
Tigecycline and Gentamicin-Combined Treatment Enhances Renal Damage: Oxidative Stress, Inflammatory Reaction, and Apoptosis Interplay

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
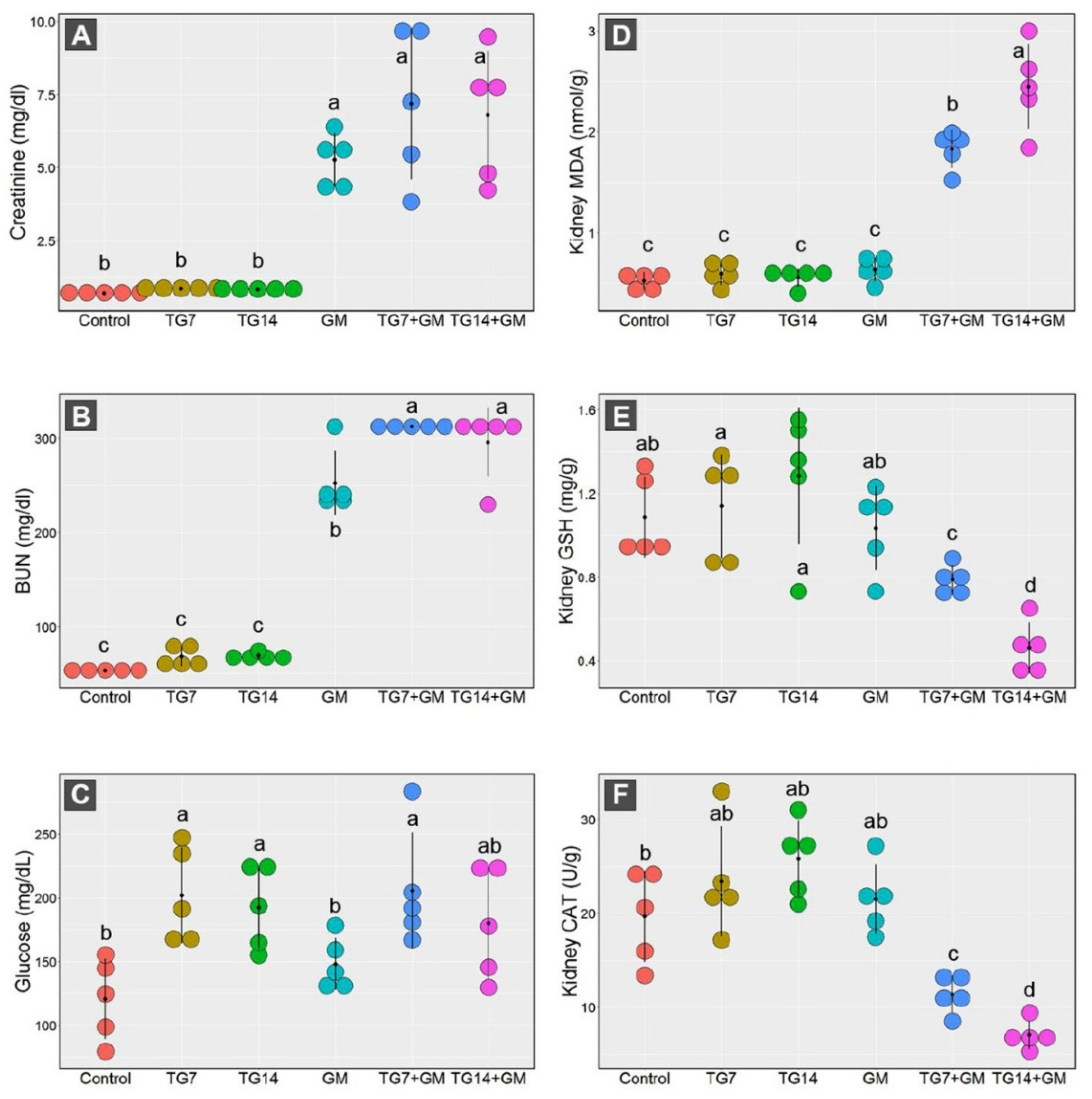
2.1. Effect of TG and GM Combination on the Kidney Function
2.2. Effect of TG and GM Combination on the Oxidative State of Renal Tissue
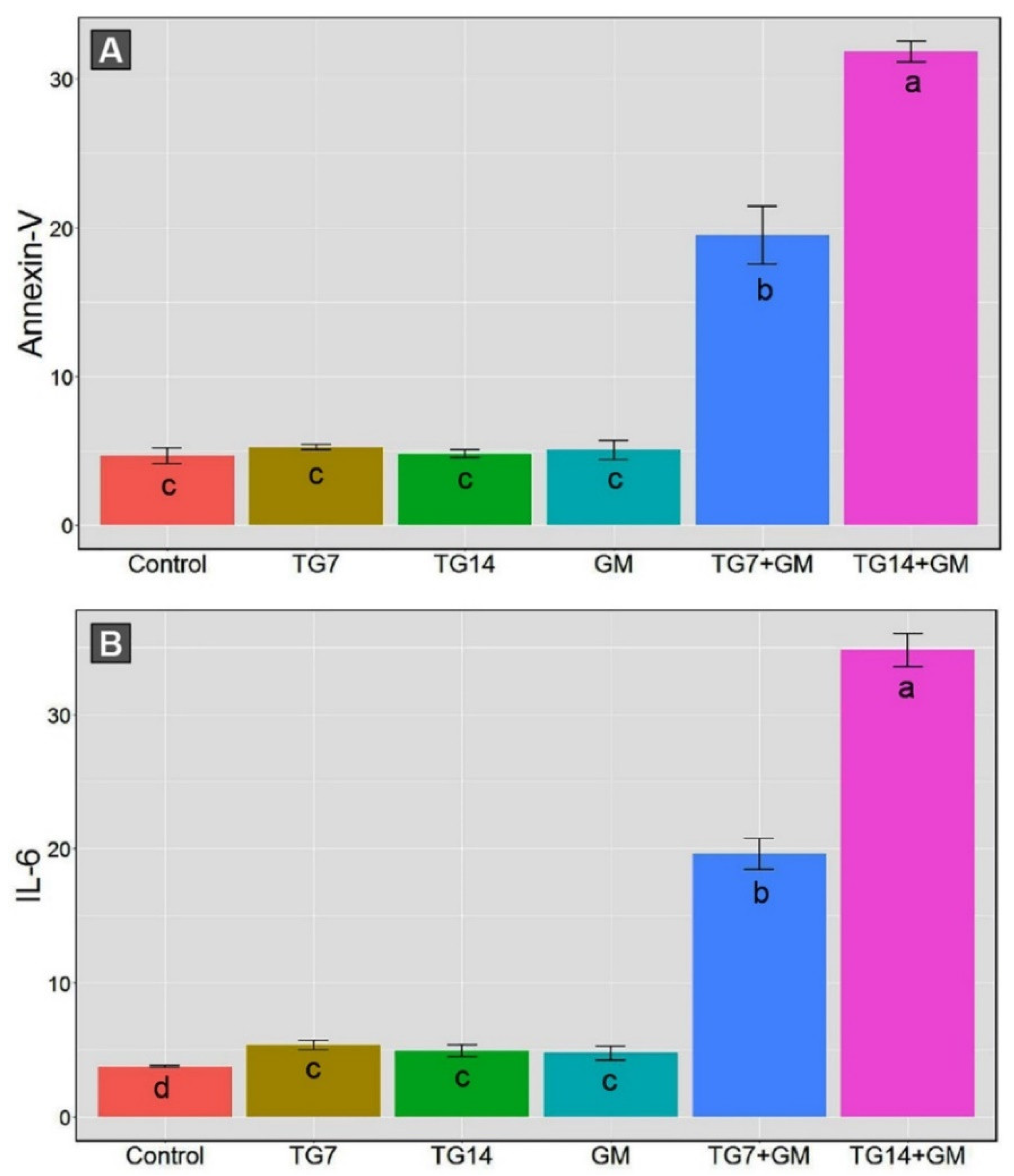
2.3. Effect of TG and GM Combination on Apoptosis and Inflammatory Response
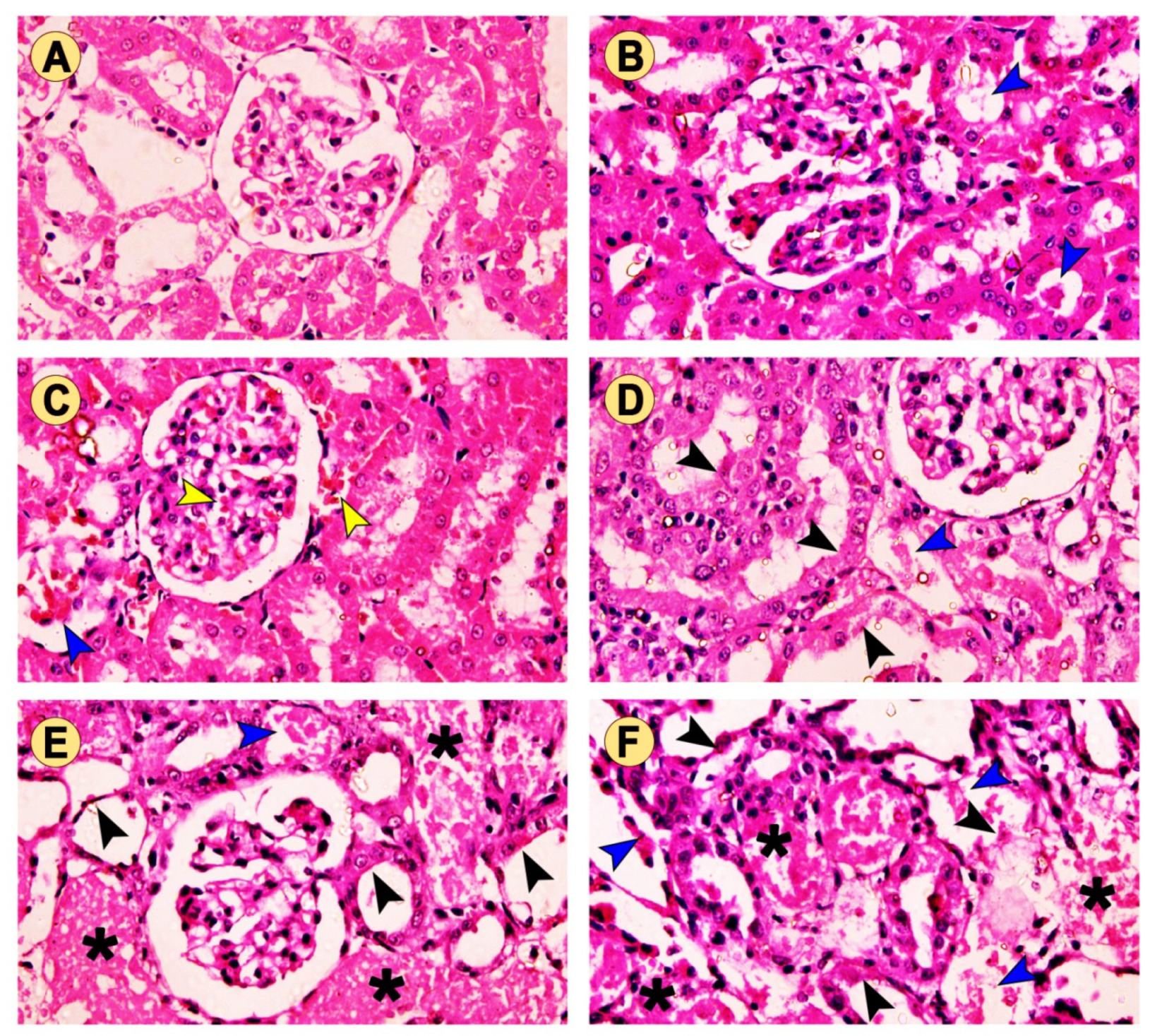
2.4. Histopathological Changes in Kidney Tissue
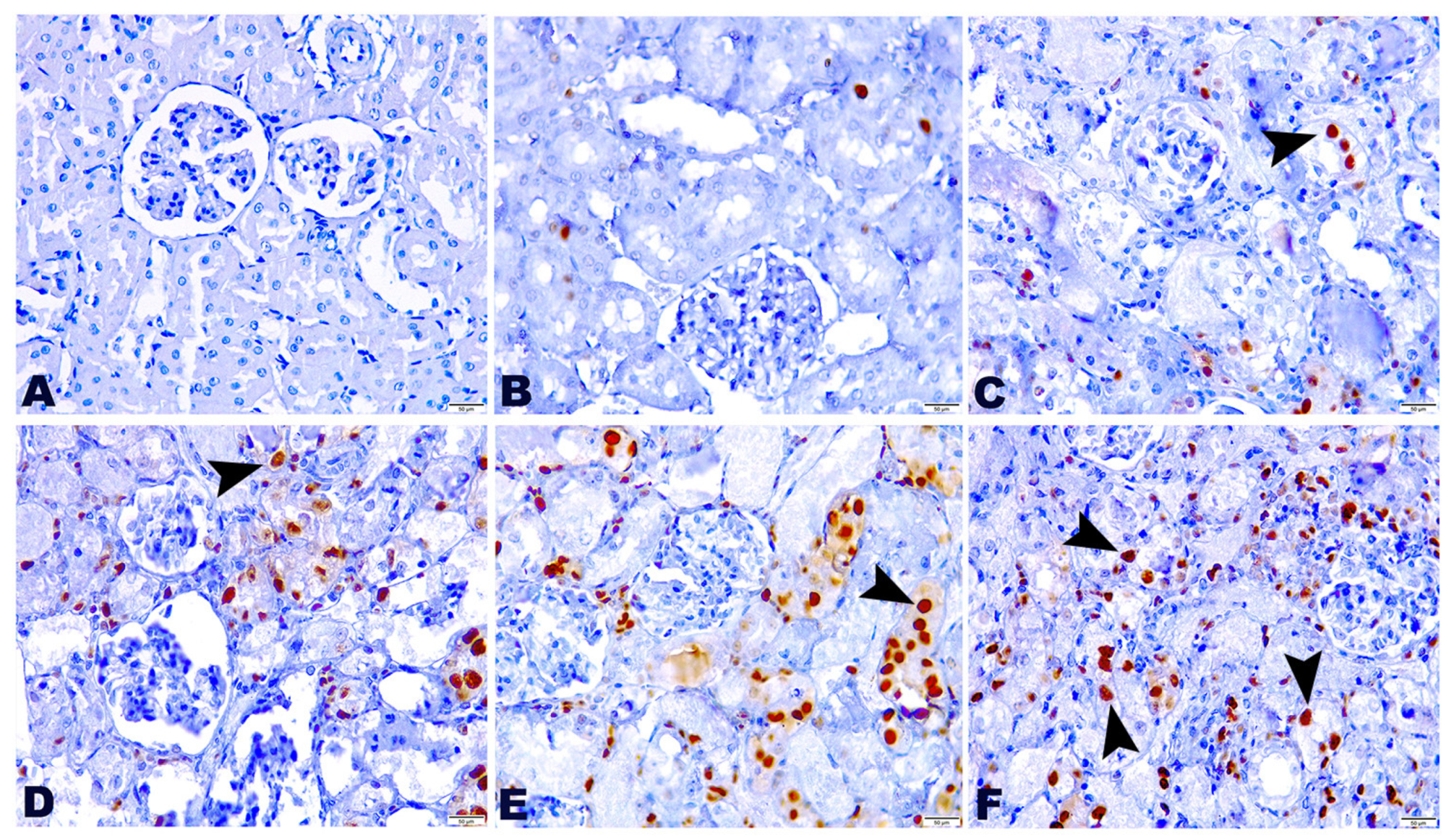
2.5. Effect of TG and GM Combination on PCNA Protein Expression Level
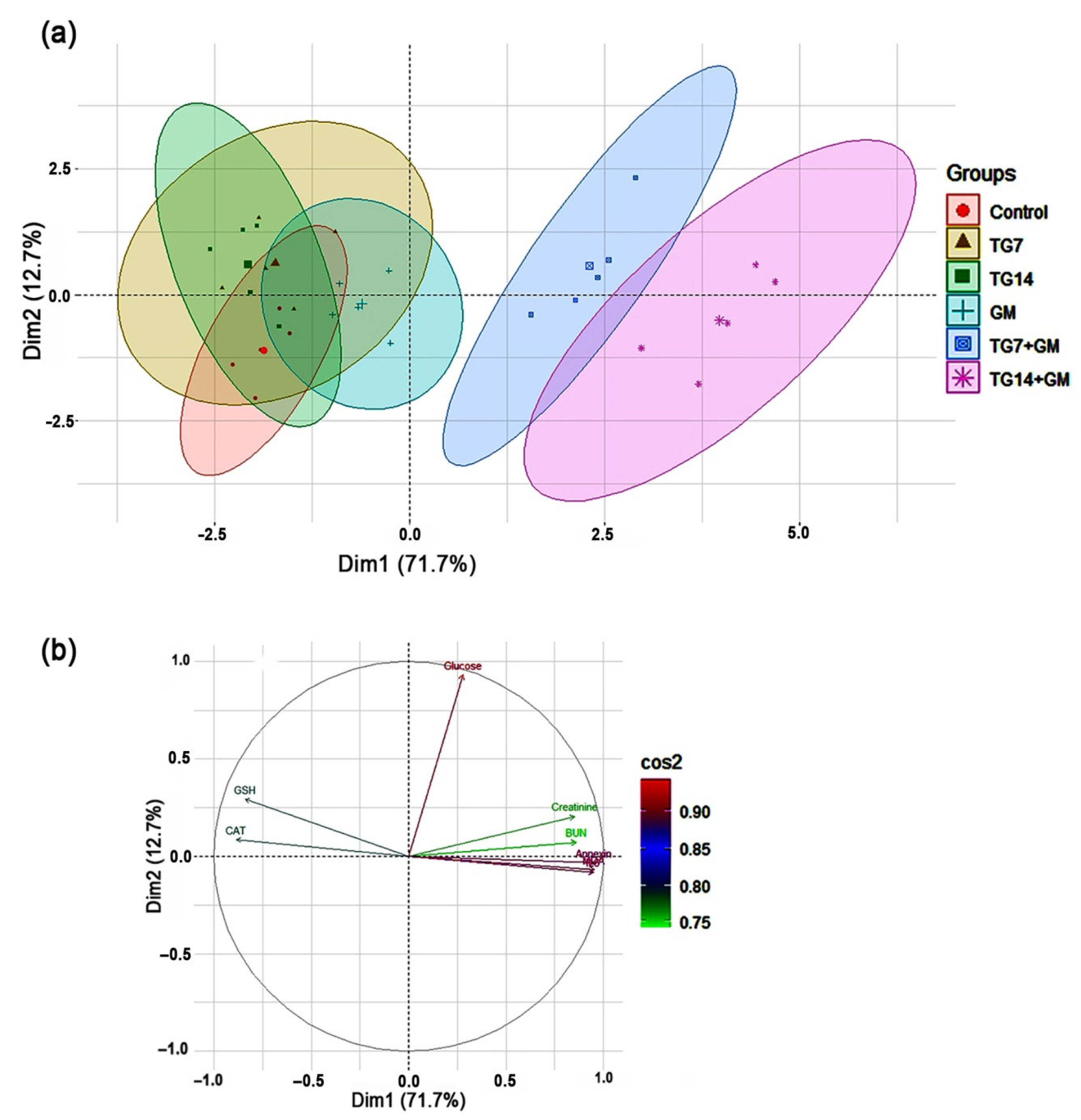
2.6. Principal Component Analysis (PCA)
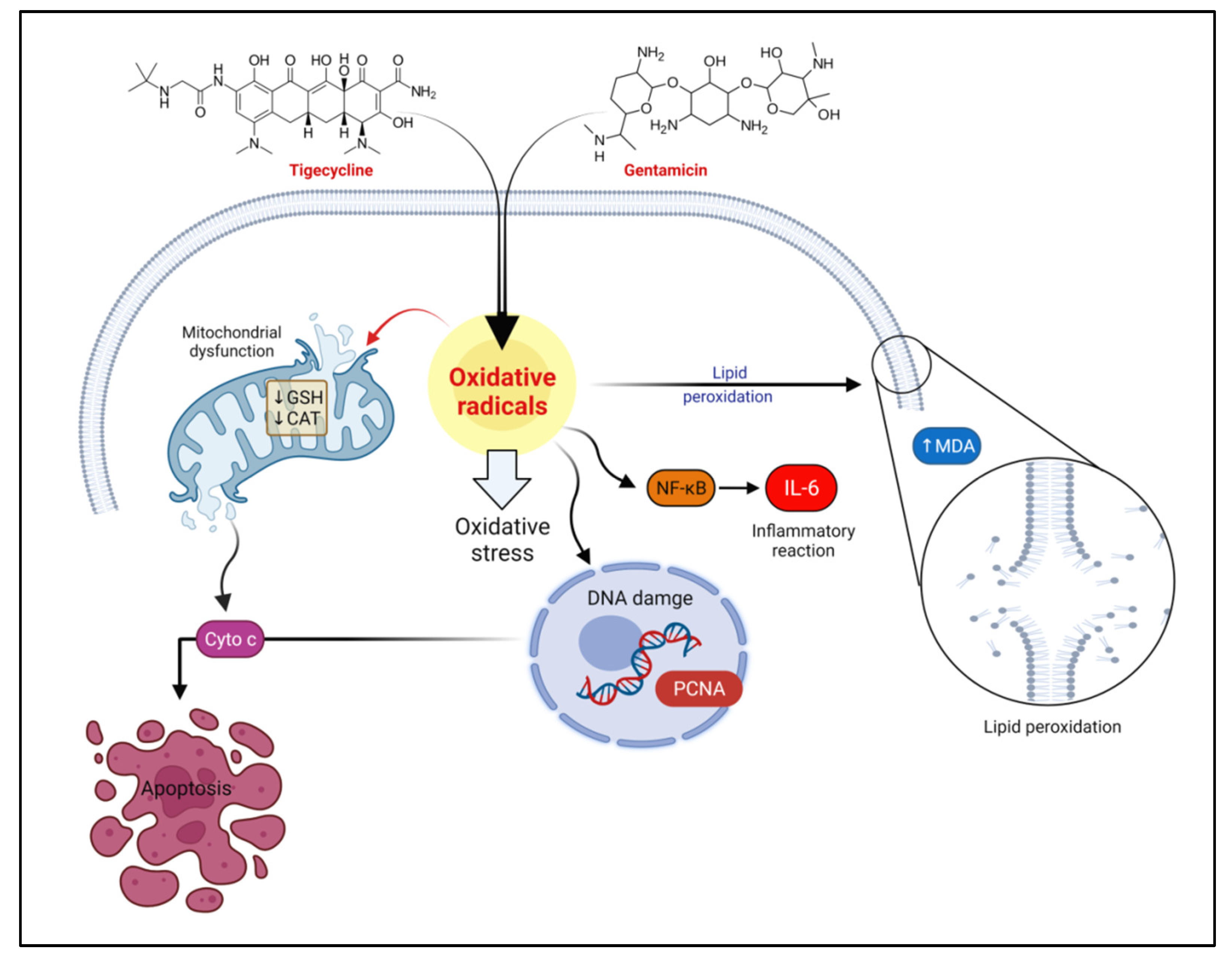
3. Discussion
4. Materials and Methods
4.1. Drugs
4.2. Experimental Animals
4.3. Experimental Design
4.4. Sampling
4.5. Serum Biochemical Studies
4.6. Tissue Oxidative Cascade Evaluation
4.7. Histopathological Examinations
4.8. Immunohistochemical Examination
4.9. Annexin-V-FITC and Interleukin-6 (IL-6) Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hu, H.; Dong, Z.; Tan, P.; Zhang, Y.; Liu, L.; Yang, L. Antibiotic drug tigecycline inhibits melanoma progression and metastasis in a p21CIP1/Waf1-dependent manner. Oncotarget 2015, 7, 3171–3185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Abbas, M.N.; Kausar, S.; Yang, J.; Li, L.; Tan, L.; Cui, H. Biological Functions and Molecular Mechanisms of Antibiotic Tigecycline in the Treatment of Cancers. Int. J. Mol. Sci. 2019, 20, 3577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, B.A.; Baron, J.; Cunha, C.B. Once daily high dose tigecycline–pharmacokinetic/pharmacodynamic based dosing for optimal clinical effectiveness: Dosing matters, revisited. Expert Rev. Anti. Infect. Ther. 2017, 15, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubi, S.; Zekiy, A.O.; Krutova, M.; Gholami, M.; Kouhsari, E.; Sholeh, M.; Ghafouri, Z.; Maleki, F. Tigecycline antibacterial activity, clinical effectiveness, and mechanisms and epidemiology of resistance: Narrative review. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 20, 4121. [Google Scholar] [CrossRef] [PubMed]
- Moghnieh, R.; Abdallah, D.; Awad, L.; Jadayel, M.; Haddad, N.; Tamim, H.; Zaiter, A.; Awwad, D.C.; Sinno, L.; El-Hassan, S.; et al. The effect of an antibiotic stewardship program on tigecycline use in a Tertiary Care Hospital, an intervention study. Annu. Clin. Microbiol. Antimicrob. 2020, 19, 35. [Google Scholar] [CrossRef] [PubMed]
- Greer, N.D. Tigecycline (Tygacil): The First in the Glycylcycline Class of Antibiotics. Baylor Univ. Med. Cent. Proc. 2006, 19, 155–161. [Google Scholar] [CrossRef]
- Kadoyama, K.; Sakaeda, T.; Tamon, A.; Okuno, Y. Adverse event profile of tigecycline: Data mining of the public version of the U.S. Food and Drug Administration adverse event reporting system. Biol. Pharm. Bull. 2012, 35, 967–970. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Xiao, Y.H.; Zheng, Y.; Lai, Y.X.; Fang, X.L.; Fang, Q. Clinical characteristics and risk factors of tigecycline-associated hypofibrinogenaemia in critically ill patients. Eur. J. Clin. Pharmacol. 2020, 76, 913–922. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Homenuik, K.; Nichol, K.; Noreddin, A.; Vercaigne, L.; Embil, J.; Gin, A.; Karlowsky, J.A.; Hoban, D.J. The Glycylcyclines: A Comparative Review with the Tetracyclines. Drugs 2004, 64, 63–88. [Google Scholar] [CrossRef]
- Sriskanthadevan, S.; Jhas, B.; Gebbia, M.; Wang, X.; Wang, Z.; Hurren, R.; Jitkova, Y.; Gronda, M.; Maclean, N.; Lai, C.K.; et al. Article Inhibition of Mitochondrial Translation as a Therapeutic Strategy for Human Acute Myeloid Leukemia. Cancer Cell 2011, 20, 674–688. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Yang, L.; Jiang, X.; Xu, C.; Wang, M.; Wang, Q.; Zhou, Z.; Xiang, Z.; Cui, H. Biochemical and Biophysical Research Communications Antibiotic drug tigecycline inhibited cell proliferation and induced autophagy in gastric cancer cells. Biochem. Biophys. Res. Commun. 2014, 446, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Ren, A.; Qiu, Y.; Cui, H.; Fu, G. Tigecycline exerts an antitumoral effect in oral squamous cell carcinoma. Oral Dis. 2015, 21, 558–564. [Google Scholar] [CrossRef]
- Zhong, X.; Zhao, E.; Tang, C.; Zhang, W.; Tan, J. Antibiotic drug tigecycline reduces neuroblastoma cells proliferation by inhibiting Akt activation in vitro and in vivo. Tumor Biol. 2016, 7615–7623. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.; Robinson, T.J.; Liu, J.C.; Shrestha, M.; Voisin, V.; Ju, Y.; Chung, P.E.D.; Pellecchia, G.; Fell, V.L.; Bae, S.; et al. RB1 deficiency in triple-negative breast cancer induces mitochondrial protein translation. J. Clin. Investig. 2016, 126, 3739–3757. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Song, M.; Zhou, M.; Hu, Y. Antibiotic tigecycline enhances cisplatin activity against human hepatocellular carcinoma through inducing mitochondrial dysfunction and oxidative damage. Biochem. Biophys. Res. Commun. 2017, 483, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Smits, P.; Smeitink, J.; van den Heuvel, L. Mitochondrial Translation and Beyond: Processes Implicated in Combined Oxidative Phosphorylation Deficiencies. J. Biomed. Biotechnol. 2010, 2010, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Wang, R.; Chen, J. Tigecycline-induced acute pancreatitis in a renal transplant patient: A case report and literature review. BMC Infect. Dis. 2018, 18, 1–6. [Google Scholar] [CrossRef]
- Li, M.X.; Li, N.; Zhu, L.Q.; Liu, W. Optimization of tigecycline dosage regimen for different infections in the patients with hepatic or renal impairment. J. Chemother. 2020, 32, 420–428. [Google Scholar] [CrossRef]
- Abdeen, A.; Samir, A.; Elkomy, A.; Aboubaker, M.; Habotta, O.A.; Gaber, A.; Alsanie, W.F.; Abdullah, O.; Elnoury, H.A.; Baioumy, B.; et al. The potential antioxidant bioactivity of date palm fruit against gentamicin-mediated hepato-renal injury in male albino rats. Biomed. Pharmacother. 2021, 143, 112154. [Google Scholar] [CrossRef]
- Xie, J.; Talaska, A.E.; Schacht, J. New developments in aminoglycoside therapy and ototoxicity. Hear. Res. 2011, 281, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Boroushaki, M.T.; Fanoudi, S.; Mollazadeh, H.; Boroumand-Noughabi, S.; Hosseini, A. Reno-protective effect of Rheum turkestanicum against gentamicin-induced nephrotoxicity. Iran. J. Basic Med. Sci. 2019, 22, 327–332. [Google Scholar] [CrossRef]
- Ghaznavi, R.; Kadkhodaee, M. Comparative effects of selective and non-selective nitric oxide synthase inhibition in gentamicin-induced rat nephrotoxicity. Arch. Toxicol. 2007, 81, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Rashid, U.; Khan, M.R. Fagonia olivieri prevented hepatorenal injuries induced with gentamicin in rat. Biomed. Pharmacother. 2017, 88, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Famurewa, A.C.; Maduagwuna, E.K.; Folawiyo, A.M.; Besong, E.E.; Eteudo, A.N.; Famurewa, O.A.; Ejezie, F.E. Antioxidant, anti-inflammatory, and antiapoptotic effects of virgin coconut oil against antibiotic drug gentamicin-induced nephrotoxicity via the suppression of oxidative stress and modulation of iNOS/NF-ĸB/caspase-3 signaling pathway in Wistar rats. J. Food Biochem. 2019, 56, e13100. [Google Scholar] [CrossRef]
- Ghaznavi, H.; Mehrzadi, S.; Dormanesh, B.; Tabatabaei, S.M.T.H.; Vahedi, H.; Hosseinzadeh, A.; Pazoki-Toroudi, H.R.; Rashidian, A. Comparison of the Protective Effects of Melatonin and Silymarin Against Gentamicin-Induced Nephrotoxicity in Rats. J. Evid. Based Complement. Altern. Med. 2016, 21, NP49–NP55. [Google Scholar] [CrossRef]
- McConeghy, K.W.; LaPlante, K.L. In vitro activity of tigecycline in combination with gentamicin against biofilm-forming Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2010, 68, 1–6. [Google Scholar] [CrossRef]
- Pontikis, K.; Pefanis, A.; Tsaganos, T.; Tzepi, I.M.; Carrer, D.P.; Giamarellou, H. Efficacy of tigecycline alone and in combination with gentamicin in the treatment of experimental endocarditis due to linezolid-resistant Enterococcus faecium. Antimicrob. Agents Chemother. 2013, 57, 3392–3394. [Google Scholar] [CrossRef] [Green Version]
- Sbrana, F.; Malacarne, P.; Viaggi, B.; Costanzo, S.; Leonetti, P.; Leonildi, A.; Casini, B.; Tascini, C.; Menichetti, F. Carbapenem-sparing antibiotic regimens for infections caused by Klebsiella pneumoniae carbapenemase-producing K. pneumoniae in intensive care unit. Clin. Infect. Dis. 2013, 56, 697–700. [Google Scholar] [CrossRef] [Green Version]
- Vergidis, P.; Schmidt-Malan, S.M.; Mandrekar, J.N.; Steckelberg, J.M.; Patel, R. Comparative activities of vancomycin, tigecycline and rifampin in a rat model of methicillin-resistant Staphylococcus aureus osteomyelitis. J. Infect. 2015, 70, 609–615. [Google Scholar] [CrossRef]
- Yin, L.Y.; Lazzarini, L.; Li, F.; Stevens, C.M.; Calhoun, J.H. Comparative evaluation of tigecycline and vancomycin with and without rifampicin, in the treatment of methicillin-resistant Staphylococcus aureus experimental osteomyelitis in a rabbit model. J. Antimicrob. Chemother. 2005, 55, 995–1002. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.A.; Raish, M.; Ahmad, A.; Ahmad, S.F.; Mudassar, S.; Mohsin, K.; Shakeel, F.; Korashy, H.M.; Bakheet, S.A. Sinapic acid mitigates gentamicin-induced nephrotoxicity and associated oxidative/nitrosative stress, apoptosis, and inflammation in rats. Life Sci. 2016, 165, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Arjinajarn, P.; Chueakula, N.; Pongchaidecha, A.; Jaikumkao, K.; Chatsudthipong, V.; Mahatheeranont, S.; Norkaew, O.; Chattipakorn, N.; Lungkaphin, A. Anthocyanin-rich Riceberry bran extract attenuates gentamicin-induced hepatotoxicity by reducing oxidative stress, inflammation and apoptosis in rats. Biomed. Pharmacother. 2017, 92, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, A.; Goudarzi, M.; Karimi, M.Y.; Khorsandi, L.; Mehrzadi, S.; Mombeini, M.A. Zingerone ameliorates gentamicin-induced nephrotoxicity in rats. Comp. Clin. Path. 2020, 29, 971–980. [Google Scholar] [CrossRef]
- Uddin, M.J.; Kim, E.H.; Hannan, M.A.; Ha, H. Pharmacotherapy against oxidative stress in chronic kidney disease: Promising small molecule natural products targeting nrf2-ho-1 signaling. Antioxidants 2021, 10, 258. [Google Scholar] [CrossRef] [PubMed]
- Kehrer, J.P. The Haber-Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Jomova, K.; Valko, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef]
- Ozbek, E. Induction of oxidative stress in kidney. Int. J. Nephrol. 2012, 2012, 5897. [Google Scholar] [CrossRef] [Green Version]
- Pillon, N.J.; Soulage, C.O. Lipid Peroxidation by-Products and the Metabolic Syndrome. In Lipid Peroxidation; InTech: Houston, TX, USA, 2012; pp. 409–436. ISBN 978-953-51-0716-3. [Google Scholar]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Abdeen, A.; Abdelkader, A.; Abdo, M.; Wareth, G.; Aboubakr, M.; Aleya, L.; Abdel-Daim, M. Protective effect of cinnamon against acetaminophen-mediated cellular damage and apoptosis in renal tissue. Environ. Sci. Pollut. Res. 2019, 26, 240–249. [Google Scholar] [CrossRef]
- Abdeen, A.; Abou-Zaid, O.A.; Abdel-Maksoud, H.A.; Aboubakr, M.; Abdelkader, A.; Abdelnaby, A.; Abo-Ahmed, A.I.; El-Mleeh, A.; Mostafa, O.; Abdel-Daim, M.; et al. Cadmium overload modulates piroxicam-regulated oxidative damage and apoptotic pathways. Environ. Sci. Pollut. Res. 2019, 26, 25167–25177. [Google Scholar] [CrossRef]
- Abdel-daim, M.M.; Abdeen, A.; Jalouli, M.; Abdelkader, A.; Megahed, A. Fucoidan supplementation modulates hepato-renal oxidative stress and DNA damage induced by a fl atoxin B1 intoxication in rats. Sci. Total Environ. 2021, 768, 144781. [Google Scholar] [CrossRef] [PubMed]
- Bayir, H.; Kagan, V.E. Bench-to-bedside review: Mitochondrial injury, oxidative stress and apoptosis–There is nothing more practical than a good theory. Crit. Care 2008, 12, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yates, S.A.; Dempster, N.M.; Murphy, M.F.; Moore, S.A. Quantitative analysis of malondialdehyde-guanine adducts in genomic DNA samples by liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2017, 31, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, P.D. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Avery, S.V. Molecular targets of oxidative stress. Biochem. J. 2011, 434, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakar, T.; Leung, W.; Nicolae, C.M.; Clements, K.E.; Shen, B.; Bielinsky, A.; Moldovan, G. Ubiquitinated-PCNA protects replication forks from fragment maturation and chromatin assembly. Nat. Commun. 2020, 11, 2147. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Sayed, A.A.; Abdeen, A.; Aleya, L.; Ali, D.; Alkahtane, A.A.; Alarifi, S.; Alkahtani, S. Piperine enhances the antioxidant and anti-inflammatory activities of thymoquinone against microcystin-LR-induced hepatotoxicity and neurotoxicity in mice. Oxid. Med. Cell. Longev. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Aboubakr, M.; Abdelkader, A.; Habotta, O.A.; Adel, N.; Emam, M.A.; Abdelhiee, E.Y.; Shanab, O.; Shoghy, K.; Elnoury, H.; Soliman, M.M.; et al. Cefepime and diclofenac sodium combined treatment-potentiated multiple organ injury: Role of oxidative damage and disrupted lipid metabolism. J. Biochem. Mol. Toxicol. 2021, 35, 1–13. [Google Scholar] [CrossRef]
- Sharma, V.C.; Kaushik, A.; Dey, Y.N.; Srivastava, B.; Wanjari, M.; Pawar, S.; Chougule, S. Nephroprotective potential of Anogeissus latifolia Roxb. (Dhava) against gentamicin-induced nephrotoxicity in rats. J. Ethnopharmacol. 2021, 273, 4001. [Google Scholar] [CrossRef]
- Aydın, G.; Gokcimen, A.; Oncü, M.; Çiçek, E.; Karahan, N.; Gökalp, O. Histopathologic Changes in Liver and Renal Tissues Induced by Different Doses of Diclofenac Sodium in Rats. Turk. J. Vet. Anim. Sci 2003, 27, 1131–1140. [Google Scholar]
- Pathak, N.; Khandelwal, S. Immunomodulatory role of piperine in cadmium induced thymic atrophy and splenomegaly in mice. Environ. Toxicol. Pharmacol. 2009, 28, 52–60. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | ||||||
|---|---|---|---|---|---|---|
| Control | TG7 | TG14 | GM | TG7+GM | TG14+GM | |
| Congestion of glomerular tuft | − | − | + | − | − | − |
| Congestion of intertubular capillaries | − | − | + | − | + | + |
| Intratubular eosinophilic debris | − | + | + | + | ++ | +++ |
| Tubular degeneration | − | − | − | + | ++ | +++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elgazzar, D.; Aboubakr, M.; Bayoumi, H.; Ibrahim, A.N.; Sorour, S.M.; El-Hewaity, M.; Elsayed, A.M.; Shehata, S.A.; Bayoumi, K.A.; Alsieni, M.; et al. Tigecycline and Gentamicin-Combined Treatment Enhances Renal Damage: Oxidative Stress, Inflammatory Reaction, and Apoptosis Interplay. Pharmaceuticals 2022, 15, 736. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15060736
Elgazzar D, Aboubakr M, Bayoumi H, Ibrahim AN, Sorour SM, El-Hewaity M, Elsayed AM, Shehata SA, Bayoumi KA, Alsieni M, et al. Tigecycline and Gentamicin-Combined Treatment Enhances Renal Damage: Oxidative Stress, Inflammatory Reaction, and Apoptosis Interplay. Pharmaceuticals. 2022; 15(6):736. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15060736
Chicago/Turabian StyleElgazzar, Dina, Mohamed Aboubakr, Heba Bayoumi, Amany N. Ibrahim, Safwa M. Sorour, Mohamed El-Hewaity, Abulmaaty M. Elsayed, Shaimaa A. Shehata, Khaled A. Bayoumi, Mohammed Alsieni, and et al. 2022. "Tigecycline and Gentamicin-Combined Treatment Enhances Renal Damage: Oxidative Stress, Inflammatory Reaction, and Apoptosis Interplay" Pharmaceuticals 15, no. 6: 736. https://0-doi-org.brum.beds.ac.uk/10.3390/ph15060736