinnity, P.; Dionne, M.; Letourneau, J.; Thonier, F.; Carvalho, G.R.; Creer, S.; Derome, N. The biogeography of the Atlantic salmon (Salmo salar) gut microbiome. ISME J 2016, 10, 1280–1284. [Google Scholar] [CrossRef] [Green Version]
- Rudi, K.; Angell, I.L.; Pope, P.B.; Vik, J.O.; Sandve, S.R.; Snipen, L.-G. Stable core gut microbiota across the freshwater-to-saltwater transition for farmed Atlantic salmon. Appl. Environ. Microbiol. 2018, 84, e01974-17. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.M.; Wiens, G.D.; Salinas, I. Analysis of the gut and gill microbiome of resistant and susceptible lines of rainbow trout (Oncorhynchus mykiss). Fish Shellfish. Immunol. 2019, 86, 497–506. [Google Scholar] [CrossRef]
- Gupta, R.S.; Sawnani, S.; Adeolu, M.; Alnajar, S.; Oren, A. Phylogenetic framework for the phylum Tenericutes based on genome sequence data: Proposal for the creation of a new order Mycoplasmoidales ord. nov., containing two new families Mycoplasmoidaceae fam. nov. and Metamycoplasmataceae fam. nov. harbouring Eperythrozoon, Ureaplasma and five novel genera. Antonie Van Leeuwenhoek 2018, 111, 1583–1630. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.R.; Watral, V.; Sichel, S.; Spagnoli, S.; Banse, A.V.; Mittge, E.; Sharpton, T.J.; Guillemin, K.; Kent, M.L. Transmission of a common intestinal neoplasm in zebrafish by cohabitation. J. Fish Dis. 2018, 41, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Bozzi, D.; Rasmussen, J.A.; Carøe, C.; Sveier, H.; Nordøy, K.; Gilbert, M.T.P.; Limborg, M.T. Salmon gut microbiota correlates with disease infection status: Potential for monitoring health in farmed animals. Anim. Microbiome 2021, 3, 30. [Google Scholar] [CrossRef]
- Rasmussen, J.A.; Villumsen, K.R.; Duchêne, D.A.; Puetz, L.C.; Delmont, T.O.; Sveier, H.; Jørgensen, L.v.G.; Præbel, K.; Martin, M.D.; Bojesen, A.M.; et al. Genome-resolved metagenomics suggests a mutualistic relationship between Mycoplasma and salmonid hosts. Commun. Biol. 2021, 4, 579. [Google Scholar] [CrossRef]
- Patil, H.J.; Benet-Perelberg, A.; Naor, A.; Smirnov, M.; Ofek, T.; Nasser, A.; Minz, D.; Cytryn, E. Evidence of increased antibiotic resistance in phylogenetically-diverse Aeromonas isolates from semi-intensive fish ponds treated with antibiotics. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tao, W.; Lee, M.H.; Wu, J.; Kim, N.H.; Kim, J.-C.; Chung, E.; Hwang, E.C.; Lee, S.-W. Inactivation of chloramphenicol and florfenicol by a novel chloramphenicol hydrolase. Appl. Environ. Microbiol. 2012, 78, 6295–6301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.-h.; Aoki, T. Sequence analysis of the florfenicol resistance gene encoded in the transferable R-plasmid of a fish pathogen, Pasteurella piscicida. Microbiol. Immunol. 1996, 40, 665–669. [Google Scholar] [CrossRef]
- Sanders, J.W.; Martin, J.W.; Hooke, M.; Hooke, J. Methylobacterium mesophilicum infection: Case report and literature review of an unusual opportunistic pathogen. Clin. Infect. Dis. 2000, 30, 936–938. [Google Scholar] [CrossRef] [Green Version]
- Kovaleva, J.; Degener, J.E.; van der Mei, H.C. Methylobacterium and its role in health care-associated infection. J. Clin. Microbiol. 2014, 52, 1317–1321. [Google Scholar] [CrossRef] [Green Version]
- Boutin, S.; Sauvage, C.; Bernatchez, L.; Audet, C.; Derome, N. Inter individual variations of the fish skin microbiota: Host genetics basis of mutualism? PLoS ONE 2014, 9, e102649. [Google Scholar] [CrossRef]
- Carda-Dieguez, M.; Mira, A.; Fouz, B. Pyrosequencing survey of intestinal microbiota diversity in cultured sea bass (Dicentrarchus labrax) fed functional diets. FEMS Microbiol. Ecol. 2014, 87, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Giatsis, C.; Sipkema, D.; Smidt, H.; Heilig, H.; Benvenuti, G.; Verreth, J.; Verdegem, M. The impact of rearing environment on the development of gut microbiota in tilapia larvae. Sci. Rep. 2015, 5, 18206. [Google Scholar] [CrossRef] [Green Version]
- Lyons, P.P.; Turnbull, J.F.; Dawson, K.A.; Crumlish, M. Effects of low-level dietary microalgae supplementation on the distal intestinal microbiome of farmed rainbow trout Oncorhynchus mykiss. Aquacult. Res. 2017, 48, 2438–2452. [Google Scholar] [CrossRef]
- Stephens, W.Z.; Burns, A.R.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J. The composition of the zebrafish intestinal microbial community varies across development. ISME J. 2016, 10, 644–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, L.; Amberg, J.; Chapman, D.; Gaikowski, M.; Liu, W.-T. Fish gut microbiota analysis differentiates physiology and behavior of invasive Asian carp and indigenous American fish. ISME J. 2014, 8, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narrowe, A.B.; Albuthi-Lantz, M.; Smith, E.P.; Bower, K.J.; Roane, T.M.; Vajda, A.M.; Miller, C.S. Perturbation and restoration of the fathead minnow gut microbiome after low-level triclosan exposure. Microbiome 2015, 3, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarkasi, K.Z.; Taylor, R.S.; Abell, G.C.J.; Tamplin, M.L.; Glencross, B.D.; Bowman, J.P. Atlantic salmon (Salmo salar L.) gastrointestinal microbial community dynamics in relation to digesta properties and diet. Microb. Ecol. 2016, 71, 589–603. [Google Scholar] [CrossRef]
- Carlson, J.M.; Hyde, E.R.; Petrosino, J.F.; Manage, A.B.; Primm, T.P. The host effects of Gambusia affinis with an antibiotic-disrupted microbiome. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 178, 163–168. [Google Scholar] [CrossRef]
- Jernberg, C.; Löfmark, S.; Edlund, C.; Jansson, J.K. Long-term ecological impacts of antibiotic administration on the human intestinal microbiota. ISME J. 2007, 1, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Buffie, C.G.; Jarchum, I.; Equinda, M.; Lipuma, L.; Gobourne, A.; Viale, A.; Ubeda, C.; Xavier, J.; Pamer, E.G. Profound alterations of intestinal microbiota following a single dose of clindamycin results in sustained susceptibility to Clostridium difficile-induced colitis. Infect. Immun. 2012, 80, 62–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarrete, P.; Mardones, P.; Opazo, R.; Espejo, R.; Romero, J. Oxytetracycline treatment reduces bacterial diversity of intestinal microbiota of Atlantic salmon. J. Aquat. Anim. Health 2008, 20, 177–183. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fish Group | Group Details | Intestinal Contents Sample Group | Day of Sampling (p.i.) | Sample Size |
|---|---|---|---|---|
| Ca | Control | Ca_n11 a | −11 | 6 |
| Ca_0 a | 0 | 9 | ||
| Ca_12 b | 12 | 9 | ||
| Ca_24 b | 24 | 9 | ||
| F | FFN-treated + FPG101-infected | F_n11 a | −11 | 6 |
| F_0 d | 0 | 3 | ||
| F_I12 | 12 | 9 | ||
| F_I24 | 24 | 9 | ||
| E | ERY-treated + FPG101-infected | E_n11 a | −11 | 6 |
| E_0 e | 0 | 2 | ||
| E_I12 | 12 | 9 | ||
| E_I24 | 24 | 9 | ||
| Cb | Untreated + FPG101-infected | Cb_n11 a | −11 | 6 |
| Cb_0 c | 0 | 9 | ||
| Cb_I12 | 12 | 9 | ||
| Cb_I24 | 24 | 9 |
| Treatment | Control + Uninfected (Ca) | Erythromycin + Infected (E) | Florfenicol + Infected (F) | Untreated + Infected (Cb) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time (days) | −11 | 0 | 12 | 24 | −11 | 0 | 12 | 24 | −11 | 0 | 12 | 24 | −11 | 0 | 12 | 24 | SEM |
| Coverage | 99.7 | 99.6 | 99.5 | 99.6 | 99.8 | 99.8 | 99.4 | 99.5 | 99.8 | 99.9 | 99.7 | 99.3 | 99.8 | 99.6 | 99.8 | 99.7 | 0.02 |
| No. OTUs | 31.5 | 50.6 | 48.3 | 37.0 | 24.5 | 25.0 | 58.6 | 43.3 | 23.5 | 32.7 | 44.8 | 64.4 | 22.3 | 30.1 | 22.6 | 26.4 | 1.9 |
| Shannon | 1.3 | 1.9 | 1.7 | 1.4 | 0.8 | 0.4 | 2.0 | 0.9 | 0.9 | 1.6 | 1.9 | 1.7 | 0.5 | 0.9 | 0.9 | 0.4 | 0.1 |
| Chao1 | 44.9 | 62.5 | 72.3 | 51.9 | 32.4 | 29.0 | 81.2 | 61.8 | 36.7 | 33.0 | 55.2 | 99.0 | 27.4 | 52.3 | 28.5 | 44.6 | 3.0 |
| Ace | 46.4 | 67.5 | 84.0 | 73.0 | 38.7 | 28.5 | 97.6 | 75.5 | 46.1 | 33.6 | 64.0 | 128.1 | 28.8 | 74.0 | 35.6 | 61.9 | 4.1 |
| Comparison | Coverage | No. OTUs | Shannon | Chao1 | Ace | ||
|---|---|---|---|---|---|---|---|
| Diet: | |||||||
| Ca | Cb | 0.188 | <0.001 | <0.001 | 0.004 | 0.048 | |
| Ca | E | 0.904 | 0.997 | 0.004 | 0.997 | 0.997 | |
| Ca | F | 0.994 | 0.947 | 0.999 | 0.976 | 0.986 | |
| Cb | E | 0.055 | <0.001 | <0.001 | 0.015 | 0.112 | |
| Cb | F | 0.356 | <0.001 | <0.001 | 0.002 | 0.028 | |
| E | F | 0.805 | 0.890 | 0.009 | 0.934 | 0.956 | |
| Time: | |||||||
| 0 | n11 | 0.379 | 0.003 | 0.016 | 0.039 | 0.038 | |
| 0 | 12 | 0.597 | 0.999 | 0.201 | 0.999 | 0.999 | |
| 0 | 24 | 0.065 | 0.973 | 0.087 | 0.992 | 0.654 | |
| 12 | 24 | 0.487 | 0.974 | <0.001 | 0.973 | 0.663 | |
| 12 | n11 | 0.015 | 0.001 | <0.001 | 0.023 | 0.011 | |
| 24 | n11 | <0.001 | 0.004 | 0.778 | 0.007 | <0.001 | |
| Diet × Time: | |||||||
| Ca | 0 | n11 | 1.000 | 0.673 | 0.927 | 0.998 | 0.996 |
| 0 | 12 | 0.990 | 1.000 | 1.000 | 1.000 | 1.000 | |
| 0 | 24 | 1.000 | 0.543 | 0.928 | 0.940 | 1.000 | |
| 12 | 24 | 0.999 | 0.753 | 0.996 | 0.656 | 0.995 | |
| 12 | n11 | 0.652 | 0.839 | 0.994 | 0.944 | 0.804 | |
| 24 | n11 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | |
| Cb | 0 | n11 | 0.818 | 0.998 | 0.987 | 0.795 | 0.302 |
| 0 | 12 | 0.655 | 0.729 | 1.000 | 0.222 | 0.309 | |
| 0 | 24 | 1.000 | 1.000 | 0.395 | 1.000 | 0.999 | |
| 12 | 24 | 0.982 | 0.988 | 0.339 | 0.837 | 0.956 | |
| 12 | n11 | 1.000 | 1.000 | 0.978 | 1.000 | 1.000 | |
| 24 | n11 | 0.995 | 1.000 | 1.000 | 0.998 | 0.920 | |
| E | 0 | n11 | 1.000 | 1.000 | 0.994 | 1.000 | 1.000 |
| 0 | 12 | 0.416 | 0.479 | 0.003 | 0.518 | 0.438 | |
| 0 | 24 | 0.606 | 0.991 | 0.997 | 0.949 | 0.746 | |
| 12 | 24 | 1.000 | 0.885 | <0.001 | 0.996 | 1.000 | |
| 12 | n11 | 0.258 | 0.032 | 0.003 | 0.179 | 0.350 | |
| 24 | n11 | 0.512 | 0.822 | 1.000 | 0.882 | 0.794 | |
| F | 0 | n11 | 0.799 | 0.997 | 0.896 | 1.000 | 1.000 |
| 0 | 12 | 0.210 | 1.000 | 1.000 | 0.994 | 0.981 | |
| 0 | 24 | <0.001 | 0.688 | 1.000 | 0.165 | 0.073 | |
| 12 | 24 | 0.115 | 0.940 | 1.000 | 0.525 | 0.368 | |
| 12 | n11 | 1.000 | 0.230 | 0.047 | 0.950 | 0.995 | |
| 24 | n11 | 0.017 | 0.002 | 0.270 | 0.018 | 0.029 | |
| Overall: | |||||||
| Diet | 0.063 | <0.001 | <0.001 | <0.001 | 0.017 | ||
| Time | <0.001 | <0.001 | <0.001 | 0.007 | <0.001 | ||
| Diet × Time | <0.001 | <0.001 | 0.002 | <0.001 | 0.002 | ||
| Group | Time (Days) | R-Value | p-Value | |
|---|---|---|---|---|
| ANOSIM: | ||||
| Ca | 0 | n11 | 0.578 | 0.001 |
| 0 | 12 | 0.692 | <0.001 | |
| 0 | 24 | 0.490 | <0.001 | |
| 12 | 24 | 0.012 | 0.345 | |
| 12 | n11 | 0.055 | 0.232 | |
| 24 | n11 | 0.065 | 0.722 | |
| Cb | 0 | n11 | 0.104 | 0.129 |
| 0 | 12 | 0.224 | 0.009 | |
| 0 | 24 | 0.276 | 0.001 | |
| 12 | 24 | 0.581 | <0.001 | |
| 12 | n11 | 0.340 | 0.012 | |
| 24 | n11 | 0.297 | 0.016 | |
| E | 0 | n11 | 0.604 | 0.070 |
| 0 | 12 | 0.530 | 0.020 | |
| 0 | 24 | 0.231 | 0.060 | |
| 12 | 24 | 0.212 | 0.026 | |
| 12 | n11 | 0.513 | 0.001 | |
| 24 | n11 | 0.170 | 0.075 | |
| F | 0 | n11 | 0.963 | 0.011 |
| 0 | 12 | 0.647 | 0.004 | |
| 0 | 24 | 0.776 | 0.001 | |
| 12 | 24 | 0.427 | 0.002 | |
| 12 | n11 | 0.987 | <0.001 | |
| 24 | n11 | 0.638 | <0.001 | |
| PERMANOVA: | ||||
| Diet | 0.176 | 0.001 | ||
| Time | 0.110 | 0.001 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huyben, D.; Jarau, M.; MacInnes, J.; Stevenson, R.; Lumsden, J. Impact of Infection with Flavobacterium psychrophilum and Antimicrobial Treatment on the Intestinal Microbiota of Rainbow Trout. Pathogens 2023, 12, 454. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12030454
Huyben D, Jarau M, MacInnes J, Stevenson R, Lumsden J. Impact of Infection with Flavobacterium psychrophilum and Antimicrobial Treatment on the Intestinal Microbiota of Rainbow Trout. Pathogens. 2023; 12(3):454. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12030454
Chicago/Turabian StyleHuyben, David, Maureen Jarau, Janet MacInnes, Roselynn Stevenson, and John Lumsden. 2023. "Impact of Infection with Flavobacterium psychrophilum and Antimicrobial Treatment on the Intestinal Microbiota of Rainbow Trout" Pathogens 12, no. 3: 454. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12030454