
A Novel Combination Therapy Using Rosuvastatin and Lactobacillus Combats Dextran Sodium Sulfate-Induced Colitis in High-Fat Diet-Fed Rats by Targeting the TXNIP/NLRP3 Interaction and Influencing Gut Microbiome Composition

 ,
,  ,
,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results
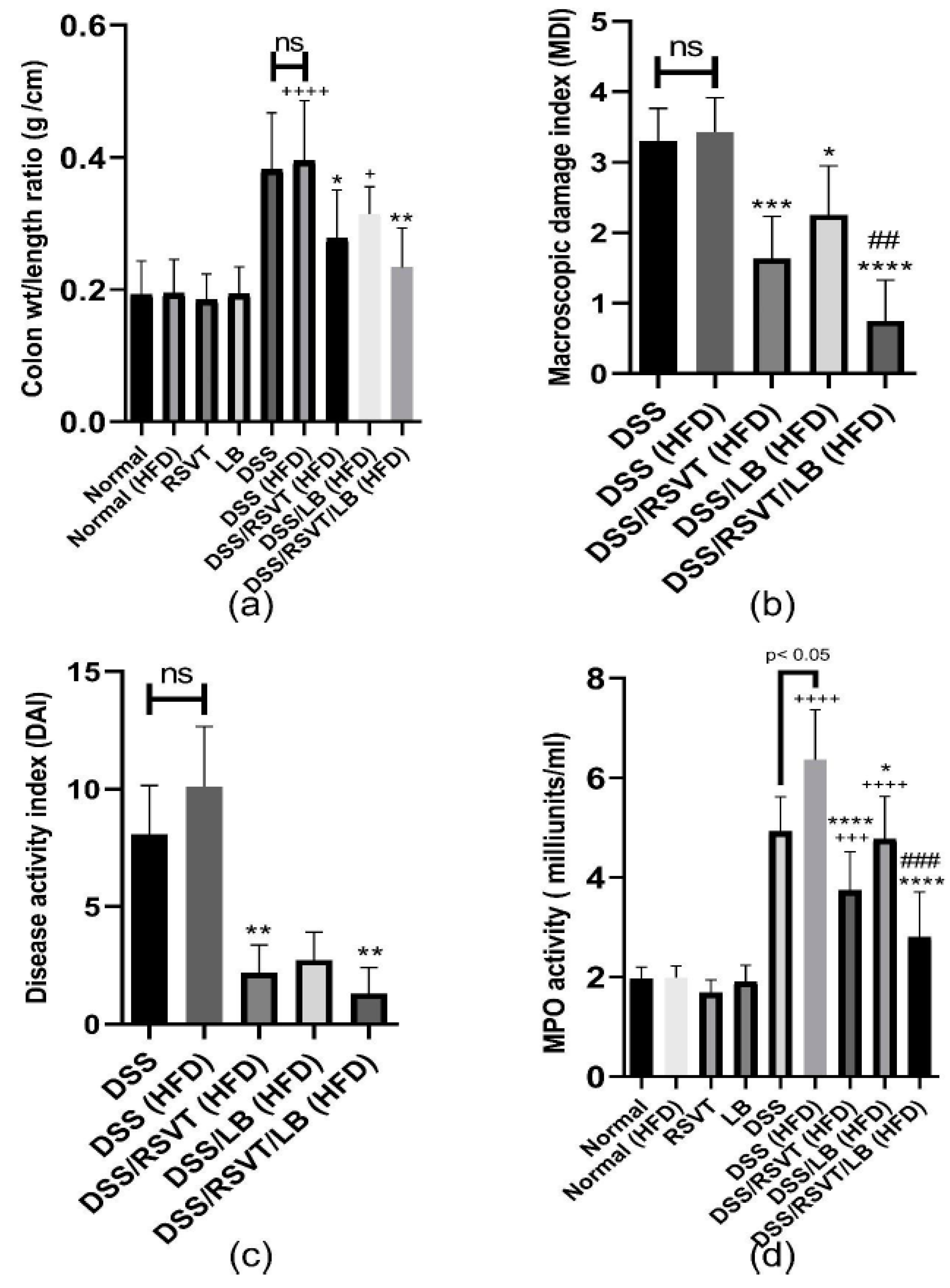
2.1. Effect of RSVT and LB on the Colon Weight/length Ratio, MDI, DAI, and Colonic Myeloperoxidase Activity
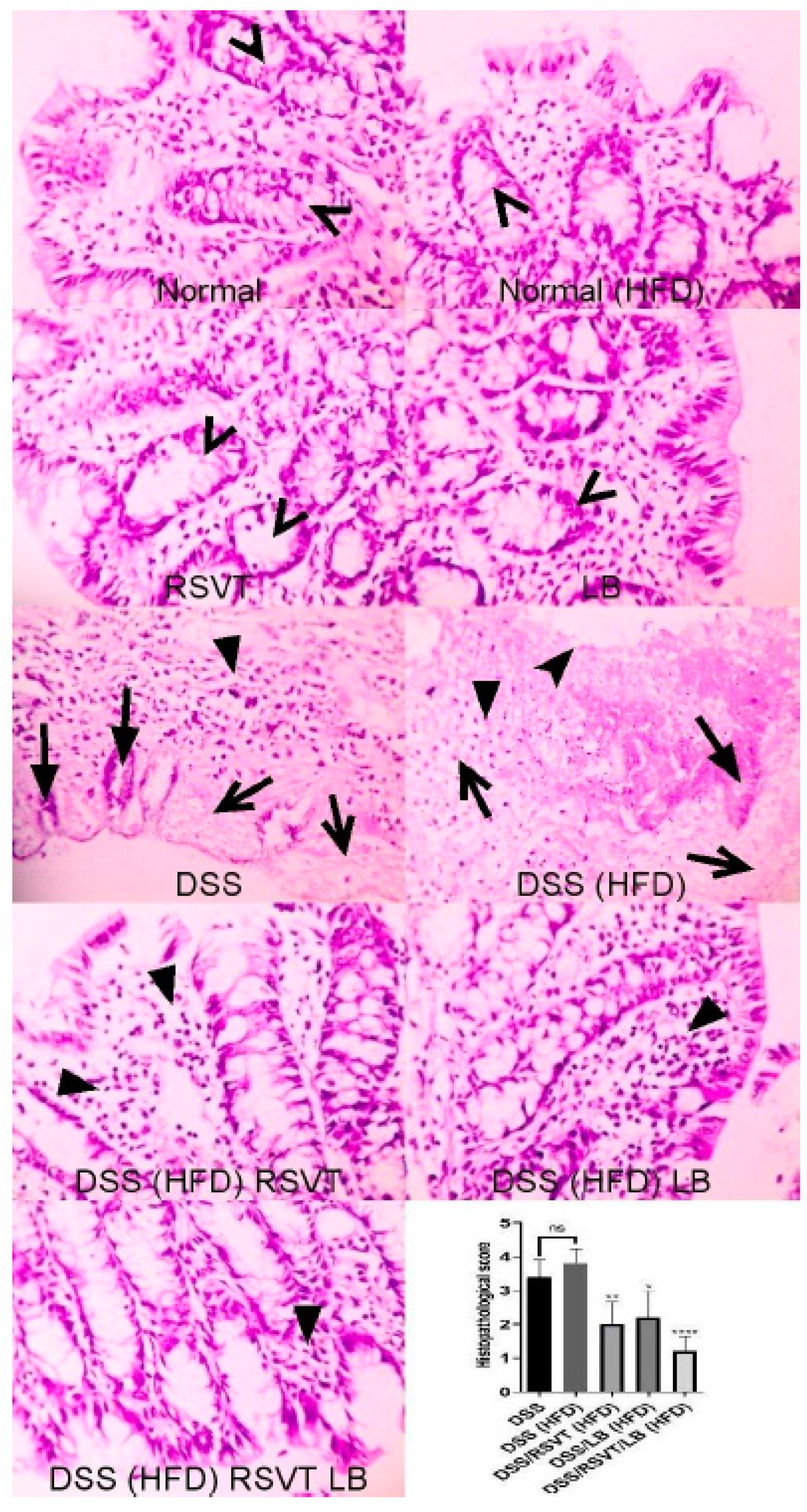
2.2. Histopathological Examination
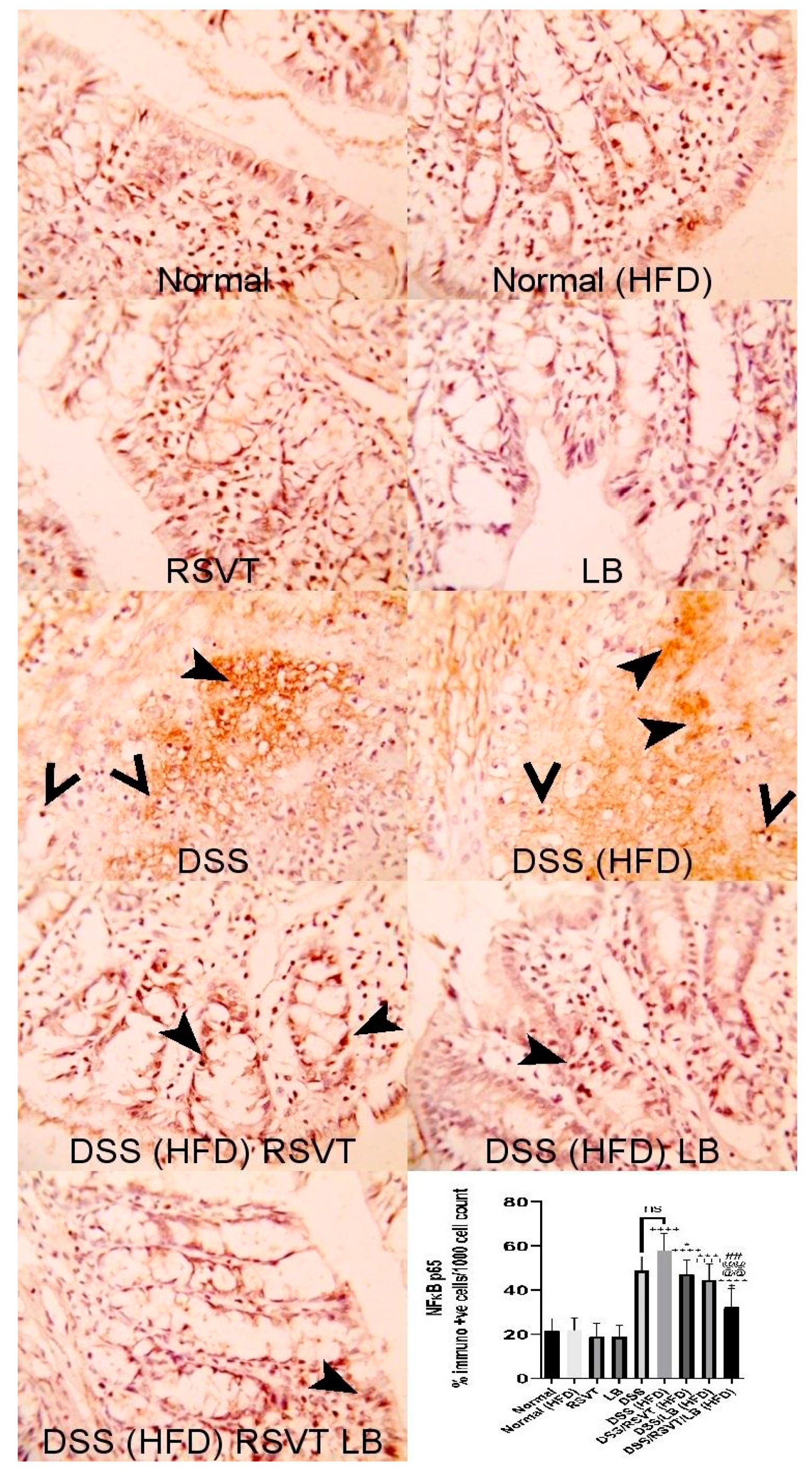
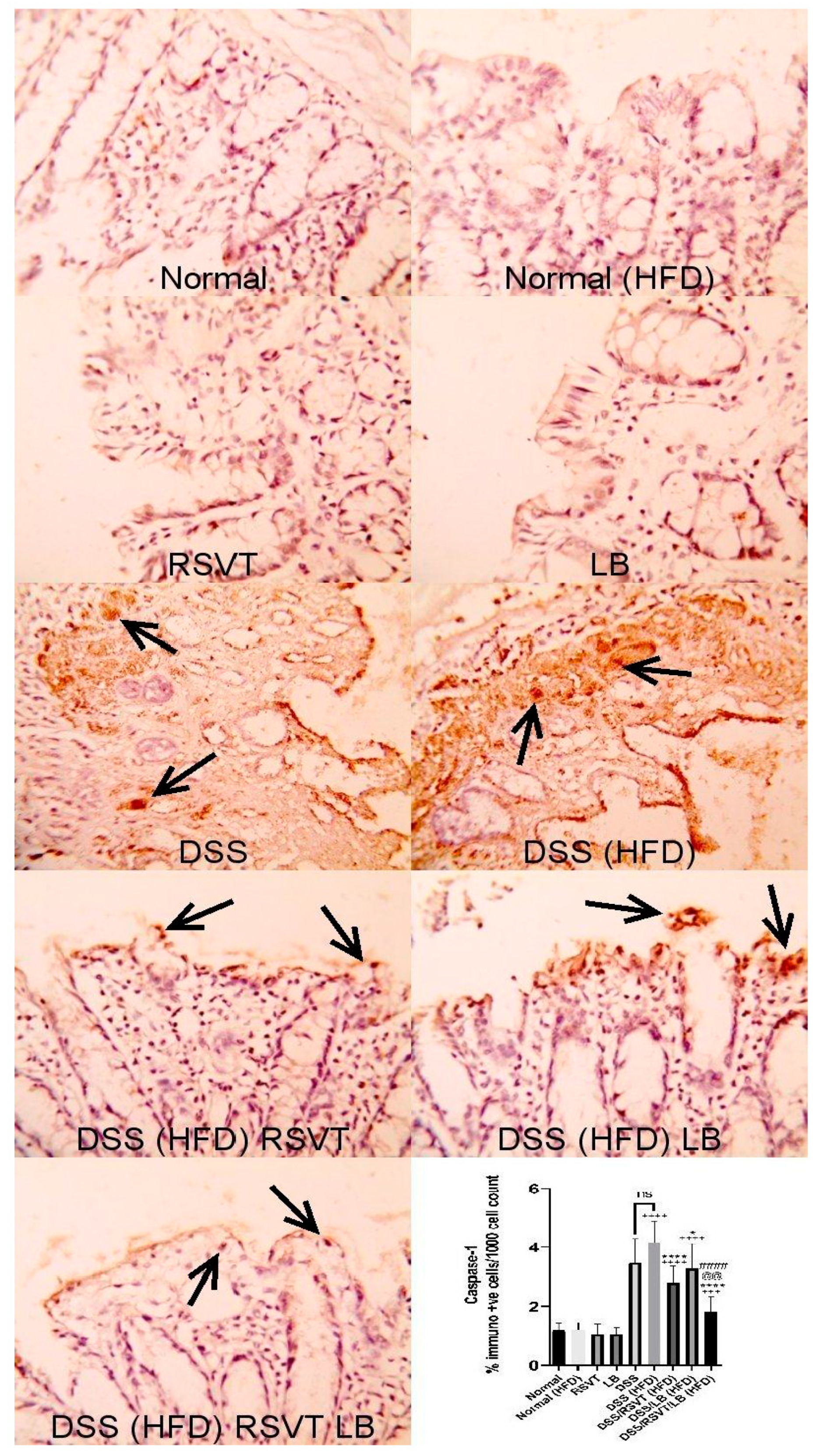
2.3. Effect of RSVT and LB on NF-ĸB p65 and Caspase-1 Immunolabeling
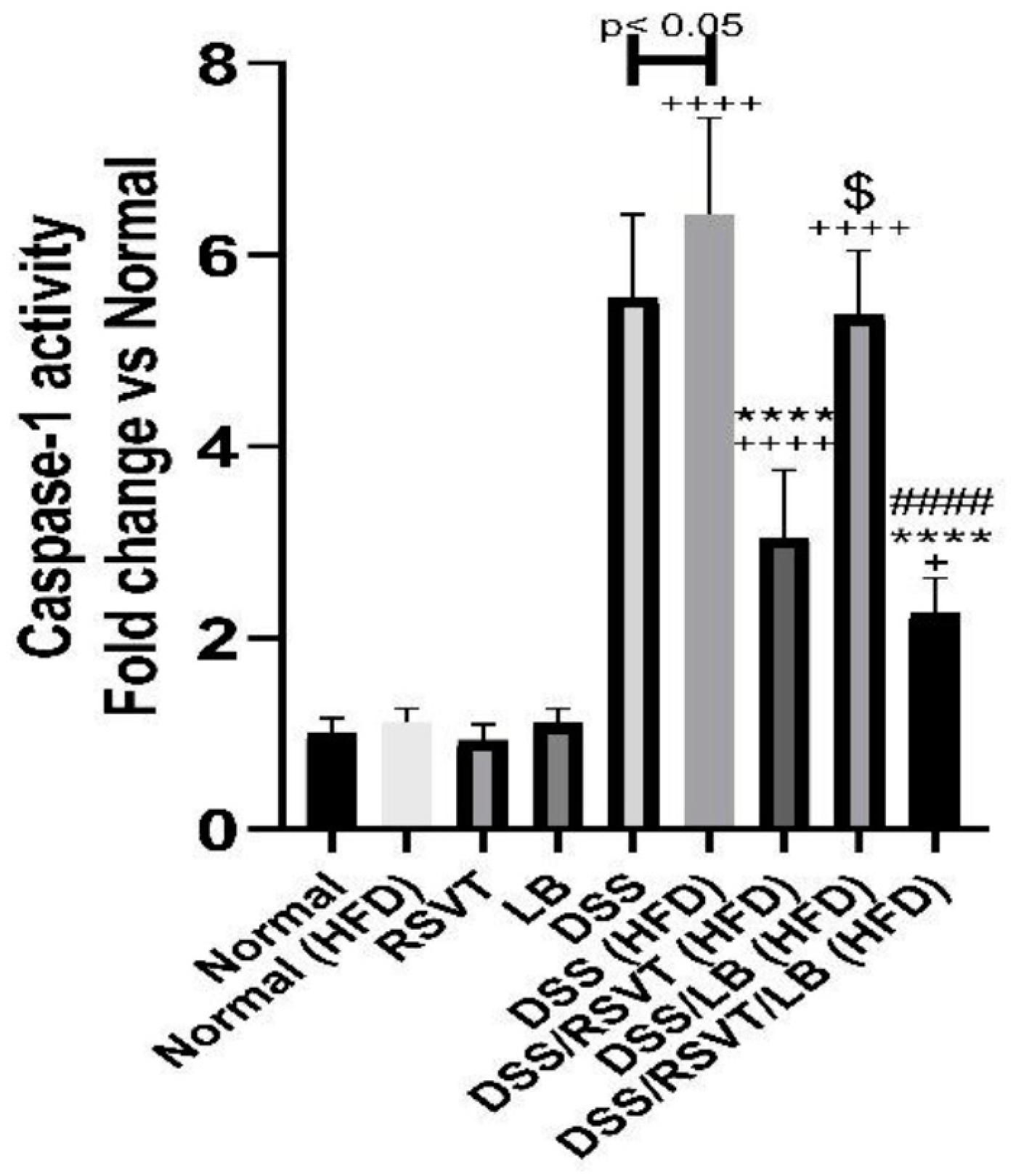
2.4. Effect of RSVT and LB on Caspase-1 Activity
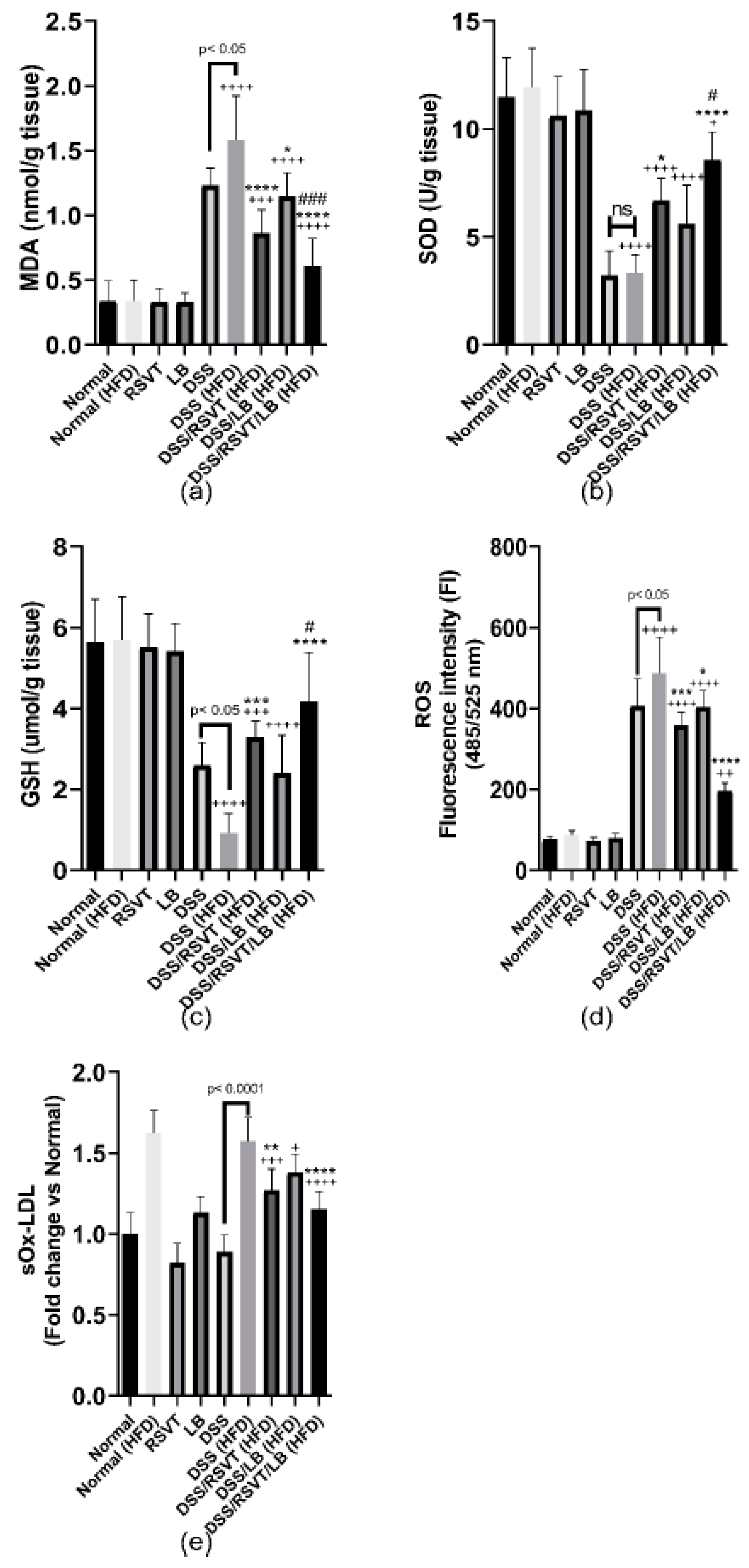
2.5. Effect of RSVT and LB on MDA, SOD, GSH, ROS, and sOx-LDL
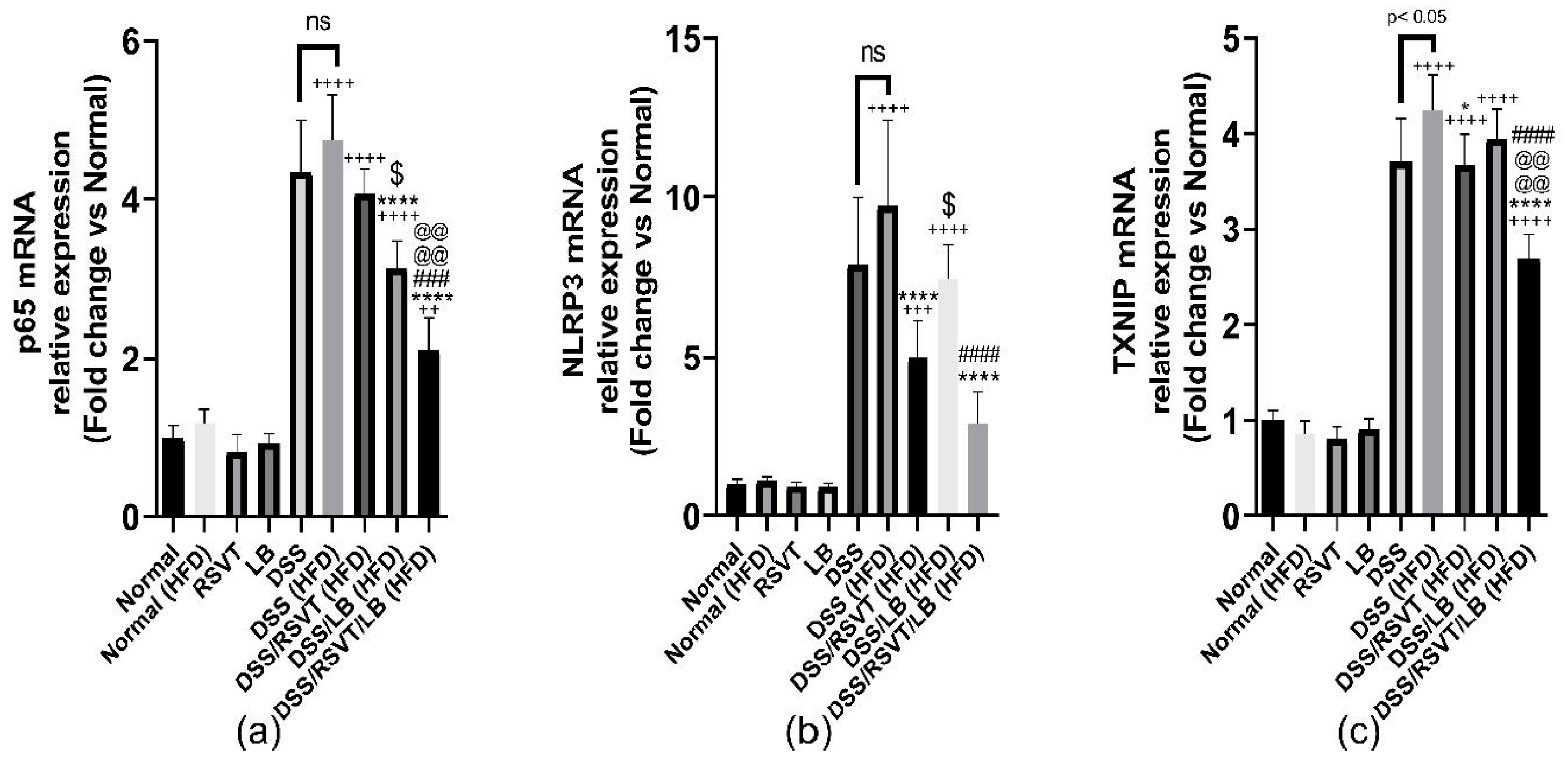
2.6. Effect of RSVT and LB on NF-κB p65, NLRP3, and TXNIP mRNA Expression
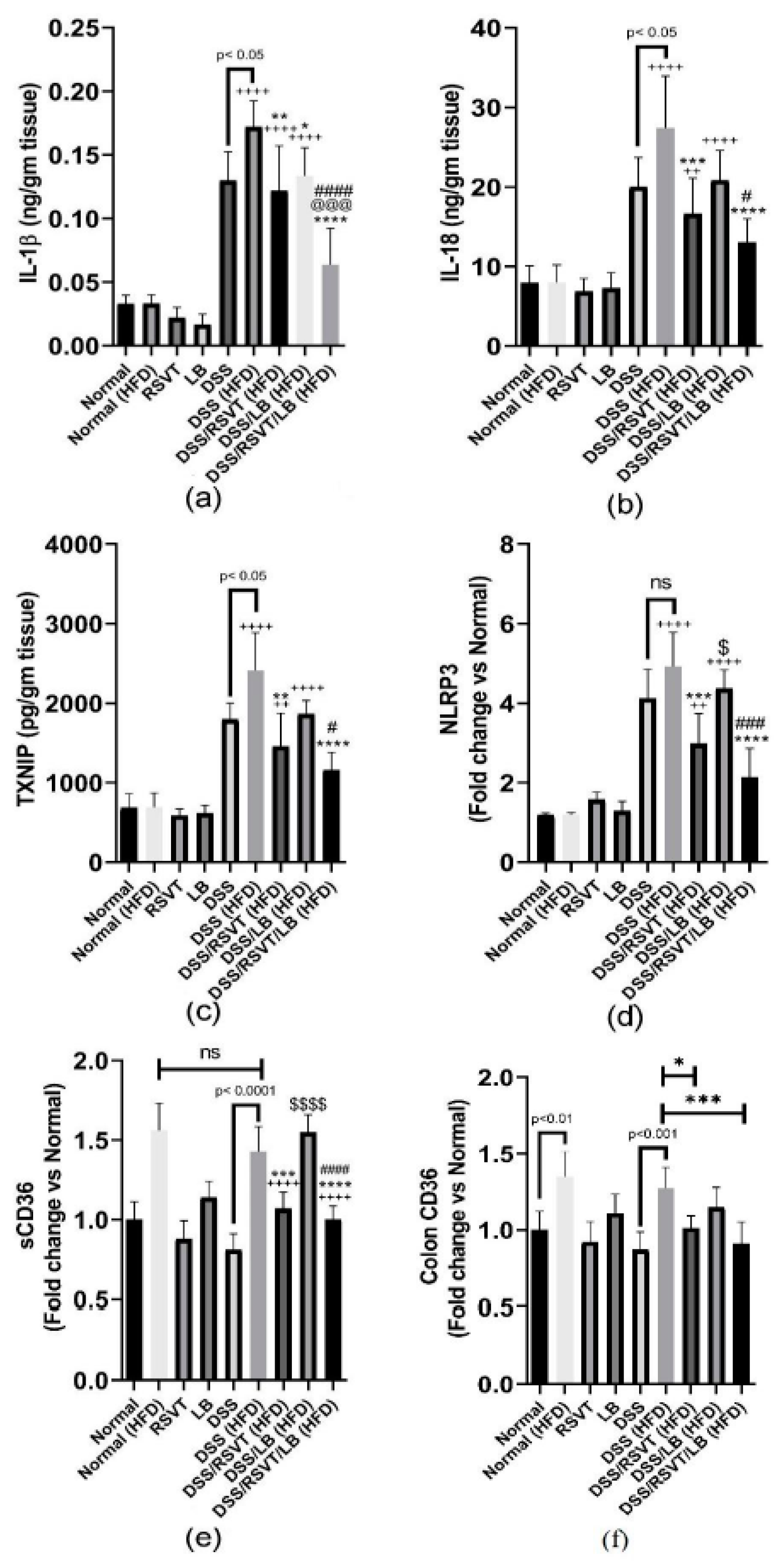
2.7. Effect of RSVT and LB on the Inflammatory Mediators TNF-α, IL-6, IL-10/IL-12 p70 Ratio, NF-κB p-p65/p65 Ratio, IL-1β, IL-18, TXNIP, NLRP3, sCD36, Colon CD36, and Ox-LDL
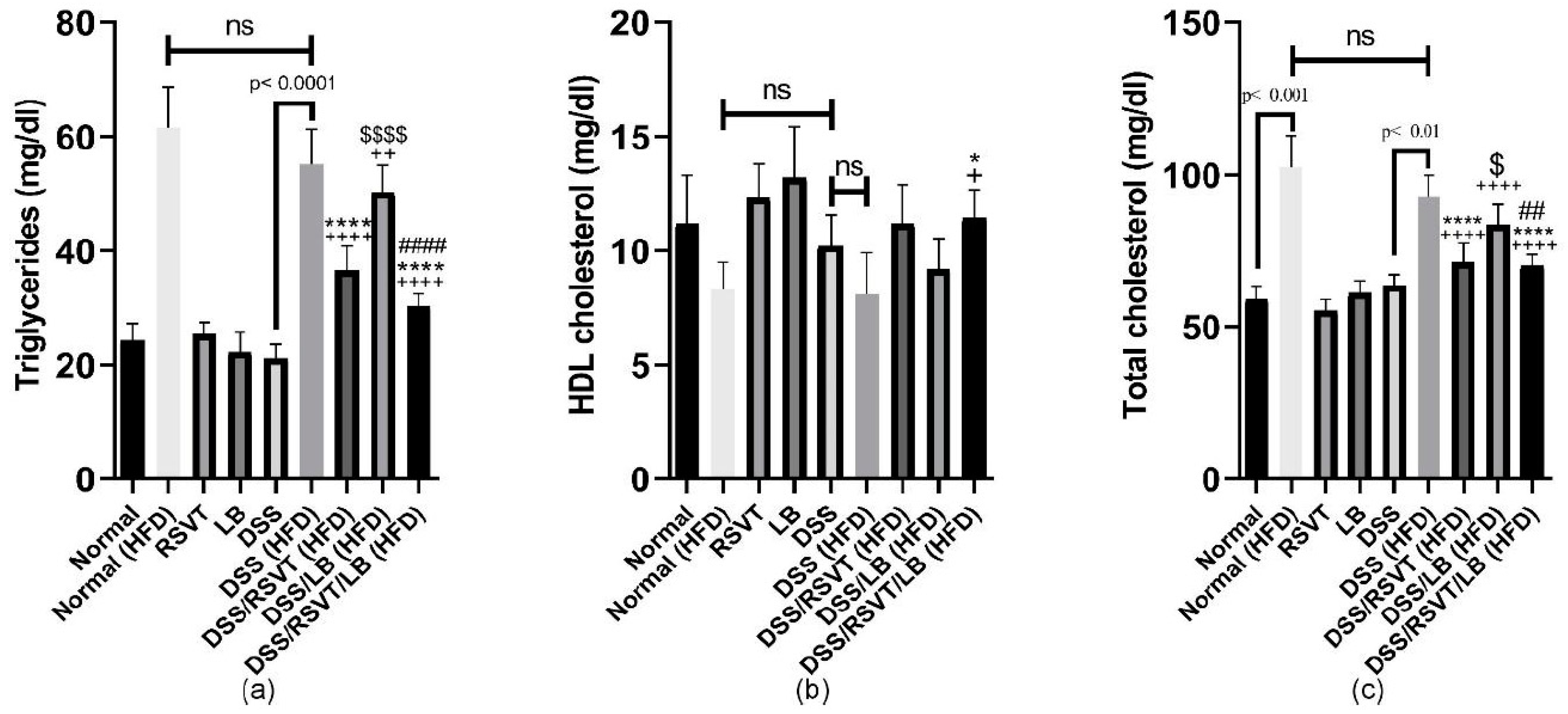
2.8. Effect of RSVT and LB on Lipid Profiles
2.9. Effect of RSVT and LB on Microbiome Composition as Determined by Conventional PCR Analysis
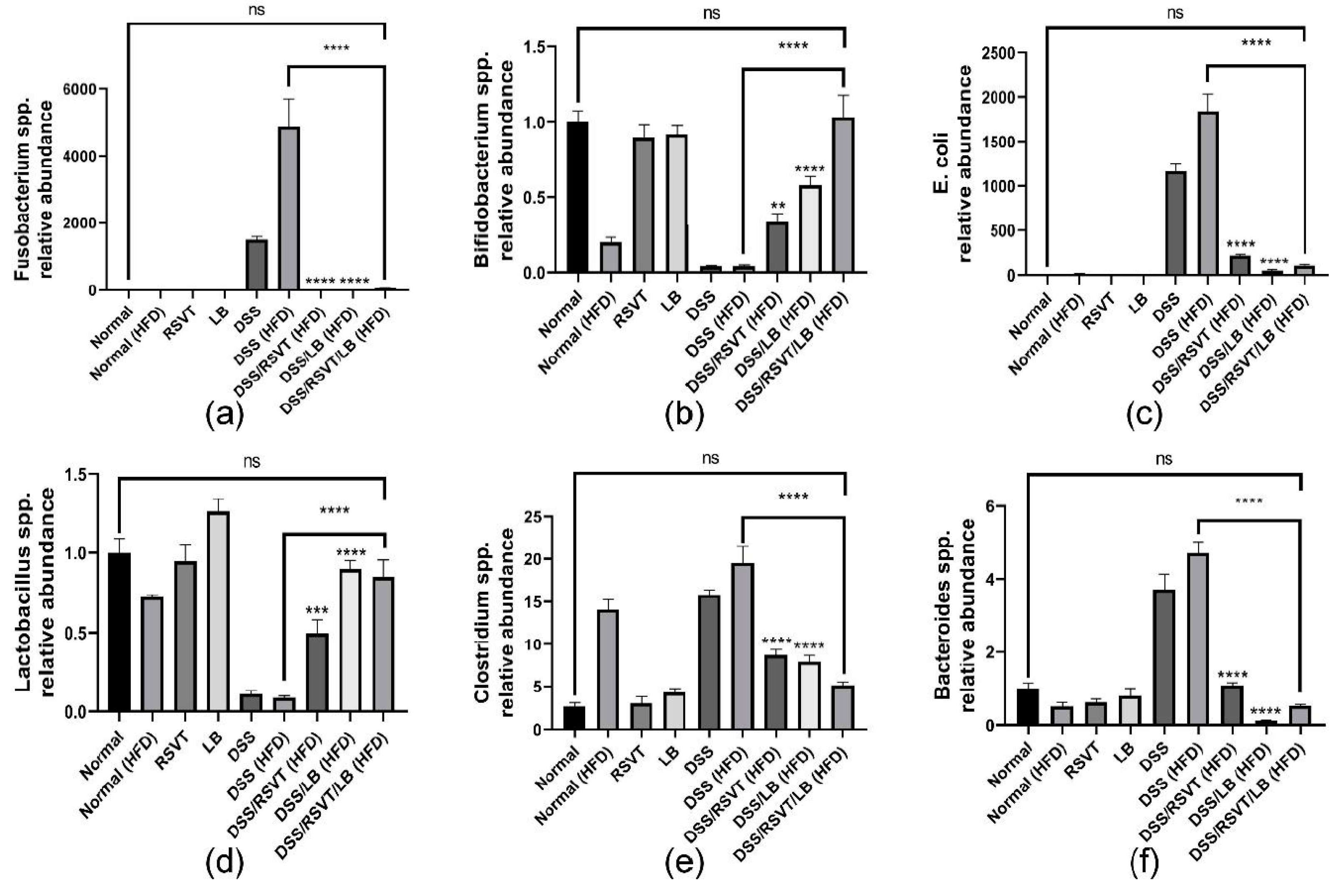
2.10. Effect of RSVT and LB on the Relative Abundance of Fusobacteria, Bifidobacteria, E. coli, Lactobacillus, Clostridium, and Bacteroides as Determined by qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Experimental Study and Treatment Protocol
4.1.1. Animals
4.1.2. HFD Composition
4.1.3. Preparation of Lactobacillus Suspension
4.1.4. Experimental Design
4.1.5. Sample Collection and Preparation
4.2. Assessment of Disease Activity Index and Macroscopic Damage Index
4.3. Histological Examination and Immunohistochemical Labeling of NF-кB p65 and Caspase-1
4.4. Biochemical Analysis
4.4.1. Assessment of Colonic Myeloperoxidase Activity
4.4.2. Assessment of Oxidative Stress Markers and Lipid Profile
4.4.3. Assessment of Caspase-1 Activity
4.4.4. Assessment of IL-1β, IL-6, IL-18, TNF-α, IL-10, IL-12 p70, TXNIP, CD36, and Ox-LDL by ELISA
4.4.5. Quantitative Real-Time PCR for the Expression of NLPR3, TXNIP, and NF-кB P65 in Colon Tissue
4.4.6. Conventional PCR for the Detection of Gut Microbiota
4.4.7. Quantitative Real-Time PCR for the Detection of the Relative Abundance of Fusobacteria, Bifidobacteria, E. coli, Lactobacillus, Clostridium, and Bacteroides
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Saber, S.; Khalil, R.M.; Abdo, W.S.; Nassif, D.; El-Ahwany, E. Olmesartan ameliorates chemically-induced ulcerative colitis in rats via modulating NFκB and Nrf-2/HO-1 signaling crosstalk. Toxicol. Appl. Pharmacol. 2019, 364, 120–132. [Google Scholar] [CrossRef]
- Saber, S.; Youssef, M.E.; Sharaf, H.; Amin, N.A.; El-Shedody, R.; Aboutouk, F.H.; El-Galeel, Y.A.; El-Hefnawy, A.; Shabaka, D.; Khalifa, A.; et al. BBG enhances OLT1177-induced NLRP3 inflammasome inactivation by targeting P2X7R/NLRP3 and MyD88/NF-κB signaling in DSS-induced colitis in rats. Life Sci. 2021, 270, 119123. [Google Scholar] [CrossRef]
- Kayal, M.; Shah, S. Ulcerative Colitis: Current and Emerging Treatment Strategies. J. Clin. Med. 2019, 9, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartor, R.B. Mechanisms of Disease: Pathogenesis of Crohn’s disease and ulcerative colitis. Nat. Clin. Pract. Gastroenterol. Hepatol. 2006, 3, 390–407. [Google Scholar] [CrossRef] [PubMed]
- Tourkochristou, E.; Aggeletopoulou, I.; Konstantakis, C.; Triantos, C. Role of NLRP3 inflammasome in inflammatory bowel diseases. World J. Gastroenterol. 2019, 25, 4796–4804. [Google Scholar] [CrossRef]
- Zhen, Y.; Zhang, H. NLRP3 Inflammasome and Inflammatory Bowel Disease. Front. Immunol. 2019, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.D.; Abreu, M.T. Diet as a Trigger or Therapy for Inflammatory Bowel Diseases. Gastroenterology 2017, 152, 398–414.e6. [Google Scholar] [CrossRef]
- Chatauret, N.; Favreau, F.; Giraud, S.; Thierry, A.; Rossard, L.; Le Pape, S.; Lerman, L.O.; Hauet, T. Diet-induced increase in plasma oxidized LDL promotes early fibrosis in a renal porcine auto-transplantation model. J. Transl. Med. 2014, 12, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, P.C.; Reigstad, C.S.; Loftus, E.V. Role of diet and gut microbiota in management of inflammatory bowel disease in an Asian migrant. J. Allergy Clin. Immunol. 2013, 132, 250.e5. [Google Scholar] [CrossRef]
- Albenberg, L.G.; Lewis, J.D.; Wu, G.D. Food and the gut microbiota in inflammatory bowel diseases. Curr. Opin. Gastroenterol. 2012, 28, 314–320. [Google Scholar] [CrossRef] [Green Version]
- Gulhane, M.; Murray, L.; Lourie, R.; Tong, H.; Sheng, Y.H.; Wang, R.; Kang, A.; Schreiber, V.; Wong, K.Y.; Magor, G.; et al. High Fat Diets Induce Colonic Epithelial Cell Stress and Inflammation that is Reversed by IL-22. Sci. Rep. 2016, 6, 28990. [Google Scholar] [CrossRef]
- Guo, X.Y.; Liu, X.J.; Hao, J.Y. Gut microbiota in ulcerative colitis: Insights on pathogenesis and treatment. J. Dig. Dis. 2020, 21, 147–159. [Google Scholar] [CrossRef]
- Saber, S. Angiotensin II: A key mediator in the development of liver fibrosis and cancer. Bull. Natl. Res. Cent. 2018, 42, 18. [Google Scholar] [CrossRef]
- Saber, S.; Ghanim, A.M.H.; El-Ahwany, E.; El-Kader, E.M.A. Novel complementary antitumour effects of celastrol and metformin by targeting IκBκB, apoptosis and NLRP3 inflammasome activation in diethylnitrosamine-induced murine hepatocarcinogenesis. Cancer Chemother. Pharmacol. 2020, 85, 331–343. [Google Scholar] [CrossRef]
- Younis, N.S.; Ghanim, A.M.H.; Saber, S. Mebendazole augments sensitivity to sorafenib by targeting MAPK and BCL-2 signalling in n-nitrosodiethylamine-induced murine hepatocellular carcinoma. Sci. Rep. 2019, 9, 19095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Wu, G.D.; Albenberg, L.; Tomov, V.T. Gut microbiota and IBD: Causation or correlation? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 573–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, M.A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. BioMed Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.J.; Sartor, R.B. Therapeutic Manipulation of the Microbiome in IBD: Current Results and Future Approaches. Curr. Treat. Options Gastroenterol. 2015, 13, 105–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.K.; Cho, J.H.; Kim, E.J.; Kim, E.-K.; Park, D.K.; Kwon, K.A.; Chung, J.-W.; Kim, K.O.; Kim, Y.J. Anti-inflammatory and anti-apoptotic effects of rosuvastatin by regulation of oxidative stress in a dextran sulfate sodium-induced colitis model. World J. Gastroenterol. 2017, 23, 4559–4568. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, X.; Wang, H.; Yang, Z.; Li, J.; Suo, H. Prevent Effects of Lactobacillus Fermentum HY01 on Dextran Sulfate Sodium-Induced Colitis in Mice. Nutrients 2017, 9, 545. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, R.; Chang, H.L.; Cote-Daigneaut, J.; Mehandru, S.; Atreja, A.; Colombel, J.-F. Statins Associated with Decreased Risk of New Onset Inflammatory Bowel Disease. Am. J. Gastroenterol. 2016, 111, 1416–1423. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, L.; Li, C.; Wu, H.; Ran, D.; Zhang, Z. Sulforaphane alter the microbiota and mitigate colitis severity on mice ulcerative colitis induced by DSS. AMB Express 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- El-Baz, A.M.; Khodir, A.E.; El-Sokkary, M.M.A.; Shata, A. The protective effect of Lactobacillus versus 5-aminosalicylic acid in ulcerative colitis model by modulation of gut microbiota and Nrf2/Ho-1 pathway. Life Sci. 2020, 256, 117927. [Google Scholar] [CrossRef] [PubMed]
- Requena, T.; Martinez-Cuesta, M.C.; Peláez, C. Diet and microbiota linked in health and disease. Food Funct. 2018, 9, 688–704. [Google Scholar] [CrossRef] [Green Version]
- Wagatsuma, K.; Nakase, H. Contradictory Effects of NLRP3 Inflammasome Regulatory Mechanisms in Colitis. Int. J. Mol. Sci. 2020, 21, 8145. [Google Scholar] [CrossRef]
- Saber, S.; El-Kader, E.M.A. Novel complementary coloprotective effects of metformin and MCC950 by modulating HSP90/NLRP3 interaction and inducing autophagy in rats. Inflammopharmacology 2020. [Google Scholar] [CrossRef]
- Saber, S.; El-Kader, E.M.A.; Sharaf, H.; El-Shamy, R.; El-Saeed, B.; Mostafa, A.; Ezzat, D.; Shata, A. Celastrol augments sensitivity of NLRP3 to CP-456773 by modulating HSP-90 and inducing autophagy in dextran sodium sulphate-induced colitis in rats. Toxicol. Appl. Pharmacol. 2020, 400, 115075. [Google Scholar] [CrossRef]
- Palmer, N.P.; Silvester, J.A.; Lee, J.J.; Beam, A.L.; Fried, I.; Valtchinov, V.I.; Rahimov, F.; Kong, S.W.; Ghodoussipour, S.; Hood, H.C.; et al. Concordance between gene expression in peripheral whole blood and colonic tissue in children with inflammatory bowel disease. PLoS ONE 2019, 14, e0222952. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat. Immunol. 2010, 11, 136–140. [Google Scholar] [CrossRef]
- Nasoohi, S.; Ismael, S.; Ishrat, T. Thioredoxin-Interacting Protein (TXNIP) in Cerebrovascular and Neurodegenerative Diseases: Regulation and Implication. Mol. Neurobiol. 2018, 55, 7900–7920. [Google Scholar] [CrossRef] [PubMed]
- Lara-Guzmán, O.J.; Gil-Izquierdo, Á.; Medina, S.; Osorio, E.; Álvarez-Quintero, R.; Zuluaga, N.; Oger, C.; Galano, J.-M.; Durand, T.; Muñoz-Durango, K. Oxidized LDL triggers changes in oxidative stress and inflammatory biomarkers in human macrophages. Redox Biol. 2018, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nyandwi, J.B.; Ko, Y.S.; Jin, H.; Yun, S.P.; Park, S.W.; Kim, H.J. Rosmarinic acid inhibits oxLDL-induced inflammasome activation under high-glucose conditions through downregulating the p38-FOXO1-TXNIP pathway. Biochem. Pharmacol. 2020, 182, 114246. [Google Scholar] [CrossRef] [PubMed]
- Cherfane, C.E.; Gessel, L.; Cirillo, D.; Zimmerman, M.B.; Polyak, S. Monocytosis and a Low Lymphocyte to Monocyte Ratio Are Effective Biomarkers of Ulcerative Colitis Disease Activity. Inflamm. Bowel Dis. 2015, 21, 1769–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Arellano, L.E.; Muñoz-Valle, J.F.; De La Cruz-Mosso, U.; Salgado-Bernabé, A.B.; Castro-Alarcón, N.; Parra-Rojas, I. Circulating CD36 and oxLDL levels are associated with cardiovascular risk factors in young subjects. BMC Cardiovasc. Disord. 2014, 14, 54. [Google Scholar] [CrossRef]
- Zhao, L.; Varghese, Z.; Moorhead, J.F.; Chen, Y.; Ruan, X.Z. CD36 and lipid metabolism in the evolution of atherosclerosis. Br. Med. Bull. 2018, 126, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Sheedy, F.J.; Grebe, A.; Rayner, K.J.; Kalantari, P.; Ramkhelawon, B.; Carpenter, S.B.; Becker, C.E.; Ediriweera, H.N.; Mullick, A.E.; Golenbock, D.T.; et al. CD36 coordinates NLRP3 inflammasome activation by facilitating intracellular nucleation of soluble ligands into particulate ligands in sterile inflammation. Nat. Immunol. 2013, 14, 812–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gizawy, S.A.; Nouh, A.; Saber, S.; Kira, A.Y. Deferoxamine-loaded transfersomes accelerates healing of pressure ulcers in streptozotocin-induced diabetic rats. J. Drug Deliv. Sci. Technol. 2020, 58, 101732. [Google Scholar] [CrossRef]
- Khalil, R.; Shata, A.; El-Kader, E.M.A.; Sharaf, H.; Abdo, W.S.; Amin, N.A.; Saber, S. Vildagliptin, a DPP-4 inhibitor, attenuates carbon tetrachloride-induced liver fibrosis by targeting ERK1/2, p38α, and NF-κB signaling. Toxicol. Appl. Pharmacol. 2020, 407, 115246. [Google Scholar] [CrossRef]
- Saber, S.; Goda, R.; El-Tanbouly, G.S.; Ezzat, D. Lisinopril inhibits nuclear transcription factor kappa B and augments sensitivity to silymarin in experimental liver fibrosis. Int. Immunopharmacol. 2018, 64, 340–349. [Google Scholar] [CrossRef]
- Saber, S.; Mahmoud, A.; Helal, N.; El-Ahwany, E.; Abdelghany, R. Liver Protective Effects of Renin-Angiotensin System Inhibition Have No Survival Benefits in Hepatocellular Carcinoma Induced by Repetitive Administration of Diethylnitrosamine in Mice. Open Access Maced. J. Med. Sci. 2018, 6, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, S.; Dudeja, P.K.; Tobacman, J.K. ROS, Hsp27, and IKKβ mediate dextran sodium sulfate (DSS) activation of IκBa, NFκB, and IL-8. Inflamm. Bowel Dis. 2009, 15, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, E.; Vykhovanets, E.V.; Vykhovanets, O.V.; MacLennan, G.T.; Singh, R.; Bhaskaran, N.; Shukla, S.; Gupta, S. High-fat diet activates pro-inflammatory response in the prostate through association of Stat-3 and NF-κB. Prostate 2012, 72, 233–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rheinheimer, J.; de Souza, B.M.; Cardoso, N.S.; Bauer, A.C.; Crispim, D. Current role of the NLRP3 inflammasome on obesity and insulin resistance: A systematic review. Metab. Clin. Exp. 2017, 74, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.-H.; Zhu, C.-X.; Quan, Y.-S.; Yang, Z.-Y.; Wu, S.; Luo, W.-W.; Tan, B.; Wang, X.-Y. Relationship between intestinal microbiota and ulcerative colitis: Mechanisms and clinical application of probiotics and fecal microbiota transplantation. World J. Gastroenterol. 2018, 24, 5–14. [Google Scholar] [CrossRef]
- Schierová, D.; Březina, J.; Mrázek, J.; Fliegerová, K.O.; Kvasnová, S.; Bajer, L.; Drastich, P.J.C. Gut Microbiome Changes in Patients with Active Left-Sided Ulcerative Colitis after Fecal Microbiome Transplantation and Topical 5-aminosalicylic Acid Therapy. Cells 2020, 9, 2283. [Google Scholar] [CrossRef] [PubMed]
- Leccese, G.; Bibi, A.; Mazza, S.; Facciotti, F.; Caprioli, F.; Landini, P.; Paroni, M.J.C. Probiotic Lactobacillus and Bifidobacterium Strains Counteract Adherent-Invasive Escherichia coli (AIEC) Virulence and Hamper IL-23/Th17 Axis in Ulcerative Colitis, but Not in Crohn’s Disease. Cells 2020, 9, 1824. [Google Scholar] [CrossRef]
- Jensen, S.R.; Mirsepasi-Lauridsen, H.C.; Thysen, A.H.; Brynskov, J.; Krogfelt, K.A.; Petersen, A.M.; Pedersen, A.E.; Brix, S. Distinct inflammatory and cytopathic characteristics of Escherichia coli isolates from inflammatory bowel disease patients. Int. J. Med. Microbiol. 2015, 305, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Ohkusa, T.; Sato, N.; Ogihara, T.; Morita, K.; Ogawa, M.; Okayasu, I. Fusobacterium varium localized in the colonic mucosa of patients with ulcerative colitis stimulates species-specific antibody. J. Gastroenterol. Hepatol. 2002, 17, 849–853. [Google Scholar] [CrossRef]
- Singh, H.; Nugent, Z.; Yu, B.N.; Lix, L.M.; Targownik, L.E.; Bernstein, C.N. Higher Incidence of Clostridium difficile Infection Among Individuals With Inflammatory Bowel Disease. Gastroenterology 2017, 153, 430–438.e2. [Google Scholar] [CrossRef]
- Keshk, W.A.; Ibrahim, M.A.; Shalaby, S.M.; Zalat, Z.A.; Elseady, W.S. Redox status, inflammation, necroptosis and inflammasome as indispensable contributors to high fat diet (HFD)-induced neurodegeneration; Effect of N-acetylcysteine (NAC). Arch. Biochem. Biophys. 2020, 680, 108227. [Google Scholar] [CrossRef]
- Garcia-Castillo, V.; Komatsu, R.; Clua, P.; Indo, Y.; Takagi, M.; Salva, S.; Islam, A.; Alvarez, S.; Takahashi, H.; Garcia-Cancino, A.; et al. Evaluation of the Immunomodulatory Activities of the Probiotic Strain Lactobacillus fermentum UCO-979C. Front. Immunol. 2019, 10, 1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fooladi, A.A.I.; Yazdi, M.H.; Pourmand, M.R.; Mirshafiey, A.; Hassan, Z.M.; Azizi, T.; Mahdavi, M.; Dallal, M.M.S. Th1 Cytokine Production Induced by Lactobacillus acidophilus in BALB/c Mice Bearing Transplanted Breast Tumor. Jundishapur J. Microbiol. 2015, 8, e17354. [Google Scholar] [CrossRef] [Green Version]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar] [PubMed]
- Saber, S.; Basuony, M.; Eldin, A.S. Telmisartan ameliorates dextran sodium sulfate-induced colitis in rats by modulating NF-κB signalling in the context of PPARγ agonistic activity. Arch. Biochem. Biophys. 2019, 671, 185–195. [Google Scholar] [CrossRef]
- Heidari, R.; Taheri, V.; Rahimi, H.R.; Yeganeh, B.S.; Niknahad, H.; Najibi, A. Sulfasalazine-induced renal injury in rats and the protective role of thiol-reductants. Ren. Fail. 2015, 38, 137–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.H.; Kwong, T.N.Y.; Chow, T.-C.; Luk, A.K.C.; Dai, R.Z.W.; Nakatsu, G.; Lam, T.Y.T.; Zhang, L.; Wu, J.C.Y.; Chan, F.K.L.; et al. Quantitation of faecalFusobacteriumimproves faecal immunochemical test in detecting advanced colorectal neoplasia. Gut 2016, 66, 1441–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Normal | Normal (HFD) | RSVT | LB | DSS | DSS (HFD) | DSS/RSVT (HFD) | DSS/LB (HFD) | DSS/RSVT/LB (HFD) |
|---|---|---|---|---|---|---|---|---|---|
| Enterococcus faecalis | − | − | − | − | − | − | − | − | − |
| Enterococcus faecium | − | − | − | − | − | − | − | − | − |
| Escherichia coli | + | + | + | + | + | + | + | + | + |
| Bifidobacterium spp. | + | + | + | + | + | + | + | + | + |
| Pseudomonasaeruginosa | − | + | − | − | + | + | − | − | − |
| Fusobacterium spp. | + | + | + | + | + | + | + | + | + |
| Providencia spp. | + | − | + | + | − | − | + | + | + |
| Prevotella intermedia | − | + | − | − | + | + | − | − | − |
| Peptostreptococcus magnus | − | − | − | − | − | − | − | − | − |
| Eubacterium saphenum | − | − | − | − | − | − | − | − | − |
| Porphyromonas gingivalis | − | + | − | − | + | + | − | − | − |
| Bacteroides spp. | + | + | + | + | + | + | + | + | + |
| Clostridium spp. | + | + | + | + | + | + | + | + | + |
| Lactobacillus spp. | + | + | + | + | + | + | + | + | + |
| Fusobacterium nucleatum | − | − | − | − | − | − | − | − | − |
| Exp. Groups | Day 1–Day 2 | Day 3–Day 9 | Day 10–Day 16 |
|---|---|---|---|
| Normal (n = 6) | ND | ND | ND |
| Normal (HFD) (n = 6) | HFD | HFD | HFD |
| RSVT (n = 6) | RSVT (20 mg/kg/day, p.o.) + ND | RSVT (20 mg/kg/day, p.o.) + ND | RSVT (20 mg/kg/day, p.o.) + ND |
| LB (n = 6) | LB (0.5 mL, p.o.) + ND | LB (0.5 mL, p.o.) + ND | LB (0.5 mL, p.o.) + ND |
| DSS (n = 10) | ND | 4% DSS in drinking water + ND | ND |
| DSS (HFD) (n = 10) | HFD | 4% DSS in drinking water + HFD | HFD |
| DSS/RSVT (HFD) (n = 8) | RSVT (20 mg/kg/day, p.o.) + HFD | RSVT (20 mg/kg/day, p.o.) + 4% DSS in drinking water + HFD | RSVT (20 mg/kg/day, p.o.) + HFD |
| DSS/LB (HFD) (n = 8) | LB (0.5 mL, p.o.) + HFD | LB (0.5 mL, p.o.) + 4% DSS in drinking water + HFD | LB (0.5 mL, p.o.) + HFD |
| DSS/RSVT/LB (HFD) (n = 8) | RSVT (20 mg/kg/day, p.o.) + LB (0.5 mL, p.o.) + HFD | RSVT (20 mg/kg/day, p.o.) + LB (0.5 mL, p.o.) + 4% DSS in drinking water + HFD | RSVT (20 mg/kg/day, p.o.) + LB (0.5 mL, p.o.) + HFD |
| Primer | GenBank Accession | F | R | Ta (°C) | Amplicon Size (bp) |
|---|---|---|---|---|---|
| TXNIP | NM_001008767.1 | 5′-AAGCTGTCCTCAGTCAGAGGCAAT-3′ | 5′-ATGACTTTCTTGGAGCCAGGGACA-3′ | 64 | 170 |
| NLRP3 | NM_001191642.1 | 5′-GAGCTGGACCTCAGTGACAATGC-3′ | 5′-ACCAATGCGAGATCCTGACAACAC-3′ | 63 | 146 |
| NFκB p65 | NM_199267.2 | 5′-TTCCCTGAAGTGGAGCTAGGA-3′ | 5′-CATGTCGAGGAAGACACTGGA-3′ | 61 | 185 |
| GAPDH | NM_017008.4 | 5′-TCAAGAAGGTGGTGAAGCAG-3′ | 5′-AGGTGGAAGAATGGGAGTTG-3′ | 57 | 111 |
| Primer Name | Forward and Reverse (5’ to 3’) | Primer Sequence | Ta (°C) | Ampilicon Size (bp) |
|---|---|---|---|---|
| (16S) | F | GAGTTTGATCCTGGCTCAG | 51 | 312 |
| R | GCTGCCTCCCGTAGGAGT | |||
| Porphyromonas gingivalis | F | AATCGTAACGGGCGACACAC | 53 | 594 |
| R | GGGTTGCTCCTTCATCACAC | |||
| E. saphenum | F | AACCACATAAAATCATAGG | 43 | 828 |
| R | ATACCCGATTAAGGGTAC | |||
| Fusobacterium | F | GGATTTATTGGGCGTAAAGC | 51.5 | 162 |
| R | GGCATTCCTACAAATATCTACGAA | |||
| E. coli | F | TGGGAGCGAAAATCCTG | 47.5 | 219 |
| R | CAGTACAGGTAGACTTCTG | |||
| Providencia | F | ACCGCATAATCTCTTAGG | 43.5 | 514 |
| R | CTACACATGGAATTCTAC | |||
| E. Faecium | F | GCAAGGCTTCTTAGAGA | 46.5 | 564 |
| R | CATCGTGTAAGCTAACTTC | |||
| Bifidobacterium | F | CTCCTGGAAACGGGTGG | 51 | 551 |
| R | GGTGTTCTTCCCGATATCTACA | |||
| Prevotella intermedia | F | CGAACCGTCAAGCATAGGC | 54 | 368 |
| R | AACAGCCGCTTTTAGAACACAA | |||
| Peptostreptococcus magnus | F | CGGGNTTTAGTAGACAGAAG | 50 | 565 |
| R | CAGTTTCCAATGCTTTACGG | |||
| P.aeruginosa | F | CGAGTACAACATGGCTCTGG | 53 | 116 |
| R | ACCGGACGCTCTTTACCATA | |||
| E. faecalis | F | ATCAAGTACAGTTAGTCTT | 44 | 940 |
| R | ACGATTCAAAGCTAACTG | |||
| Bacteroides spp. | F | AAGGGAGCGTAGATGGATGTTTA | 55 | 193 |
| R | CGAGCCTCAATGTCAGTTGC | |||
| Clostridium spp. | F | CGGTACCTGACTAAGAAGC | 50 | 429 |
| R | AGTTTGATTCTTGCGAACG | |||
| Lactobacillus spp. | F | AGCAGTAGGGAATCTTCCA | 50 | 334 |
| R | CACCGCTACACATGGAG | |||
| Fusobacterium nucleatum | F | CAACCATTACTTTAACTCTACCATGTTCA | 57 | 356 |
| R | GTTGACTTTACAGAAGGAGATTATGTAAAAATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saber, S.; Abd El-Fattah, E.E.; Yahya, G.; Gobba, N.A.; Maghmomeh, A.O.; Khodir, A.E.; Mourad, A.A.E.; Saad, A.S.; Mohammed, H.G.; Nouh, N.A.; et al. A Novel Combination Therapy Using Rosuvastatin and Lactobacillus Combats Dextran Sodium Sulfate-Induced Colitis in High-Fat Diet-Fed Rats by Targeting the TXNIP/NLRP3 Interaction and Influencing Gut Microbiome Composition. Pharmaceuticals 2021, 14, 341. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14040341
Saber S, Abd El-Fattah EE, Yahya G, Gobba NA, Maghmomeh AO, Khodir AE, Mourad AAE, Saad AS, Mohammed HG, Nouh NA, et al. A Novel Combination Therapy Using Rosuvastatin and Lactobacillus Combats Dextran Sodium Sulfate-Induced Colitis in High-Fat Diet-Fed Rats by Targeting the TXNIP/NLRP3 Interaction and Influencing Gut Microbiome Composition. Pharmaceuticals. 2021; 14(4):341. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14040341
Chicago/Turabian StyleSaber, Sameh, Eslam E. Abd El-Fattah, Galal Yahya, Naglaa A. Gobba, Abdalkareem Omar Maghmomeh, Ahmed E. Khodir, Ahmed A. E. Mourad, Ahmed S. Saad, Hager G. Mohammed, Nehal A. Nouh, and et al. 2021. "A Novel Combination Therapy Using Rosuvastatin and Lactobacillus Combats Dextran Sodium Sulfate-Induced Colitis in High-Fat Diet-Fed Rats by Targeting the TXNIP/NLRP3 Interaction and Influencing Gut Microbiome Composition" Pharmaceuticals 14, no. 4: 341. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14040341