Evaluation of Biocontrol Activities of Streptomyces spp. against Rice Blast Disease Fungi

1
Division of Biotechnology, Faculty of Science, Maejo University, Chiang Mai 50290, Thailand
2
Department of Biology and Biotechnology, Faculty of Science and Technology, Nakhon Sawan Rajabhat University, Nakhon Sawan 60000, Thailand
3
Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai 50200, Thailand
4
Center of Excellence in Microbial Diversity and Sustainable Utilization, Faculty of Science, Chiang Mai University, Chiang Mai 50200, Thailand
*
Author to whom correspondence should be addressed.
Pathogens 2020, 9(2), 126; https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9020126
Submission received: 15 December 2019
/
Revised: 11 February 2020
/
Accepted: 12 February 2020
/
Published: 15 February 2020
(This article belongs to the Special Issue Streptomyces Microbiomes in Agriculture)
Abstract
:Rhizosphere bacteria can positively influence plant growth by direct and indirect mechanisms. A total of 112 bacterial strains were isolated from the rhizosphere of rice and tested for plant beneficial activities such as siderophore production, cell-wall-degrading enzyme production, hydrogen cyanide (HCN) production and antifungal activity against rice blast disease fungus. The actinomycetes count was 3.8 × 106 CFU/g soil. Streptomyces strains PC 12, D 4.1, D 4.3 and W1 showed strong growth inhibition of blast disease fungus, Pyricularia sp. (87.3%, 82.2%, 80.0% and 80.5%) in vitro. Greenhouse experiments revealed that rice plants treated with Streptomyces strain PC 12 recorded maximum plant height, root length and root dry weight compared to the control. Taxonomic characterization of this strain on the basis of 16S rRNA gene sequence led to its identification as Streptomyces palmae PC 12. Streptomyces palmae PC 12 may be used as biofertilizer to enhance the growth and productivity of commercially important rice cultivar RD6 and the biocontrol of blast disease fungus.
1. Introduction
Rice (Oryza sativa) is an important food crop and it is the staple diet of people around the world, especially in Asia. However, rice is susceptible to diseases by phytopathogenic fungi that cause crop yield losses. Some of them also produce toxic compounds which are harmful upon consumption [1,2]. The rice blast fungus Pyricularia sp. is one of the most devastating airborne pathogens [3]. The fungal spores attach to the host surface by mucilage secreted from the spore tip. The germinated spores produce an extracellular matrix (ECM), and firmly attach to the plant surface [4]. The attached spores develop a germination tube, an aspersorium and a penetration peg, and complete infection by signal exchange with the host plant via ECM [4]. The ECM of Pyricularia sp. helps the fungal spore adhesion to the cellulose membrane and suppressed disease occurrence in the plant cell [5,6]. The fungus can infect rice plants at any growth stage and infects the aerial parts of rice including leaves, nodes, stems and panicles [2,3,7]. The mycelium may survive within the tissues of embryo, endosperm and glumes. Rice blast symptoms include leaf blast, node blast, collar rot, neck rot and panicle blast, which manifests as grayish/brownish spots or lesions as well as the withering of leaves [2,3,8]. The fungus infects the roots of the rice plant and spreads to the aerial tissues, causing rice blast diseases [9]. Yield loss due to rice blast ranges from approximately 10–30% in various rice-producing countries and can increase by up to 50% during disease outbreak [2,8,10]. Many fungicides have been used against blast disease, i.e., azoxystrabin, benomyl, carbendazim, carpropamid, dithiocarbamate, edifenphose, fenoxanil, tiadinil, tricyclazole, pyroquilon, probenazole, iprobenfos, isoprothiolane, metominostrobin and propiconazole [3,11,12]. However, the synthetic fungicides cause environmental pollution, residual problems, the development of pesticide resistance, soil quality reduction and damage to natural ecosystems [11,12]. The control of blast disease using fungicide adversely affects both the environment and soil microbiota [13]. Additionally, human exposure to pesticides may be harmful to health, causing skin irritation, headache, eye irritation and shortness of breath due to the exposure to various pesticides during mixing and spraying [14], and biological processes such as the reproductive system and hormone levels [15].
Current strategies used to control rice blast disease include the use of disease-resistant rice varieties and synthetic fungicides. However, Pyricularia sp. develops new strains rapidly, resulting in a breakdown in rice resistance [12,13]. Nowadays, natural products which are safe for the environment and have low toxicity to living organisms are gaining interest as important sources for the development of fungicides. Streptomyces are natural soil-dwelling bacteria that have been largely used as biological control agents [16]. Streptomyces species have excellent capacities to produce a variety of bioactive compounds, such as antibacterial, antifungal, antiviral, anticancer and antioxidant properties [16]. Some antibiotics produced by Streptomyces have been used as fungicides for the control of rice blast, i.e., Blasticidin-S and Kasugamycin [12,16,17]. For example, Kasugamycin was safely used to protect rice plants against blast disease with low mammalian toxicity and a lack of phytotoxicity toward rice plants and most crops [17]. Oligomycin A, Rapamycin and Pyrroles (Pyrroles [1,2-a] pyrazine-1,4-dione, hexahydro) were found to control the development of rice blast [18]. Streptomyces spp. can be an alternative to synthetic chemical fungicides and used as a biocontrol agent. This biological approach to plant disease management offers a better alternative to control blast disease due to its safety for human use and the environment.
The use of Streptomyces as biocontrol agents of rice blast disease is still limited, especially in Thailand. Boukaew and Prasertsan reported the isolation of S. philanthi RM1-138 from rhizosphere soil of chili pepper in southern Thailand [19]. This Streptomyces showed strong in vitro antifungal activity (88.73% inhibition) on the mycelial growth of Magnaporthe oryzae (P. oryzae PTRRC-18). Other studies also indicated the potential of Streptomyces species to control the disease [12,16,20,21,22,23]. The present study aimed to isolate antagonistic Streptomyces from rice rhizosphere soil and investigate their plant growth promotion and antifungal activity against rice blast fungus Pyricularia sp. in vitro and under greenhouse conditions.
2. Results
2.1. Isolation of Rhizospheric Actinomycetes
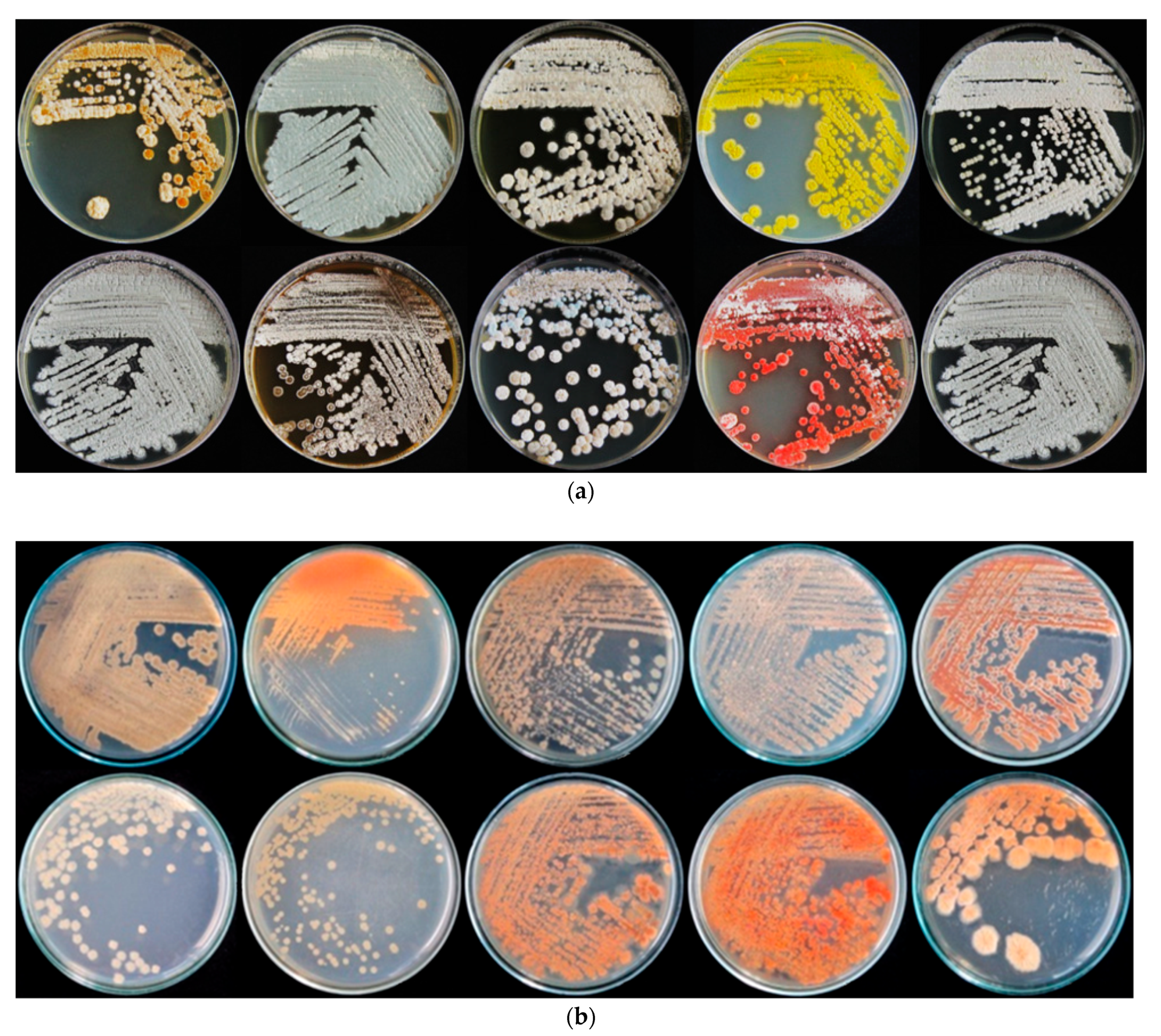
A total of 112 actinomycetes were isolated from the rhizospheric soil of rice in Maerim district, Chiang Mai province, Thailand (18° 56′ 17.2104” N and 98° 53′ 1.7520” E). Isolated actinomycetes showed well-developed substrate mycelium, were filamentous branched, and most aerial mycelium appeared floccose, granular or powdery. Colonies with characteristic features, such as a powdery appearance and color ranging from white or gray to pinkish and yellowish, were selected. The isolates obtained formed colored tough, leathery and filamentous colonies that were hard to pick from the culture media, as a characteristic of genus Streptomyces, and produced colored pigments. Preliminary assignment of Streptomyces according to color of aerial mycelium and presence of soluble pigments is shown in Table 1. Ten main classes of color were observed with grey, brown, white-brown and white as the main colors (Table 1 and Figure 1). Cell wall composition analysis of actinomycetes using thin-layer chromatography (TLC) revealed a type I cell wall with LL-DAP isomers.
2.2. Isolation of Blast Disease Fungi from Rice Leaves and Pathogenicity Test
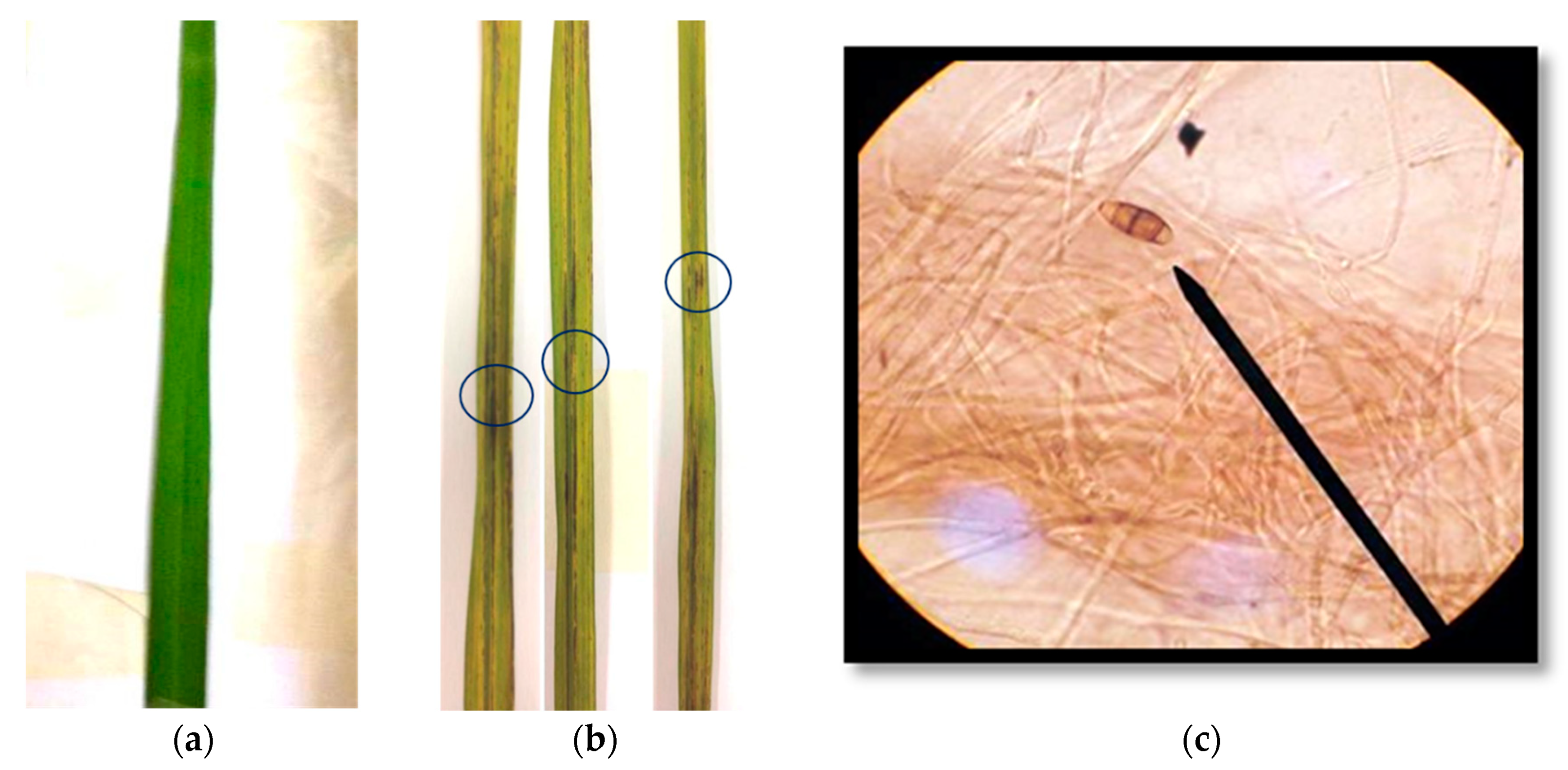
Fifteen isolates of Pyricularia sp. were isolated from the diseased leaves of rice (Oryza sativa L.) cultivar RD 6 and cultured on a potato dextrose agar (PDA) medium at 28 °C for 7 days for 10 days. Morphological characterization on the PDA plate showed that all fungal isolates developed a white, light gray or dark gray mycelium that formed concentric rings on the growth medium. Microscopic observation showed pear-shaped conidiospore with a tail-like structure at the posterior end. The conidia consisted of 2–3 septations with a hyaline brownish color (Figure 2c). Pure culture of isolated Pyricularia strains were inoculated onto rice (O. sativa L.) cultivar RD 6. Pyricularia strain WPP09 reduced the plant height and weight of RD 6 and induced blast disease symptom compared to control plants. Fourteen days after inoculation, 80% diseased plants were recorded (Figure 2b) in Pyricularia inoculated plants, whereas disease was not developed in un-inoculated plants (Figure 2a).
2.3. Plant Growth Properties of the Actinomycete Isolates
In the present study, some isolated strains were positive for plant growth promotion and biocontrol activities. Only strains with a significant antifungal activity in vitro against Pyricularia sp. were shown in Table 2. Streptomyces strain PC 12, Streptomyces strain D 4.1, Streptomyces strain D 4.3 and Streptomyces strain W1 showed maximum growth inhibition of Pyricularia sp. (87.3%, 80.0%, 82.2% and 80.5%) in a dual-culture plate. Based on percent inhibition of the pathogen, four actinomycetes strains, namely W1, PC 12, D 4.1 and D 4.3, were selected for greenhouse experiment (Table 2). All strains were positive for HCN production, had strong siderophores production and produced cell-wall-degrading enzymes, thus indicating their potential for antifungal activities (Table 2). Siderophore production was screened by observing the change in color from blue to orange in CAS agar plates. Twenty-seven isolated strains were siderophore positive with varying intensities of orange zones. Strain D 4.1 (52.3 mm), strain D 4.3 (51.4 mm), strain PC-117 (50.7 mm) and strain PC-94 (50.2 mm) exhibited the maximum siderophore producer on the CAS agar plate (Table 2). All isolates were positive for protease activity with a zone diameter ranging from 24.2 mm to 73.7 mm. Nine isolates were positive for cellulase, with strain D 4.1 as the best producer (70.8 mm), followed by strain D 4.3 (69.5 mm) and strain PC-55 (64.2 mm). Chitinase activity was positive for seven strains. Maximum chitinase activity was observed in strain D 4.1 (57.4 mm), followed by strain D 4.3 (48.5 mm) (Table 2).
2.4. Greenhouse Experiment
2.4.1. Pathogenicity Test of Selected Streptomyces
We evaluated the pathogenicity of four selected Streptomyces strains, W 1, PC 12, D 4.1 and D 4.3, on rice RD 6. No sign of abnormalities, such as lesion formulation or wilting, were observed in seedlings at 14 days.
2.4.2. Evaluation of Streptomyces against Pyricularia sp. under Greenhouse Condition
Four Streptomyces (strain W 1, strain PC 12, strain D 4.1 and strain D 4.3) were selected and screened under greenhouse conditions. The initial disease symptoms appeared five days post-inoculation (dpi). Typical symptoms of blast on the rice leaves were diamond-shaped lesion with a grey or white center and brown border (Figure 2). Symptoms of blast increased daily from 4 dpi (day post inoculation) and reached their peak on 60 dpi in the inoculated rice plant. The progression of disease in terms of disease severity varied among the treatments (Table 3). Disease severity in pathogen-inoculated control plants was 87.5% at 60 dpi, while in pathogen + Streptomyces, it ranged from 51.9% to 31.4%. PC 12-inoculated plants showed the lowest disease severity (31.4%). No disease development was observed in the untreated control. Compared to pathogen-inoculated plants, Pathogen + Streptomyces PC 12-inoculated plants had a significantly lower disease severity (35.8%) compared to the control (Table 3). In addition, treatments with Streptomyces PC 12 and Streptomyces D 4.3 showed slower blast disease progression than the other treatments (Table 3).
2.4.3. Plant Growth Promotion effect
Lesion length and number of dried leaves
Plants treated with Streptomyces strain PC 12 showed the lowest number of leaf lesions and dried leaves compared to the other tested isolates. All isolates significantly reduced the disease compared to the control. Strain PC 12 showed the lowest mean lesion length (0.55 cm) and number of dried leaves (3.10 leaves/plant) (Table 3). These low disease parameters indicate the healthy status of PC 12-inoculated plants.
Plant height
All tested strains enhanced the plant height compared to control (Table 3). At 60 days, the highest plant height (76.93 cm) was recorded in the treatment of Streptomyces strain PC 12. This was followed by the treatment of Streptomyces strain D 4.3 (52.69 cm). The control plant treated with pathogen showed the lowest height. In addition, the height of rice in PC 12-inoculated plants was almost double that of un-inoculated plants, which indicated that rice was in a healthy condition.
Root length and root dry weight
In general, the root length of plants from all treatments with selected Streptomyces was longer than the control (Table 3). The longest root length (41.60 cm) was recorded in rice inoculated with Streptomyces strain PC 12. Similarly, the dry weight of root was enhanced in all treatments with selected Streptomyces. Again, Streptomyces strain PC 12 yielded the highest root dry weight of 2.68 g. Plants treated with the fungal pathogen showed the lowest dry weight (1.88 g) (Table 3). These observations suggested the healthy status of PC 12-inoculated plants compared to the pathogen-treated control.
Number of tiller
The average number of tiller was significantly enhanced by all tested Streptomyces PC 12 in pathogen + Streptomyces-inoculated plants. Streptomyces PC 12 -inoculated plants yielded the highest number of tiller (19.4) over pathogen-inoculated control and untreated control plants, suggesting that the rice was healthy.
2.5. Rhizospheric Colonization by Streptomyces
At the beginning of the experiment, the actinomycetes isolates had an average cell number of 3.0 × 106 CFU/mL. After the selected Streptomyces strains were applied to the plants for 2 months, the amount of Streptomyces strain PC 12 decreased to 4.5 × 104 CFU/mL (Table 4). Other Streptomyces strains also showed a decreasing trend. The persistence of the strains in the rhizosphere may account in part for the varied level of disease suppression between different Streptomyces sp. treatments.
2.6. Identification of Streptomyces PC 12
Strain PC 12 was observed to grow on a variety of ISP agar, including yeast-extract-malt-extract agar (ISP-2), oatmeal agar (ISP-3), inorganic salts/starch agar (ISP-4) and peptone/yeast agar (ISP-6). Aerial mycelium color was white and substrate mycelium was grey on ISP-2 media (Figure 3a,b). Melanin pigments were not observed on any of the media tested. Spiral spore chains were observed under light microscope (Figure 3d). Spore chain and spore surface morphology of strain PC 12 was determined by scanning electron microscope (SEM). Spores were spherical with a spiny spore surface (Figure 4). TLC analysis of whole-cell hydrolysates of strain PC 12 showed LL-diaminopimelic acid (LL-DAP) (data not shown).
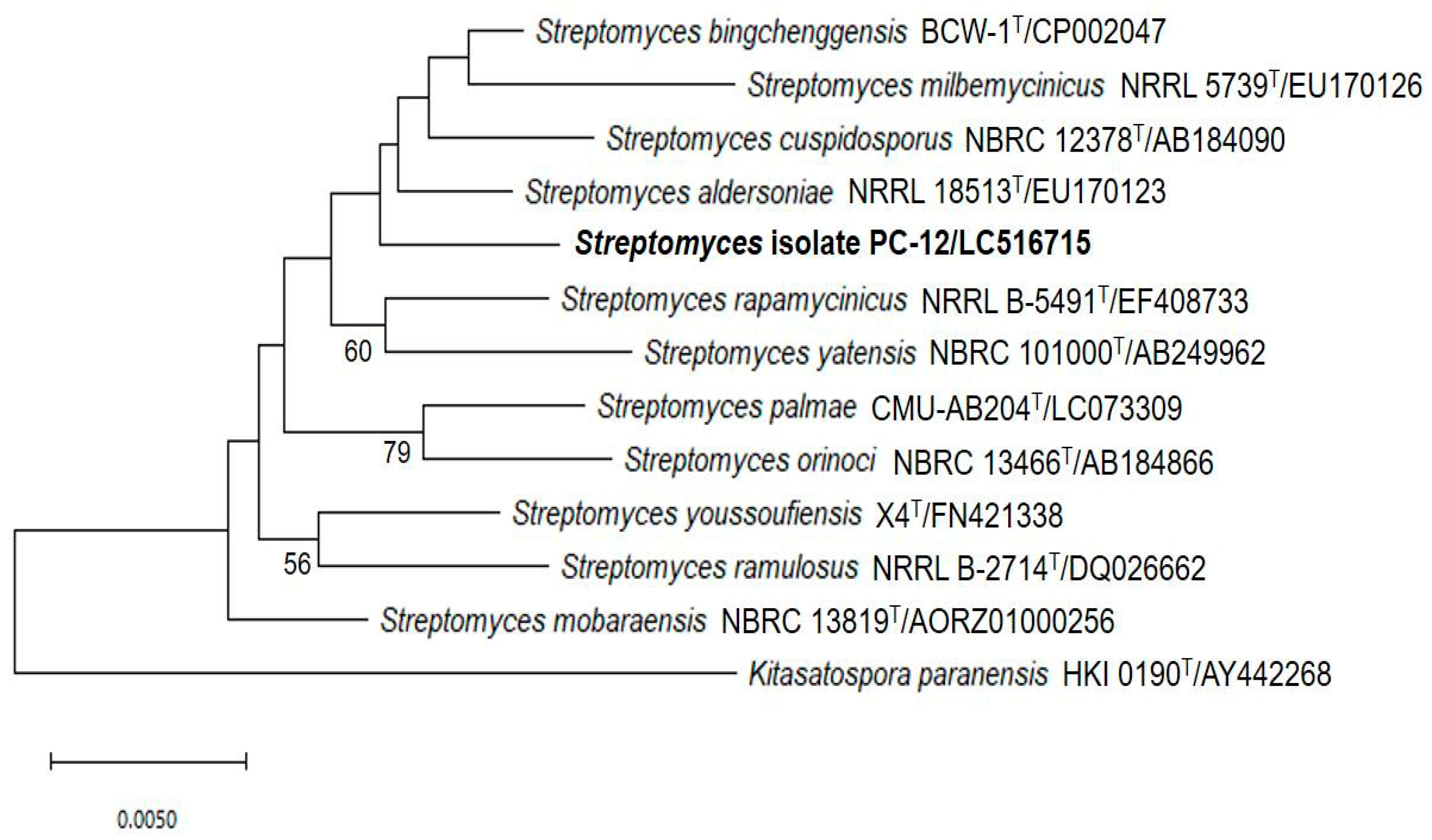
Based on chemical, cultural and morphological characteristics, strain PC 12 belongs to the genus Streptomyces according to Nonomura’s key [24]. The 16S rRNA gene of strain PC 12 was sequenced and compared with related Streptomyces species deposited in the GenBank database which indicated that, phylogenetically, strain PC 12 belonged to genus Streptomyces. BLAST analysis showed that strain PC 12 was closely related to Streptomyces palmae CMU-AB204T, with 98.82% similarity. Phylogenic analysis based on 16S rRNA gene sequences using the neighbor-joining methods was shown in Figure 5. Strain PC 12 formed an independent clade separated from S. palmae T CMU-AB204T.
3. Discussion
The present study was designed to isolate strains of actinomycetes from rice rhizosphere in an attempt to investigate their potential to control rice blast fungus Pyricularia sp in vitro and under greenhouse conditions. In this study, 112 isolates were recovered. Morphologically, all isolates were assigned to the genus Streptomyces [24]. Actinomycetes are commonly found in rhizosphere soil, especially members of the genus Streptomyces [16]. These microorganisms are well documented as potential candidates to inhibit the growth of several fungal plant pathogens including Pyricularia spp. [12,16]. Rice blast caused by Pyricularia oryzae is the most serious disease in all rice-growing regions worldwide [2,3]. In the present work, fifteen fungal strains were isolated from disease leaves of rice RD 6 and showed a typical morphology of Pyricularia species according to Mew and Gonzales (2002) [25] and Bussaban et al. [26]. The conidial shape can be used to differentiate Pyricularia from closely related genera and the spore morphology is consistently correlated with phylogenetic analysis [26].
Controlling rice blast disease has become a major concern, as it impacts rice productivity worldwide [2]. In this study, Streptomyces PC 12 was found to be capable of suppressing rice blast disease and promoting plant growth. When tested in vitro, strain PC 12 greatly inhibited Pyricularia WPP009 mycelial growth (87.3%), suggesting its capability of inhibiting the rice blast pathogen (Table 2). Our results are supported by with other works which reported the potential of Streptomyces strains as biocontrol agents against Pyricularia spp. under laboratory conditions. S. vinaceusdrappus was isolated from sediment of Loktak lake in India [20]. It showed 53.5% growth inhibition against P. oryzae MTCC1477. Simialrly, S. philanthi RM-1-138 isolated from chili pepper rhizosphere soil of southern Thailand could inhibit growth of P. oryzae PTRRC-18 in vitro [19].
Some studies also reported the evaluation of Streptomyces as a biocontrol agent for Pyricularia spp. under greenhouse or field experiments. For example, Streptomyces UPMR54 was reported to reduce rice blast disease by 67.9% and promoted rice growth and yield [21]. Antifungal compound, SPM5C-1 from Streptomyces strain PM5 completely inhibited mycelial growth of P. oryzae in vitro at concentrations of 25 μg/ml [22]. In addition, blast disease was reduced by 76.1% in greenhouse experiment at 500 μg/ml of SPM5C-1. Recently, foliar treatment of Streptomyces hygroscopicus OsiSh-2 culture filtrate showed 23.5% and 28.3% disease reduction of P. oryzae in rice seedlings under greenhouse and field trials [23]. In the present study, it was evident that Streptomyces strain PC 12 efficiently reduced blast severity from 87.5% to 31.4% in pathogen-infected plants under greenhouse conditions (Table 3).
The ability of potential biocontrol agents to promote plant growth is considered as an added advantage. Streptomyces PC 12 inoculation was found to enhance rice growth, as indicated by plant height, root length, root dry weight and number of tiller (Table 3). It is known that Streptomyces can enhance plant growth while suppressing disease using different mechanisms. Streptomyces sp. promotes plant growth by the production of plant growth hormones and siderophores, nitrogen fixation, and mineral solubilization, especially phosphates [16]. Streptomyces, with both antagonistic activity against pathogens and growth promoting ability, are attractive for development as a biocontrol agent to replace chemical fertilizers and pesticides in agriculture [21,27,28].
Biocontrol bacteria inhibit plant pathogens via several mechanisms, such as the production of antimicrobial compounds (antibiosis), iron sequestration (siderophores), production of extracellular enzymes that interfere with cell wall synthesis (chitinases, cellulase, proteases) and induction of plant-resistance mechanisms [12,16,23]. Rhizosphere Streptomyces in this study showed antagonistic activity against the rice blast fungus, Pyricularia sp., which correlated with their ability to produce siderophores, hydrolytic enzymes and antibiotics (Table 2). All actinomycete isolates were able to grow on CAS agar in the present study, which indicated their ability to produce siderophores. Siderophore-producing bacteria suppress some soil-borne fungal pathogens through iron competition [1,27]. All tested actinomycetes were also found to produce HCN. The production of HCN in excess may play a critical role in the control of fungal disease [29]. Microbial production of HCN has been suggested as an important antifungal activity to control root pathogens. Cyanide acts as a general metabolic inhibitor to avoid predation or competition [28]. The rice seedlings are not harmfully affected by inoculation with HCN-producing strains and the selected Streptomyces can act as a biological control agent. It is clear from the present investigation that rhizosphere Streptomyces sp. are able to produce antifungal substances as all isolates showed antagonistic activity against Pyricularia sp., though the active compound has yet to be purified and characterized (Table 2). Different levels of disease suppression in rice seedlings might be due to the different colonization pattern and secretion of secondary metabolites and/or cell-wall-degrading enzymes by the antagonistic bacteria. In the present study, all selected Streptomyces strains were found to colonize well in the rhizosphere of rice (Table 4). Streptomyces strain PC 12 was found to persist better than the other strains, and this might be the reason for the better performance in controlling blast disease.
The cultural and morphological characteristics assigned the selected strain under the genus Streptomyces. Chemotaxonomic characteristics indicated that they belonged to genus Streptomyces, as the cell wall contained LL-diaminopimelic acid (cell wall type-I). BLAST analysis indicated that strain PC12 was closely related to Streptomyces palmae CMU-AB204T with 98.82% similarity. It is recommended that 98.65% 16S rRNA gene sequence similarity can be used to differentiate between closely related species [30]. Therefore, in this study, the potent Streptomyces strain PC 12 was identified by 16S rRNA gene sequence analysis as Streptomyces palmae PC 12. However, from the position of strain PC12 in the phylogenetic tree, it formed a separate branch to its closest neighbor, S. palmae CMU-AB204T (Figure 5). It is likely that strain PC12 may represent a novel Streptomyces species. However, a detailed polyphasic taxonomic characterization including whole genome sequence is required to confirm its status, which is clearly not the objective of the current study.
The results from this in vitro and greenhouse study suggest that the actinomycete isolates have potential to be used as biocontrol agents for the inhibition of rice blast disease by Pyricularia spp.
4. Materials and Methods
4.1. Isolation of Rhizospheric Actinomycetes
The isolation of actinomycetes was performed by conventional serial dilution spread-plate technique. The suspension from an appropriate dilution was inoculated on Actinomycetes Isolation Agar (AIA, Difco) and incubated at 30 °C for 7 days. The AIA medium was supplemented with 40 μg/mL of cyclohexamide to inhibit fungal growth and 10 μg/ml of nalidixic acid to inhibit other bacterial growth without affecting the actinomycetes [31]. Pure cultures were obtained by re-streaking on AIA medium and were stored as spore suspension in 20% glycerol for long-term preservation at -80 °C.
4.2. Isolation of Rice Blast Disease Fungi
Five infected rice plants (RD 6) were collected from the rice field in Maerim district, Chiang Mai province, Thailand (18° 56′ 17.2104” N and 98° 53′ 1.7520” E). Leaves with disease symptoms were cleaned under running tap water and cut into 1 × 1 cm segments. Surface sterilization was done by washing with 70% ethanol for 5 min followed by five rinses in sterile distilled water. Leaves were placed on potato dextrose agar (PDA) and final rinsing water was spread onto PDA medium to check the effectiveness of surface sterilization. The absence of microbial growth on the PDA medium confirmed that the surface sterilization procedure was effective in removing the surface bacteria [32]. Inoculated plates were incubated at 30 °C for 5 days. The fungal colony growing out from plant tissues was transferred onto fresh PDA medium. Pure cultures were observed under light microscope and were examined macro- and microscopically to identify the fungus as Pyricularia spp [25,26].
4.3. Pathogenicity Assay
Pyricularia sp. isolate was incubated on PDA medium and incubated at 28 °C for 5 days. The 14 day old plantlets at two-leaf growth stage were inoculated with 5 mm fungal plugs of Pyricularia sp. culture. The plugs were dropped on four leaves and wrapped with cotton moistened with distilled water. Humidity and moisture were maintained through the use of aluminum foil. The disease symptoms appeared 72 hours post-inoculation (hpi) and leaves were harvested [33].
4.4. In vitro Antifungal Assay
All the isolated actinomycetes were tested in an antagonistic assay conducted using the dual-culture technique. A 5 mm agar plug of Pyricularia sp. was placed on a side of the plate and another 5 mm agar plug with an actinomycete was placed on the opposite side [34]. Plates were incubated at 28 °C for 5 days and the antagonistic activity was scored according to the scale developed by Alfredo and Aleli (2011) [35]. The diameter of the mycelium growing out from the plug was measured and reported as the percentage inhibition of radial growth (PIRG) using the following formula [36]
where R 1= Radial growth of Pyricularia sp. (mm) in the control plates
PIRG = R 1-R 2/R 1 × 100
R 2 = Radial growth of P. oryzae interacting with antagonistic bacteria (mm)
Three biological replicates were performed and an average was taken.
4.5. Actinomycetes Identification
4.5.1. Cultural and Morphological Characterizations
Morphological characteristics of the selected strain, including spore size and surface ornamentation, were assessed by compound microscope and scanning electron microscope (SEM) of 10 day old cultures grown on ISP-2 agar. The color of aerial spore mass, substrate mycelium and diffusible pigments of the selected strain were recorded after incubation at 30℃ for 10 days on ISP (International Streptomyces Project) medium (Difco) [37]. All characteristics were compared with Nonomura’s key [24].
4.5.2. Cell Wall Composition Analysis
In order to determine the genus of antagonistic actinomycetes, the 2,6- diaminopimelic acid, one of the cell wall components of actinomycetes mycelia was analyzed using the method of Hasegawa et al. (1983) [38]. The selected strain was cultured in ISP-2 medium (4.0 g yeast extract, 10.0 g malt extract, 4.0 g dextrose, and 1 L sterile distilled water, pH 7.3) at 150 rpm, 28℃ for 14 days. After cultivation, the culture broth was centrifuged to collect cells. 6N HCl was used to hydrolyze the cells by heating at 70ºC for 18 h in a water bath. The hydrolysate was filtered through Whatman No.1 filter paper and evaporated to dryness in order to remove the HCl residue. Dried hydrolysate was dissolved in 1 mL of distilled water and applied onto TLC plate (15 x 20 cm, Merck Co., USA). A total of 20 ul of 0.01 M DL-DAP (Sigma Chemical Co., USA) containing both meso- and LL-DAP isomers and amino acids (alanine, glycine and glutamate) was also loaded on the TLC plates as a standard.
4.5.3. Molecular Characterization of Potential Antagonistic Isolate
The sequence of 16S rDNA of potential strain was determined after genomic DNA extraction and PCR amplification using eubacterial 16S rRNA gene-specific primers 27F (5’-AGT TTG ATC CTG GCT CAG GAC GAA CG-3’) and 1525R (5’-AGC CGG TCC CCC TGC AAG-3’) [21]. PCR amplification was carried out in 20 µL reaction mixture and amplification cycles, as described previously [27]. Amplified DNA was purified using a polymerase chain reaction (PCR) purification kit (Promega, Madison, USA). The sequencing was performed by commercial service of Pacific Science Co., Ltd., Thailand. A comparison of the obtained sequence with related Streptomyces strains in EzBiocloud database was performed using the Basic Local Alignment Search Tool (BLAST). The obtained 16S rRNA gene sequence was aligned with nucleotide sequences of related Streptomyces species, and the phylogenic tree was constructed by the neighbor-joining method using a MEGA X software package [39]. The resultant tree topology was evaluated by bootstrap analysis of the neighbor-joining data, based on 1000 resampled datasets [40]. The evolutionary distances were computed using the Maximum Composite Likelihood method [41] and are in the units of the number of base substitutions per site. This analysis involved 13 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. All ambiguous positions were removed for each sequence pair (pairwise deletion option). There were a total of 1339 positions in the final dataset.
4.5.4. Scanning Electron Microscopy
Spore surface ornamentation was observed by scanning electron microscopy (SEM). Mycelia were taken after 10 days culture and washed in 0.1 M sodium cacodylate buffer (pH 7.4). They were fixed in 2.5% gluraraldehyde in 0.1 M sodium cacodylate buffer for 4 h at 4 °C, followed by post-fixation with 1% osmium tetraoxide in 0.1 M sodium cacodylate buffer (pH 7.4) and dried in a critical point dryer (EMITECH model K850, Hitachi). The specimens were mounted onto aluminium holders, sputter-coated with 10 nm Au and observed by SEM (Hitachi model S3400 at 15-30 kv, 2-5.00 μM).
4.6. Screening of Plant Growth Promoting Activities
4.6.1. Siderophore Production
Siderophore production was assayed on Chrome Azurol S blue agar (CAS) [42] and overnight bacteria culture were spotted on CAS plates and incubated at 30 °C for 5 days in the dark. The cultures showing a yellow to orange colored zone around colonies indicated siderophore production.
4.6.2. HCN Production
Actinomyctes were streaked into ISP-2 agar supplemented with glycine. The plates were inverted and a piece of filter paper impregnated with 0.5% picric acid and 2% sodium carbonate was placed on the lid. After incubation for a week at 30 °C, the color change of the filter paper from yellow to orange was an indicator for HCN production [28].
4.6.3. Hydrolytic Enzyme Production
Cellulase Production
Tested strains were grown on CMC agar for 5 days, flooded with 1% Congo red solution and washed with distilled water. The clear zone around the colony was observed and measured [28].
Chitinases Assay
Screening for chitinase production was done by agar plate assay on colloidal chitin medium containing 1.5% colloidal chitin, yeast extract 0.5 g, (NH4)2SO4 1 g, MgSO4 6H2O 0.3 g, KH2PO4 1.36 g, agar 15 g and distilled water 1000 mL. The plates were incubated for 5 days at 30 °C and a clear zone around the colony indicated chitinase activity [28].
Proteases Assay
Protease activity was screened on skim milk agar containing (per liter): 5 g pancreatic digest of casein, 2.5 g yeast extract, 1 g glucose, 7% skim milk solution and 15 g of agar (pH 7.0). Streptomyces plug (6 mm) were placed on the medium and incubated at 30 °C for 5 days. Proteolytic activity was identified by the clear zone around the cell [28].
4.7. Greenhouse Experiments
The effect of the antagonistic Streptomyces sp. on controlling rice blast in vivo was assayed in greenhouse conditions. Rice (Oryza sativa L.) cultivar RD6 was used as a host plant as it is particularly susceptible to rice blast disease. The spore suspension of antagonistic actinobacterium (106 CFU/mL) and the conidial suspension of Pyricularia sp. (105 spores/mL) were prepared the same as for the in vitro assay.
In greenhouse conditions, surface-sterilized rice seeds (RD 6) were sown in pots containing autoclaved soil (soil: sand; 3:1). The spore suspension of the antagonistic actinobacterium containing 0.2% (v/v) Tween 20 was sprayed on 10 day old rice leaves using a spray bottle (50 mL spore suspension per 100 seedlings). The rice leaves were sprayed with water containing 0.2% (v/v) Tween 20 as a control. The conidial suspension of Pyricularia sp. was sprayed 7 days later in the same way as described for the antagonistic actinobacterium. High humidity was maintained by the constant spray of moisture and the plants were kept under polythene shading. The experiment was carried out in pot culture with eleven treatments in four replications, following a completely randomized design (CRD). The treatments were prepared as shown below:
- T 1 = Healthy control (no fungus, no actinomycetes);
- T 2 = Healthy rice plant inoculate with Streptomyces strain W 1;
- T 3 = Rice plant inoculate with Streptomyces strain W 1+ Pyricularia sp.;
- T 4 = Healthy rice plant inoculate with Streptomyces strain D 4.3;
- T 5 = Rice plant inoculate with Streptomyces strain D 4.3 + Pyricularia sp.;
- T 6 = Healthy rice plant inoculate with Streptomyces strain PC 12;
- T 7 = Rice plant inoculate with Streptomyces strain PC 12 + Pyricularia sp.;
- T 8 = Healthy rice plant inoculate with Streptomyces strain D 4.1;
- T 9 = Rice plant inoculate with Streptomyces strain D 4.1+ Pyricularia sp.;
- T 10 = Disease control (Pyricularia sp. alone);
- T 11 = Rice plant inoculate with Fungicide + Pyricularia sp.
The disease severity was evaluated 50 days after inoculating with Pyricularia sp. according to the standard evaluation system for rice [43]. The disease level was scored on a scale from 0 to 9 as follows:
- 0. no lesions observed;
- 1. small brown specks of pin-point size without sporulating centre;
- 2. larger brown specks but less than 1 mm in diameter;
- 3. small roundish to slightly elongated, necrotic grey spots 1– 2 mm;
- 4. typical susceptible blast lesions (spindle-shaped) 3 mm or longer, infecting less than 4.0% of the leaf area;
- 5. typical blast lesions infecting 4.1–10.0% of the leaf area;
- 6. typical blast lesions infecting 10.1–25.0% of the leaf area;
- 7. typical blast lesions infecting 25.1–50.0% of the leaf area;
- 8. typical blast lesions infecting 50.1–75.0% of the leaf area;
- 9. typical blast lesions infecting more than 75.1% of the leaf area.
The disease severity was computed using the following equation
The experiment was repeated twice with three replicates per treatment and 100 plants per replicate.
4.8. Effect of Streptomyces Application on Plant Growth Promotion under Greenhouse Conditions
4.8.1. Measurement of Shoot and Root Weight and Tiller Number
The plants were uprooted carefully from the pot 60 days after transplantation. Tiller numbers were obtained. The root region was cut, separated from the plants and washed thoroughly to remove adhered soil particles. The fresh shoot and fresh root were dried by hot air oven at 50℃ for 48 h. The dry shoot weight and dry root weight of the plants of each treatment were measured in grams.
4.8.2. Rhizospheric Colonization by Streptomyces
Root apex samples from rice plants inoculated with each tested actinomycetes strains were taken 40 days after inoculation to check for rhizospheric colonization of bacteria. Root extracts and serial dilutions of 10-3-10-5 were made for the selection of bacteria. For both types of samples, 0.1 ml of each dilution was placed on ISP-2 medium and incubated at 30ºC for 72 h or until colony development was observed.
4.9. Statistical Analysis
The data were subjected to analysis of variance using SPSS 14.0 software. Mean values among treatments were compared by the least-significant difference (LSD) test at 5% level (P ≤ 0.05) of significance and presented as the mean values ± standard deviation (SD).
5. Conclusions
It is evident that selected rhizosphere actinomycetes successfully suppressed or reduced disease symptoms under both in vitro and in vivo greenhouse conditions. S. palmae PC 12 significantly reduced the disease severity by 56%. This strain also significantly increased plant growth attributes in infected plants. The results obtained in this study show the potential of Streptomyces palmae PC 12 as a biocontrol agent against rice blast fungus, Pyricularia sp., as well as in promoting the growth of rice variety RD6. The fungal inhibitory effect could be due to the production of cell-wall-degrading enzymes, such as chitinase, and HCN production. However, additional field trials are required to confirm the feasibility of S. palmae PC12 as an inexpensive, safe, and sustainable biopesticide for blast disease management.
Author Contributions
Conceptualization, M.C.; T.T. and W.P.-a; Methodology, M.C and T.T.; Investigation, M.C.; T.T. and W.P.-a; Writing—original draft preparation, M.C.; Writing—revision and editing, M.C.; T.T. and W.P.-a All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Research Council of Thailand (NRCT research number: MJU-1-60-055) and partially supported by Chiang Mai University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chaiharn, M.; Chunhaleuchanon, S.; Lumyong, S. Screening siderophore producing bacteria as potential biological control agent for fungal rice pathogens in Thailand. World J. Microbiol. Biotechnol. 2009, 25, 1919–1928. [Google Scholar] [CrossRef]
- Asibi, A.E.; Chai, Q.; Coulter, J.A. Rice blast: A disease with implications for global food security. Agronomy 2019, 9, 451. [Google Scholar] [CrossRef] [Green Version]
- Kato, H. Rice blast disease. Pestic. Outlook. 2001, 12, 23–25. [Google Scholar] [CrossRef]
- Hamer, J.E.; Howard, R.J.; Chumley, F.G.; Valent, B. A mechanism for surface attachment in spores of a plant pathogenic fungus. Science 1988, 239, 288–290. [Google Scholar] [CrossRef]
- Vidhyasekaran, P. Fungal pathogenesis in plants and crops. In Molecular Biology and Host Defense Mechanisms, 2th ed.; Vidhyasekaran, P., Ed.; CRC Press: New York, NY, USA, 2008; pp. 1–53. [Google Scholar]
- Bae, C.Y.; Kim, S.; Choi, W.; Lee, Y.H. Involvement of extracellular matrix and integrin-like proteins on conidal adhesion and appressorium differentiation in Magnaporthe oryzae. J. Microbiol. Biotechnol. 2007, 17, 1198–1203. [Google Scholar] [PubMed]
- Inoue, K.; Suzuki, T.; Ikeda, K.; Jiang, S.; Hosogi, N.; Hyon, G.S.; Hida, S.; Yamada, T.; Park, P. Extracellular matrix of Magnaporthe oryzae may have a role in host adhesion during fungal penetration and is digested by matrix metalloproteinases. J. Gen. Plant Pathol. 2007, 73, 388–398. [Google Scholar] [CrossRef]
- Wilson, R.A.; Talbot, N.J. Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Sesma, A.; Osbourn, A.E. The rice leaf blast pathogen undergoes developmental processes typical of root-infecting fungi. Nature 2004, 431, 582–586. [Google Scholar] [CrossRef]
- Ashkani, S.; Yusop, M.R.; Shabanimofrad, M.; Haron, A.R.; Sahebi, M.; Latif, M.A. Genetic analysis of resistance to rice blast: A study on the inheritance of resistance to the blast disease pathogen in an F3 population of rice. J. Phytopathol. 2015, 163, 300–309. [Google Scholar] [CrossRef]
- Pooja, K.; Katoch, A. Past, present and future of rice blast management. Plant Sci. Today 2014, 1, 165–173. [Google Scholar] [CrossRef]
- Law, J.W.; Ser, H.L.; Khan, T.M.; Chuah, L.H.; Pusparajah, P.; Chan, K.G.; Goh, B.H.; Lee, L.H. The potential of Streptomyces as biocontrol agents against the rice blast fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, M.Y.; Cha, B.; Kim, J.C. Recent trends in studies on botanical fungicides in agriculture. Plant Pathol. J. 2013, 29, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, S.; Meisner, C.; Wheeler, D.; Xuyen, K.; Lam, N.T. Pesticide poisoning of farm workers-implications of blood test results from Vietnam. Int. J. Hyg. Environ. Health 2007, 210, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattahi, F.; Mousavi Moghadam, M.; Khanbabaei, R. The effect of tricyclazole on testosterone changes and testicular structure in mice. J. Babol. Univ. Med. Sci. 2015, 17, 43–49. [Google Scholar] [CrossRef]
- Newitt, J.T.; Prudence, S.M.M.; Hutchings, M.I.; Worsley, S.F. Biocontrol of cereal crop diseases using streptomycetes. Pathogens 2019, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef]
- Yang, P.; Li, M.; Zhao, J.; Zhu, M.; Shan, H.; Li, J. Oligomycins A and C, major secondary metabolites isolated from the newly isolated strain Streptomyces diastaticus. Folia Microbiol. 2010, 55, 10–16. [Google Scholar] [CrossRef]
- Boukaew, S.; Prasertsan, P. Suppression of rice sheath blight disease using a heat stable culture filtrate from Streptomyces philanthi RM-1-138. Crop Protect. 2014, 61, 1–10. [Google Scholar] [CrossRef]
- Ningthoujam, S.; Sanasam, S.; Tamreihao, K.; Nimaich, S. Antagonistic activities of local actinomycete isolates against rice fungal pathogens. Afr. J. Microbiol. Res. 2009, 3, 737–742. [Google Scholar]
- Awla, H.K.; Kadir, J.; Othman, R.; Rashid, T.S.; Hamid, S.; Wong, M.-Y. Plant growth-promoting abilities and biocontrol efficacy of Streptomyces sp. UPMRS4 against Pyricularia oryzae. Biol. Control. 2017, 112, 55–63. [Google Scholar] [CrossRef]
- Prabavathy, V.R.; Mathivanan, N.; Muragesan, K. Control of blast and sheath blight disease of rice using antifungal metabolites produced by Streptomyces sp. PM 5. Biol. Control. 2006, 3, 313–319. [Google Scholar] [CrossRef]
- Xu, T.; Cao, L.; Zeng, J.; Franco, C.M.M.; Yang, Y.; Hu, X.; Liu, Y.; Wang, X.; Gao, Y.; Bu, Z.; et al. The antifungal action mode of the rice endophyte Streptomyces hygroscopicus OsiSh-2 as a potential biocontrol agent against the rice blast pathogen. Pestic Biochem. Phys. 2019, 160, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Nonomura, H. Key for classification and identification of 458 species of the Streptomycetes included in ISP. J. Ferment. Technol. 1974, 52, 78–92. [Google Scholar]
- Mew, T.W.; Gonzales, P. A Handbook of Rice Seed Borne Fungi. Los Banos, Laguna: IRRI; Enfield, N.H., Ed.; Science Publishers, Inc.: New York, NY, USA, 2012; pp. 27–31. [Google Scholar]
- Bussaban, B.; Lumyong, S.; Lumyong, P.; Seelanan, T.; Park, D.C.; McKenzie, E.H.C.; Hyde, K.D. Molecular and morphological characterization of Pyricularia and allied genera. Mycologia 2005, 97, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Chaiharn, C.; Sujada, N.; Pathom-aree, W.; Lumyong, S. Biological control of Rigidoporus microporus the casue of white root disease in rubber using PGPRs in vivo. Chiang Mai J. Sci. 2019, 46, 850–866. [Google Scholar]
- Hata, E.M.; Sijam, K.; Ahmad, Z.A.M.; Yusof, M.T.; Azman, N.A. In vitro antimicrobial assay of actinomycetes in rice against Xanthomonas oryzae pv. oryzicola and as potential plant growth promoter. Braz. Arch. Biol. Technol. 2015, 58, 821–832. [Google Scholar] [CrossRef]
- Banchio, E.; Bogino, P.C.; Zygadlo, J.; Giordano, W. Plant growth promoting rhizobacteria improve growth and essential oil yield in Origanum majorana L. Biochem. Syst. Ecol. 2008, 36, 766–771. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Loqman, S.; Barka, E.E.A.; Clement, C.E.; Ouhdouch, E.Y. Antagonistic actinomycetes from Moroccan soil to control the grapevine gray mold. World J. Microbiol. Biotechnol. 2009, 25, 81–91. [Google Scholar] [CrossRef]
- Schulz, B.; Wanke, U.; Draeger, S. Endophytes from herbaceous plants and shrubs: Effectiveness of surface sterilization methods. Mycol. Res. 1993, 97, 1447–1450. [Google Scholar] [CrossRef]
- Plodpai, P.; Chuenchitt, S.; Petcharat, V.; Chakthong, S.; Piyawan, S.; Voravuthikunchai, S.S. Anti-Rhizoctonia solani activity by Desmos chinensis extracts and its mechanism of action. Crop Prot. 2013, 43, 65–71. [Google Scholar] [CrossRef]
- Hamdia, Z.A.; Kalaivani, N. Evaluating efficacy of Trichoderma isolates and Bacillus subtilis as biological control agents against Rhizoctonia solani. Res. J. Appl. Sci. 2013, 8, 72–81. [Google Scholar]
- Alfredo, M.S.; Aleli Cornelia, R.P. Biological control of sheath blight of upland rice with Trichoderma species. J. Trop. Plant Pathol. 2011, 69, 1–9. [Google Scholar]
- Lee, J.M.; Tan, W.S.; Ting, A.S.Y. Revealing the antimicrobial and enzymatic potentials of culturable fungal endophytes from tropical pitcher plants (Nepenthes spp.). Mycosphere 2014, 5, 364–377. [Google Scholar] [CrossRef]
- Shirling, E.B.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, T.; Takizawa, M.; Tanida, S. A rapid analysis for chemical grouping of aerobic actinomycetes. J. Gen. Appl. Microbiol. 1983, 29, 319–322. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogeny: An appropriate use of the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Nat. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- International Rice Research Institute. Standards Evaluation System for Rice; IRRI: Manila, Philippines, 2013. [Google Scholar]
Figure 1.
Characteristic of the aerial (a) and substrate mycelium (b) of some rhizospheric actinomycetes isolated on ISP-2 medium after being incubated at 30 °C for 14 days.
Figure 1.
Characteristic of the aerial (a) and substrate mycelium (b) of some rhizospheric actinomycetes isolated on ISP-2 medium after being incubated at 30 °C for 14 days.

Figure 2.
Typical blast symptom on rice (Oryza sativa L.) cultivar RD6 leaves 14 days after inoculation (a) Healthy leaf, (b) infected sick leaves and (c) Pyricularia sp. WPP09 spores at magnification 400 X by Compound light microscope (Olympus CX 23).
Figure 2.
Typical blast symptom on rice (Oryza sativa L.) cultivar RD6 leaves 14 days after inoculation (a) Healthy leaf, (b) infected sick leaves and (c) Pyricularia sp. WPP09 spores at magnification 400 X by Compound light microscope (Olympus CX 23).

Figure 3.
Macroscopic determination and Microscopic determination of Streptomyces strain PC 12 (a)Streptomyces strain PC 12 culture on ISP2 medium at 28 °C for 7 days showed substrate mycelium (b,c) Streptomyces strain PC 12 culture on ISP2 medium at 28 °C for 7 days showed white aerial mycelium (d) Photomicrograph of spiral spore chains of Streptomyces strain PC 12 under Compound light microscope.
Figure 3.
Macroscopic determination and Microscopic determination of Streptomyces strain PC 12 (a)Streptomyces strain PC 12 culture on ISP2 medium at 28 °C for 7 days showed substrate mycelium (b,c) Streptomyces strain PC 12 culture on ISP2 medium at 28 °C for 7 days showed white aerial mycelium (d) Photomicrograph of spiral spore chains of Streptomyces strain PC 12 under Compound light microscope.

Figure 4.
Scanning electron micrograph of the spore chain and the spiny spores of Streptomyces strain PC-12 culture on the ISP 2 agar for 14 days.
Figure 4.
Scanning electron micrograph of the spore chain and the spiny spores of Streptomyces strain PC-12 culture on the ISP 2 agar for 14 days.

Figure 5.
Neighbor-joining tree showing the position of strain PC 12 compared to its related Streptomyces species. Numbers at the nodes indicate the percentage of bootstrap values based on 1000 replicates. Only values greater than 50% are shown.
Figure 5.
Neighbor-joining tree showing the position of strain PC 12 compared to its related Streptomyces species. Numbers at the nodes indicate the percentage of bootstrap values based on 1000 replicates. Only values greater than 50% are shown.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Morphological characterization of one-hundred and twelve Streptomyces isolated from the rhizospheric soil of rice plant.
Table 1.
Morphological characterization of one-hundred and twelve Streptomyces isolated from the rhizospheric soil of rice plant.
| No. | Color of Aerial Mycelium | Soluble Pigment | Number of Isolates | Percentage of all Isolates (%) |
|---|---|---|---|---|
| 1 | Gray | Brown | 41 | 36.6 |
| 2 | Brown | Brown | 23 | 20.5 |
| 3 | White-Gray | Brown | 14 | 12.5 |
| 4 | White | Brown | 12 | 10.7 |
| 5 | Yellow | - | 10 | 8.9 |
| 6 | Green | - | 3 | 2.7 |
| 7 | Orange | Orange | 3 | 2.7 |
| 8 | Red | Brown | 3 | 2.7 |
| 9 | Pink | - | 2 | 1.8 |
| 10 | Black | - | 1 | 0.9 |
| Total | 112 | 100 |
Table 2.
Plant growth promoting and antifungal activities of rhizosphere Streptomyces spp.
| Isolate. | Inhibition Zone (mm) | HCN Production | ||||
|---|---|---|---|---|---|---|
| Antagonistic to Pyricularia sp. (%) | Siderophore Production | Protease Production | Cellulase Production | Chitinase Production | ||
| PC-1 | 39.5 ± 2.7 f | 11.4 ± 0.9 e | 51.3 ± 1.9 b | - | - | + |
| PC-5 | 41.3 ± 1.8 e | 8.9 ± 0.7 f | 50.6 ± 1.5 b | - | - | + |
| PC-7 | 29.7 ± 3.5 g | 10.4 ± 0.5 e | 40.7 ± 1.1 c | - | - | + |
| PC-9 | 22.6 ± 2.9 g | 12.5 ± 0.9 e | 43.2 ± 1.3 c | - | - | + |
| PC-12 | 87.3 ± 1.8 b | 24.5 ± 0.9 d | 25.7 ± 1.3 e | 57.2 ± 3.1 c | 40.3 ± 2.7 c | + |
| PC-17 | 45.6 ± 3.1 e | 18.5 ± 1.1 e | 28.6 ± 1.5 e | 43.1 ± 2.7 d | 35.7 ± 2.1 d | + |
| PC-23 | 31.2 ± 4.9 f | 17.3 ± 1.3 e | 33.5 ± 1.3 d | 27.9 ± 1.7 e | - | + |
| PC-24 | 71.6 ± 1.4 bc | 32.4 ± 1.3 c | 73.7 ± 1.5 a | - | - | + |
| PC-37 | 38.4 ± 7.8 f | 28.9 ± 1.5 d | 38.4 ± 1.7 d | - | - | + |
| PC-39 | 51.7 ± 1.9 d | 17.5 ± 1.1 e | 34.7 ± 1.5 d | - | - | + |
| PC-42 | 66.0 ± 3.5 c | 32.8 ± 1.7 c | 46.2 ± 1.1 c | - | - | + |
| PC-47 | 33.5 ± 3.1 f | 36.7 ± 1.9 c | 51.5 ± 1.9 b | 40.5 ± 3.7 d | 21.4 ± 1.3 e | + |
| PC-55 | 27.7 ± 7.5 g | 41.0 ± 1.7 b | 44.3 ± 1.7 c | 64.2 ± 2.7 b | 27.9 ± 2.7 e | + |
| PC-63 | 69.3 ± 3.7 c | 40.5 ± 1.3 b | 52.4 ± 1.5 b | - | - | + |
| PC-71 | 21.0 ± 1.7 h | 34.6 ± 1.1 c | 47.2 ± 1.9 c | - | - | + |
| PC-73 | 38.6 ± 2.1 f | 43.4 ± 1.3 b | 24.2 ± 1.3 e | - | - | + |
| PC-88 | 26.1 ± 0.8 g | 42.8 ± 1.5 b | 35.7 ± 1.7 d | - | - | + |
| PC-94 | 71.3 ± 1.4 bc | 50.2 ± 1.3 a | 28.9 ± 1.5 e | - | - | + |
| PC-102 | 38.3 ± 4.0 f | 34.5 ± 1.5 c | 47.6 ± 1.3 c | 57.6 ± 4.1 c | - | + |
| PC-110 | 49.5 ± 2.3 e | 37.4 ± 1.3 c | 28.7 ± 1.7 e | - | - | + |
| PC-117 | 67.2 ± 4.2 c | 50.7 ± 1.1 a | 37.8 ± 1.9 d | - | - | + |
| PC-119 | 75.8 ± 3.7 b | 40.8 ± 1.5 b | 45.4 ± 1.5 c | - | - | + |
| PC-120 | 74.3 ± 4.2 b | 31.7 ± 1.3 c | 50.2 ± 1.9 b | - | - | + |
| D 4.1 | 82.2 ± 2.5 a | 52.3 ± 1.9 a | 53.5 ± 1.9 b | 70.8 ± 4.7 a | 57.4 ± 3.1 a | + |
| D 4.3 | 80.0 ± 2.7 a | 51.4 ± 1.1a | 52.2 ± 1.9 b | 69.5 ± 3.7 b | 48.5 ± 2.7 b | + |
| W 1 | 80.5 ± 1.7 a | 47.8 ± 1.5 c | 50.7 ± 1.5 b | 57.8 ± 1.9 c | 34.8 ± 2.1 d | + |
| W 9 | 76.4 ± 3.2 b | 41.5 ± 1.1 c | 34.7 ± 1.3 d | - | - | + |
“Positive” (+): Having trait; “Negative” (-): Not Having trait. Data are means ± standard error over four replicates. Numbers in column followed by the same letter are not significantly different according to Duncan’s multiple range test (DMRT) at P ≤ 0.05.
Table 3.
Effect of selected Streptomyces application on blast disease and plant growth promotion under greenhouse conditions.
Table 3.
Effect of selected Streptomyces application on blast disease and plant growth promotion under greenhouse conditions.
| Treatment | Blast disease Parameters 60 Days | Plant growth Parameters60 Days | |||||
|---|---|---|---|---|---|---|---|
| Lesion Length (cm) | No. of Dried Leaves per Plant | Disease Severity (%) | Plant Height (cm) | Root Length (cm) | Root Dry Weight (g) | No of Tiller | |
| Streptomyces W1 | 1.19 ± 0.21 b | 3.67 ± 0.27 c | 51.9 ± 0.31 c | 45.08 ± 0.57 c | 29.43 ± 0.17 c | 2.13 ± 0.07 ab | 12.8 ± 0.31 bc |
| Streptomyces D 4.3 | 0.98 ± 0.19 b | 3.37 ± 0.21 c | 44.5 ± 0.37 b | 52.69 ± 0.47 b | 37.00 ± 0.19 b | 2.53 ± 0.11 a | 15.2 ± 0.27 b |
| Streptomyces PC 12 | 0.57 ± 0.19 c | 3.10 ± 0.19 c | 31.4 ± 0.37 a | 76.93 ± 0.59 a | 41.60 ± 0.27 a | 2.68 ± 0.17 a | 19.4 ± 0.45 a |
| Streptomyces D 4.1 | 1.07 ± 0.15 b | 4.50 ± 0.29 b | 42.8 ± 0.31 b | 43.57 ± 0.37 c | 31.89 ± 0.21 b | 2.05 ± 0.19 ab | 14.7 ± 0.21 b |
| Pathogen (treated control) | 6.78 ± 1.17 a | 8.38 ± 0.35 a | 87.5 ± 0.47 d | 36.59 ± 0.41 d | 26.17 ± 0.15 c | 1.88 ± 0.13 b | ND |
| Healthy (Untreated control) | ND | ND | ND | 40.70 ± 0.35 cd | 29.43 ± 0.29 c | 1.96 ± 0.17 b | 9.20 ± 0.31 d |
ND = Not detectable; data are means ± standard error over four replicates. Numbers in column followed by the same letter are not significantly different according to Duncan’s multiple range test (DMRT) at P ≤ 0.05.
Table 4.
Cell viability of selected Streptomyces sp. at 60 dpi.
| Streptomyces Strains | Cell Number (CFU/mL) |
|---|---|
| Streptomyces W1 | 3.2 × 103 |
| Streptomyces D 4.3 | 3.4 × 104 |
| Streptomyces PC 12 | 4.5 × 104 |
| Streptomyces D 4.1 | 2.7 × 103 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chaiharn, M.; Theantana, T.; Pathom-aree, W. Evaluation of Biocontrol Activities of Streptomyces spp. against Rice Blast Disease Fungi. Pathogens 2020, 9, 126. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9020126
AMA Style
Chaiharn M, Theantana T, Pathom-aree W. Evaluation of Biocontrol Activities of Streptomyces spp. against Rice Blast Disease Fungi. Pathogens. 2020; 9(2):126. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9020126
Chicago/Turabian StyleChaiharn, Mathurot, Teerayut Theantana, and Wasu Pathom-aree. 2020. "Evaluation of Biocontrol Activities of Streptomyces spp. against Rice Blast Disease Fungi" Pathogens 9, no. 2: 126. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9020126
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.