
Beneficial Effects of Polydeoxyribonucleotide (PDRN) in an In Vitro Model of Fuchs Endothelial Corneal Dystrophy

 , , and
, , and
Abstract
:1. Introduction
2. Results
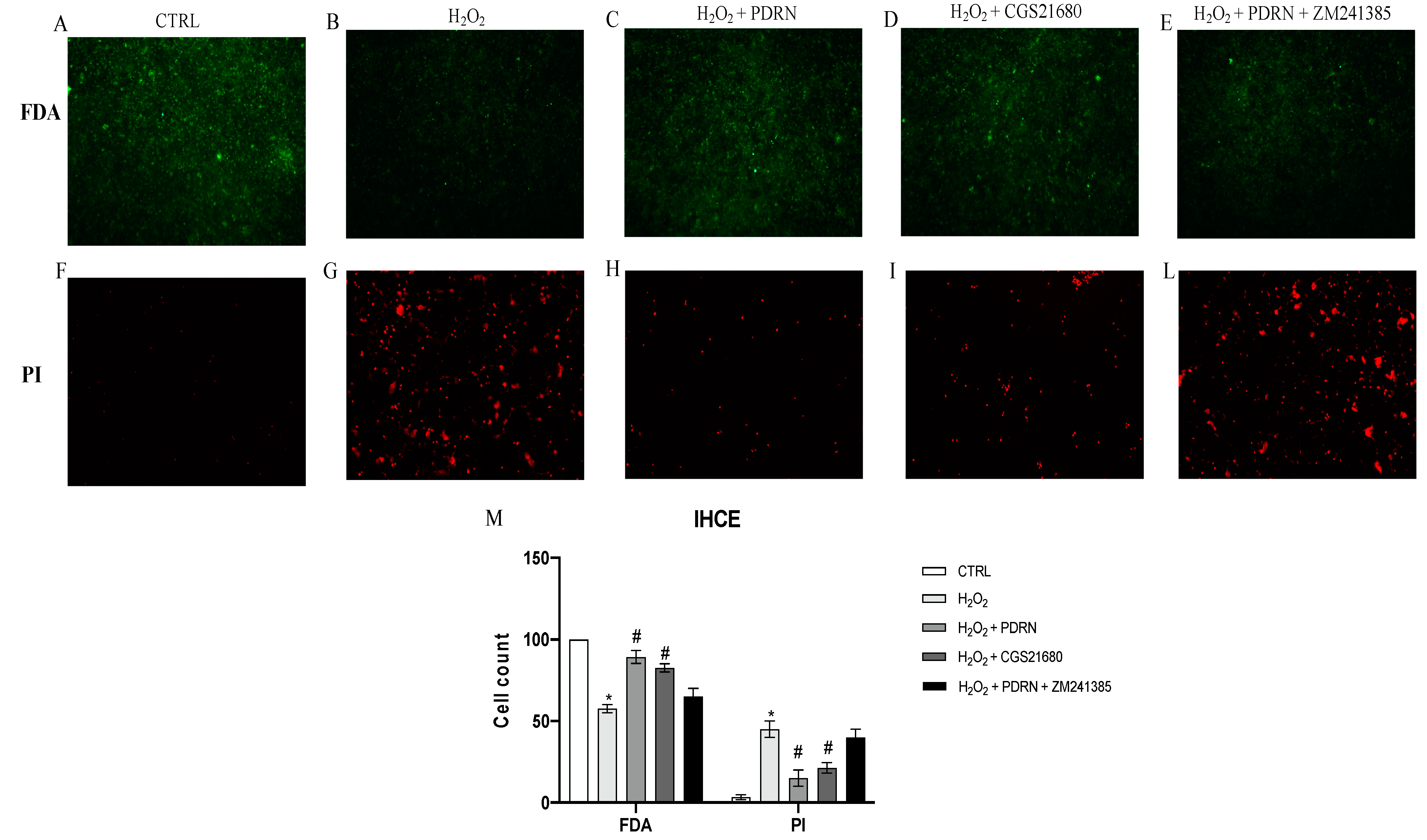
2.1. Effects of PDRN on Cell Viability
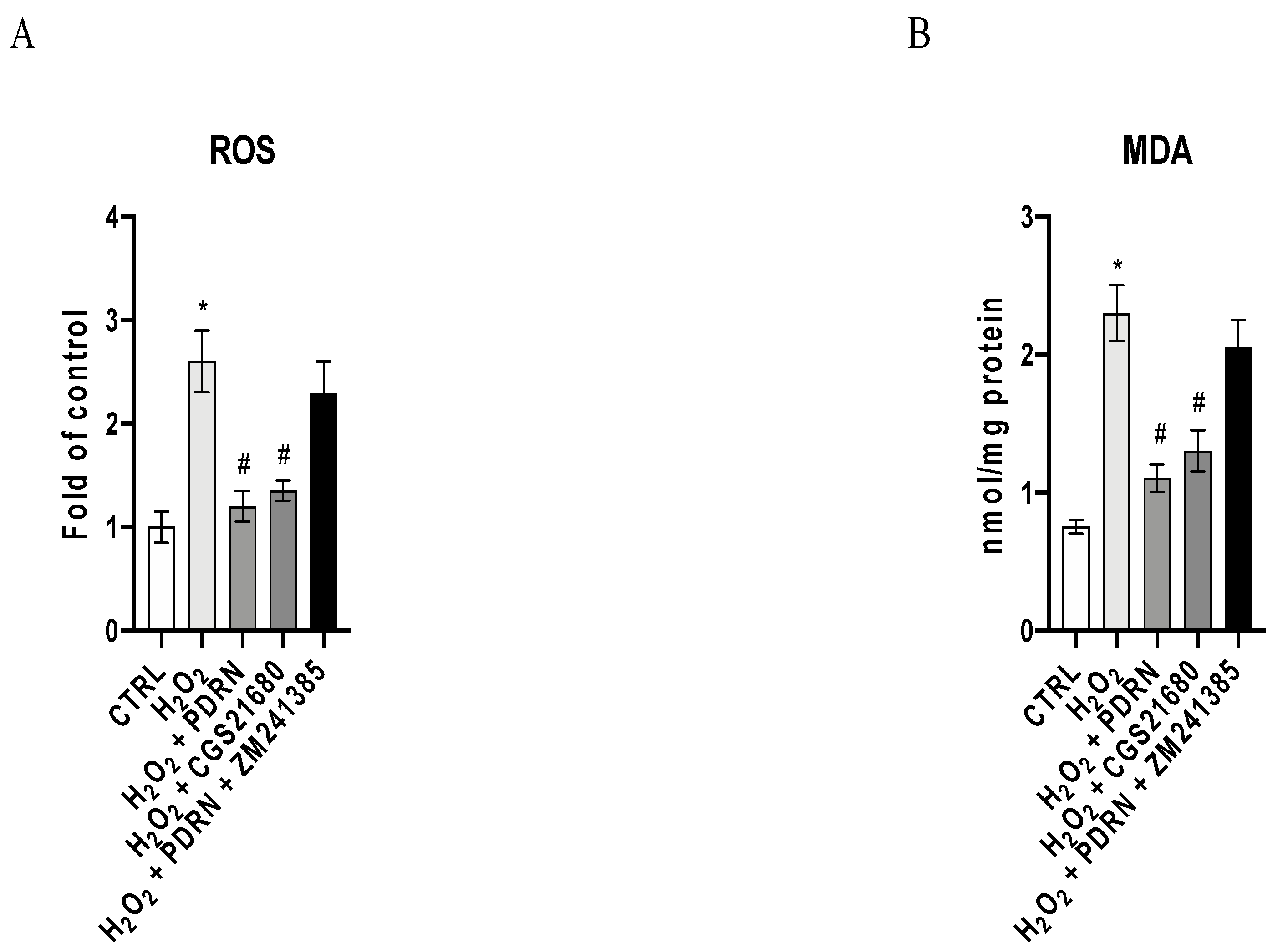
2.2. The Effects of PDRN on Oxidative Stress
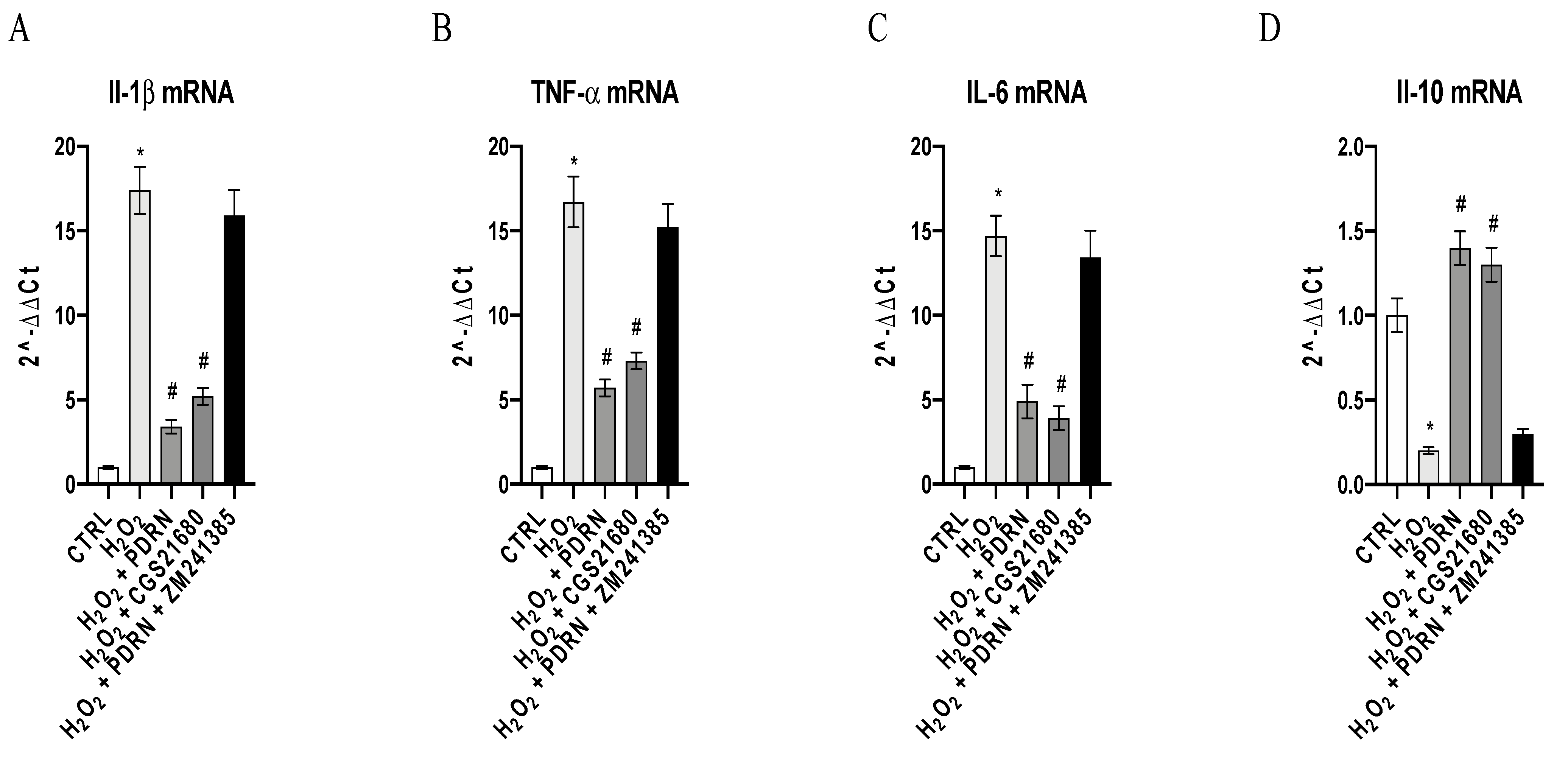
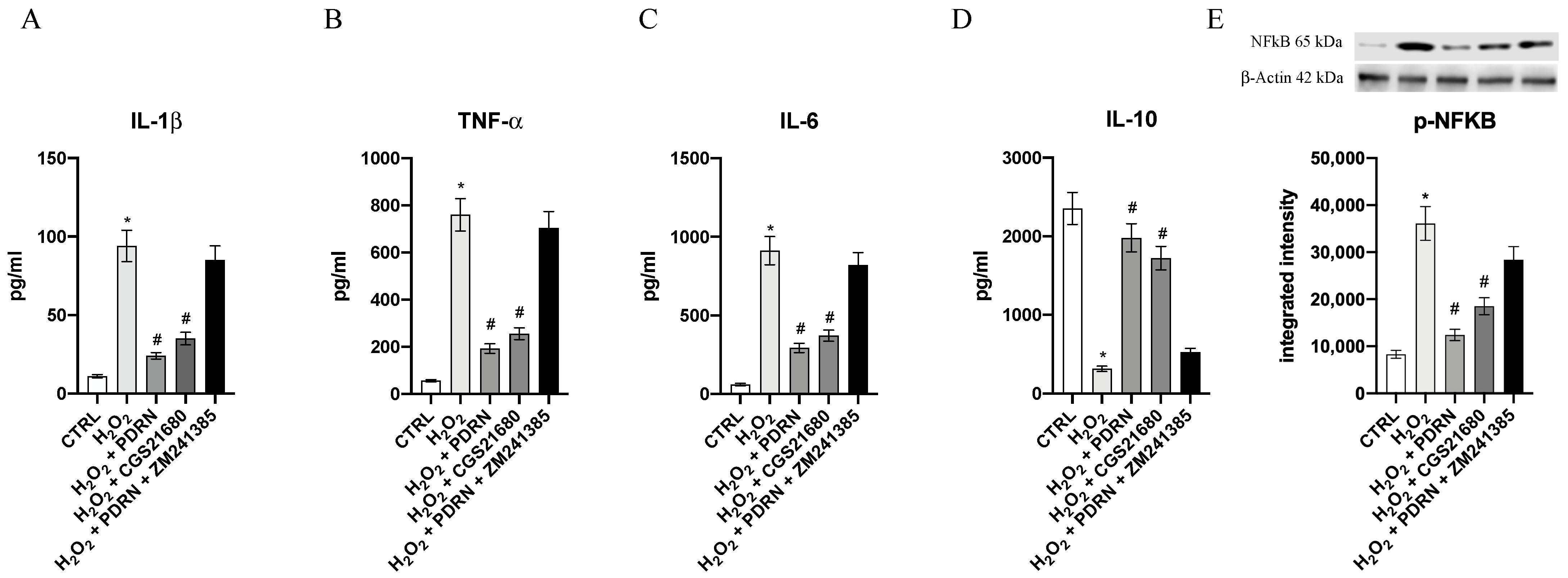
2.3. The Effects of PDRN on Inflammatory Markers
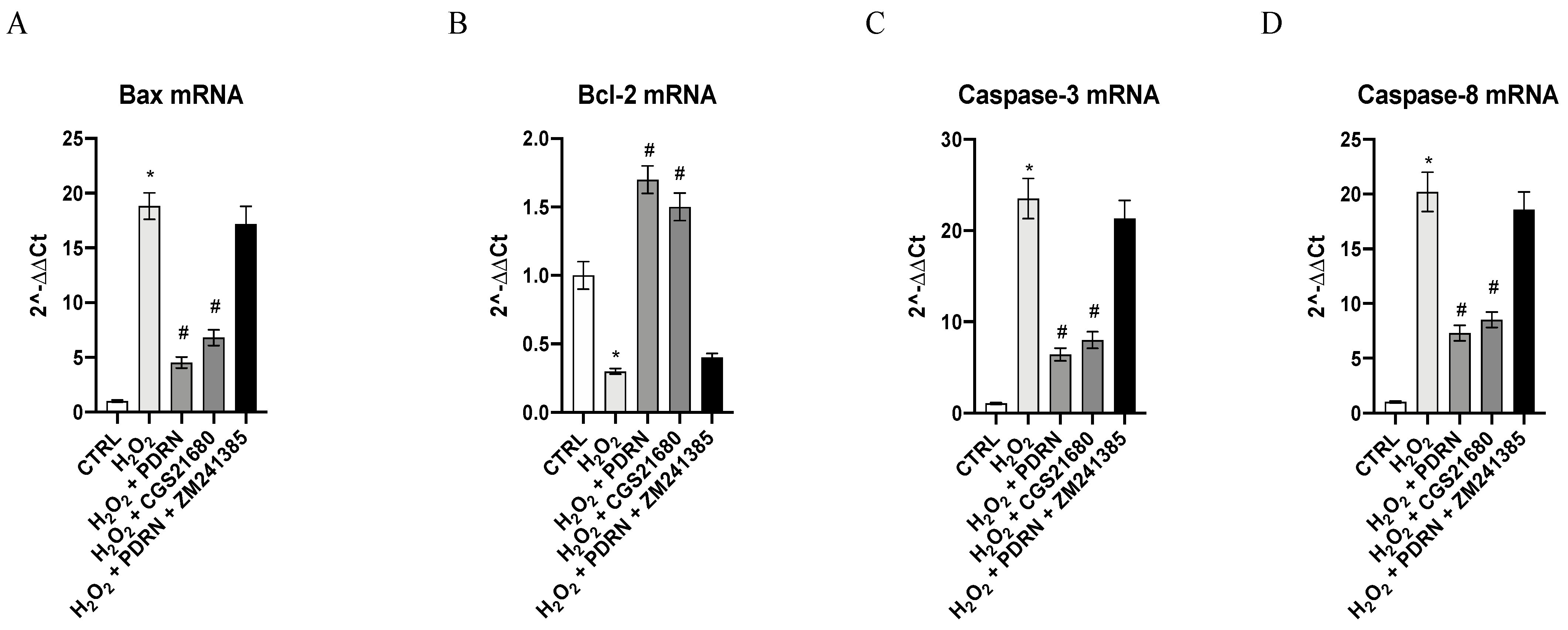
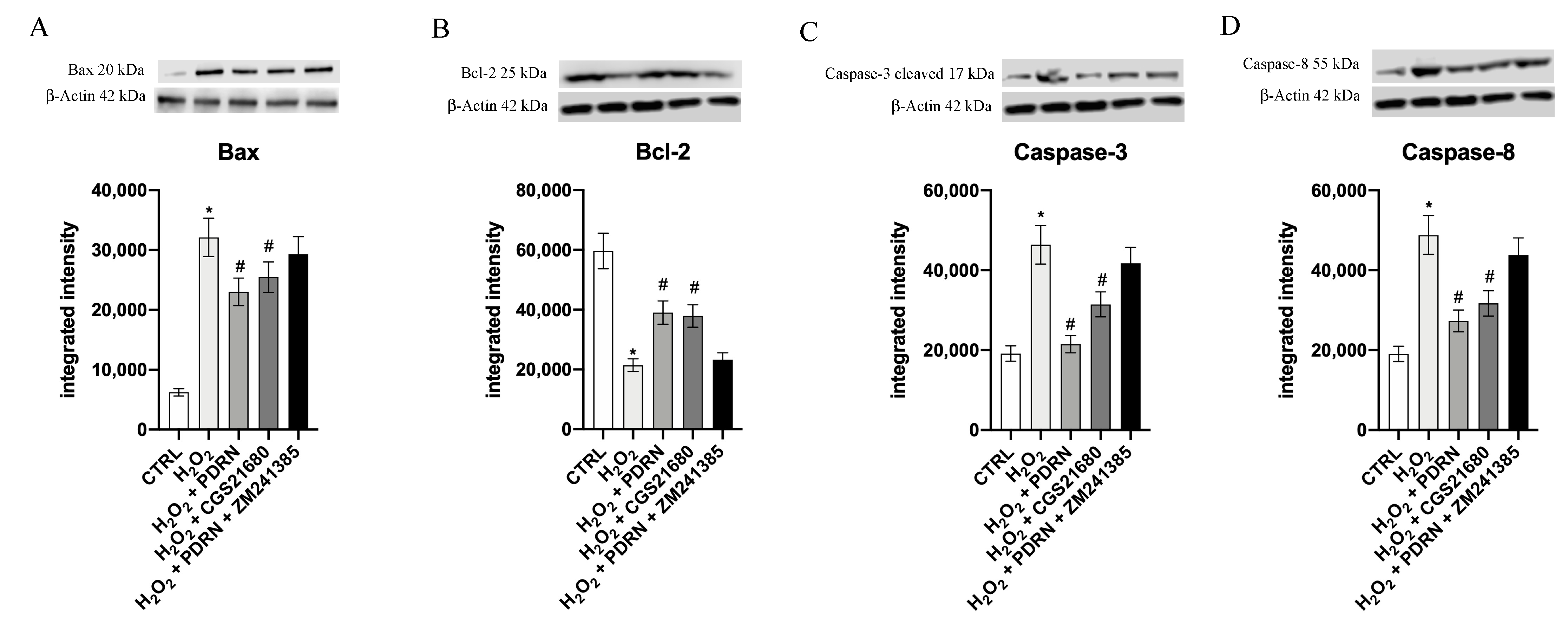
2.4. Effects of PDRN on Apoptosis
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Cell Treatments
4.3. FDA/PI Staining
4.4. ROS Measurement
4.5. Malondialdehyde Assay
4.6. Real-Time Quantitative PCR Amplification (RT-qPCR)
4.7. Measurements of the Cytokines
4.8. Western Blot Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elhalis, H.; Azizi, B.; Jurkunas, U.V. Fuchs endothelial corneal dystrophy. Ocul. Surf. 2010, 8, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Rolev, K.; Coussons, P.; King, L.; Rajan, M. Experimental models of corneal endothelial cell therapy and translational challenges to clinical practice. Exp. Eye Res. 2019, 188, 107794. [Google Scholar] [CrossRef]
- Matthaei, M.; Hribek, A.; Clahsen, T.; Bachmann, B.; Cursiefen, C.; Jun, A.S. Fuchs Endothelial Corneal Dystrophy: Clinical, Genetic, Pathophysiologic, and Therapeutic Aspects. Annu. Rev. Vis. Sci. 2019, 5, 151–175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Igo, R.P., Jr.; Fondran, J.; Mootha, V.V.; Oliva, M.; Hammersmith, K.; Sugar, A.; Lass, J.H.; Iyengar, S.K. Fuchs’ Genetics Multi-Center Study Group. Association of smoking and other risk factors with Fuchs’ endothelial corneal dystrophy severity and corneal thickness. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5829–5835. [Google Scholar] [CrossRef] [PubMed]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurkunas, U.V.; Bitar, M.S.; Funaki, T.; Azizi, B. Evidence of oxidative stress in the pathogenesis of fuchs endothelial corneal dystrophy. Am. J. Pathol. 2010, 177, 2278–2289. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.C.; Meng, H.; Jun, A.S. N-Acetylcysteine increases corneal endothelial cell survival in a mouse model of Fuchs endothelial corneal dystrophy. Exp. Eye Res. 2014, 127, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocaba, V.; Katikireddy, K.R.; Gipson, I.; Price, M.O.; Price, F.W.; Jurkunas, U.V. Association of the Gutta-Induced Microenvironment with Corneal Endothelial Cell Behavior and Demise in Fuchs Endothelial Corneal Dystrophy. JAMA Ophthalmol. 2018, 136, 886–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Zong, R.; Bao, X.; Zheng, X.; Cui, H.; Liu, Z.; Zhou, Y. Oxidative Stress Suppresses Cellular Autophagy in Corneal Epithelium. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3286–3293. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Zeng, Q.; Guo, Y.; Liu, S.; Wang, P.; Xie, M.; Chen, J. Pathological molecular mechanism of symptomatic late-onset Fuchs endothelial corneal dystrophy by bioinformatic analysis. PLoS ONE 2018, 13, e0197750. [Google Scholar] [CrossRef] [PubMed]
- Gomez, A.; Serrano, A.; Salero, E.; Tovar, A.; Amescua, G.; Galor, A.; Keane, R.W.; de Rivero Vaccari, J.P.; Sabater, A.L. Tumor necrosis factor-alpha and interferon-gamma induce inflammasome-mediated corneal endothelial cell death. Exp. Eye Res. 2021, 207, 108574. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, M.J.; Kweon, D.K.; Lim, S.T.; Lee, S.J. Polydeoxyribonucleotide Activates Mitochondrial Biogenesis but Reduces MMP-1 Activity and Melanin Biosynthesis in Cultured Skin Cells. Appl. Biochem. Biotechnol. 2020, 191, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Pallio, G.; Bitto, A.; Ieni, A.; Irrera, N.; Mannino, F.; Pallio, S.; Altavilla, D.; Squadrito, F.; Scarpignato, C.; Minutoli, L. Combined Treatment with Polynucleotides and Hyaluronic Acid Improves Tissue Repair in Experimental Colitis. Biomedicines 2020, 8, 438. [Google Scholar] [CrossRef]
- Muratore, O.; Cattarini, G.; Gianoglio, S.; Tonoli, E.L.; Saccà, S.C.; Ghiglione, D.; Venzano, D.; Ciurlo, C.; Lantieri, P.B.; Schito, G.C. A human placental polydeoxyribonucleotide (PDRN) may promote the growth of human corneal fibroblasts and iris pigment epithelial cells in primary culture. New Microbiol. 2003, 26, 13–26. [Google Scholar] [PubMed]
- Lazzarotto, M.; Tomasello, E.M.; Caporossi, A. Clinical evaluation of corneal epithelialization after photorefractive keratectomy in a patient treated with Polydeoxyribonucleotide (PDRN) eye drops: A randomized, double-blind, placebo-controlled trial. Eur. J. Ophthalmol. 2004, 14, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Picciolo, G.; Mannino, F.; Irrera, N.; Altavilla, D.; Minutoli, L.; Vaccaro, M.; Arcoraci, V.; Squadrito, V.; Picciolo, G.; Squadrito, F.; et al. PDRN, a natural bioactive compound, blunts inflammation and positively reprograms healing genes in an “in vitro” model of oral mucositis. Biomed. Pharmacother. 2021, 138, 111538. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, A. A2A Adenosine Receptor Agonists and their Potential Therapeutic Applications. An Update. Curr. Med. Chem. 2018, 25, 3597–3612. [Google Scholar] [CrossRef]
- Squadrito, F.; Bitto, A.; Irrera, N.; Pizzino, G.; Pallio, G.; Minutoli, L.; Altavilla, D. Pharmacological Activity and Clinical Use of PDRN. Front. Pharmacol. 2017, 8, 224. [Google Scholar] [CrossRef]
- Baek, A.; Kim, Y.; Lee, J.W.; Lee, S.C.; Cho, S.R. Effect of Polydeoxyribonucleotide on Angiogenesis and Wound Healing in an In Vitro Model of Osteoarthritis. Cell Transplant. 2018, 27, 1623–1633. [Google Scholar] [CrossRef] [Green Version]
- Pallio, G.; Bitto, A.; Pizzino, G.; Galfo, F.; Irrera, N.; Squadrito, F.; Squadrito, G.; Pallio, S.; Anastasi, G.P.; Cutroneo, G.; et al. Adenosine Receptor Stimulation by Polydeoxyribonucleotide Improves Tissue Repair and Symptomology in Experimental Colitis. Front. Pharmacol. 2016, 7, 273. [Google Scholar] [CrossRef] [Green Version]
- Beach, K.M.; Hung, L.F.; Arumugam, B.; Smith, E.L., 3rd; Ostrin, L.A. Adenosine receptor distribution in Rhesus monkey ocular tissue. Exp. Eye Res. 2018, 174, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Gain, P.; Jullienne, R.; He, Z.; Aldossary, M.; Acquart, S.; Cognasse, F.; Thuret, G. Global Survey of Corneal Transplantation and Eye Banking. JAMA Ophthalmol. 2016, 134, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.J.; Ashraf, M.F.; Shen, D.F.; Green, W.R.; Stark, W.J.; Chan, C.C.; O’Brien, T.P. The role of apoptosis in the pathogenesis of Fuchs endothelial dystrophy of the cornea. Arch. Ophthalmol. 2001, 119, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.K. Adenosine A2A receptors regulate oxidative stress formation in rat pheochromocytoma PC12 cells during serum deprivation. Neurosci. Lett. 2003, 350, 127–131. [Google Scholar] [CrossRef]
- Chiu, F.L.; Lin, J.T.; Chuang, C.Y.; Chien, T.; Chen, C.M.; Chen, K.H.; Hsiao, H.Y.; Lin, Y.S.; Chern, Y.; Kuo, H.C. Elucidating the role of the A2A adenosine receptor in neurodegeneration using neurons derived from Huntington’s disease iPSCs. Hum. Mol. Genet. 2015, 24, 6066–6079. [Google Scholar] [CrossRef] [Green Version]
- Hwang, L.; Jin, J.J.; Ko, I.G.; Kim, S.; Cho, Y.A.; Sung, J.S.; Choi, C.W.; Chang, B.S. Polydeoxyribonucleotide Attenuates Airway Inflammation through A2AR Signaling Pathway in PM10-Exposed Mice. Int. Neurourol. J. 2021, 25, 19–26. [Google Scholar] [CrossRef]
- Ko, I.G.; Jin, J.J.; Hwang, L.; Kim, S.H.; Kim, C.J.; Jeon, J.W.; Chung, J.Y.; Han, J.H. Adenosine A2A receptor agonist polydeoxyribonucleotide ameliorates short-term memory impairment by suppressing cerebral ischemia-induced inflammation via MAPK pathway. PLoS ONE 2021, 16, e0248689. [Google Scholar] [CrossRef]
- Ko, I.G.; Jin, J.J.; Hwang, L.; Kim, S.H.; Kim, C.J.; Won, K.Y.; Na, Y.G.; Kim, K.H.; Kim, S.J. Adenosine A2A Receptor Agonist Polydeoxyribonucleotide Alleviates Interstitial Cystitis-Induced Voiding Dysfunction by Suppressing Inflammation and Apoptosis in Rats. J. Inflamm. Res. 2021, 14, 367–378. [Google Scholar] [CrossRef]
- Lee, S.; Won, K.Y.; Joo, S. Protective Effect of Polydeoxyribonucleotide against CCl4-Induced Acute Liver Injury in Mice. Int. Neurourol. J. 2020, 24, 88–95. [Google Scholar] [CrossRef]
- Kim, S.E.; Ko, I.G.; Jin, J.J.; Hwang, L.; Kim, C.J.; Kim, S.H.; Han, J.H.; Jeon, J.W. Polydeoxyribonucleotide Exerts Therapeutic Effect by Increasing VEGF and Inhibiting Inflammatory Cytokines in Ischemic Colitis Rats. BioMed Res. Int. 2020, 2020, 2169083. [Google Scholar] [CrossRef]
- Ceravolo, I.; Mannino, F.; Irrera, N.; Squadrito, F.; Altavilla, D.; Ceravolo, G.; Pallio, G.; Minutoli, L. Health Potential of Aloe vera against Oxidative Stress Induced Corneal Damage: An “In Vitro” Study. Antioxidants 2021, 10, 318. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, S.; Miao, Q.; Jin, K.; Lou, L.; Ye, X.; Xi, Y.; Ye, J. Protective Role of Hinokitiol against H2O2-Induced Injury in Human Corneal Epithelium. Curr. Eye Res. 2017, 42, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Choi, J.H.; Cui, L.; Li, Y.; Yang, J.M.; Yun, J.J.; Jung, J.E.; Choi, W.; Yoon, K.C. Anti-Inflammatory and Antioxidative Effects of Camellia japonica on Human Corneal Epithelial Cells and Experimental Dry Eye: In Vivo and In Vitro Study. Investig. Ophthalmol. Vis Sci. 2017, 58, 1196–1207. [Google Scholar] [CrossRef] [Green Version]
- Varani, K.; Padovan, M.; Vincenzi, F.; Targa, M.; Trotta, F.; Govoni, M.; Borea, P.A. A2A and A3 adenosine receptor expression in rheumatoid arthritis: Upregulation, inverse correlation with disease activity score and suppression of inflammatory cytokine and metalloproteinase release. Arthritis Res. Ther. 2011, 13, R197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumdar, S.; Aggarwal, B. Adenosine suppresses activation of nuclear factor-κB selectively induced by tumor necrosis factor in different cell types. Oncogene 2003, 22, 1206–1218. [Google Scholar] [CrossRef] [PubMed]
- Irrera, N.; Bitto, A.; Vaccaro, M.; Mannino, F.; Squadrito, V.; Pallio, G.; Arcoraci, V.; Minutoli, L.; Ieni, A.; Lentini, M.; et al. PDRN, a Bioactive Natural Compound, Ameliorates Imiquimod-Induced Psoriasis through NF-κB Pathway Inhibition and Wnt/β-Catenin Signaling Modulation. Int. J. Mol. Sci. 2020, 21, 1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, I.G.; Jin, J.J.; Hwang, L.; Kim, S.H.; Kim, C.J.; Han, J.H.; Lee, S.; Kim, H.I.; Shin, H.P.; Jeon, J.W. Polydeoxyribonucleotide Exerts Protective Effect against CCl4-Induced Acute Liver Injury through Inactivation of NF-κB/MAPK Signaling Pathway in Mice. Int. J. Mol. Sci. 2020, 21, 7894. [Google Scholar] [CrossRef] [PubMed]
- Campo, G.M.; Micali, A.; Avenoso, A.; D’Ascola, A.; Scuruchi, M.; Pisani, A.; Bruschetta, A.; Calatroni, A.; Puzzolo, D.; Campo, S. Inhibition of small HA fragment activity and stimulation of A2A adenosine receptor pathway limit apoptosis and reduce cartilage damage in experimental arthritis. Histochem. Cell Biol. 2015, 143, 531–543. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Park, S.H.; Ko, I.G.; Jin, J.J.; Hwang, L.; Ji, E.S.; Kim, S.H.; Kim, C.J.; Park, S.Y.; Hwang, J.J.; et al. Polydeoxyribonucleotide Ameliorates Lipopolysaccharide-Induced Lung Injury by Inhibiting Apoptotic Cell Death in Rats. Int. J. Mol. Sci. 2017, 18, 1847. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Chung, J.Y. Effectiveness of polydeoxyribonucleotide injection versus normal saline injection for treatment of chronic plantar fasciitis: A prospective randomised clinical trial. Int. Orthop. 2015, 39, 1329–1334. [Google Scholar] [CrossRef]
- Lee, D.O.; Yoo, J.H.; Cho, H.I.; Cho, S.; Cho, H.R. Comparing effectiveness of polydeoxyribonucleotide injection and corticosteroid injection in plantar fasciitis treatment: A prospective randomized clinical study. Foot Ankle Surg. 2020, 26, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Cho, R.K.; In, Y. The efficacy and safety of polydeoxyribonucleotide for the treatment of knee osteoarthritis: Systematic review and meta-analysis of randomized controlled trials. Medicine 2019, 98, e17386. [Google Scholar] [CrossRef] [PubMed]
- Picciolo, G.; Pallio, G.; Altavilla, D.; Vaccaro, M.; Oteri, G.; Irrera, N.; Squadrito, F. β-Caryophyllene Reduces the Inflammatory Phenotype of Periodontal Cells by Targeting CB2 Receptors. Biomedicines 2020, 8, 164. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Galfo, F.; Pallio, G.; Mannino, F.; D’amore, A.; Pellegrino, E.; Ieni, A.; Russo, G.T.; Calapai, M.; et al. Effects of the antagomiRs 15b and 200b on the altered healing pattern of diabetic mice. Br. J. Pharmacol. 2018, 175, 644–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irrera, N.; Russo, M.; Pallio, G.; Bitto, A.; Mannino, F.; Minutoli, L.; Altavilla, D.; Squadrito, F. The Role of NLRP3 Inflammasome in the Pathogenesis of Traumatic Brain Injury. Int. J. Mol. Sci. 2020, 21, 6204. [Google Scholar] [CrossRef] [PubMed]
- Ferlito, M.; Romanenko, O.G.; Ashton, S.; Squadrito, F.; Halushka, P.V.; Cook, J.A. Effect of cross-tolerance between endotoxin and TNF-alpha or IL-1beta on cellular signaling and mediator production. J. Leukoc. Biol. 2001, 70, 821–829. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| β-actin | Fw:5′AGAGCTACGAGCTGCCTGAC3′ |
| Rw:5′AGCACTGTGTTGGCGTACAG3′ | |
| IL-1β | Fw:5′TGAGCTCGCCAGTGAAATGA3′ |
| Rw:5′AGATTCGTAGCTGGATGCCG3′ | |
| TNF-α | Fw:5′CAGAGGGCCTGTACCTCATC3′ |
| Rw:5′GGAAGACCCCTCCCAGATAG3′ | |
| IL-6 | Fw:5′TTCGGTCCAGTTGCCTTCTC3′ |
| Rw:5′CAGCTCTGGCTTGTTCCTCA3′ | |
| IL-10 | Fw:5′TGGCGCGGTGGATTCATAC3′ |
| Rw:5′AGGGGTCTGTTTTGTTGGCA3′ | |
| Bcl-2 | Fw:5′GCTCTTGAGATCTCCGGTTG3′ |
| Rw:5′AATGCATAAGGCAACGATCC3′ | |
| Bax | Fw:5′TTTGCTTCAGGGTTTCATCC3′ |
| Rw:5′CAGTTGAAGTTGCCGTCAGA3′ | |
| Caspase-3 | Fw:5′CCTGGTTCATCCAGTCGCTT |
| Rw:5′ TCTGTTGCCACCTTTCGGTT | |
| Caspase-8 | Fw:5′GGTTAGGGGACTCGGAGACT3′ |
| Rw:5′CAGGCTCAGGAACTTGAGGG3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceravolo, I.; Mannino, F.; Irrera, N.; Minutoli, L.; Arcoraci, V.; Altavilla, D.; Cavallini, G.M.; Guarini, S.; Squadrito, F.; Pallio, G. Beneficial Effects of Polydeoxyribonucleotide (PDRN) in an In Vitro Model of Fuchs Endothelial Corneal Dystrophy. Pharmaceuticals 2022, 15, 447. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15040447
Ceravolo I, Mannino F, Irrera N, Minutoli L, Arcoraci V, Altavilla D, Cavallini GM, Guarini S, Squadrito F, Pallio G. Beneficial Effects of Polydeoxyribonucleotide (PDRN) in an In Vitro Model of Fuchs Endothelial Corneal Dystrophy. Pharmaceuticals. 2022; 15(4):447. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15040447
Chicago/Turabian StyleCeravolo, Ida, Federica Mannino, Natasha Irrera, Letteria Minutoli, Vincenzo Arcoraci, Domenica Altavilla, Gian Maria Cavallini, Salvatore Guarini, Francesco Squadrito, and Giovanni Pallio. 2022. "Beneficial Effects of Polydeoxyribonucleotide (PDRN) in an In Vitro Model of Fuchs Endothelial Corneal Dystrophy" Pharmaceuticals 15, no. 4: 447. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15040447