The Effect of Bacillus licheniformis MH48 on Control of Foliar Fungal Diseases and Growth Promotion of Camellia oleifera Seedlings in the Coastal Reclaimed Land of Korea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Antagonistic Bacteria Growth
2.2. Lytic Enzyme Assays
2.3. Antagonistic Activity of B. licheniformis MH48 against Foliar Fungal Pathogens
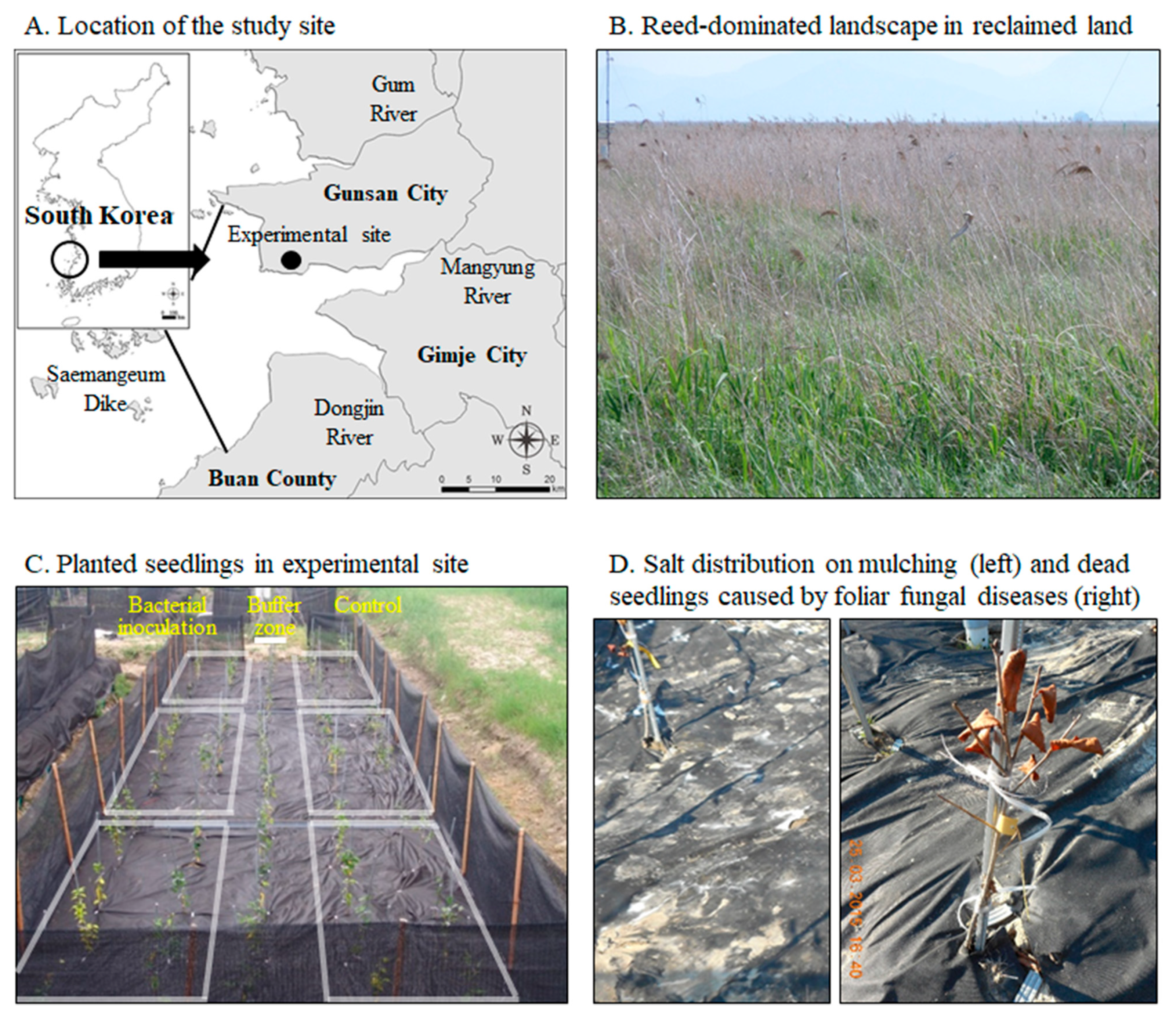
2.4. Study Area and Field Experimental Conditions
2.5. Chemical Properties in Soils and Nutrient Content in Seedlings
2.6. Analysis of C. oleifera Seedling Survival Rate
2.7. Statistical Analyses
3. Results
3.1. Effect of B. licheniformis MH48 on Foliar Fungal Pathogens
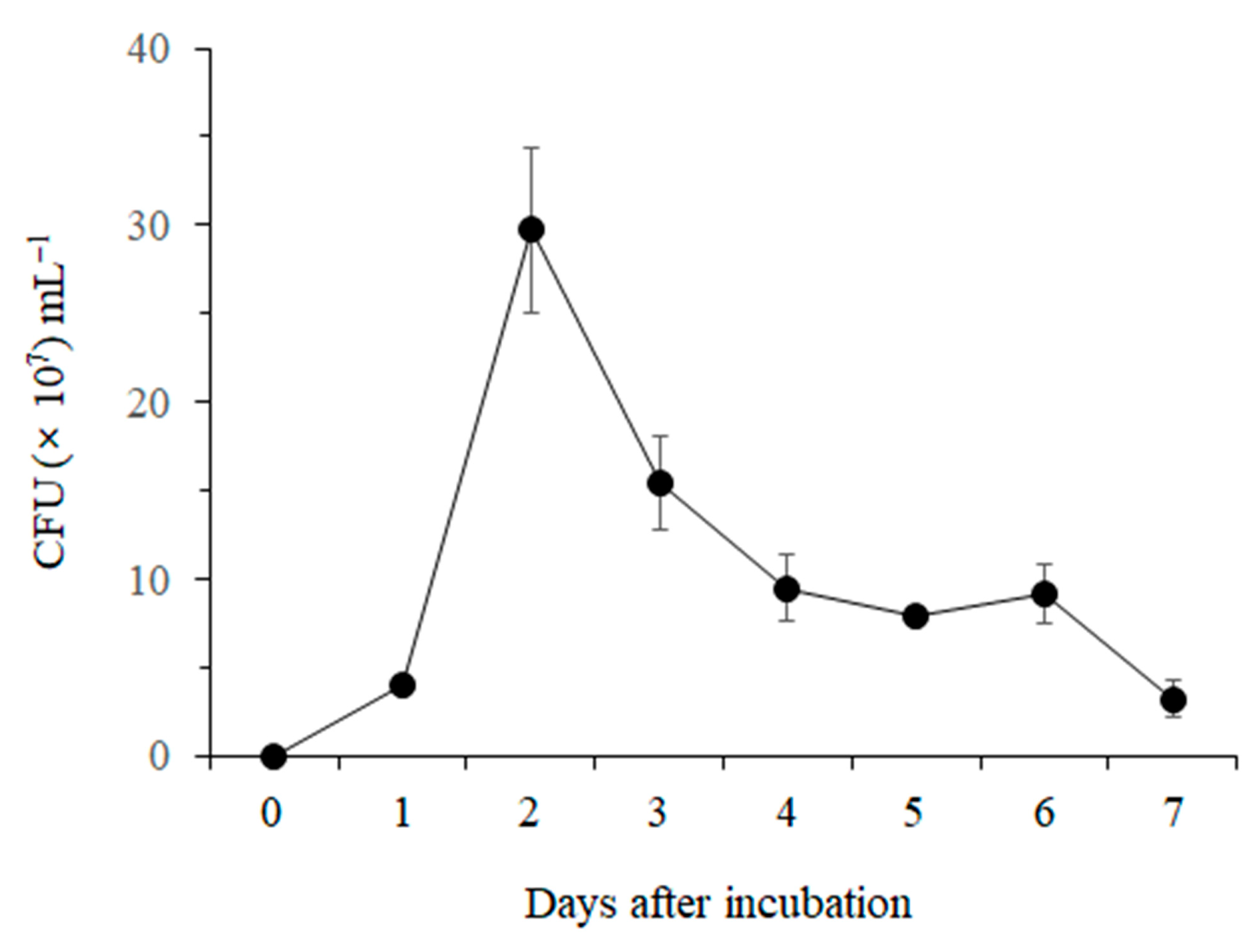
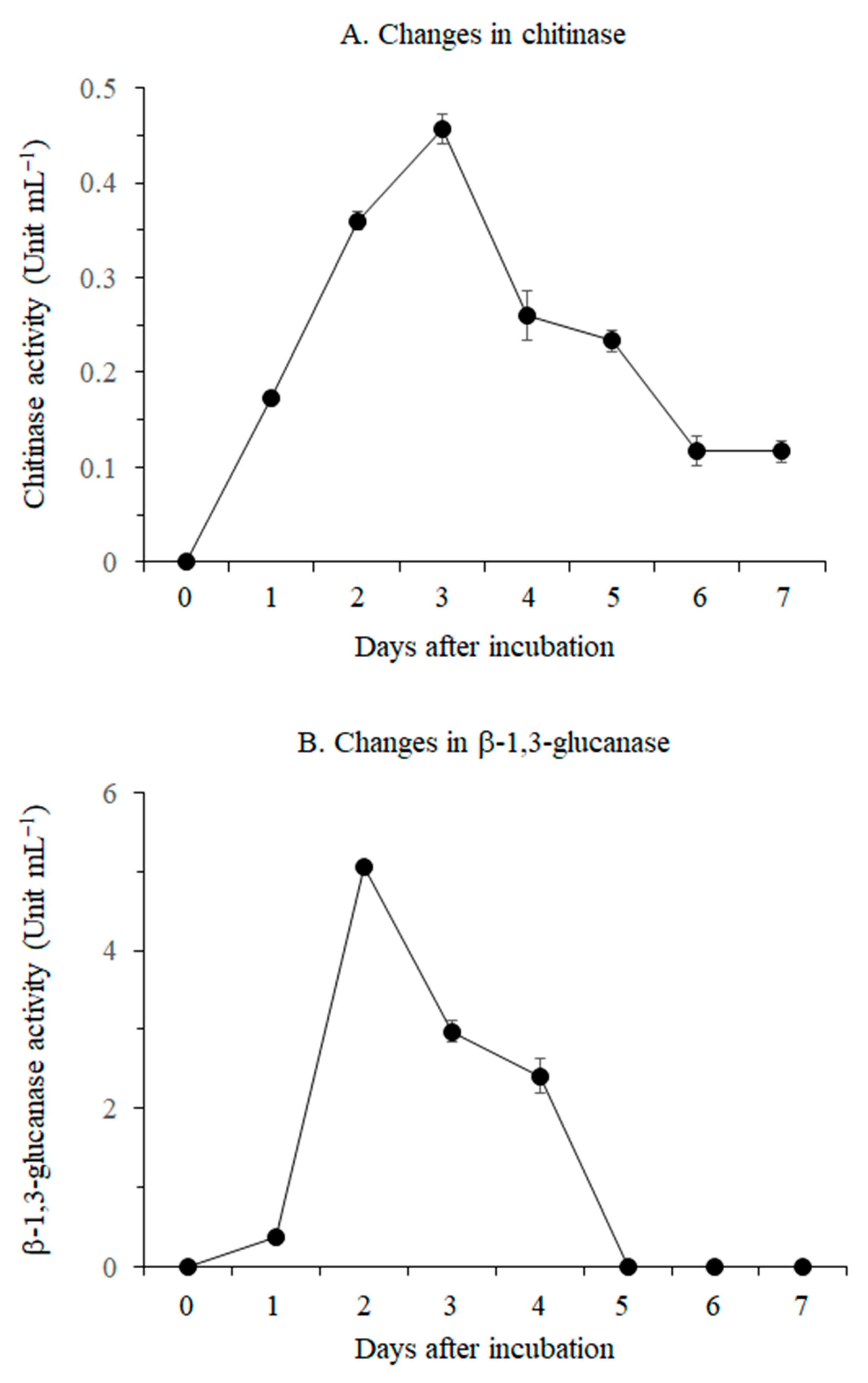
3.1.1. Lytic Enzyme Production
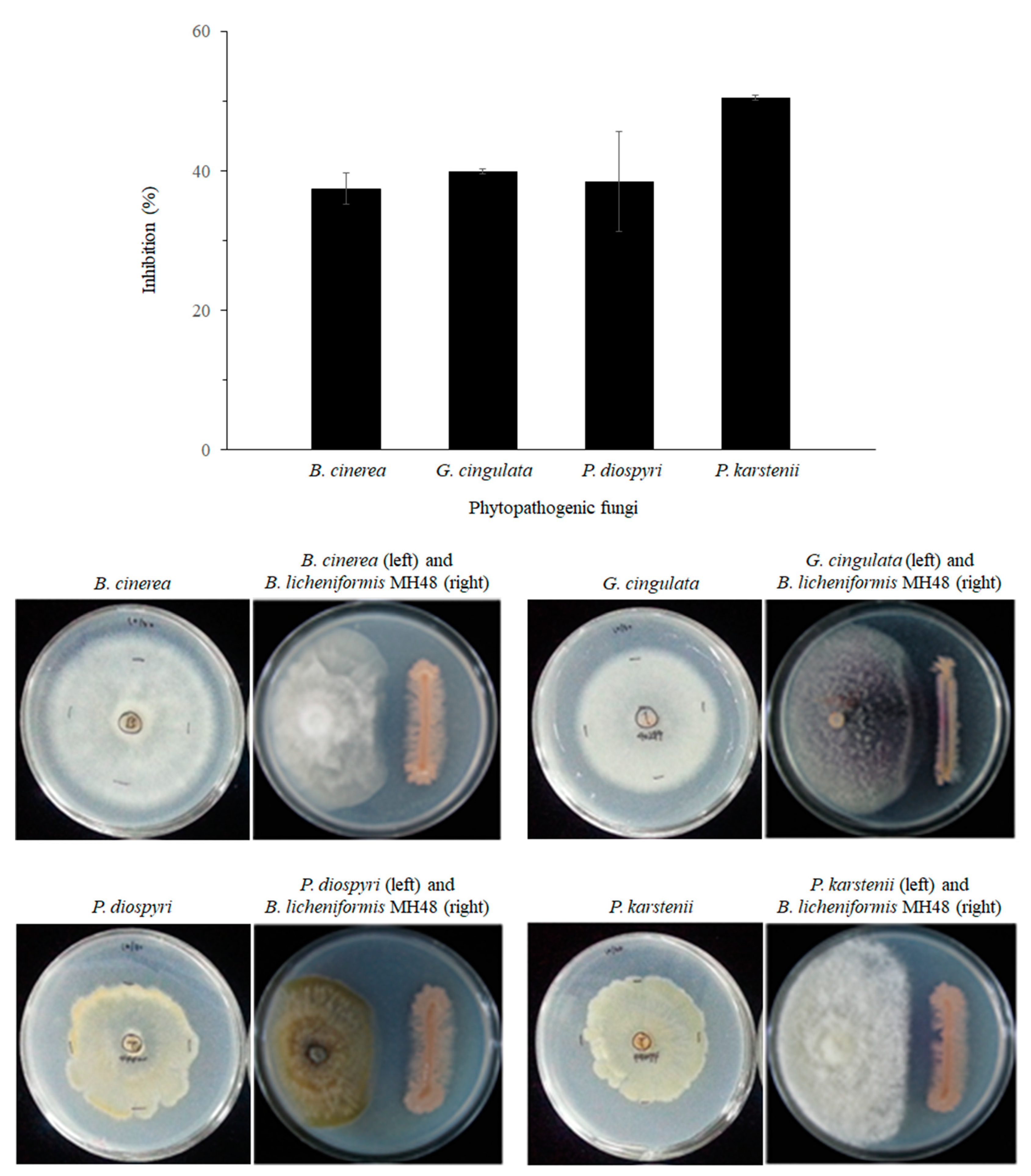
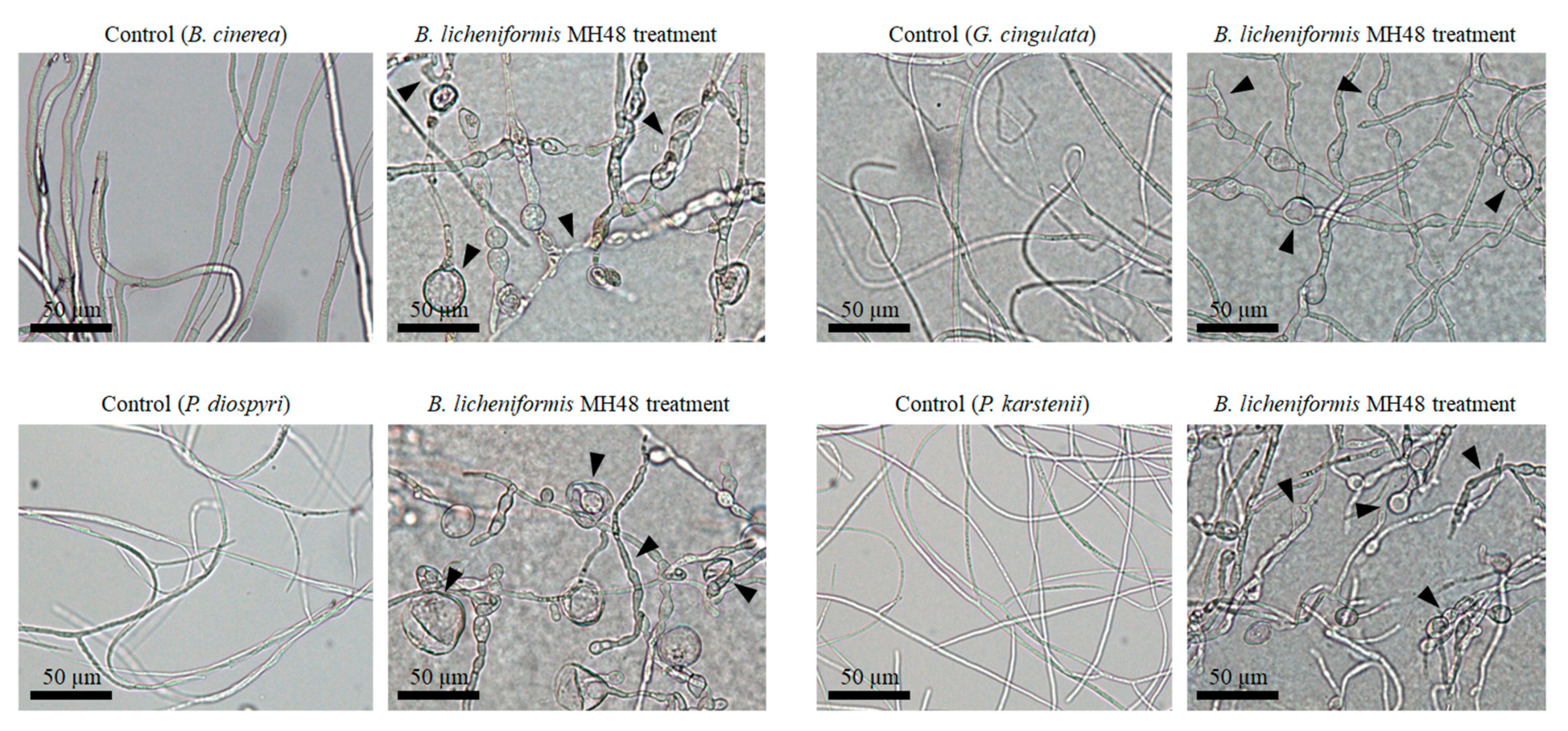
3.1.2. Antagonistic Activity against Foliar Fungal Pathogens
3.2. Effect of B. licheniformis MH48 on Growth Promotion of C. oleifera Seedlings
3.2.1. Chemical Properties in Soils
3.2.2. Dry Weight, Nutrient Concentration and Nutrient Content of C. oleifera Seedlings
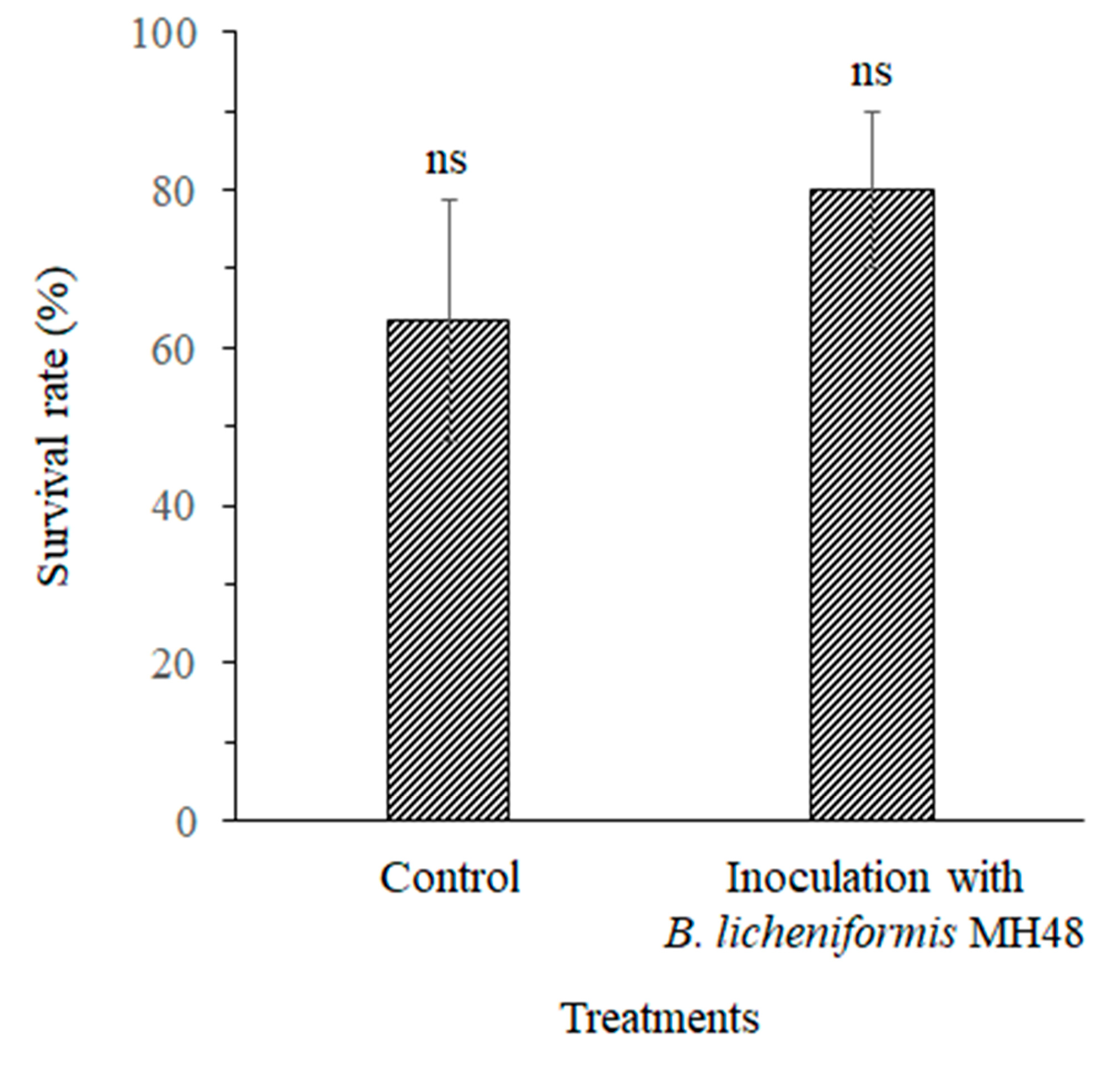
3.2.3. Survival Rate of C. oleifera Seedlings
4. Discussion
4.1. Antagonistic Activity of B. licheniformis MH48 against Foliar Fungal Pathogens
4.2. Growth Promotion of C. oleifera Seedlings by B. licheniformis MH48
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tai, Y.; Wei, C.; Yang, H.; Zhang, L.; Chen, Q.; Deng, W.; Wei, S.; Zhang, J.; Fang, C.; Ho, C.; et al. Transcriptomic and phytochemical analysis of the biosynthesis of characteristic constituents in tea (Camellia sinensis) compared with oil tea (Camellia oleifera). BMC Plant Biol. 2015, 15, 190. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; He, L.; Chen, Y.; Wu, L.; Wang, L.; Liu, Z. Anti-inflammatory and antioxidative effects of Camellia oleifera Abel components. Future Med. Chem. 2017, 9, 2069–2079. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.A.; Li, H.; Zhou, G.Y. Specific and rapid detection of Camellia oleifera anthracnose pathogen by nested-PCR. Afr. J. Biotechnol. 2009, 8, 1056–1061. [Google Scholar]
- Park, H.G.; Jeong, M.H.; Ahn, Y.S. Inoculation with Bacillus licheniformis MH48 to improve Camellia japonica seedling development in coastal lands. Turk. J. Agric. 2017, 41, 381–388. [Google Scholar] [CrossRef]
- Park, H.G.; Lee, Y.S.; Kim, K.Y.; Park, Y.S.; Park, K.H.; Han, T.H.; Park, C.M.; Ahn, Y.S. Inoculation with Bacillus licheniformis MH48 promotes nutrient uptake in seedlings of the ornamental plant Camellia japonica grown in Korean reclaimed coastal lands. Hortic. Sci. Technol. 2017, 35, 11–20. [Google Scholar] [CrossRef]
- Qiu, N.; Lu, Q.; Lu, C. Photosynthesis, photosystem II efficiency and the xanthophyll cycle in the salt-adapted halophyte Atriplex centralasiatica. New Phytol. 2003, 159, 479–486. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Koyro, H.W. Effect of salinity on growth, photosynthesis, water relations and solute composition of the potential cash crop halophyte Plantago coronopus (L.). Environ. Exp. Bot. 2006, 56, 136–146. [Google Scholar] [CrossRef]
- Rojas-Tapias, D.; Moreno-Galván, A.; Pardo-Díaz, S.; Obando, M.; Rivera, D.; Bonilla, R. Effect of inoculation with plant growth-promoting bacteria (PGPB) on amelioration of saline stress in maize (Zea mays). Appl. Soil Ecol. 2012, 61, 264–272. [Google Scholar] [CrossRef]
- Bai, Y.; Kissoudis, C.; Yan, Z.; Visser, R.G.F.; van der Linden, G. Plant behaviour under combined stress: Tomato responses to combined salinity and pathogen stress. Plant J. 2018, 93, 781–793. [Google Scholar] [CrossRef]
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical pesticides and human health: The urgent need for a new concept in agriculture. Front. Public Health 2016, 4, 148. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012. [Google Scholar] [CrossRef]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Jeong, M.H.; Lee, Y.S.; Cho, J.Y.; Ahn, Y.S.; Moon, J.H.; Hyun, H.N.; Cha, G.S.; Kim, K.Y. Isolation and characterization of metabolites from Bacillus licheniformis MH48 with antifungal activity against plant pathogens. Microb. Pathog. 2017, 110, 645–653. [Google Scholar] [CrossRef]
- van Loon, L.C.; Bakker, P.A.; Pieterse, C.M. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Maung, C.E.H.; Choi, T.G.; Nam, H.H.; Kim, K.Y. Role of Bacillus amyloliquefaciens Y1 in the control of Fusarium wilt disease and growth promotion of tomato. Biocontrol Sci. Technol. 2017, 27, 1400–1415. [Google Scholar] [CrossRef]
- Li, H.; Zhou, G.; Zhang, H.; Song, G.; Liu, J. Study on isolated pathogen of leaf blight and screening antagonistic bacteria from healthy leaves of Camellia oleifera. Afr. J. Agric. Res. 2011, 6, 4560–4566. [Google Scholar]
- Yu, J.; Wu, Y.; He, Z.; Li, M.; Zhu, K.; Gao, B. Diversity and antifungal activity of endophytic fungi associated with Camellia oleifera. Mycobiology 2018, 46, 85–91. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Kloepper, J.W. Plant-microbes interactions in enhanced fertilizer-use efficiency. Appl. Microbiol. Biotechnol. 2009, 85, 1–12. [Google Scholar] [CrossRef]
- Weyens, N.; van der Lelie, D.; Taghavi, S.; Newman, L.; Vangronsveld, J. Exploiting plant-microbe partnerships to improve biomass production and remediation. Trends Biotechnol. 2009, 27, 591–598. [Google Scholar] [CrossRef]
- Abbasi, M.K.; Sharif, S.; Kazmi, M.; Sultan, T.; Aslam, M. Isolation of plant growth promoting rhizobacteria from wheat rhizosphere and their effect on improving growth, yield, and nutrient uptake of plants. Plant Biosyst. 2011, 145, 159–168. [Google Scholar] [CrossRef]
- Karakurt, H.; Kotan, R. Effects of plant growth promoting rhizobacteria on fruit set, pomological and chemical characteristics, color values, and vegetative growth of sour cherry (Prunus cerasus cv. Kutahya). Turk. J. Biol. 2011, 35, 283–291. [Google Scholar] [CrossRef]
- Lingappa, Y.; Lockwood, J. Chitin media for selective isolation and culture of Actinomycetes. Phytopathology 1962, 52, 317–323. [Google Scholar]
- Liang, Z.C.; Hseu, R.S.; Wang, H.H. Partial purification and characterization of a 1,3-β-d-glucanase from Ganoderma tsugae. J. Ind. Microbiol. Biotechnol. 1995, 14, 5–9. [Google Scholar] [CrossRef]
- Skidmore, A.; Dickinson, C. Colony interactions and hyphal interference between Septoria nodorum and phylloplane fungi. Trans. Br. Mycol. Soc. 1976, 66, 57–64. [Google Scholar] [CrossRef]
- Annual Climatological Report (2014). Available online: http://www.kma.go.kr/index.jsp (accessed on 1 May 2018). (In Korean).
- Annual Climatological Report (2015). Available online: http://www.kma.go.kr/index.jsp (accessed on 1 May 2018). (In Korean).
- Mulvaney, R.L. Nitrogen-Inorganic Forms. In Methods of Soil Analysis, Part 3, Chemical Methods; Spark, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpoor, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 1123–1184. ISBN 978-0-89118-825-4. [Google Scholar]
- Jamal, Q.; Lee, Y.S.; Jeon, H.D.; Park, Y.S.; Kim, K.Y. Isolation and biocontrol potential of Bacillus amyloliquefaciens Y1 against fungal plant pathogens. Korean J. Soil Sci. Fertil. 2015, 48, 485–491. [Google Scholar] [CrossRef]
- Jeong, M.H.; Yang, S.Y.; Lee, Y.S.; Ahn, Y.S.; Park, Y.S.; Han, H.R.; Kim, K.Y. Selection and characterization of Bacillus licheniformis MH48 for the biocontrol of pine wood nematode (Bursaphelenchus xylophilus). J. Korean For. Soc. 2015, 104, 512–518, (In Korean with English Abstract). [Google Scholar] [CrossRef]
- Jeon, H.D. Characterization and Study on Onsite Mass Cultivation of Bacillus amyloliquefaciens Y1. Master’s Thesis, Chonnam National University, Gwangju, Korea, February 2017. (In Korean with English Abstract). [Google Scholar]
- Salama, R.B.; Otto, C.J.; Fitzpatrick, R.W. Contributions of groundwater conditions to soil and water salinization. Hydrogeol. J. 1999, 7, 46–64. [Google Scholar] [CrossRef]
- Rengasamy, P. World salinization with emphasis on Australia. J. Exp. Bot. 2006, 57, 1017–1023. [Google Scholar] [CrossRef] [Green Version]
- Morgan, P.W.; Drew, M.C. Ethylene and plant response to stress. Physiol. Plant. 1997, 100, 620–630. [Google Scholar] [CrossRef]
- Amira, M.S.; Qados, A. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar] [CrossRef]
- Yao, L.; Wu, Z.; Zheng, Y.; Kaleem, I.; Li, C. Growth promotion and protection against salt stress by Pseudomonas putida Rs-198 on cotton. Eur. J. Soil Biol. 2010, 46, 49–54. [Google Scholar] [CrossRef]
- Puente, M.E.; Bashan, Y.; Li, C.Y.; Lebsky, V.K. Microbial populations and activities in the rhizoplane of rock-weathering desert plants. I. Root colonization and weathering of igneous rocks. Plant Biol. 2004, 6, 629–642. [Google Scholar] [CrossRef]
- Puente, M.E.; Li, C.Y.; Bashan, Y. Microbial populations and activities in the rhizoplane of rock-weathering desert plants. II. Growth promotion of cactus seedlings. Plant Biol. 2004, 6, 643–650. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Inoculation with B. licheniformis MH48 | |
|---|---|---|
| pH | 6.30 ± 0.15 * | 7.29 ± 0.21 * |
| Total nitrogen (g kg−1) | 0.47 ± 0.21 * | 1.35 ± 0.57 * |
| Total phosphorus (g kg−1) | 0.21 ± 0.07 * | 2.48 ± 0.93 * |
| Control | Inoculation with B. licheniformis MH48 | |
|---|---|---|
| Dry weight (g plant−1) | ||
| Leaf | 0.91 ± 0.23 * | 2.46 ± 1.00 * |
| Shoot | 5.25 ± 1.51 * | 10.07 ± 4.60 * |
| Root | 4.45 ± 0.83 * | 7.42 ± 0.93 * |
| Total nitrogen concentration (% plant−1) | ||
| Leaf | 1.57 ± 0.18 * | 2.30 ± 0.37 * |
| Shoot | 0.94 ± 0.10 | 1.28 ± 0.28 |
| Root | 1.09 ± 0.13 * | 1.72 ± 0.32 * |
| Total phosphorus concentration (% plant−1) | ||
| Leaf | 0.17 ± 0.03 | 0.21 ± 0.05 |
| Shoot | 0.21 ± 0.05 | 0.24 ± 0.04 |
| Root | 0.19 ± 0.03 | 0.21 ± 0.03 |
| Nutrient content (mg plant−1) | ||
| Total nitrogen | 112.95 ± 18.11 * | 317.57 ± 126.71 * |
| Total phosphorus | 20.84 ± 3.11 * | 46.86 ± 13.11 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Won, S.-J.; Kwon, J.-H.; Kim, D.-H.; Ahn, Y.-S. The Effect of Bacillus licheniformis MH48 on Control of Foliar Fungal Diseases and Growth Promotion of Camellia oleifera Seedlings in the Coastal Reclaimed Land of Korea. Pathogens 2019, 8, 6. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8010006
Won S-J, Kwon J-H, Kim D-H, Ahn Y-S. The Effect of Bacillus licheniformis MH48 on Control of Foliar Fungal Diseases and Growth Promotion of Camellia oleifera Seedlings in the Coastal Reclaimed Land of Korea. Pathogens. 2019; 8(1):6. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8010006
Chicago/Turabian StyleWon, Sang-Jae, Jun-Hyeok Kwon, Dong-Hyun Kim, and Young-Sang Ahn. 2019. "The Effect of Bacillus licheniformis MH48 on Control of Foliar Fungal Diseases and Growth Promotion of Camellia oleifera Seedlings in the Coastal Reclaimed Land of Korea" Pathogens 8, no. 1: 6. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8010006