Enniatin Production Influences Fusarium avenaceum Virulence on Potato Tubers, but not on Durum Wheat or Peas

 ,
,
Abstract
:1. Introduction
2. Results
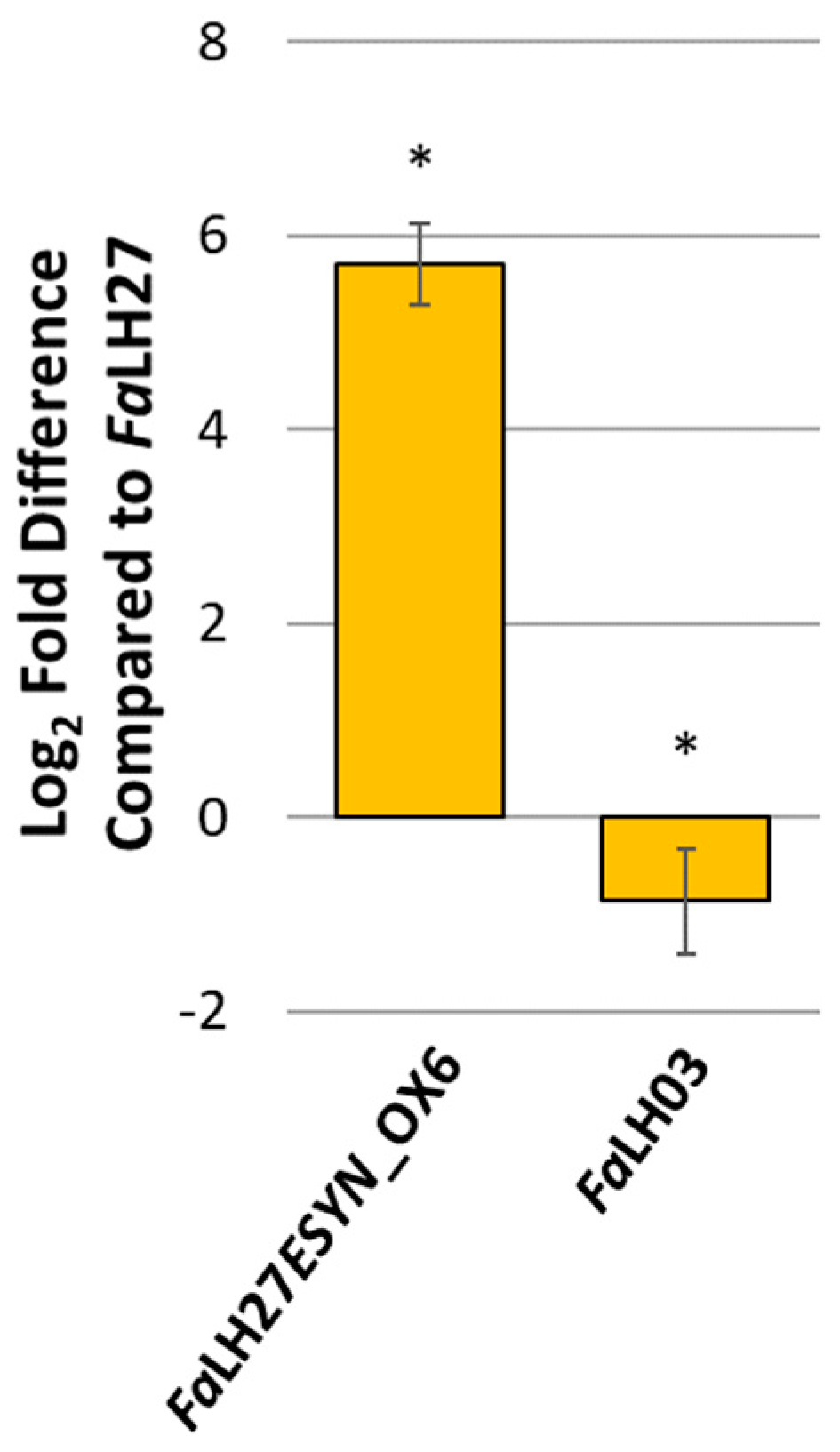
2.1. Generation of F. avenaceum ESYN1 Disruption and Overexpression Strains
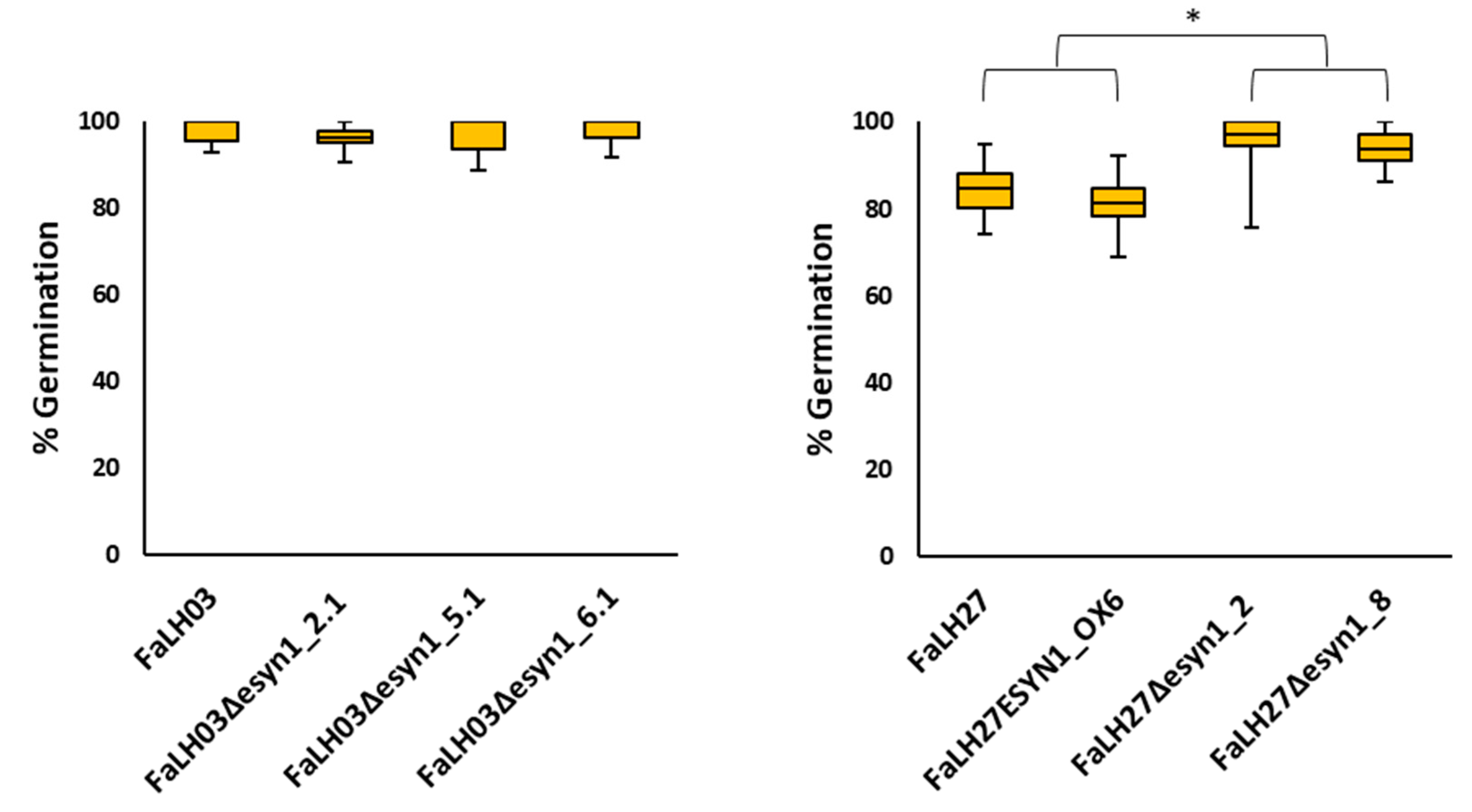
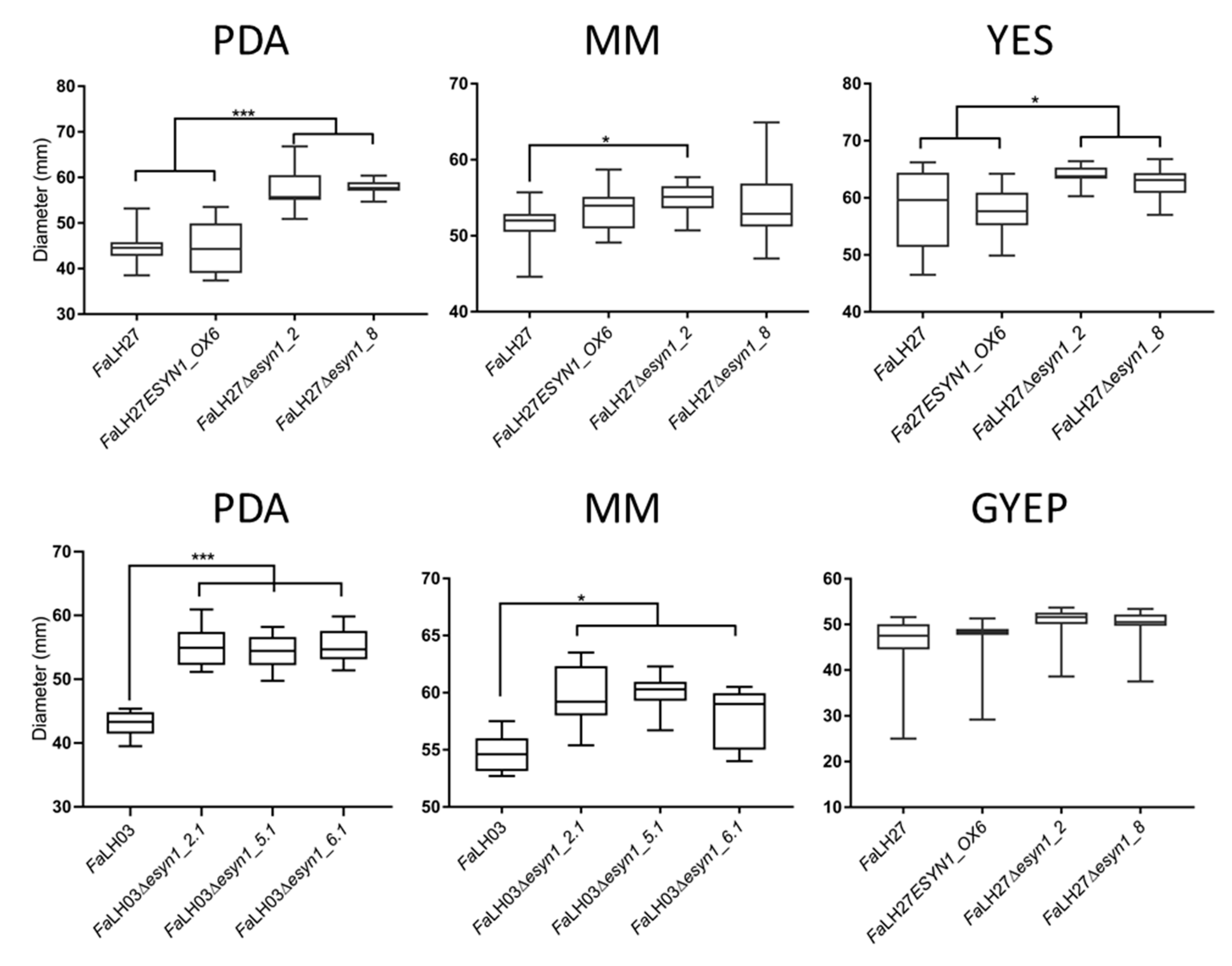
2.2. Genetic Disruption of ESYN1 Has a Small Positive Effect on Some Fungal Growth Parameters
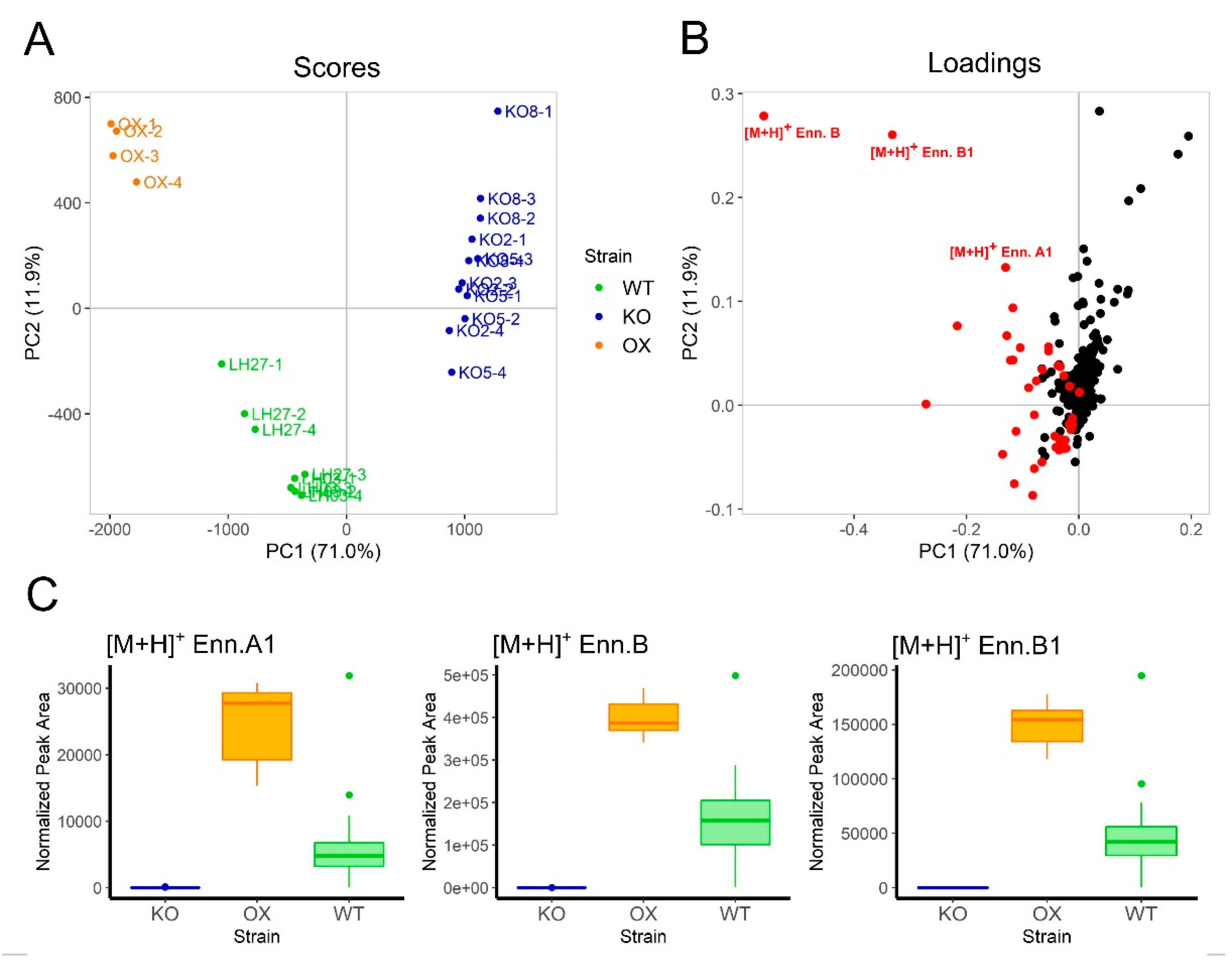
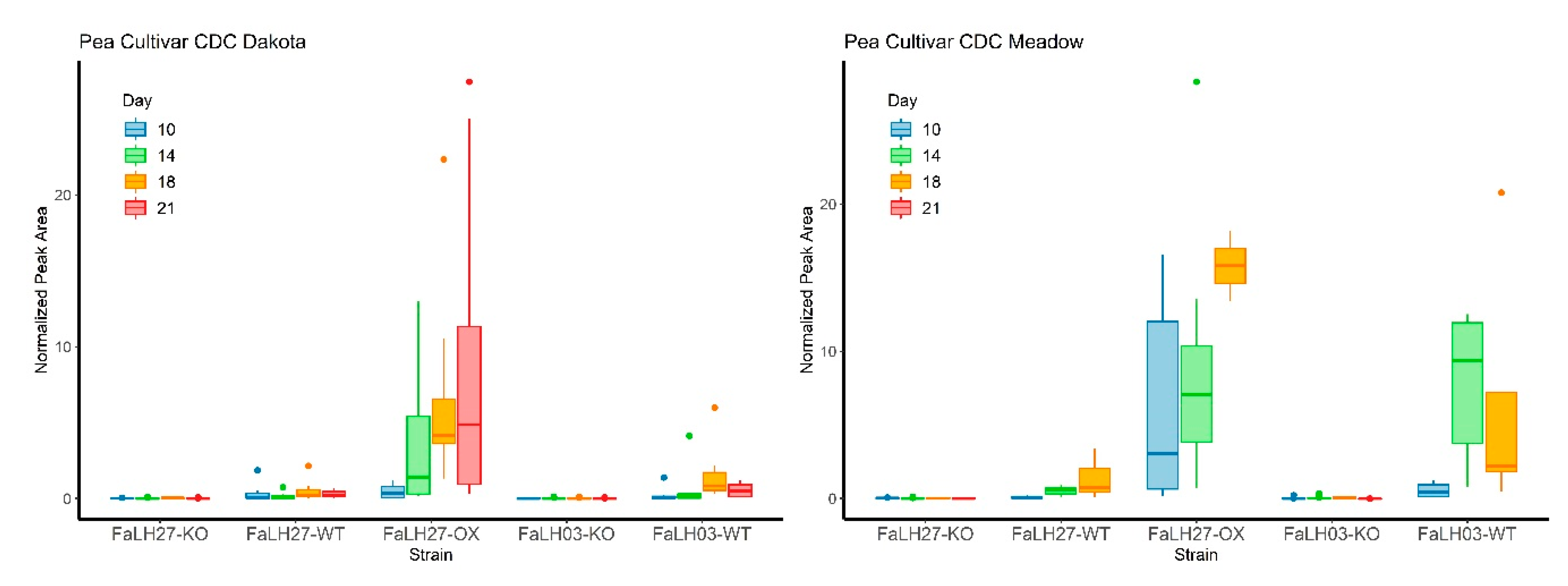
2.3. Metabolic Analysis of Wild-type, ∆esyn1, and ESYN1_OX Isolates
2.4. Impact of ESYN1 Disruption or Overexpression on F. avenaceum Aggressiveness in Different Field Crops
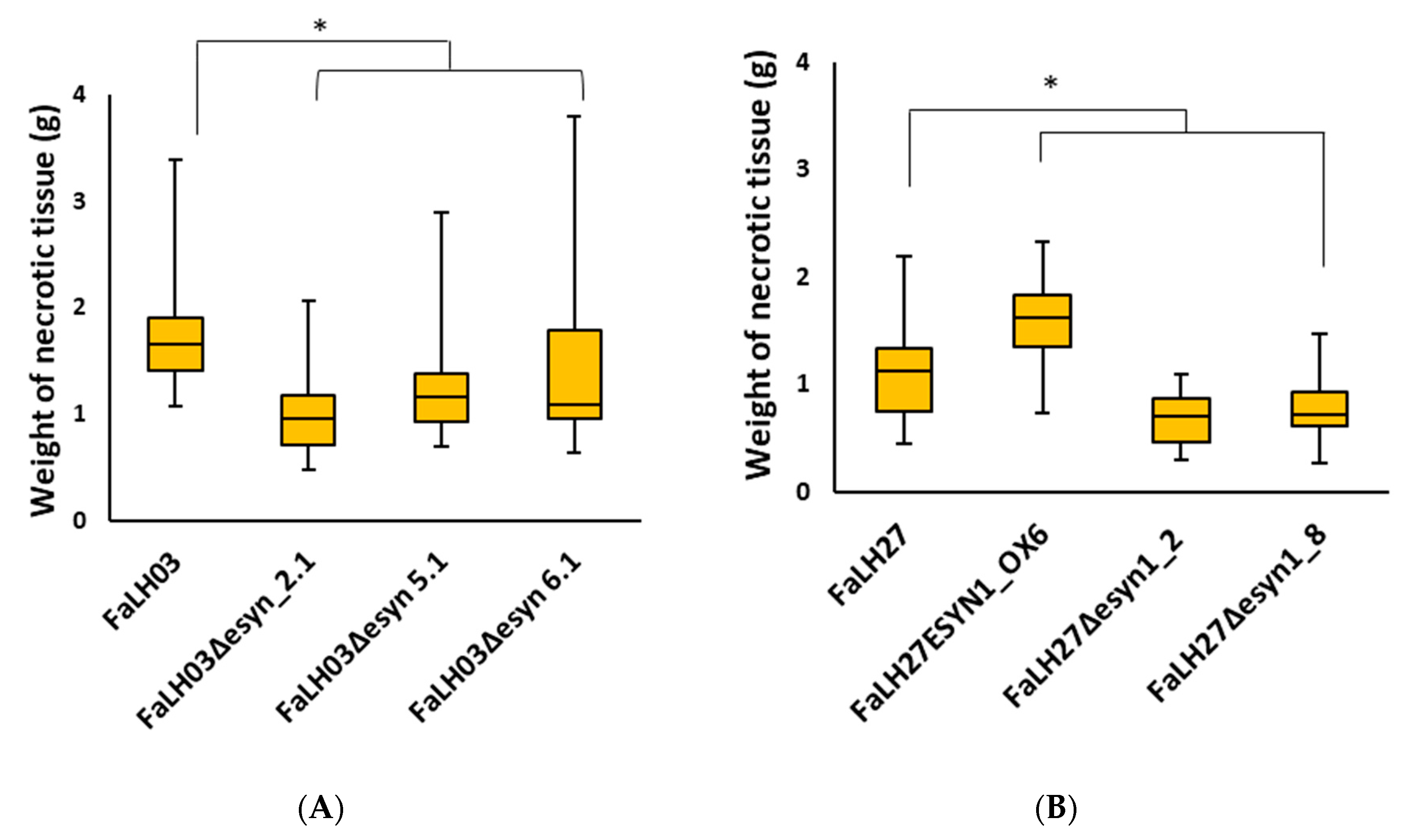
2.4.1. Potato Tuber Necrosis is Increased in the ESYN1 Overexpression Isolate and Reduced in the Genetic Disruption Mutants
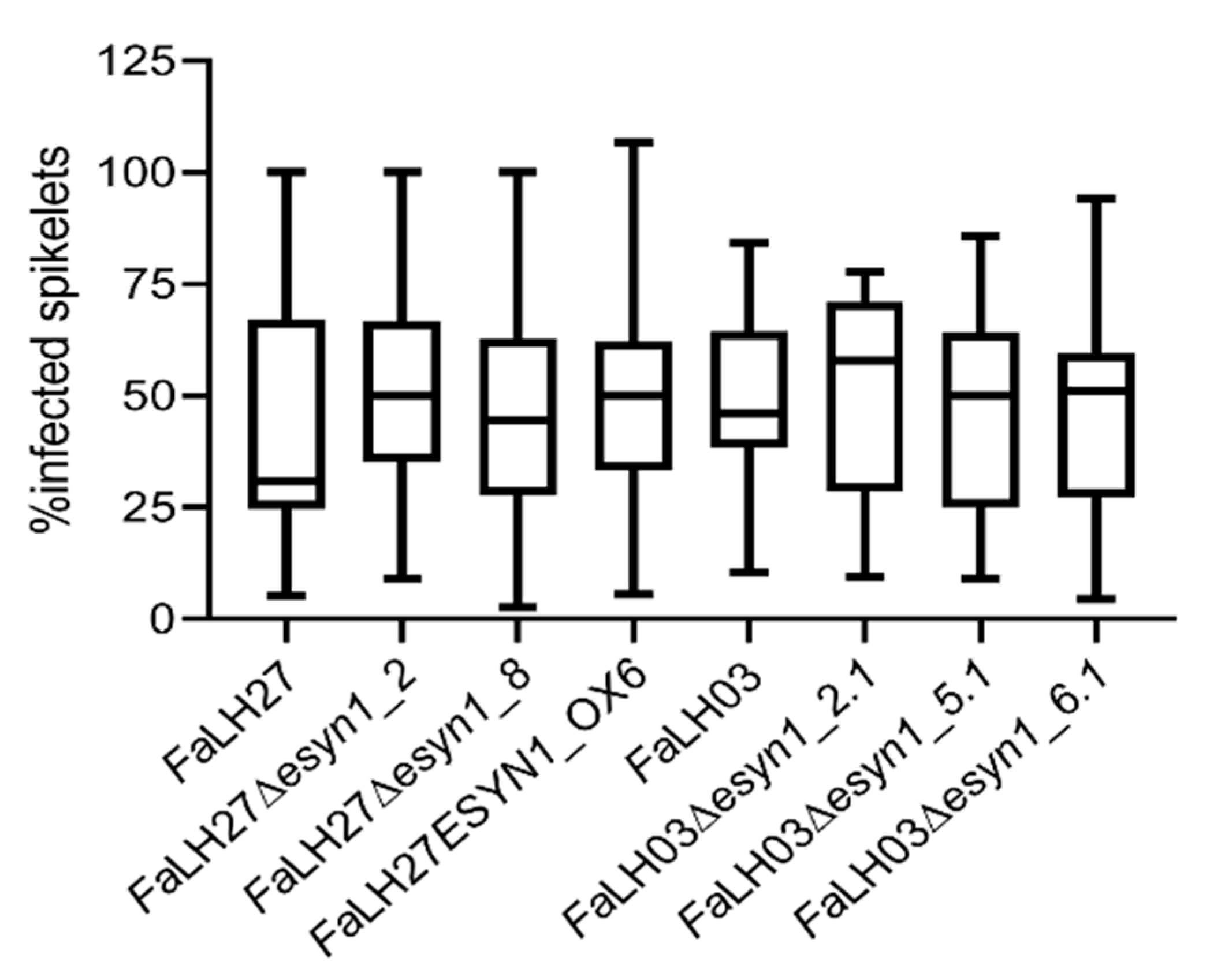
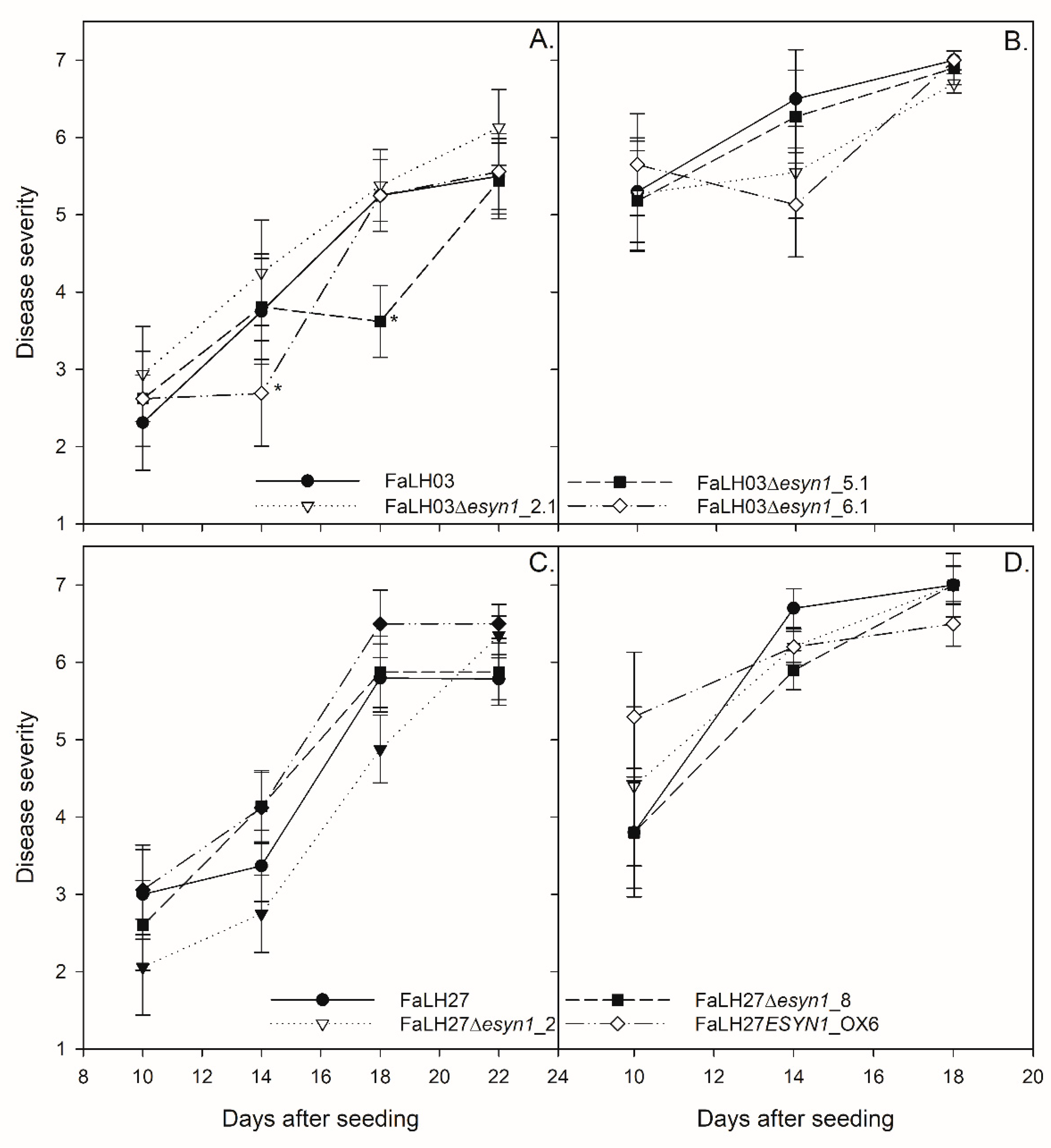
2.4.2. No Difference Was Observed in FHB Disease Spread Among ESYN1-Modified Isolates and Wild-Type F. avenaceum
2.4.3. No Difference Was Observed in Pea Emergence or Root Rot Symptoms Among ESYN1-modified Isolates and Wild-type F. avenaceum
3. Discussion
4. Materials and Methods
4.1. F. avenaceum Strains and Spore Production
4.2. Generation and Confirmation of ESYN1 Knockout and Overexpression Mutants
4.3. Spore Germination, Fungal Culturing, and Mycelial Growth Assays
4.4. Potato Tuber Necrosis Assays
4.5. FHB Assays in Durum Wheat
4.6. Pea Root Assays
4.7. Metabolic Profiling
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Esmaeili Taheri, A.; Chatterton, S.; Foroud, N.A.; Gossen, B.D.; McLaren, D.L. Identification and community dynamics of fungi associated with root, crown, and foot rot of field pea in western Canada. Eur. J. Plant Pathol. 2017, 147, 489–500. [Google Scholar] [CrossRef]
- García-Núñez, H.; Martínez-Campos, A.R.; López-Orona, C.A.; Díaz-Mínguez, J.M. First report of Fusarium avenaceum causing stalk and root rot in commercial potato crops (Solanum tuberosum) in the Toluca Valley, Mexico. Plant Dis. 2016, 100, 2170. [Google Scholar] [CrossRef]
- Koike, S.T.; Daugovish, O.; Kirkpatrick, S.C.; Henry, P.M.; Gordon, T.R. First report of crown and root rot of leek caused by Fusarium avenaceum in California. Plant Dis. 2015, 99, 1864. [Google Scholar] [CrossRef]
- Foroud, N.A.; Chatterton, S.; Reid, L.M.; Turkington, T.K.; Tittlemier, S.A.; Gräfenhan, T. Fusarium diseases of Canadian grain crops: impact and disease management strategies. In Future Challenges in Crop Protection Against Fungal Pathogens; Goyal, A., Manoharachary, C., Eds.; Springer: New York, NY, USA, 2014; pp. 267–316. [Google Scholar]
- Stevenson, W.R.; Loria, R.; Franc, G.D.; Weingartner, D.P. Compendium of Potato Diseases, 2nd ed.; American Phytopathological Society Press: Saint Paul, MN, USA, 2001. [Google Scholar]
- Choiseul, J.; Allen, L.; Carnegie, S.F. Fungi causing dry tuber rots of seed potatoes in storage in Scotland. Potato Res. 2006, 49, 241–253. [Google Scholar] [CrossRef]
- Wharton, P.; Hammerschmidt, R.; Kirk, W. Fusarium Dry Rot. Available online: https://www.canr.msu.edu/outreach/uploads/files/fusarium-dry-rot-bulletin.pdf (accessed on 14 January 2020).
- Sandipan, P.B.; Solanki, B.P.; Patel, N.N.; Patel, R.L.; Verma, P.D.; Desai, H.R. Efficacy of different fungicides against dry rot pathogen of potato caused by Fusarium sp. under in vitro condition. Cercet. Agron. Mold. 2016, 49, 69–74. [Google Scholar]
- Estrada, R., Jr.; Gudmestad, N.C.; Rivera, V.V.; Secor, G.A. Fusarium graminearum as a dry rot pathogen of potato in the USA: Prevalence, comparison of host isolate aggressiveness and factors affecting aetiology. Plant Pathol. 2010, 59, 1114–1120. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Plattner, R.D. Trichothecene toxin production by strains of Gibberella pulicaris (Fusarium sambucinum) in liquid culture and in potato tubers. J. Agr. Food. Chem. 1989, 37, 388–392. [Google Scholar] [CrossRef]
- Herrmann, M.; Zocher, R.; Haese, A. Enniatin production by Fusarium strains and its effect on potato tuber tissue. Appl. Environ. Microbiol. 1996, 62, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, M.; Zocher, R.; Haese, A. Effect of disruption of the enniatin synthetase gene on the virulence of Fusarium avenaceum. Mol. Plant-Microbe Interact. 1996, 9, 226–232. [Google Scholar] [CrossRef]
- Song, H.-H.; Lee, H.-S.; Jeong, J.-H.; Park, H.-S.; Lee, C. Diversity in beauvericin and enniatins H, I, and MK1688 by Fusarium oxysporum isolated from potato. Int. J. Food Microbiol. 2008, 122, 296–301. [Google Scholar] [CrossRef]
- Peters, J.C.; Lees, A.K.; Cullen, D.W.; Sullivan, L.; Stroud, G.P.; Cunnington, A.C. Characterization of Fusarium spp. responsible for causing dry rot of potato in Great Britain. Plant Pathol. 2008, 57, 262–271. [Google Scholar] [CrossRef]
- Hanson, L.; Schwager, S.; Loria, R. Sensitivity to thiabendazole in Fusarium species associated with dry rot of potato. Phytopathology 1996, 86378, 384. [Google Scholar]
- Gachango, E.; Hanson, L.E.; Rojas, A.; Hao, J.J.; Kirk, W.W. Fusarium spp. causing dry rot of seed potato tubers in Michigan and their sensitivity to fungicides. Plant Dis. 2012, 96, 1767–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterton, S.; Bowness, R.; Harding, M.W. First report of root rot of field pea caused by Aphanomyces euteiches in Alberta, Canada. Plant Dis. 2015, 99, 288. [Google Scholar] [CrossRef]
- Gossen, B.D.; Conner, R.L.; Chang, K.-F.; Pasche, J.S.; McLaren, D.L.; Henriquez, M.A.; Chatterton, S.; Hwang, S.-F. Identifying and managing root rot of pulses on the northern Great Plains. Plant Dis. 2016, 100, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Chatterton, S.; Harding, M.W.; Bowness, R.; McLaren, D.L.; Banniza, S.; Gossen, B.D. Importance and causal agents of root rot on field pea and lentil on the Canadian prairies, 2014–2017. Can. J. Plant Pathol. 2019, 41, 98–114. [Google Scholar] [CrossRef]
- Chittem, K.; Mathew, F.M.; Gregoire, M.; Lamppa, R.S.; Chang, Y.W.; Markell, S.G.; Bradley, C.A.; Barasubiye, T.; Goswami, R.S. Identification and characterization of Fusarium spp. associated with root rots of field pea in North Dakota. Eur. J. Plant Pathol. 2015, 143, 641–649. [Google Scholar] [CrossRef]
- Baćanović-Šišić, J.; Šišić, A.; Schmidt, J.H.; Finckh, M.R. Identification and characterization of pathogens associated with root rot of winter peas grown under organic management in Germany. Eur. J. Plant. Pathol. 2018, 151, 745–755. [Google Scholar] [CrossRef]
- Persson, L.; Bødker, L.; Larsson-Wikström, M. Prevalence and pathogenicity of foot and root rot pathogens of pea in southern Scandinavia. Plant Dis. 1997, 81, 171–174. [Google Scholar] [CrossRef] [Green Version]
- Safarieskandari, S. Towards development of a disease risk model for pea root disease. Doctor’s Dissertation, The University of Alberta, Edmonton, AB, Canada, 2019. [Google Scholar]
- Kazan, K.; Gardiner, D.M. Fusarium crown rot caused by Fusarium pseudograminearum in cereal crops: Recent progress and future prospects. Mol. Plant Pathol. 2018, 19, 1547–1562. [Google Scholar] [CrossRef] [Green Version]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (scab) in small grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Khan, M.R.; Fischer, S.; Egan, D.; Doohan, F.M. Biological control of Fusarium seedling blight disease of wheat and barley. Phytopathology 2006, 96, 386–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windels, C.E. Economic and social impacts of Fusarium head blight: Changing farms and rural communities in the Northern Great Plains. Phytopathology 2000, 90, 17–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestka, J. Toxicological mechanisms and potential health effects of deoxynivalenol and nivalenol. World Mycotoxin J. 2010, 3, 323–347. [Google Scholar] [CrossRef]
- Santini, A.; Meca, G.; Uhlig, S.; Ritieni, A. Fusaproliferin, beauvericin and enniatins: Occurrence in food – a review. World Mycotoxin J. 2012, 5, 71–81. [Google Scholar] [CrossRef]
- Bai, G.-H.; Desjardins, A.E.; Plattner, R.D. Deoxynivalenol-nonproducing Fusarium graminearum causes initial infection, but does not cause disease spread in wheat spikes. Mycopathologia 2002, 153, 91–98. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Gräfenhan, T.; Patrick, S.K.; Roscoe, M.; Trelka, R.; Gaba, D.; Chan, J.M.; McKendry, T.; Clear, R.M.; Tittlemier, S.A. Fusarium damage in cereal grains from Western Canada. 1. Phylogenetic analysis of moniliformin-producing Fusarium species and their natural occurrence in mycotoxin-contaminated wheat, oats, and rye. J. Agric. Food Chem. 2013, 61, 5425–5437. [Google Scholar]
- Köhl, J.; de Haas, B.H.; Kastelein, P.; Burgers, S.L.G.E.; Waalwijk, C. Population dynamics of Fusarium spp. and Microdochium nivale in crops and crop residues of winter wheat. Phytopathology 2007, 97, 971–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlig, S.; Jestoi, M.; Parikka, P. Fusarium avenaceum—The North European situation. Int. J. Food Microbiol. 2007, 119, 17–24. [Google Scholar] [CrossRef]
- Tittlemier, S.A.; Roscoe, M.; Trelka, M.; Gaba, D.; Chan, J.; Patrick, S.; Sulyok, M.; Krska, R.; McKendry, T.; Gräfenhan, T. Fusarium damage in cereal grains from western Canada. 2. Occurrence of mycotoxins and their source organisms in Fusarium damaged durum wheat harvested in 2010. J. Agr. Food Chem. 2013, 61, 5438–5448. [Google Scholar] [CrossRef] [PubMed]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-occurrence of regulated, masked and emerging mycotoxins and secondary metabolites in finished feed and maize—An extensive survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aboukhaddour, R.; Kim, Y.M.; Strelkov, S.E. RNA-mediated gene silencing of ToxB in Pyrenophora tritici-repentis. Mol. Plant Pathol. 2012, 13, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef] [PubMed]
- Lysøe, E.; Harris, L.J.; Walkowiak, S.; Subramaniam, R.; Divon, H.H.; Riiser, E.S.; Llorens, C.; Gabaldón, T.; Kistler, H.C.; Jonkers, W.; et al. The genome of the generalist plant pathogen Fusarium avenaceum is enriched with genes involved in redox, signaling and secondary metabolism. PLoS ONE 2014, 9, e112703. [Google Scholar]
- Jestoi, M. Emerging Fusarium-mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- Kamyar, M.; Rawnduzi, P.; Studenik, C.R.; Kouri, K.; Lemmens-Gruber, R. Investigation of the electrophysiological properties of enniatins. Arch. Biochem. Biophys. 2004, 429, 215–223. [Google Scholar] [CrossRef]
- Cui, C.; Kim, K.S. Cation affinities of cyclohexadepsipeptide: Ab initio study. J. Phys. Chem. A 1999, 103, 2751–2755. [Google Scholar] [CrossRef]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A review of the mycotoxin enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef]
- Maranghi, F.; Tassinari, R.; Narciso, L.; Tait, S.; Rocca, C.L.; Felice, G.D.; Butteroni, C.; Corinti, S.; Barletta, B.; Cordelli, E.; et al. In vivo toxicity and genotoxicity of beauvericin and enniatins. Combined approach to study in vivo toxicity and genotoxicity of mycotoxins beauvericin (BEA) and enniatin B (ENNB). EFSA Support. Publ. 2018, 15, 1406E. [Google Scholar] [CrossRef]
- Huang, C.-H.; Wang, F.-T.; Chan, W.-H. Enniatin B1 exerts embryotoxic effects on mouse blastocysts and induces oxidative stress and immunotoxicity during embryo development. Environ. Toxicol. 2019, 34, 48–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, L.; Skjerve, E.; Eriksen, G.S.; Uhlig, S. Cytotoxicity of enniatins A, A1, B, B1, B2 and B3 from Fusarium avenaceum. Toxicon 2006, 47, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Renaud, J.B.; Kelman, M.J.; McMullin, D.R.; Yeung, K.K.-C.; Sumarah, M.W. Application of C8 liquid chromatography-tandem mass spectrometry for the analysis of enniatins and bassianolides. J. Chromatogr. A 2017, 1508, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, H.R.; Plattner, R.D. Enniatin production by Fusarium tricinctum and its effect on germinating wheat seeds. Phytopathology 1987, 77, 1483–1487. [Google Scholar] [CrossRef]
- Klymiuk, V.; Fatiukha, A.; Huang, L.; Wei, Z.-z.; Kis-Papo, T.; Saranga, Y.; Krugman, T.; Fahima, T. Durum wheat as a bridge between wild emmer wheat genetic resources and bread wheat. In Applications of Genetic and Genomic Research in Cereals; Miedaner, T., Korzun, V., Eds.; Woodhead Publishing: Cambrige, UK, 2019; pp. 201–230. [Google Scholar]
- Howlett, B.J. Secondary metabolite toxins and nutrition of plant pathogenic fungi. Curr. Opin. Plant Biol. 2006, 9, 371–375. [Google Scholar] [CrossRef]
- Nasmith, C.G.; Walkowiak, S.; Wang, L.; Leung, W.W.Y.; Gong, Y.; Johnston, A.; Harris, L.J.; Guttman, D.S.; Subramaniam, R. Tri6 is a global transcription regulator in the phytopathogen Fusarium graminearum. PLoS Pathog. 2011, 7, e1002266. [Google Scholar] [CrossRef]
- Wang, L.; Mogg, C.; Walkowiak, S.; Joshi, M.; Subramaniam, R. Characterization of NADPH oxidase genes NoxA and NoxB in Fusarium graminearum. Can. J. Plant Pathol. 2014, 36, 12–21. [Google Scholar] [CrossRef]
- Voigt, C.A.; Schafer, W.; Salomon, S. A secreted lipase of Fusarium graminearum is a virulence factor required for infection of cereals. Plant J. 2005, 42, 364–375. [Google Scholar] [CrossRef]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant-Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Harris, L.J.; Alexander, N.J.; Saparno, A.; Blackwell, B.; McCormick, S.P.; Desjardins, A.E.; Robert, L.S.; Tinker, N.; Hattori, J.; Piche, C.; et al. A novel gene cluster in Fusarium graminearum contains a gene that contributes to butenolide synthesis. Fungal Genet. Biol. 2007, 44, 293–306. [Google Scholar] [CrossRef]
- Lysøe, E.; Bone, K.R.; Klemsdal, S.S. Real-time quantitative expression studies of the zearalenone biosynthetic gene cluster in Fusarium graminearum. Phytopathology 2009, 99, 176–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eudes, F.; Comeau, A.; Rioux, S.; Collin, J. Impact of trichothecenes on Fusarium head blight (Fusarium graminearum) development in spring wheat (Triticum aestivum). Can. J. Plant Pathol. 2001, 23, 318–322. [Google Scholar] [CrossRef]
- Maier, F.J.; Miedaner, T.; Hadeler, B.; Felk, A.; Salomon, S.; Lemmens, M.; Kassner, H.; Schaefer, W. Involvement of trichothecenes in fusarioses of wheat, barley and maize evaluated by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of different chemotype and virulence. Mol. Plant Pathol. 2006, 7, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Jansen, C.; von Wettstein, D.; Schaefer, W.; Kogel, K.H.; Felk, A.; Maier, F.J. Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum. Proc. Natl. Acad. Sci. USA 2005, 102, 16892–16897. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, A.E. Fusarium Mycotoxins: Chemistry, Genetics, and Biology; APS Press: Saint Paul, MN, USA, 2006. [Google Scholar]
- Fernandez, M.R.; Chen, Y. Pathogenicity of Fusarium species on different plant parts of spring wheat under controlled conditions. Plant Dis. 2005, 89, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willsey, T.L.; Chatterton, S.; Heynen, M.; Erickson, A. Detection of interactions between the pea root rot pathogens Aphanomyces euteiches and Fusarium spp. using a multiplex qPCR assay. Plant Pathol. 2018, 67, 1912–1923. [Google Scholar] [CrossRef]
- Kosiak, B.; Torp, M.; Skjerve, E.; Thrane, U. The prevalence and distribution of Fusarium species in Norwegian cereals: A Survey. Acta Agric. Scand. B 2003, 53, 168–176. [Google Scholar]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in cereal grains—An update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [Green Version]
- Tittlemier, S.A.; Blagden, R.; Chan, J.; Gaba, D.; McKendry, T.; Pleskach, K.; Roscoe, M. Fusarium and Alternaria mycotoxins present in Canadian wheat and durum harvest samples. Can. J. Plant Pathol. 2019, 41, 403–414. [Google Scholar] [CrossRef]
- Fernández-Blanco, C.; Font, G.; Ruiz, M.-J. Interaction effects of enniatin B, deoxinivalenol and alternariol in Caco-2 cells. Toxicol. Lett. 2016, 241, 38–48. [Google Scholar] [CrossRef]
- Alonso-Garrido, M.; Escrivá, L.; Manyes, L.; Font, G. Enniatin B induces expression changes in the electron transport chain pathway related genes in lymphoblastic T-cell line. Food Chem. Toxicol. 2018, 121, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.-J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium Pathogenomics. Annu. Rev. Plant Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foroud, N.A.; Pordel, R.; Goyal, R.K.; Ryabova, D.; Chatterton, S.; Kovalchuk, I. Chemical activation of the ethylene signalling pathway promotes wheat resistance to Fusarium graminearum. Phytopathology 2019, 109, 796–803. [Google Scholar] [CrossRef]
- Xue, A.G.; Armstrong, K.C.; Voldeng, H.D.; Fedak, G.; Babcock, C. Comparative aggressiveness of isolates of Fusarium spp. causing head blight on wheat in Canada. Can. J. Plant Pathol. 2004, 26, 81–88. [Google Scholar] [CrossRef]
- Frandsen, R.J.N.; Andersson, J.A.; Kristensen, M.B.; Giese, H. Efficient four fragment cloning for the construction of vectors for targeted gene replacement in filamentous fungi. BMC Mol. Biol. 2008, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, L.; Lysøe, E.; Larsen, J.; Khorsand-Jamal, P.; Nielsen, K.; Frandsen, R.J. Genetic transformation of Fusarium avenaceum by Agrobacterium tumefaciens mediated transformation and the development of a USER-Brick vector construction system. BMC Mol. Biol. 2014, 15, 15. [Google Scholar] [CrossRef] [Green Version]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006. [Google Scholar]
- Goyal, R.K.; Tulpan, D.; Chomistek, N.; González-Peña Fundora, D.; West, C.; Ellis, B.E.; Frick, M.; Laroche, A.; Foroud, N.A. Analysis of MAPK and MAPKK gene families in wheat and related Triticeae species. BMC Genom. 2018, 19, 178. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Bilgi, V.N.; Bradley, C.A.; Khot, S.D.; Grafton, K.F.; Rasmussen, J.B. Response of dry bean genotypes to Fusarium root rot, caused by Fusarium solani f. sp. phaseoli, under field and controlled conditions. Plant Dis. 2008, 92, 1197–1200. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial 1 | Trial 2 | ||||
|---|---|---|---|---|---|
| Dakota | Meadow | Dakota | Meadow | ||
| FaLH27 | 40.6a † | 29.2 | 81.2 | 6.2 | |
| FaLH27Δesyn1_2 | 71.9ab | 4.2 | 78.1 | 12.5 | |
| FaLH27Δesyn1_8 | 81.2b | 25.0 | 53.1 | 6.2 | |
| FaLH27ESYN1_OX6 | 71.9ab | 41.7 | 62.5 | 21.9 | |
| SE * | 6.48 | 10.38 | 8.87 | 7.50 | |
| FaLH03 | 81.2 | 29.2 | 50.0 | 0 | |
| FaLH03Δesyn1_2.1 | 75.0 | 20.8 | 40.6 | 0 | |
| FaLH03Δesyn1_5.1 | 87.5 | 12.5 | 50.0 | 9.4 | |
| FaLH03Δesyn1_6.1 | 71.9 | 25.0 | 62.5 | 3.1 | |
| SE * | 7.16 | 9.66 | 9.54 | 3.89 | |
| Control | 75.0 | 79.2 | 100 | 75 | |
| Rating | Lesion Description | Root Discolored (%) | Root Mass Reduction |
|---|---|---|---|
| 1 | 0 | 0 | 0 |
| 2 | Small (0.1–0.2 cm) reddish brown discoloration at point of seed attachment | 0 | 0 |
| 3 | Localized tap root/epicotyl lesions (0.5 to 1 cm) coalescing around ½ of tap root | 10–20% | 0 |
| 4 | Lesions encircle tap root/epicotyl, 1–2 cm long | 95% | 5–10% |
| 5 | Tap root lesion 2–3 cm long, encircle root | 100% | 20–50% |
| 6 | Lesions > 3 cm long, root girdled | 100% | 50–80% |
| 7 | Total decay of tap root/epicotyl | Dead | Dead |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eranthodi, A.; Schneiderman, D.; Harris, L.J.; Witte, T.E.; Sproule, A.; Hermans, A.; Overy, D.P.; Chatterton, S.; Liu, J.; Li, T.; et al. Enniatin Production Influences Fusarium avenaceum Virulence on Potato Tubers, but not on Durum Wheat or Peas. Pathogens 2020, 9, 75. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9020075
Eranthodi A, Schneiderman D, Harris LJ, Witte TE, Sproule A, Hermans A, Overy DP, Chatterton S, Liu J, Li T, et al. Enniatin Production Influences Fusarium avenaceum Virulence on Potato Tubers, but not on Durum Wheat or Peas. Pathogens. 2020; 9(2):75. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9020075
Chicago/Turabian StyleEranthodi, Anas, Danielle Schneiderman, Linda J. Harris, Thomas E. Witte, Amanda Sproule, Anne Hermans, David P. Overy, Syama Chatterton, Jiajun Liu, Tao Li, and et al. 2020. "Enniatin Production Influences Fusarium avenaceum Virulence on Potato Tubers, but not on Durum Wheat or Peas" Pathogens 9, no. 2: 75. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9020075