Identification and Genome Analysis of Vibrio coralliilyticus Causing Mortality of Pacific Oyster (Crassostrea gigas) Larvae


 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Rule-Out and Identification of Causative Agent
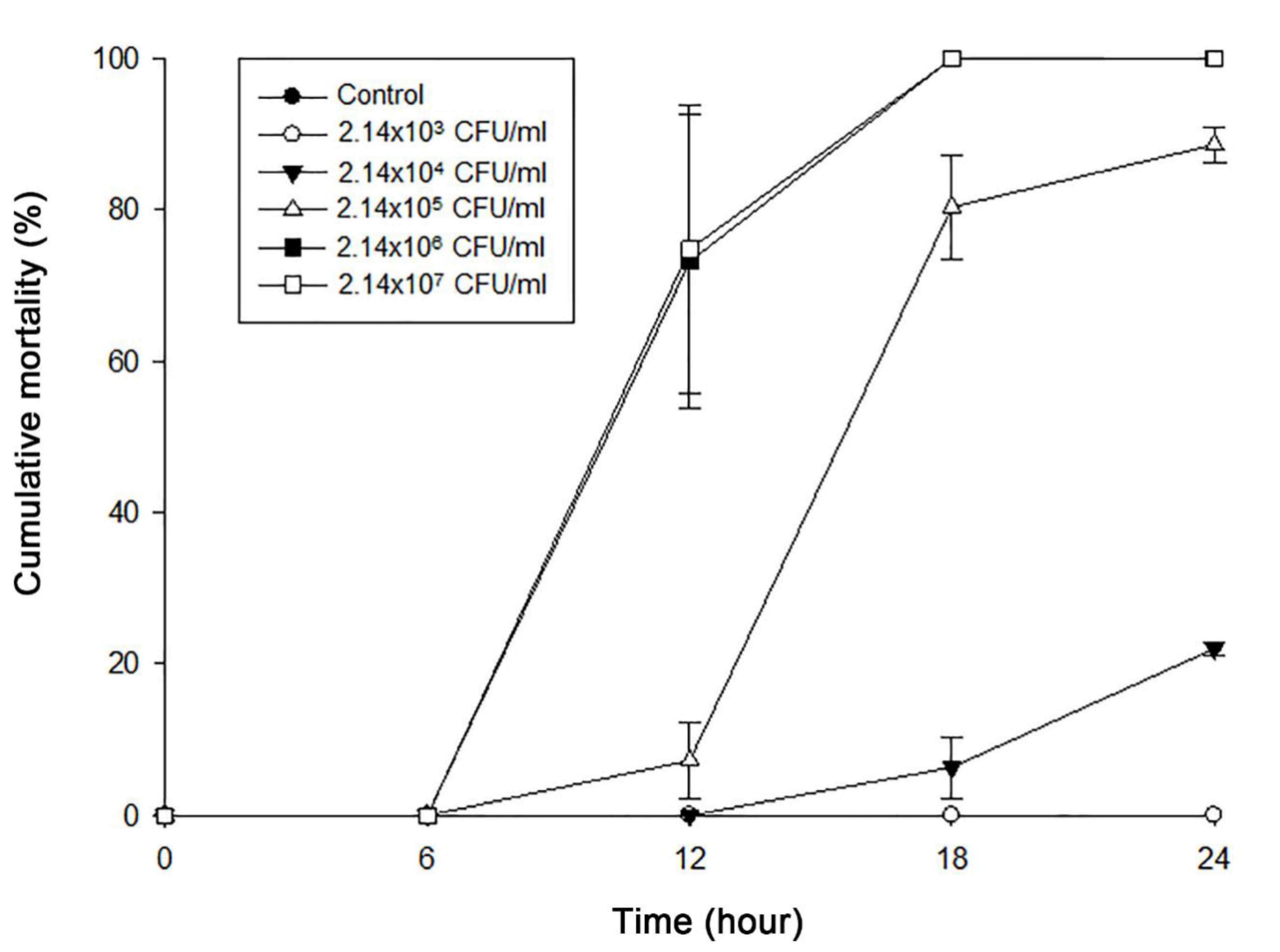
2.2. Pathogenicity of Isolated V. Coralliilyticus SNUTY-1
2.3. Anti-Microbial Susceptibility and Biochemical Analysis of V. coralliilyticus SNUTY-1
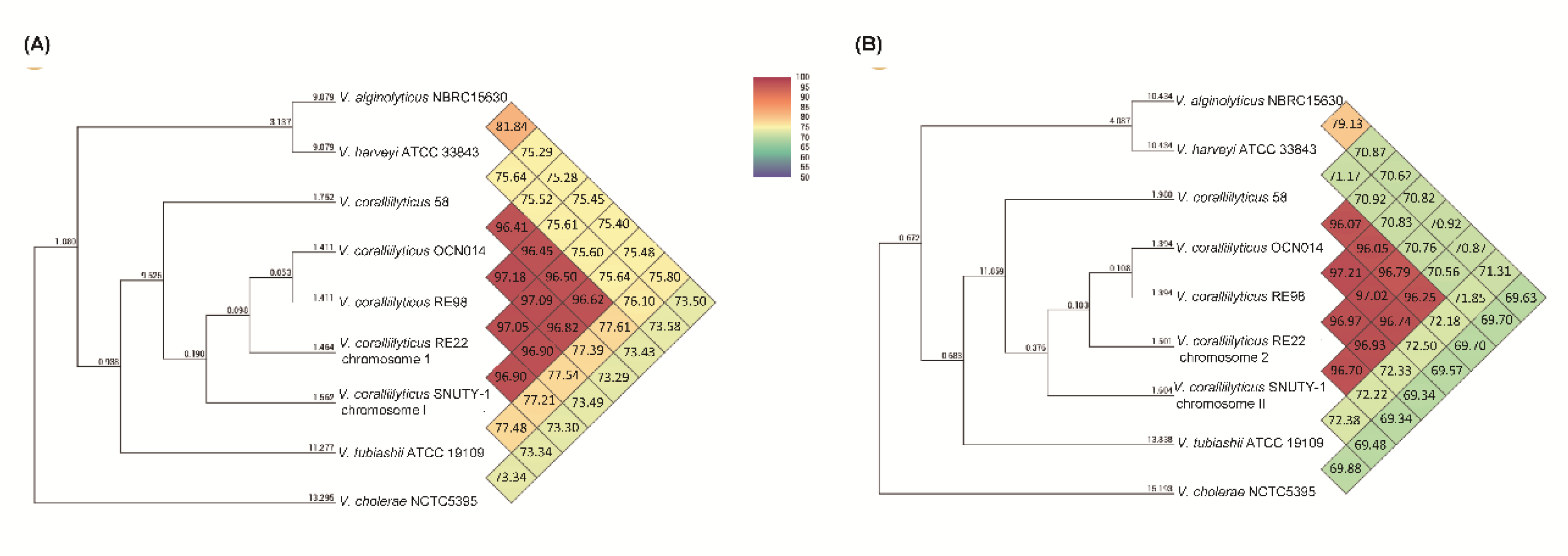
2.4. Genome of V. coralliilyticus SNUTY-1
3. Materials and Methods
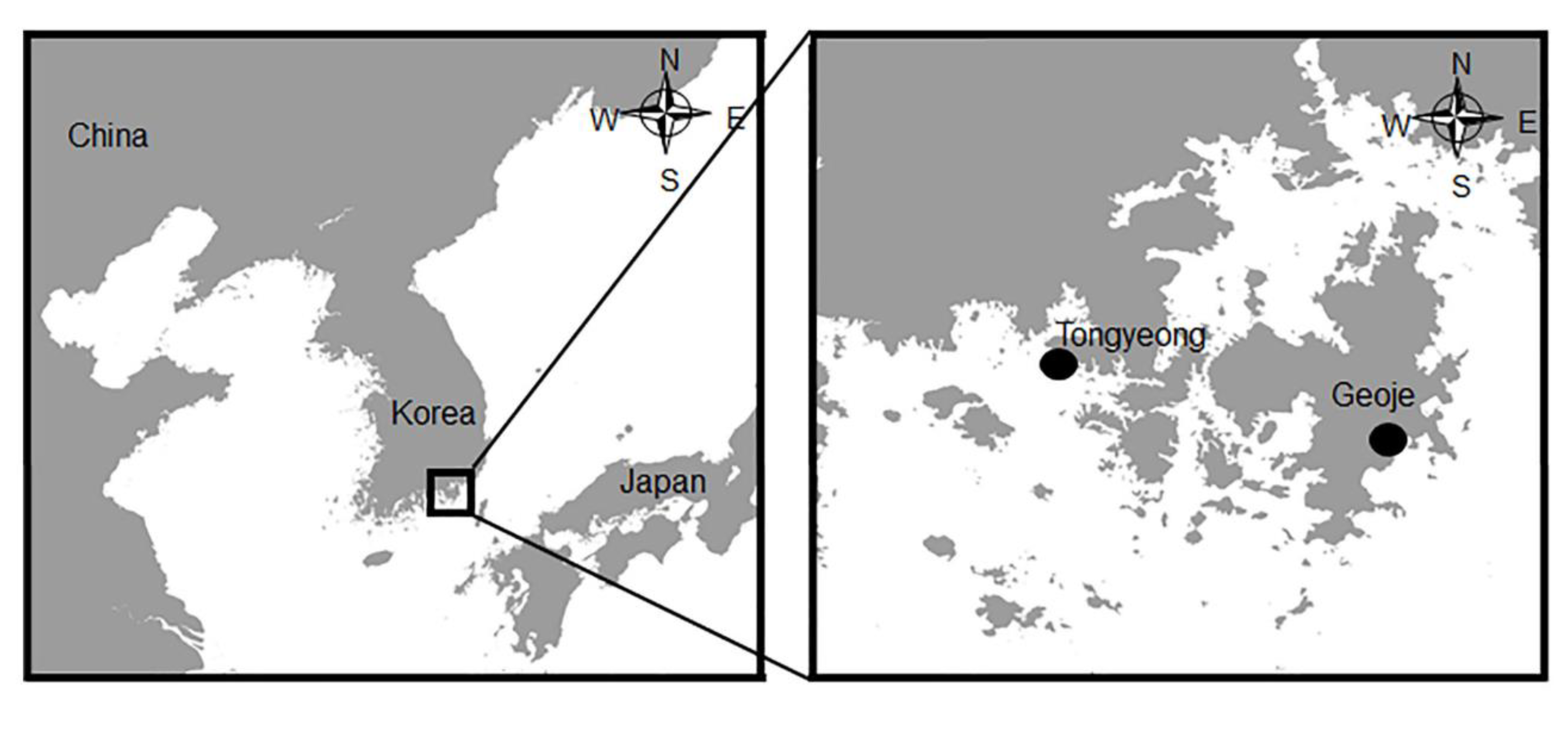
3.1. Sampling and Clinical Examination
3.2. Molecular Detection of Pathogen
3.3. Bacteria Isolation and Identification
3.4. Pathogenicity Test of Isolated Bacterium
3.5. Anti-Microbial Susceptibility Test and Biochemical Analysis
3.6. Genome Sequencing
4. Conclusions
5. Culture Deposition and Nucleotide Sequence Accession Numbers
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO FishStat. Global Aquaculture Production for Species (Tonnes): Pacific Oyster. 2019. Available online: http://www.fao.org/fishery/topic/16140/en (accessed on 31 October 2019).
- Le Roux, F.; Zouine, M.; Chakroun, N.; Binesse, J.; Saulnier, D.; Bouchier, C.; Zidane, N.; Ma, L.; Rusniok, C.; Lajus, A.; et al. Genome sequence of Vibrio splendidus: An abundant planktonic marine species with a large genotypic diversity. Environ. Microbiol. 2009, 11, 1959–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Salvo, L.H.; Blecka, J.; Zebal, R. Vibrio anguillarum and larval mortality in a California coastal shellfish hatchery. Appl. Environ. Microbiol. 1978, 35, 219–221. [Google Scholar] [CrossRef] [Green Version]
- Elston, R.A. Infectious diseases of the Pacific oyster Crassostrea gigas. Annu. Rev. Fish Dis. 1993, 3, 259–276. [Google Scholar] [CrossRef]
- Gómez-León, J.; Villamil, L.; Lemos, M.L.; Novoa, B.; Figueras, A. Isolation of Vibrio alginolyticus and Vibrio splendidus from aquacultured carpet shell clam (Ruditapes decussatus) larvae associated with mass mortalities. Appl. Environ. Microbiol. 2005, 71, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Lodeiros, C.; Bolinches, J.; Dopazo, C.P.; Toranzo, A.E. Bacillary necrosis in hatcheries of Ostrea edulis in Spain. Aquaculture 1987, 65, 15–29. [Google Scholar] [CrossRef]
- Sugumar, G.; Nakai, T.; Hirata, Y.; Matsubara, D.; Muroga, K. Vibrio splendidus biovar II as the causative agent of bacillary necrosis of Japanese oyster Crassostrea gigas larvae. Dis. Aquat. Organ. 1998, 33, 111–118. [Google Scholar] [CrossRef]
- Mersni-Achour, R.; Cheikh, Y.B.; Pichereau, V.; Doghri, I.; Etien, C.; Dégremont, L.; Saulnier, D.; Fruitier-Arnaudin, I.; Travers, M.-A. Factors other than metalloprotease are required for full virulence of French Vibrio tubiashii isolates in oyster larvae. Microbiology 2015, 161, 997–1007. [Google Scholar] [CrossRef]
- Richards, G.P.; Watson, M.A.; Needleman, D.S.; Church, K.M.; Häse, C.C. Mortalities of Eastern and Pacific oyster larvae caused by the pathogens Vibrio coralliilyticus and Vibrio tubiashii. Appl. Environ. Microbiol. 2015, 81, 292–297. [Google Scholar] [CrossRef] [Green Version]
- Ben-Haim, Y.; Thompson, F.L.; Thompson, C.C.; Cnockaert, M.C.; Hoste, B.; Swings, J.; Rosenberg, E. Vibrio coralliilyticus sp. nov., a temperature-dependent pathogen of the coral c. Int. J. Syst. Evol. Microbiol. 2003, 53, 309–315. [Google Scholar] [CrossRef]
- Ben-Haim, Y.; Zicherman-Keren, M.; Rosenberg, E. Temperature-Regulated Bleaching and Lysis of the Coral Pocillopora damicornis by the Novel Pathogen Vibrio coralliilyticus. Appl. Environ. Microbiol. 2003, 69, 4236–4242. [Google Scholar] [CrossRef] [Green Version]
- Austin, B.; Austin, D.; Sutherland, R.; Thompson, F.; Swings, J. Pathogenicity of Vibrios to rainbow trout (Oncorhynchus mykiss, Walbaum) and Artemia nauplii. Environ. Microbiol. 2005, 7, 1488–1495. [Google Scholar] [CrossRef] [PubMed]
- Genard, B.; Miner, P.; Nicolas, J.-L.; Moraga, D.; Boudry, P.; Pernet, F.; Tremblay, R. Integrative study of physiological changes associated with bacterial infection in Pacific oyster larvae. PLoS ONE 2013, 8, e64534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesarcodi-Watson, A.; Miner, P.; Nicolas, J.-L.; Robert, R. Protective effect of four potential probiotics against pathogen-challenge of the larvae of three bivalves: Pacific oyster (Crassostrea gigas), flat oyster (Ostrea edulis) and scallop (Pecten maximus). Aquaculture 2012, 344, 29–34. [Google Scholar] [CrossRef]
- Elston, R.A.; Hasegawa, H.; Humphrey, K.L.; Polyak, I.K.; Häse, C.C. Re-emergence of Vibrio tubiashii in bivalve shellfish aquaculture: Severity, environmental drivers, geographic extent and management. Dis. Aquat. Organ. 2008, 82, 119–134. [Google Scholar] [CrossRef]
- Travers, M.; Achour, R.M.; Haffner, P.; Tourbiez, D.; Cassone, A.; Morga, B.; Doghri, I.; Garcia, C.; Renault, T.; Fruitier-Arnaudin, I.; et al. First description of French V. tubiashii strains pathogenic to mollusk: I. Characterization of isolates and detection during mortality events. J. Invertebr. Pathol. 2014, 123, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Tubiash, H.S.; Chanley, P.E.; Leifson, E. Bacillary necrosis, a disease of larval and juvenile bivalve mollusks I. Etiology and epizootiology. J. Bacteriol. 1965, 90, 1036–1044. [Google Scholar] [CrossRef] [Green Version]
- Renault, T.; Arzul, I. Herpes-like virus infection in hatchery-reared bivalve larvae in Europe: Specific viral DNA detection by PCR. J. Fish Dis. 2001, 24, 161–167. [Google Scholar] [CrossRef]
- OIE. Manual of Diagnostic Tests for Aquatic Animals: Chapter 2.4.5. Infection with Ostreid Herpesvirus 1 Microvariants. 2018. Available online: https://www.oie.int/index.php?xml:id=2439&L=0&htmfile=chapitre_ostreid_herpesvirus_1.htm (accessed on 31 October 2019).
- Pollock, F.J.; Morris, P.J.; Willis, B.L.; Bourne, D.G. Detection and quantification of the coral pathogen Vibrio coralliilyticus by real-time PCR with TaqMan fluorescent probes. Appl. Environ. Microbiol. 2010, 76, 5282–5286. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Jun, J.W.; Giri, S.S.; Chi, C.; Yun, S.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Han, S.J.; Kwon, J.; et al. Application of the bacteriophage pVco-14 to prevent Vibrio coralliilyticus infection in Pacific oyster (Crassostrea gigas) larvae. J. Invertebr. Pathol. 2019, 167, 107244. [Google Scholar] [CrossRef]
- Stager, C.E.; Davis, J.R. Automated systems for identification of microorganisms. Clin. Microviol. Rev. 1992, 5, 302–327. [Google Scholar] [CrossRef]
- O’Hara, C.M.; Westbrook, G.L.; Miller, J.M. Evaluation of Vitek GNI+ and Becton dickinson microbiology system crystal E/NF identification system for identification of members of the Family Enterobacteriaceae and other gram-negative glucose-fermenting and non-glucose-fermenting Bacilli. J. Clin. Microbiol. 1997, 35, 3269–3273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushijima, B.; Videau, P.; Poscablo, D.; Vine, V.; Salcedo, M.; Aeby, G.; Callahan, S.M. Complete genome sequence of Vibrio coralliilyticus strain OCN014, isolated from a diseased coral at Palmyra atoll. Genome Announc. 2014, 2, e01318-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, G.P.; Bono, J.L.; Watson, M.A.; Needleman, D.S. Complete genome sequence for the shellfish pathogen Vibrio coralliilyticus RE98 isolated from a shellfish hatchery. Genome Announc. 2014, 2, e01253-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinard, E.; Kessner, L.; Gomez-Chiarri, M.; Rowley, D.C.; Nelson, D.R. Draft genome sequence of the marine pathogen Vibrio coralliilyticus RE22. Genome Announc. 2015, 3, e01432-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Kim, J.H.; Jun, J.W.; Giri, S.S.; Chi, C.; Yun, S.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Jeong, D.G.; et al. Complete genome sequence of Vibrio coralliilyticus 58, isolated from Pacific oyster (Crassostrea gigas) larvae. Genome Announc. 2017, 5, e00437-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, G.P.; Needleman, D.S.; Watson, M.A.; Bono, J.L. Complete genome sequence of the larval shellfish pathogen Vibrio tubiashii type strain ATCC 19109. Genome Announc. 2014, 2, e01252-14. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Kim, Y.O.; Park, S.-C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef]
- Aguirre-Guzmán, G.; Mejia Ruíz, H.; Ascencio, F. A review of extracellular virulence product of Vibrio species important in diseases of cultivated shrimp. Aquac. Res. 2004, 35, 1395–1404. [Google Scholar] [CrossRef]
- Hasegawa, H.; Lind, E.J.; Boin, M.A.; Häse, C.C. The extracellular metalloprotease of Vibrio tubiashii is a major virulence factor for Pacific oyster (Crassostrea gigas) larvae. Appl. Environ. Microbiol. 2008, 74, 4101–4110. [Google Scholar] [CrossRef] [Green Version]
- Miranda, C.D.; Kehrenberg, C.; Ulep, C.; Schwarz, S.; Roberts, M.C. Diversity of tetracycline resistance genes in bacteria from Chilean salmon farms. Antimicrob. Agents Chemother. 2003, 47, 883–888. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria M45, 3rd ed.; CLSI guideline M45; CLSI: Wayne, PA, USA, 2015. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Antibiotics | Strain | Antibiotics | Strain | ||
|---|---|---|---|---|---|
| SNUTY-1 | 58 | SNUTY-1 | 58 | ||
| Ampicillin | R * | R | Meropenem | R | R |
| Amoxicillin-clavulanate | R | I * | Amikacin | S * | I |
| Ampicillin-sulbactam | R | R | Gentamicin | S | S |
| Piperacillin | R | R | Tetracycline | S | S |
| Piperacillin-tazobactam | S | I | Ciprofloxacin | S | S |
| Cefepime | R | R | Levofloxacin | S | S |
| Cefotaxime | R | R | Ofloxacin | S | S |
| Cefoxitin | I | R | Sulfonamides | S | S |
| Ceftazidime | R | R | Trimethoprim-sulfamethoxazole | S | S |
| Cefuroxime sodium | S | S | Chloramphenicol | S | S |
| Imipenem | S | S | |||
| Items | Description |
|---|---|
| Classification | Domain Bacteria |
| Phylum Proteobacteria | |
| Order Vibrionales | |
| Genus Vibrio | |
| Species coralliilyticus | |
| Strain: SNUTY-1 | |
| General features | |
| Gram stain | Gram negative |
| Cell shape | Curved rod |
| Motility | Motile with polar flagella |
| Temperature | 4–37 °C |
| Pigmentation | Non-pigmented |
| MIGS data | |
| Investigation_type | Bacteria_archaea |
| Project_name | Genome sequence of Vibrio coralliilyticus SNUTY-1 |
| Lat_lon | 34˚47’13”N, 128˚25’22”E |
| Geo_loc_name | South Korea: Tongyeong |
| Collection_date | 2015-04 |
| Env_biome | landlocked sea [ENVO:00,000,219] |
| Env_feature | coastal water [ENVO:00,002,150] |
| Env_material | oyster [ENVO:02,000,079] |
| Num_replicons | 4 |
| Extrachrom_elements | 2 |
| Estimated_size | 5,842,676 |
| Ref_biomaterial | PMID: 9684317 |
| Source_mat_id | KCCM 43251 |
| biotic_relationship | Infectious (or commensal) |
| host | Crassostrea gigas (Pacific oyster) larvae |
| host_disease | bacillary necrosis |
| health_state | inactivated |
| Pathogenicity | Pathogenic in Pacific oyster (Crassostrea gigas) |
| Trophic_level | Chemoorganotroph |
| Rel_to_oxygen | Facultative anaerobic |
| Isol_growth_condt | PMID: 9684317 |
| Seq_meth | PacBio RSII |
| Annot_source | GenBank |
| Finishing_strategy | Complete; 160× coverage, 4 contigs |
| Genome assembly data | |
| Assembly method | HGAP |
| Assembly name | HGAP algorithm ver. 3 |
| Genome coverage | 237× |
| Sequencing technology | PacBio RSII |
| Attribute | Value | |||
|---|---|---|---|---|
| Chromosome I | Chromosome II | Plasmid pVs58 | Plasmid pVs58 | |
| Size (bp) | 3,474,874 | 1,976,676 | 254,703 | 136,423 |
| Coding regions (%) | 87.9 | 87.6 | 81.9 | 84.2 |
| G+C content | 45.7 | 45.1 | 49.3 | 44.7 |
| Total genes | 3358 | 1783 | 254 | 130 |
| tRNA genes | 111 | 5 | - | - |
| rRNA genes | 37 | - | - | - |
| ncRNA genes | 4 | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.J.; Jun, J.W.; Giri, S.S.; Chi, C.; Yun, S.; Kim, S.G.; Kim, S.W.; Han, S.J.; Kwon, J.; Oh, W.T.; et al. Identification and Genome Analysis of Vibrio coralliilyticus Causing Mortality of Pacific Oyster (Crassostrea gigas) Larvae. Pathogens 2020, 9, 206. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9030206
Kim HJ, Jun JW, Giri SS, Chi C, Yun S, Kim SG, Kim SW, Han SJ, Kwon J, Oh WT, et al. Identification and Genome Analysis of Vibrio coralliilyticus Causing Mortality of Pacific Oyster (Crassostrea gigas) Larvae. Pathogens. 2020; 9(3):206. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9030206
Chicago/Turabian StyleKim, Hyoun Joong, Jin Woo Jun, Sib Sankar Giri, Cheng Chi, Saekil Yun, Sang Guen Kim, Sang Wha Kim, Se Jin Han, Jun Kwon, Woo Taek Oh, and et al. 2020. "Identification and Genome Analysis of Vibrio coralliilyticus Causing Mortality of Pacific Oyster (Crassostrea gigas) Larvae" Pathogens 9, no. 3: 206. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9030206