1. Introduction
Brucellosis is a zoonotic bacterial disease that exhibits pathogenesis consistent with inflammation. Transmitted through
Brucella spp. primarily from agricultural animals to humans in unpasteurized dairy products, brucellosis symptoms in humans often include inflammatory or influenza-like characteristics such as arthritis, undulant fevers, and neurological manifestations [
1,
2]. Because there is currently no human vaccine for brucellosis and effective antibiotic regiments for the disease require long treatment durations, these symptoms often persist throughout the infected individual’s lifetime due to the well-adapted ability of
Brucella to evade immune recognition [
3]. Unlike the classical lipopolysaccharide (LPS) layer of Gram-negative bacteria such as
Escherichia coli that contain a glucosamine backbone with short acyl groups,
Brucella spp. contain a modified lipid A layer that consists of a diaminoglucose backbone with long branching acyl groups [
2]. This deviation from a consistent molecular structure has the potential to subvert immune recognition by the innate immune system through complement interference and decreased cytokine production, leading to enhanced
Brucella replication and pathogenesis. Despite its mechanisms of immune avoidance, there are some aspects of
Brucella spp. that are recognized by the innate immune system, making the understanding of these mechanisms essential for targeting future treatments for brucellosis.
As hypothesized by Janeway (1989), the innate immune system has evolved over time to recognize consistent molecular structures in pathogens known as Pathogen or Damage-Associated Molecular Patterns (PAMPs or DAMPs). These PAMPs and DAMPs are recognized by protein structures known as pattern recognition receptors (PRRs) [
4]. From previous studies,
Brucella genomic DNA (gDNA) is known to be recognized by the PRR absent in melanoma 2 (AIM2) and subsequently promotes inflammation, making it an excellent PAMP for immune recognition [
5,
6,
7]. PRRs include membrane-bound receptors, which consist of Toll-like receptors (TLRs) and C-type Lectin receptors (CLRs), as well as cytosolic receptors made up of a Nucleotide-Binding Domain and Leucine-Rich Repeat Containing receptors (NLRs), Aim-2-Like receptors (ALRs), Rig-I-Like Helicase receptors (RLRs), and the X-LR class of uncategorized receptors [
8,
9]. After the recognition of a PAMP or DAMP, PRRs generally serve as scaffolding proteins to promote the initiation or inhibition of immune signaling pathways [
9]. Of the PRRs that have been described in brucellosis, the best characterized have been the TLRs. From previous literature, many TLRs have been implicated with
Brucella detection, which plays a role in bacterial signaling, host resistance, and dendritic cell activation [
10,
11,
12,
13,
14,
15,
16]. TLRs also play important roles in the transcriptional generation of inactive inflammatory cytokines in response to
Brucella infections that can be activated by NLR or ALR immune signaling complexes [
17,
18]. This indicates that multiple PRRs work in tandem to attenuate brucellosis pathogenesis.
Inflammasome-forming NLRs and AIM2 have also been reported to play a role in
Brucella sensing [
5,
6,
7]. After recognition, the NLR or ALR is able to bind the apoptosis-associated speck-like protein containing a caspase activation recruitment domain (CARD) (ASC) and procaspase-1 to form the canonical inflammasome [
8]. The inflammasome then cleaves caspase-1, which subsequently cleaves the cytokines pro-IL-1β and pro-IL-18, produced through TLR signaling, to their active forms to promote inflammation [
8,
9,
19,
20,
21,
22,
23]. Inflammasome signaling can also lead to a form of inflammatory cell death known as pyroptosis. Pyroptosis occurs when activated capase-1 cleaves the protein gasdermin D, releasing the gasdermin N subunit [
24]. This subunit binds with phosphoinositides on the cell membrane and oligomerizes, creating membrane pores that lead to an osmotic imbalance in the cell that eventually leads to cell lysis [
24]. Recently, the formation of a non-canonical inflammasome has been described that utilizes caspase-11 to mediate the cleavage of gasdermin D to initiate pyroptosis.
Previous studies evaluating inflammasome activation in response to
Brucella have predominately focused on characterizing the activation of inflammatory cytokine signaling associated with canonical inflammasome activation. The best described inflammasomes involved in
Brucella infections are NLR Family Pyrin Domain Containing 3 (NLRP3) and AIM2. In mouse models, the NLRP3 inflammasome promotes survival and decreased bacterial load through enhanced cytokine secretion, in addition to sensing mitochondrial reactive oxygen species (ROS) generated from
Brucella [
6,
7]. Looking at the
Brucella PAMPs, AIM2, as a known sensor of bacterial DNA, becomes activated from
Brucella gDNA recognition and initiates inflammatory cytokine signaling and pyroptosis [
5,
6,
25]. These inflammasomes are ASC-dependent, as shown in the formation of punctate ASC structures during infection [
6], indicating that ASC-dependent inflammasomes are important in
Brucella recognition and targeting through inflammatory cytokine signaling. Despite these advancements in studying inflammasome-mediated inflammatory cytokine signaling, pyroptosis and the role of gasdermin D have not been extensively evaluated in response to
Brucella.In this study, we used Asc−/− and Caspase-11−/− mice to further elucidate the role of the canonical and non-canonical inflammasomes following B. abortus infection. We sought to assess survival, histopathology, bacterial load, and cell death to provide a more holistic view of cytokine responses and pyroptosis, both in vivo and in vitro. Additionally, we reassessed Brucella PAMPs using B. abortus gDNA and LPS to better define the mechanisms associated with pathogen recognition. Ultimately, we found that ASC functions to attenuate B. abortus pathogenesis through the modulation of inflammation and pyroptosis, requiring gasdermin D through a mechanism independent of caspase-11. Additionally, we determined that Brucella gDNA, rather than LPS, provoked an elevated inflammasome response that augmented pyroptosis. This report contributes to the current literature and provides some additional novel insights into potential mechanisms of inflammasome activation during brucellosis.
3. Discussion
In this report, we assessed inflammasome activation following
Brucella abortus infection. During canonical inflammasome activation, a pathogen is sensed by a NLR or ALR pattern recognition receptor and forms a multi-protein complex with the binding protein ASC and caspase-1. This process can initiate the cleavage of IL-1β and IL-18 in addition to pyroptosis [
8]. While highly related to the canonical inflammasome pathway, non-canonical inflammasome activation is often more closely associated with the promotion of pyroptosis and the activation of caspase-11 [
26]. Together, our in vivo data reveal a more robust phenotype in the
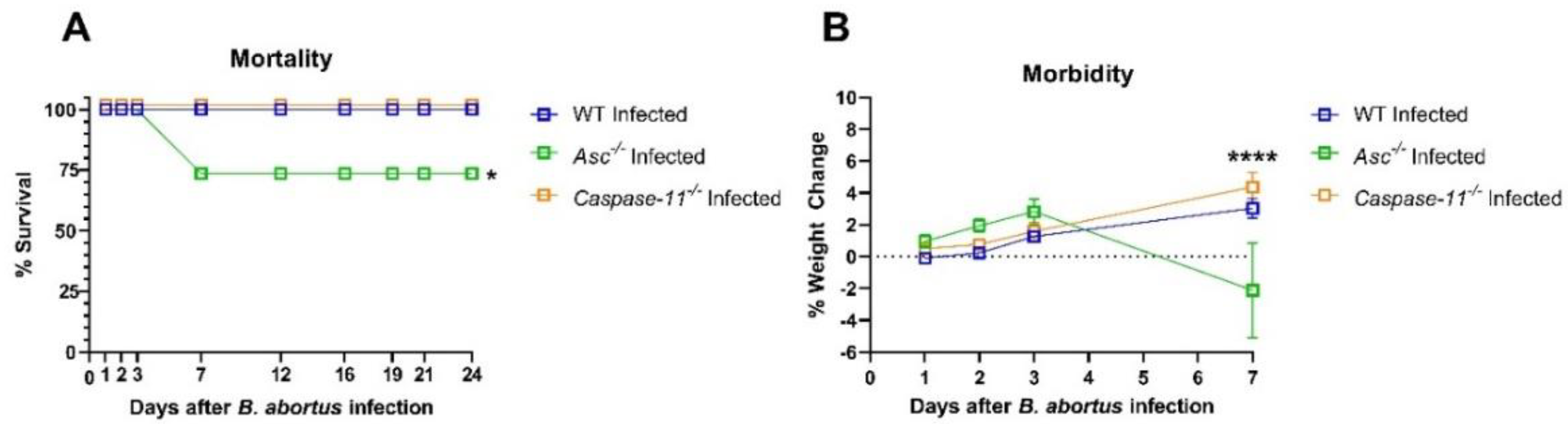
Asc−/− mice compared to the
Caspase-11−/− animals, suggesting that the canonical inflammasome plays a significantly greater role in host pathogen defense following
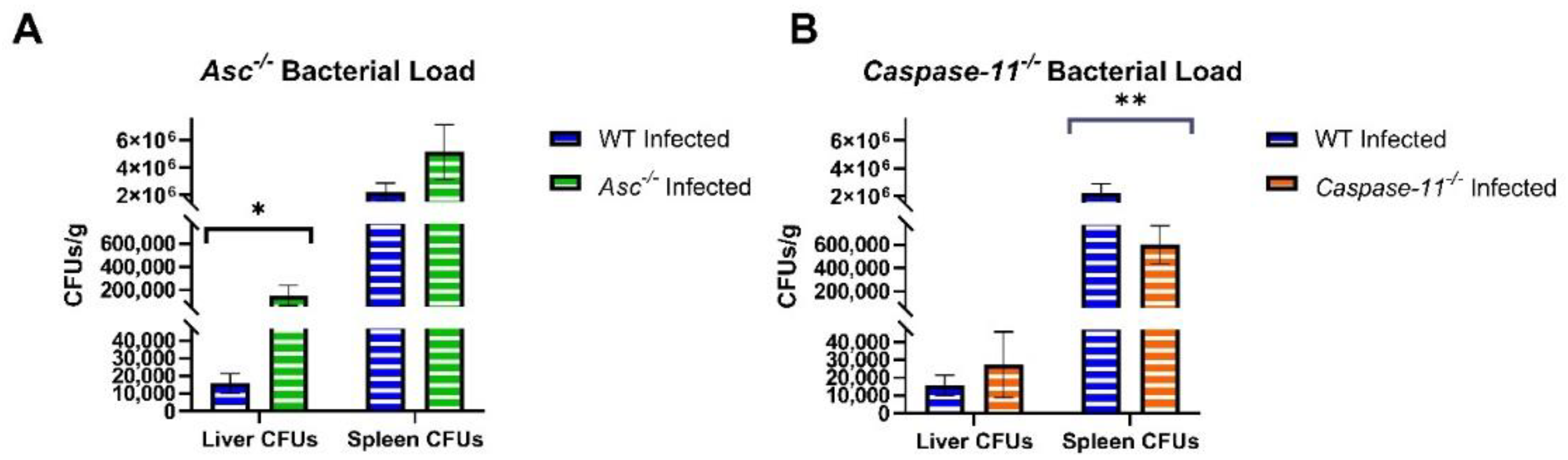
B. abortus infection. ASC and, by extension, the canonical inflammasome promotes survival, augments inflammation, and attenuates bacterial load
in vivo. This result is consistent with others in the field that have used myriad of other inflammasome knockout models [
5,
6,
7,
27]. However, there are certainly exceptions to these findings. For example, Gomes et al. (2103) recently reported no inflammasome knockout mice (
n = 5 per group) exhibited mortality under their experimental conditions [
6]. One difference to note is that our study utilized a much larger sample size per group (
n = 19). Because many of our animals recovered, it is certainly possible that greater power through that increased sample size is necessary to better reflect the mortality data for the
Asc−/− animals. Ultimately, our comparison between
Asc−/− and
Caspase-11−/− mice expands upon the findings of many of these prior studies and provides a more direct assessment of the pathobiological effects of the canonical and non-canonical inflammasome in
B. abortus host defense.
Although non-canonical signaling through the
Brucella LPS activation of caspase-11-mediated pyroptosis has been previously indicated [
28], we found inconsistent results in inflammasome activation under our conditions. In
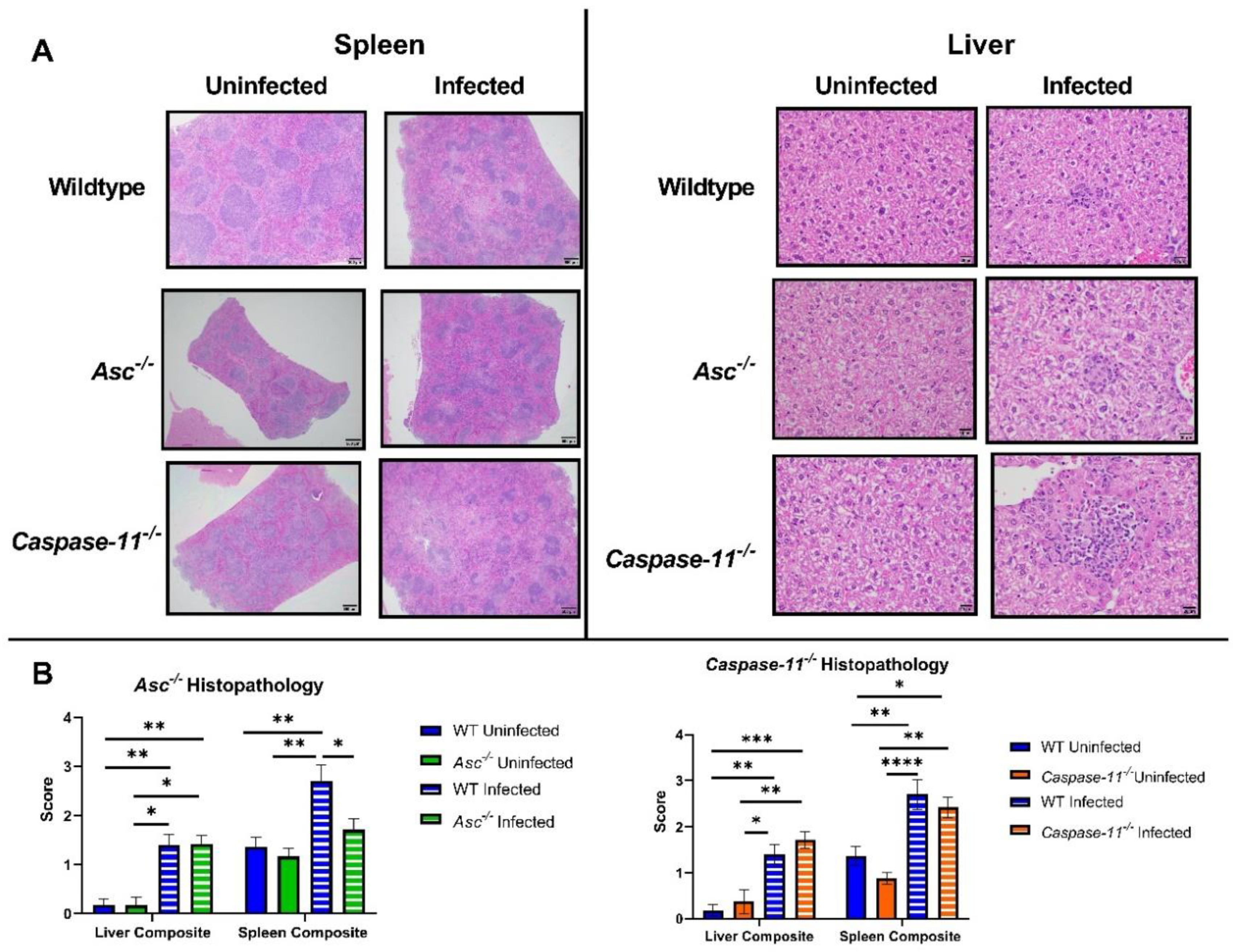
Caspase-11−/− mice, we found no loss in survival or morbidity, no significant inflammation through histopathology scoring, and no protective role in promoting inflammatory signaling. Previous studies using
Caspase-11−/− mice mimicked our non-significant results in bacterial load at 3 days post-infection and only found significant bacterial load 1–2 weeks after infection [
28]. Although we did not assess the bacterial load or inflammatory signaling after 3 d.p.i in this report, we did assess the morbidity and mortality of
Caspase-11−/− mice over 3 weeks, in which knockouts exhibited no decrease in morbidity or loss in survival. This indicates that caspase-11 may play a small but relatively insignificant role in promoting pyroptosis that perhaps may be slightly amplified 1–2 weeks after
Brucella infection. Additionally, these studies utilized immune cell priming in macrophages with PAMPs, such as
E. coli LPS and Pam3CSK4, and subsequently observed high cytokine signaling in their challenge [
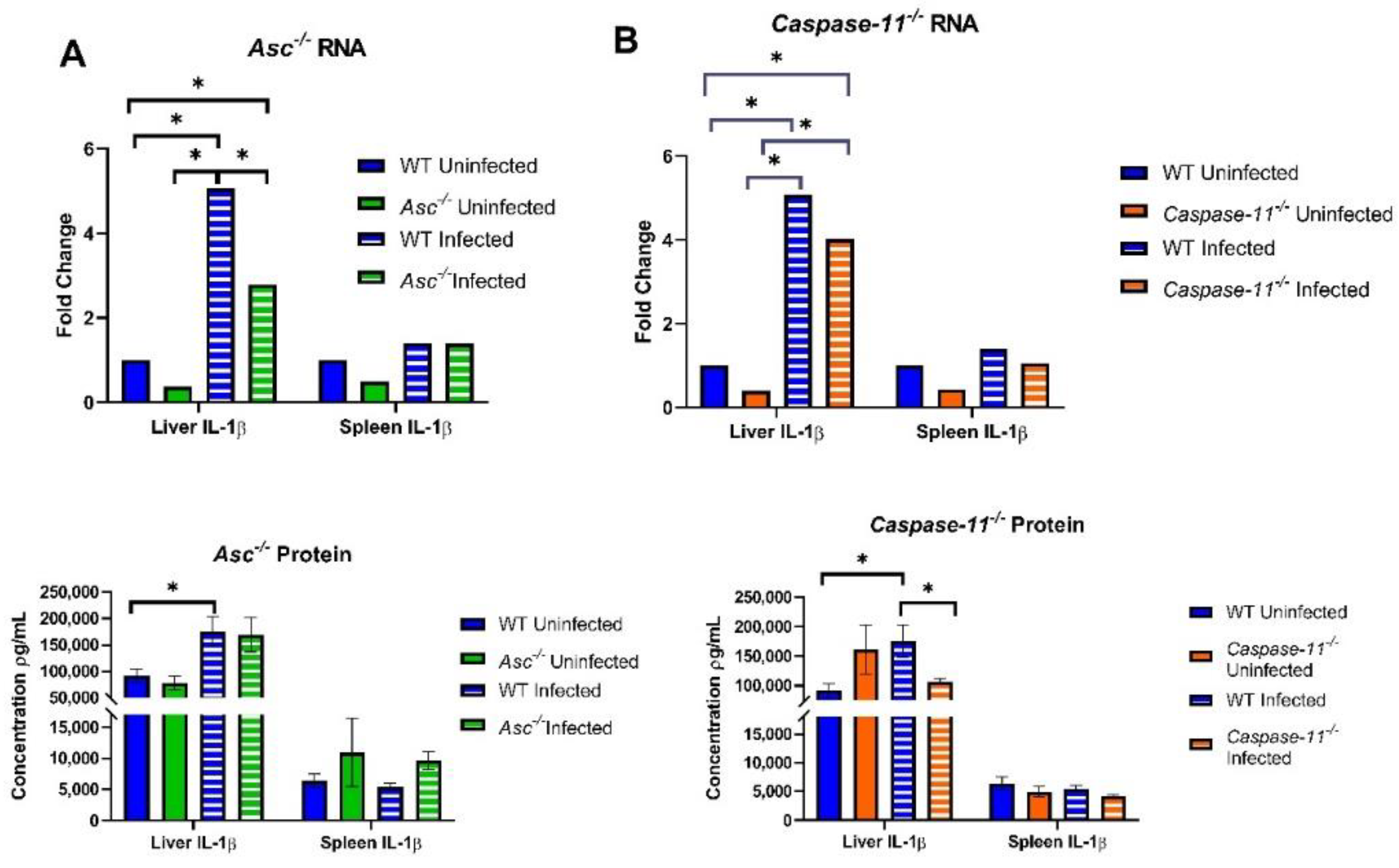
28]. Contrasting this data, our results indicated no IL-1β protein response to
Brucella LPS stimulation in unprimed macrophages. These data suggest that the immune adjuvants directly impact the activation of caspase-11 and that the role of non-canonical inflammasome in host defense against
B. abortus is minimal in the absence of macrophage priming.
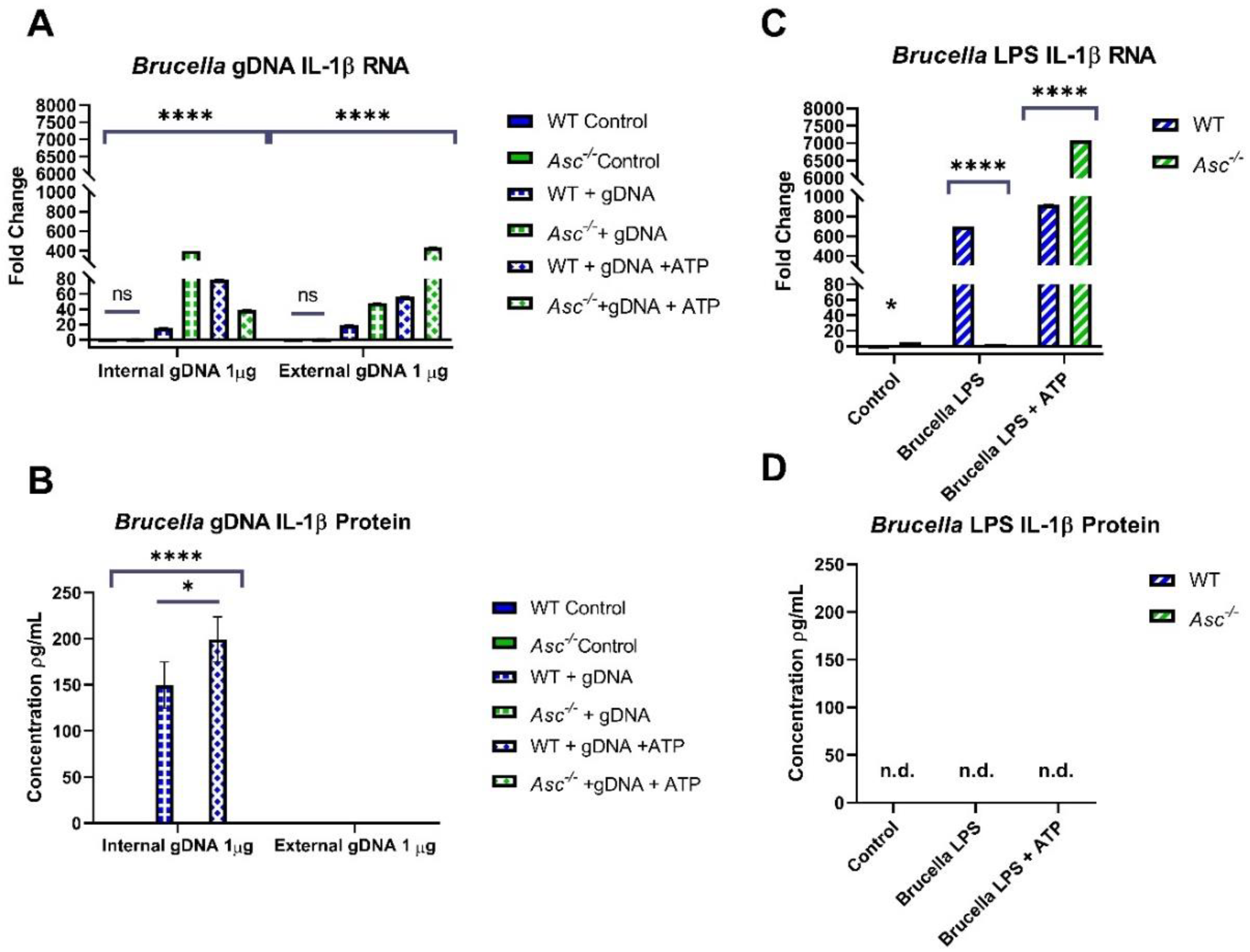
Previously, pyroptosis had only been attributed to non-canonical inflammasome activation by
Brucella spp. [
6,
28]. This is further confirmed in this report through the presence of cleaved gasdermin D bands in
Asc−/− macrophages, indicating that ASC-independent pyroptosis occurs in response to
Brucella. However, our research demonstrates that the removal of ASC-dependent inflammasome activation significantly decreases the activation of gasdermin D to form pyroptotic pores. As described in the literature, ASC specks serve as recruitment factors for procaspase-1 through the polymerization of its caspase activation recruitment domain (CARD). Caspase-1 only becomes activated through this process during ASC-dependent inflammasome formation [
29]. In turn, caspase-1 cleaves gasdermin D, which has been identified as the most significant gene initiating caspase-1 induced pyroptosis, and initiates inflammatory cell death [
30]. Our results are consistent with this ASC-mediated pathway of the capsase-1 activation of pyroptosis that is dependent on gasdermin D cleavage. To date, we know that
Brucella initiates the caspase-1 and -11 activation of pyroptosis in joints of animal models [
31]. Additionally, pyroptosis is activated by gDNA in dendritic cells [
5]. Our data supports a model where both caspase-1 and -11 promote pyroptosis, and where gDNA from
B. abortus functions as a robust PAMP that specifically activates the canonical inflammasome, driving ASC-dependent inflammation and pyroptosis.
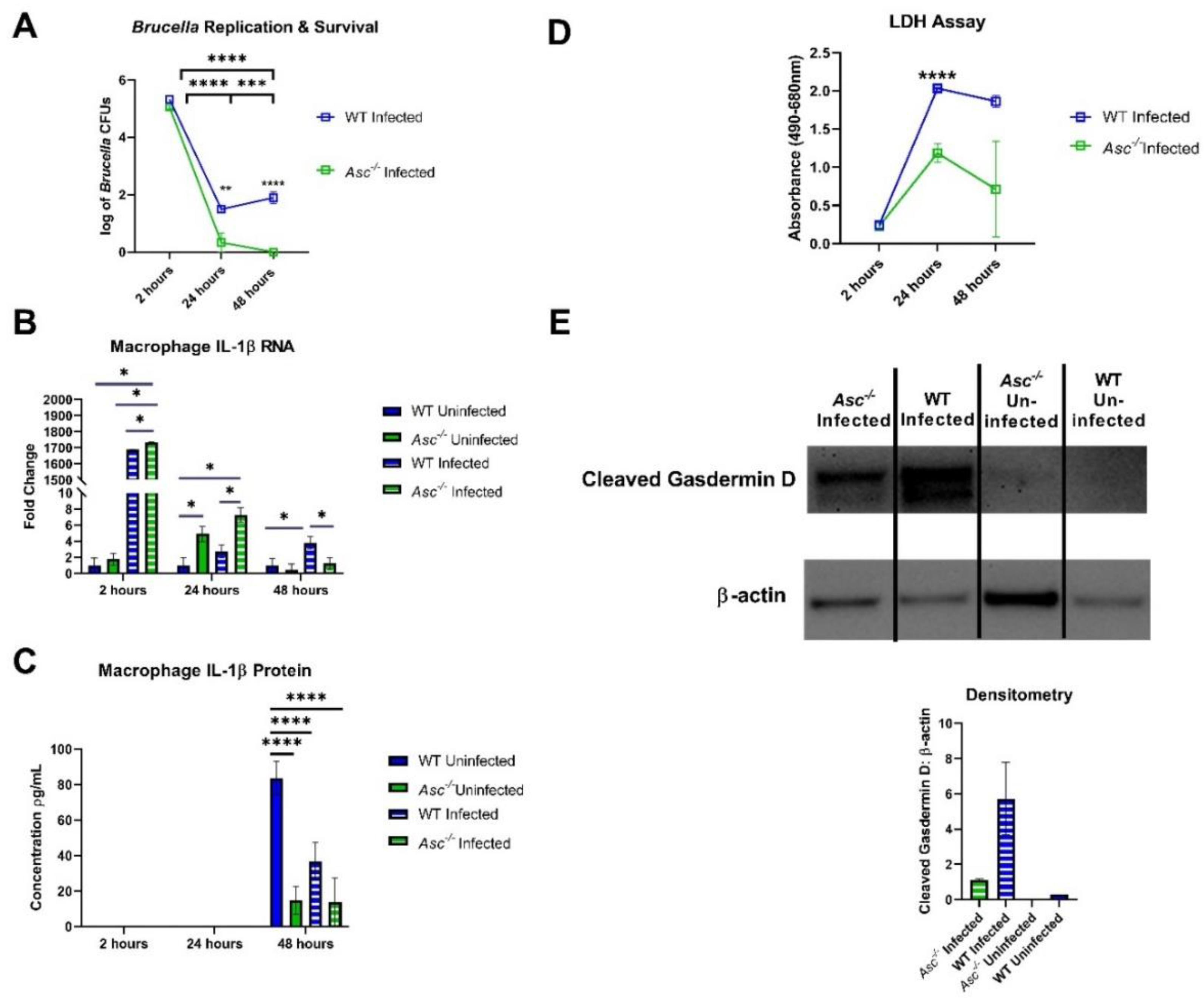
Previous literature indicates that the role of pyroptosis in brucellosis serves to restrict
Brucella growth in macrophages of the joints and control infection [
31]. Our findings are consistent with this previous study. Under our conditions, the ASC-mediated initiation of pyroptosis appears to ensure mouse survival, immune cell recruitment, and inflammatory signaling. However, we should also point out the bacteria clearance and IL-1β data in the BMDM studies (
Figure 5A,C). In
Figure 5A, these data would suggest that the lack of ASC and canonical inflammasome signaling actually improved the bacteria clearance from these BMDMs, despite having reduced IL-1β and pyroptosis. While these data seem to conflict with each other, several recent studies have reported similar findings for other bacterial pathogens. For example,
Citrobacter rodentium infection results in significant osmotic changes in targeted cells that can augment inflammasome signaling [
32]. However, the clearance of the pathogen itself appears to be independent of the inflammasome and the ASC modulation of inflammation and pyroptosis [
33]. Thus, it is possible that a similar mechanism is associated here with
B. abortus. Looking at the IL-1β graph in
Figure 5C, it suggests that
B. abortus infection in macrophages suppresses IL-1β production from wildtype cells. It is possible that the attenuation of total IL-1β in this figure may be due to the subversion of TLR signaling generating pro- IL-1β. Many studies have shown
Brucella subversion of TLR signaling through proteins such as Tcbp, which leads to decreased proinflammatory cytokine expression [
10]. Additionally,
Brucella microRNAs can lead to the downregulation of the mRNA and protein expression of innate immune PRRs [
34]. These mechanisms likely did not occur in our gDNA studies as there was no inclusion of these immunosuppressive proteins or production of inhibitory microRNAs. Therefore, it is possible that the full
Brucella bacterium may be utilizing these methods of immunosuppression to contribute to decreased total IL-1β.
Our data suggests that ASC and the canonical inflammasome contribute to host defense in response to B. abortus. These results are consistent with several others in the field and provide additional insight into host defense against this highly intriguing pathogen. Currently, brucellosis is having a significantly negative impact on a growing number of human populations worldwide. Therefore, it is essential that we expand our understanding of the underlying disease mechanisms and host immune response to B. abortus. This finding that the canonical inflammasome plays a dominate role in driving the host innate immune response and pyroptosis following the sensing of gDNA is an encouraging discovery that may contribute to the development of future therapeutics or strategic approaches to combat this disease and its underlying pathogen.
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
Brucella abortus 2308 was routinely grown on Schaedler blood agar (SBA), which is composed of Schaedler agar (BD, Franklin Lakes, NJ, USA) containing 5% defibrinated bovine blood (Quad Five, Ryegate, MT, USA). All work with live Brucella strains was performed in a biosafety level 3 (BSL3) facility. Animal work conducted in ABSL3 conditions was conducted under IACUC protocol # 14-055 at Virginia Tech following the ethical standards of animal use in research.
4.2. In Vivo Brucella Studies
C57BL/6 WT mice (n = 26), Asc−/− mice (C57BL/6 background; n = 19), and Caspase-11−/− (C57BL/6 background; n = 19) mice were inoculated intraperitoneally with 1 × 105 CFU of Brucella abortus. The percent weight change was measured each day post-Brucella inoculation to determine morbidity warranting euthanasia (>20%) following the IACUC protocol.
Additionally, 20 canonical knockout (Asc−/−, n = 6 U, 14 I), 27 non-canonical (Caspase-11−/−, n = 8 U, 19 I), and 34 C57BL/6 WT (n = 11 U, 23 I) mice were euthanized at 3 days post infection and harvested for the liver and spleen. Both organs were sectioned into three equal parts. The first section of both organs was taken for histopathology analysis to determine the scoring of extramedullary hematopoiesis (EMH) and inflammation. The remaining sections were homogenized in 1×PBS and analyzed for the number of CFUs per gram and the RNA and protein concentrations of IL-1β. RNA was isolated from the liver and spleen with TRIzol reagents (Invitrogen) followed by ethanol precipitation. Genomic DNA was removed with DNase I, and samples were cleaned using phenol-chloroform extractions and precipitated with ethanol. RNA samples were then resuspended in nuclease-free H2O, and the purity of samples was checked with a NanoDrop 1000 spectrophotometer (ThermoFisher).
After isolation, RNA was converted into 1 μg cDNA through a High-Capacity cDNA Reverse Transcription Kit (ThermoFisher). This cDNA was analyzed for IL-1β by RT-qPCR (40× cycles) using Taqman Fast MasterMix (ThermoFisher). Protein was determined through a sandwich ELISA kit (R&D systems).
4.3. Bone Marrow-Derived Macrophage (BMDM) Isolation
Bone marrow-derived macrophages (BMDMs) were derived from the bone marrow of C57BL/6 WT and
Asc−/− mice under the
IACUC protocol #18-104 at Virginia Tech. Two adult mice from each group were sacrificed using CO
2 fixation followed by cervical dislocation. Bone marrow was extracted from the tibias and femora of the mice. Cells were cultured in non-TC culture dishes in our formulation of culture media (Dulbecco’s Modified Eagle Medium (DMEM) (ThermoFisher) containing 10% Fetal Bovine Serum (FBS), 1% penicillin/streptomycin, 1% nonessential amino acids, and 20% L929 conditioned media [
35]). These culture dishes were incubated at 37 °C with 5% CO
2. After 6–7 days of culture, these cells had differentiated into macrophages. BMDMs were collected from the bottom of the plates through a cold 1× PBS solution containing 5 mM of EDTA. After 1 h on ice, we checked for macrophage detachment via microscope. Macrophages were collected and seeded at 500,000 cells/well in 24-well plates in media without antibiotics and left to adhere overnight in culture media.
4.4. Live Brucella Challenge in BMDMs
Macrophages were infected with a MOI 100:1 (10
7 CFUs/5 × 10
5 BMDMs) with
B. abortus 2308. At the 2, 24, and 48 h time points, intracellular
B. abortus was determined through a gentamicin protection assay [
36]. At 2 h, the infected macrophages were treated with gentamicin (50 μg/mL) for 1 h. Macrophages were then lysed with 0.1% deoxycholate in PBS, and serial dilutions were plated on Schaedler blood agar (SBA) containing 5% bovine blood (Quad Five). For the 24 and 48 h time points, macrophages were washed with PBS and fresh cell culture medium containing gentamicin (20 μg/mL) was added to the macrophages. At the indicated time points, macrophages were lysed, and serial dilutions were plated on SBA in triplicates.
The supernatant of BMDMs was sterile-filtered for later IL-1β protein analysis through a sandwich ELISA kit (R&D systems). Macrophages at each time point were lysed with 0.1% deoxycholate and isolated for RNA. After isolation, RNA was converted into 0.5μg cDNA through a High-Capacity cDNA Reverse Transcription Kit (ThermoFisher). This cDNA was analyzed for IL-1β by RT-qPCR (40× cycles) using Taqman Fast MasterMix (ThermoFisher).
4.5. LDH Assay
Macrophages were infected with a MOI 100:1 (107 CFUs/5 × 105 BMDMs) with B. abortus. At the 2, 24, and 48 h time points, extracellular B. abortus was killed through gentamicin (50 µg/mL). The supernatant of BMDMs was collected and centrifuged at 1000× g for 10 min to remove cell debris. The remaining supernatant (50 μL) was used for the CyQUANT™ LDH Cytotoxicity Assay kit (Invitrogen) and read at a corrected absorbance of 490–680 nm.
4.6. Brucella PAMP Isolation
4.6.1. Brucella gDNA
B. abortus gDNA was isolated from the 2308 strain by phenol:chloroform extraction. Approximately 3 mL of an overnight culture of B. abortus was pelleted by centrifugation. The pellet was resuspended in 200 μL of 0.04 M sodium acetate, 200 μL of 10% SDS, and 600 μL of TRIzol. A total of 250 μL of chloroform was added to this mixture in a Phase Lock tube (5PRIME) and centrifuged at 20,000× g (max speed) for 2 min. A second chloroform wash was performed in a Phase Lock tube. The resulting aqueous layer is then removed and added to 1 mL of 100% ethanol. DNA precipitation is carried out from this point.
4.6.2. Brucella LPS
B. abortus LPS was isolated from the
B. abortus 2308 using hot-phenol extraction, as described previously [
31]. Bacteria were killed with ethanol: acetone, and the cells were recovered by centrifugation. The pellet was suspended in deionized water at 66 °C and then mixed with 90% phenol
w/v that was heated to 66 °C. After stirring for 20 min, the suspension was chilled on ice. The solution was then subjected to centrifugation (15 min at 13,000×
g). The phenol layer was aspirated, filtered through a Whatman #1 filter, and the LPS was precipitated with methanol containing 1% methanol saturated with sodium acetate. Following incubation at 4 °C for 1 h, the mixture was subjected to centrifugation (10,000×
g for 10 min). The precipitate was stirred with deionized water for 12 h at 4 °C. Following centrifugation (10,000×
g for 10 min), the supernatant was precipitated with trichloroacetic acid, and the resulting supernatant following centrifugation (10,000×
g for 10 min) was dialyzed with deionized water and stored at −20 °C. The concentration of LPS was determined through the Pierce™ Chromogenic Endotoxin Quant Kit (ThermoFisher).
4.7. Brucella PAMP Challenge
After PAMP isolation, B. abortus gDNA (2 μg/mL) and LPS (1 μg/mL) were introduced to BMDMs. gDNA was introduced both intracellularly, through the Lipofectamine 3000 Transfection Reagent (Invitrogen), and extracellularly in media. LPS was only introduced extracellularly. Timepoints for this challenge included 24 h for gDNA and 8 h for LPS post-challenge. Samples were run with and without the priming of 300 μM of ATP 45 min before each time point to stimulate IL-1β protein release after transcription. At each time point, supernatant was collected from each well and centrifuged at 1000× g for 10 min to remove cell debris. The supernatant was later used for protein quantification using an IL-1β sandwich ELISA (R&D Systems). Macrophages were lysed with 200 μL of TRIzol Reagent (Invitrogen) and followed the TRIzol RNA isolation protocol. After isolation, RNA was converted into 1 μg of cDNA through a High-Capacity cDNA Reverse Transcription Kit (ThermoFisher). This cDNA was analyzed for IL-1β by RT-qPCR (40× cycles) using the Taqman Fast MasterMix (ThermoFisher).
4.8. Western Blot
Macrophages from live Brucella challenge were lysed using a sodium lysis buffer (0.3% SDS, 200 mM dithiothreitol, 22 mM Tris-base, and 28 mM Tris-HCl pH 8.0). Samples were then boiled for a period of 1 h, vortexing the samples every 10 min. Samples were frozen at −20 °C until use. Prior to running the gel, samples were sonicated for 10 s each. Protein quantification was determined using the Pierce™ Detergent Compatible Bradford Assay Kit (ThermoFisher). Samples (20 μg of protein) were heated at 97 °C with a reducing buffer for 7 min and run on pre-cast Bolt™ 4 to 12%, Bis-Tris, 1.0 mm, Mini Protein Gel, 10-well gels (Invitrogen) for 45 min at 165 V with a 1× Micro Extraction Packet Sorbent (MEPS) buffer (ThermoFisher). Gel was transferred onto a polyvinylidene difluoride (PVDF) membrane using a transfer chamber with transfer buffer (20% methanol in 1× Tris Glycine (TGE)). The membrane was then blocked in 5% milk for 1 h, and then incubated overnight with a cleaved gasdermin D rabbit antibody (diluted 1:1000, Cell Signaling #36425S). Membrane was washed in Tris-buffered saline with Tween 20 (1× TBST) 4× for 15 min each and then blocked with goat anti-rabbit IgG antibody conjugated with horseradish peroxidase (HRP) (diluted 1:2000, Cell Signaling #7074) for 1 h. Membrane was washed again with 1× TBST 4× for 15 min each and then imaged using West Pico Substrate for imaging (ThermoFisher). Gels were imaged using an IBright CL1500 imaging machine. Sample bands were normalized using the β-actin rabbit antibody (Cell Signaling #4970) using the same protocol above. The density of bands was calculated using the IBright Analysis software (ThermoFisher) to calculate a cleaved gasdermin D/β-actin ratio.
4.9. Graphing and Statistical Analyses
All the figures and statistical analyses were generated in GraphPad 8.4.3 (Prism). Statistical tests included two-way ANOVAs, using the Tukey or Sidak post-hoc tests, and two sample t-tests when appropriate. All the data are contained within the article.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





