
Activation of Tenofovir Alafenamide and Sofosbuvir in the Human Lung and Its Implications in the Development of Nucleoside/Nucleotide Prodrugs for Treating SARS-CoV-2 Pulmonary Infection


Abstract
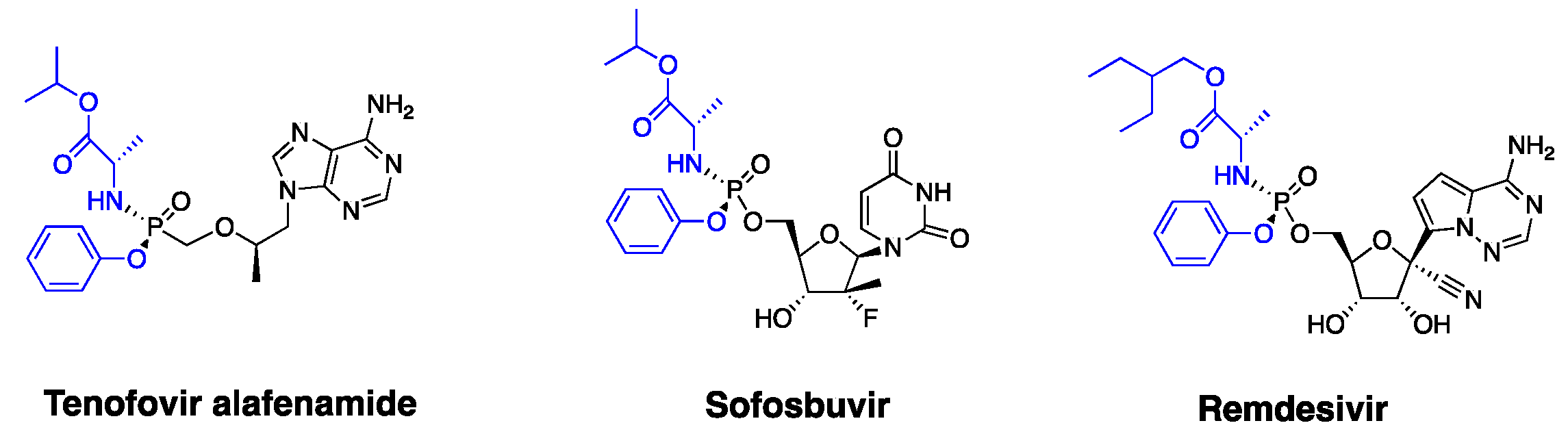
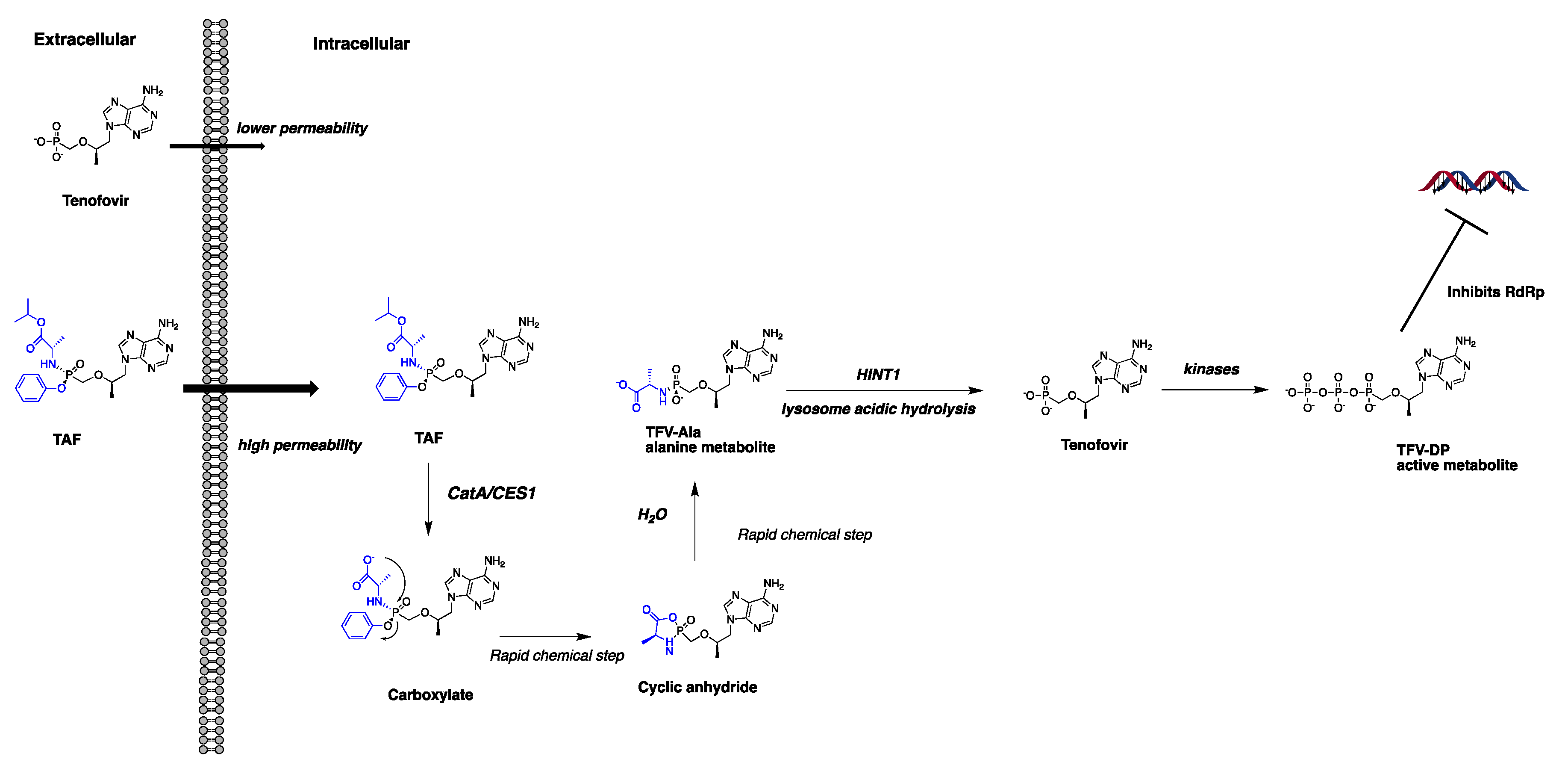
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. ABPP of Serine Hydrolases HlungS9
2.3. Enzyme and Tissue S9 Activity Assays
2.4. Inhibition Assay
2.5. LC-MS/MS-Based Proteomics Analysis
2.6. LC-MS/MS Analysis of TAF and SBV
2.7. Meta-Analysis of CatA, CES1, HINT1 Protein Expression in Human Respiratory and Hepatic Systems
2.8. Data Analysis
3. Results
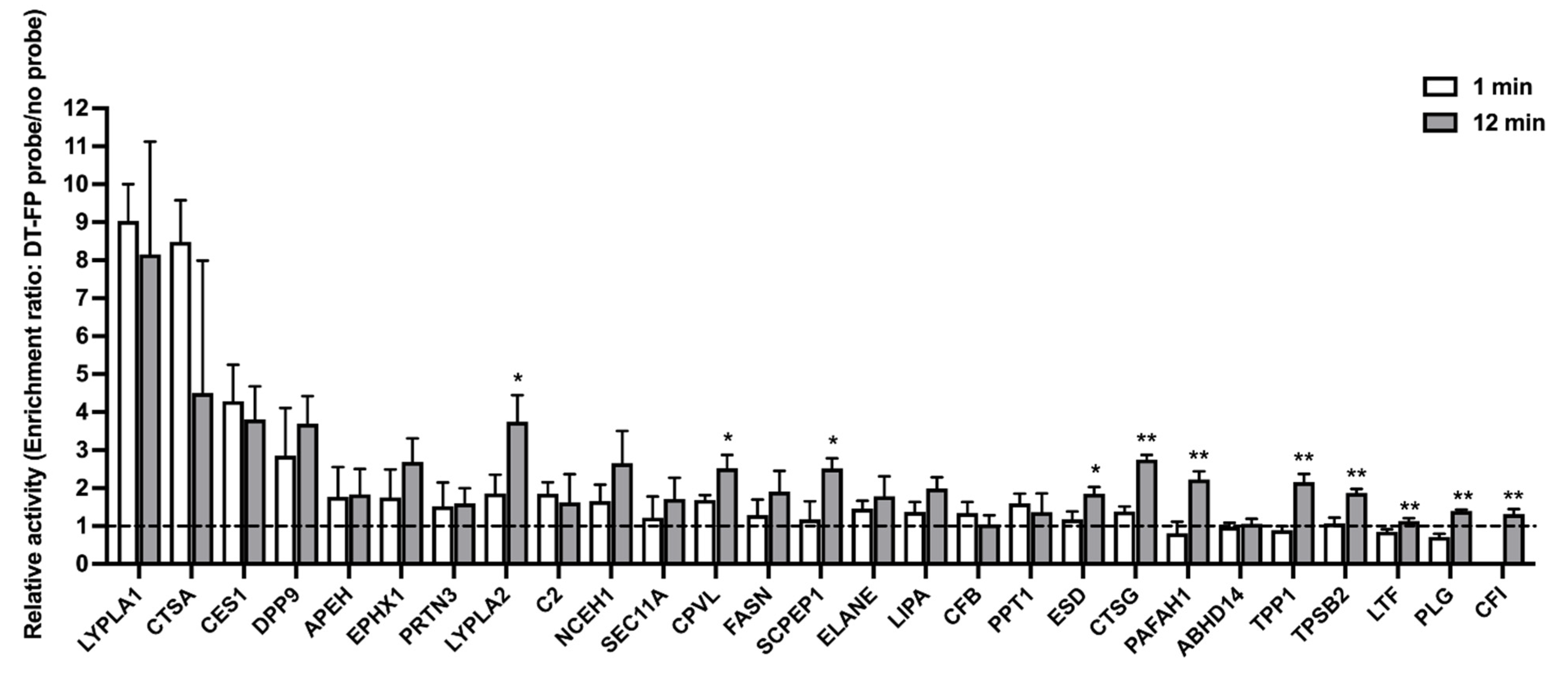
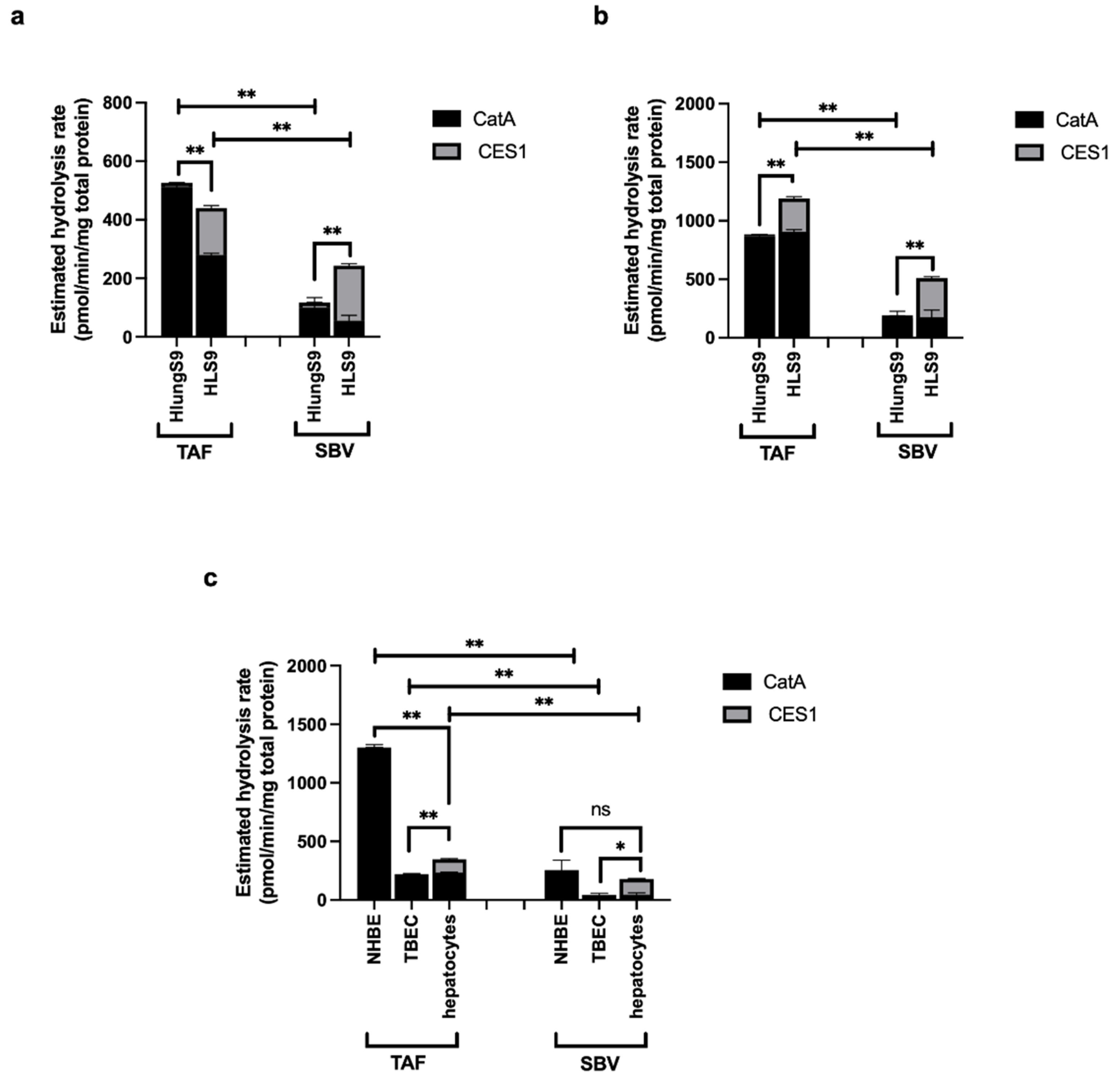
3.1. Serine Hydrolase Activity Profiles in HlungS9
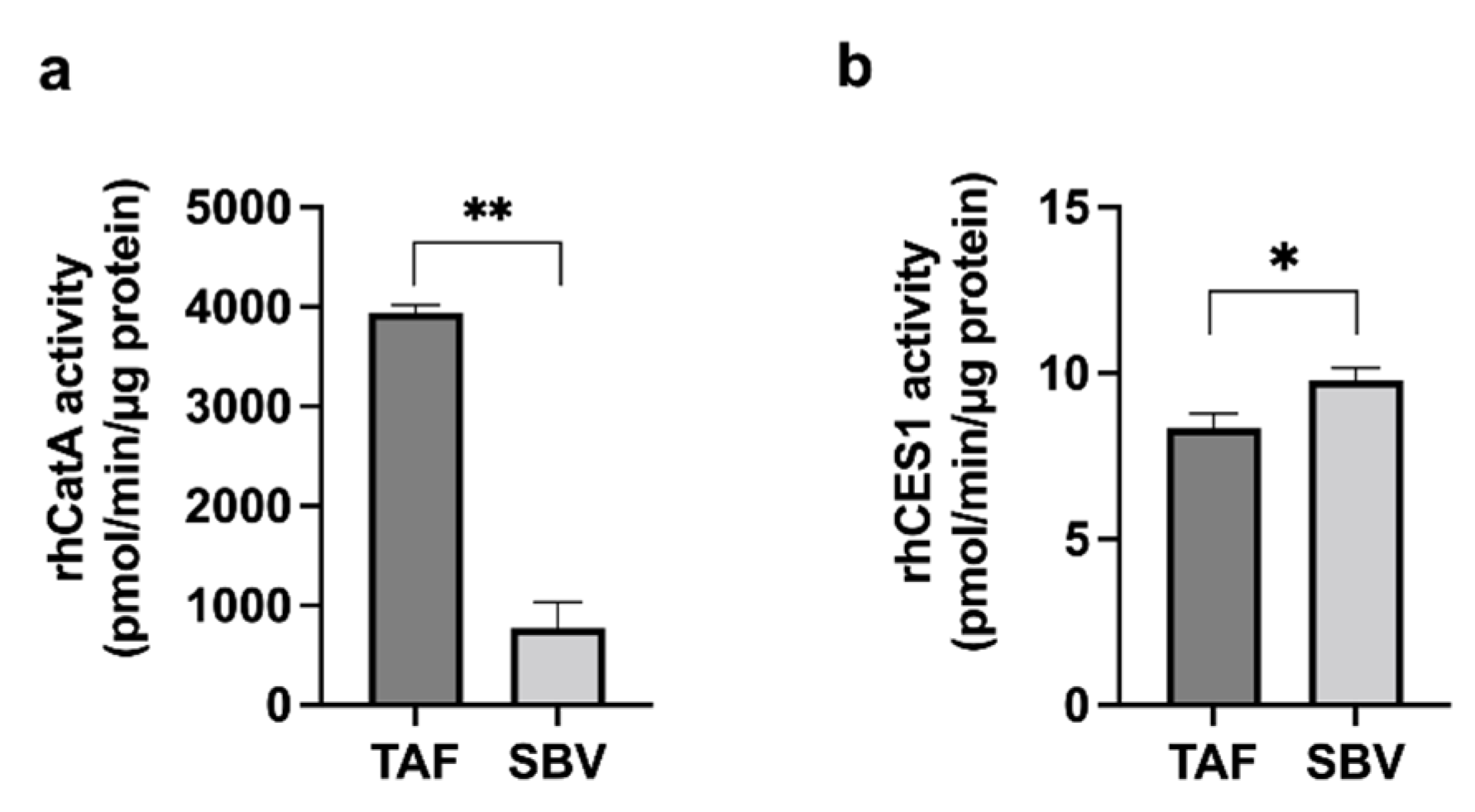
3.2. Enzyme Activity
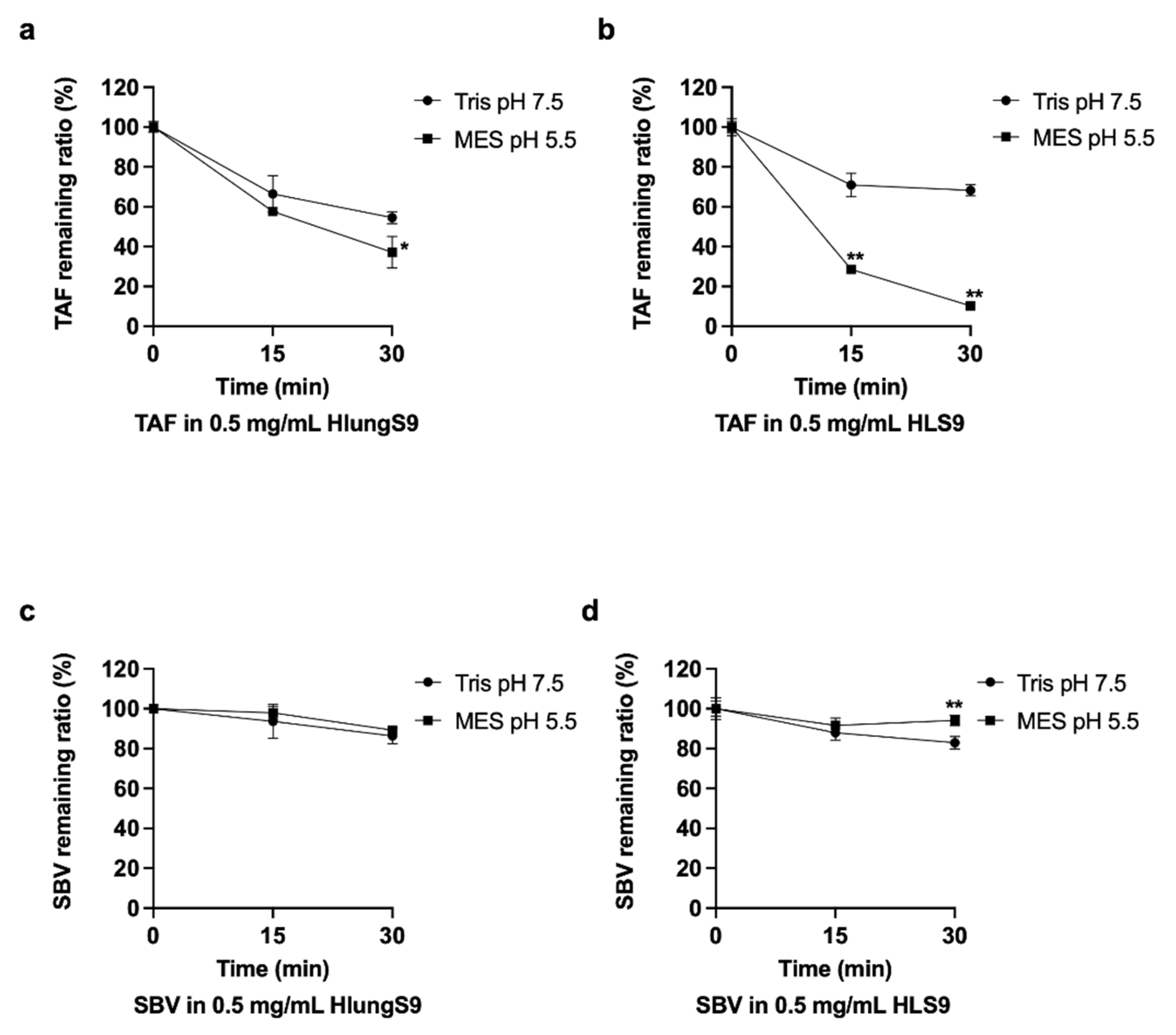
3.3. TAF and SBV Hydrolysis in HlungS9 and HLS9
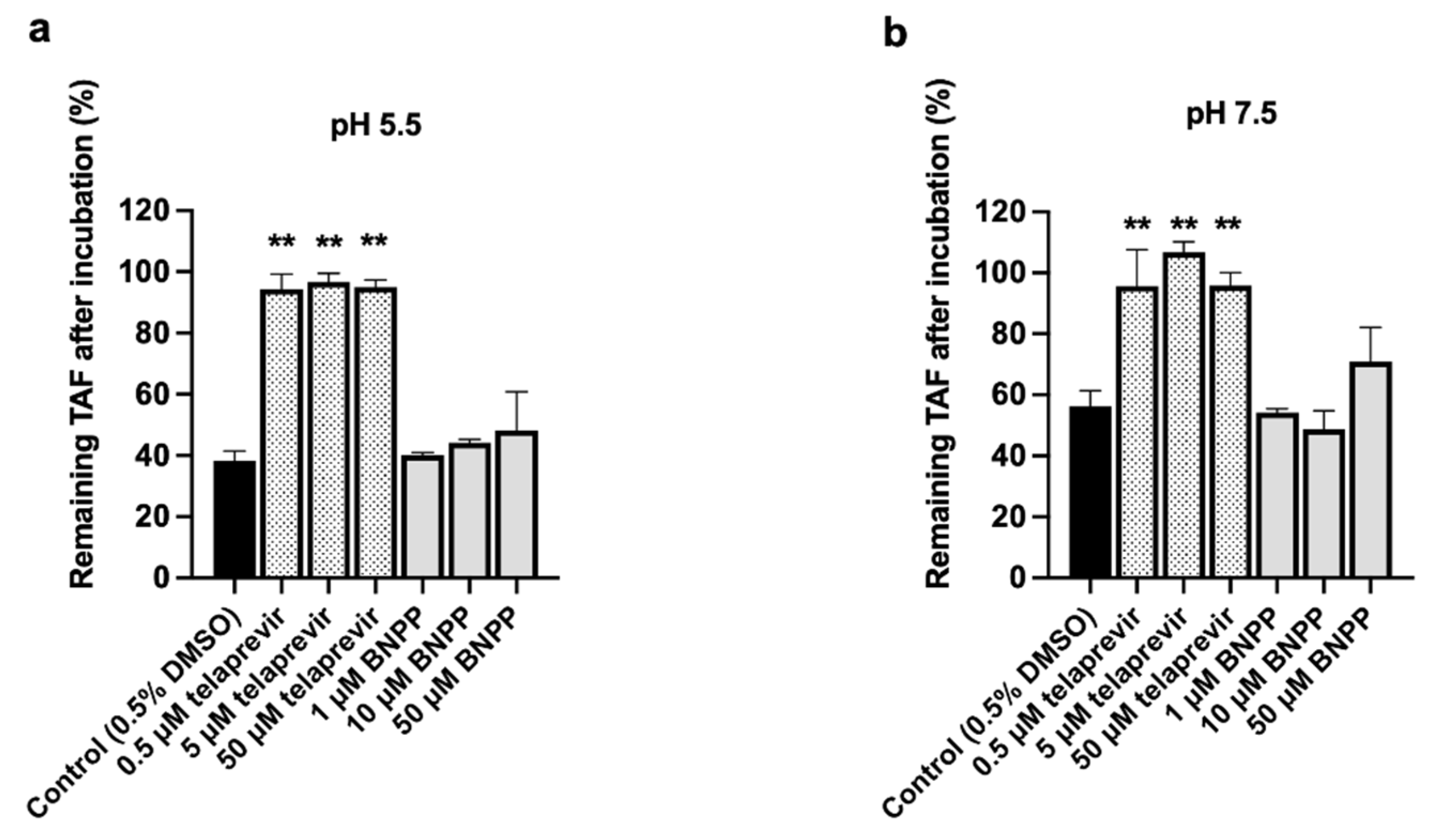
3.4. CatA and CES1 Inhibitor Effects on TAF Hydrolysis in HlungS9
3.5. Meta-Analysis of the Abundance of CatA and CES1 in Human Respiratory and Hepatic Systems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clercq, E.D.; Li, G. Approved Antiviral Drugs over the Past 50 Years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Schutter, C.; Ehteshami, M.; Hammond, E.T.; Amblard, F.; Schinazi, R.F. Metabolism of Nucleosides and Nucleotides Prodrugs. Curr. Pharm. Des. 2017, 23, 6984–7002. [Google Scholar] [CrossRef]
- Mehellou, Y.; Rattan, H.S.; Balzarini, J. The ProTide Prodrug Technology: From the Concept to the Clinic. J. Med. Chem. 2018, 61, 2211–2226. [Google Scholar] [CrossRef] [PubMed]
- Serpi, M.; Pertusati, F. An overview of ProTide technology and its implications to drug discovery. Expert Opin. Drug Discov. 2021, 16, 1149–1161. [Google Scholar] [CrossRef]
- Hao, Z.; Cooney, D.A.; Farquhar, D.; Perno, C.F.; Zhang, K.; Masood, R.; Wilson, Y.; Hartman, N.R.; Balzarini, J.; Johns, D.G. Potent DNA chain termination activity and selective inhibition of human immunodeficiency virus reverse transcriptase by 2′,3′-dideoxyuridine-5′-triphosphate. Mol. Pharmacol. 1990, 37, 157–163. [Google Scholar] [PubMed]
- Balzarini, J.; Herdewijn, P.; De Clercq, E. Differential Patterns of Intracellular Metabolism of 2′,3′-Didehydro-2′,3′-dideoxythymidine and 3′-Azido-2′,3′-dideoxythymidine, Two Potent Anti-human Immunodeficiency Virus Compounds. J. Biol. Chem. 1989, 264, 6127–6133. [Google Scholar] [CrossRef]
- Wiemer, A.J. Metabolic Efficacy of Phosphate Prodrugs and the Remdesivir Paradigm. ACS Pharmacol. Transl. Sci. 2020, 3, 613–626. [Google Scholar] [CrossRef]
- Chien, M.; Anderson, T.K.; Jockusch, S.; Tao, C.; Li, X.; Kumar, S.; Russo, J.J.; Kirchdoerfer, R.N.; Ju, J. Nucleotide Analogues as Inhibitors of SARS-CoV-2 Polymerase, a Key Drug Target for COVID-19. J. Proteome Res. 2020, 19, 4690–4697. [Google Scholar] [CrossRef]
- Jockusch, S.; Tao, C.; Li, X.; Chien, M.; Kumar, S.; Morozova, I.; Kalachikov, S.; Russo, J.J.; Ju, J. Sofosbuvir terminated RNA is more resistant to SARS-CoV-2 proofreader than RNA terminated by Remdesivir. Sci. Rep. 2020, 10, 16577. [Google Scholar] [CrossRef]
- Birkus, G.; Wang, R.; Liu, X.; Kutty, N.; MacArthur, H.; Cihlar, T.; Gibbs, C.; Swaminathan, S.; Lee, W.; McDermott, M. Cathepsin A is the major hydrolase catalyzing the intracellular hydrolysis of the antiretroviral nucleotide phosphonoamidate prodrugs GS-7340 and GS-9131. Antimicrob. Agents Chemother. 2007, 51, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Birkus, G.; Kutty, N.; He, G.-X.; Mulato, A.; Lee, W.; McDermott, M.; Cihlar, T. Activation of 9-[(R)-2-[[(S)-[[(S)-1-(Isopropoxycarbonyl) ethyl] amino] phenoxyphosphinyl]-methoxy] propyl] adenine (GS-7340) and other tenofovir phosphonoamidate prodrugs by human proteases. Mol. Pharmacol. 2008, 74, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Murakami, E.; Tolstykh, T.; Bao, H.; Niu, C.; Steuer, H.M.M.; Bao, D.; Chang, W.; Espiritu, C.; Bansal, S.; Lam, A.M. Mechanism of activation of PSI-7851 and its diastereoisomer PSI-7977. J. Biol. Chem. 2010, 285, 34337–34347. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Liclican, A.; Xu, Y.; Pitts, J.; Niu, C.; Zhang, J.; Kim, C.; Zhao, X.; Soohoo, D.; Babusis, D.; et al. Key Metabolic Enzymes Involved in Remdesivir Activation in Human Lung Cells. Antimicrob. Agents Chemother. 2021, 65, e0060221. [Google Scholar] [CrossRef] [PubMed]
- Birkus, G.; Bam, R.A.; Willkom, M.; Frey, C.R.; Tsai, L.; Stray, K.M.; Yant, S.R.; Cihlar, T. Intracellular activation of tenofovir alafenamide and the effect of viral and host protease inhibitors. Antimicrob. Agents Chemother. 2016, 60, 316–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, V.C.; Muller, F.L. Advantages of the Parent Nucleoside GS-441524 over Remdesivir for Covid-19 Treatment. ACS Med. Chem. Lett. 2020, 11, 1361–1366. [Google Scholar] [CrossRef]
- Li, J.; Liu, S.; Shi, J.; Wang, X.; Xue, Y.; Zhu, H.-J. Tissue-Specific Proteomics Analysis of Anti-COVID-19 Nucleoside and Nucleotide Prodrug-Activating Enzymes Provides Insights into the Optimization of Prodrug Design and Pharmacotherapy Strategy. ACS Pharmacol. Transl. Sci. 2021, 4, 870–887. [Google Scholar] [CrossRef]
- Simon, G.M.; Cravatt, B.F. Activity-based proteomics of enzyme superfamilies: Serine hydrolases as a case study. J. Biol. Chem. 2010, 285, 11051–11055. [Google Scholar] [CrossRef] [Green Version]
- Tallman, K.R.; Levine, S.R.; Beatty, K.E. Small-Molecule Probes Reveal Esterases with Persistent Activity in Dormant and Reactivating Mycobacterium tuberculosis. ACS Infect. Dis. 2016, 2, 936–944. [Google Scholar] [CrossRef]
- Bachovchin, D.A.; Ji, T.; Li, W.; Simon, G.M.; Blankman, J.L.; Adibekian, A.; Hoover, H.; Niessen, S.; Cravatt, B. Superfamily-wide portrait of serine hydrolase inhibition achieved by library-versus-library screening. Proc. Natl. Acad. Sci. USA 2010, 107, 20941–20946. [Google Scholar] [CrossRef] [Green Version]
- van Rooden, E.J.; van Esbroeck, A.C.M.; Baggelaar, M.P.; Deng, H.; Florea, B.I.; Marques, A.R.A.; Ottenhoff, R.; Boot, R.G.; Overkleeft, H.S.; Aerts, J.M.F.G.; et al. Chemical Proteomic Analysis of Serine Hydrolase Activity in Niemann-Pick Type C Mouse Brain. Front. Neurosci. 2018, 12, 440. [Google Scholar] [CrossRef] [Green Version]
- van Rooden, E.J.; Florea, B.I.; Deng, H.; Baggelaar, M.P.; van Esbroeck, A.C.M.; Zhou, J.; Overkleeft, H.S.; van der Stelt, M. Mapping in vivo target interaction profiles of covalent inhibitors using chemical proteomics with label-free quantification. Nat. Protoc. 2018, 13, 752–767. [Google Scholar] [CrossRef]
- Hirano, T.; Kishi, M.; Sugimoto, H.; Taguchi, R.; Obinata, H.; Ohshima, N.; Tatei, K.; Izumi, T. Thioesterase activity and subcellular localization of acylprotein thioesterase 1/lysophospholipase 1. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2009, 1791, 797–805. [Google Scholar] [CrossRef]
- He, B.; Shi, J.; Wang, X.; Jiang, H.; Zhu, H.-J. Label-free absolute protein quantification with data-independent acquisition. J. Proteom. 2019, 200, 51–59. [Google Scholar] [CrossRef]
- Wang, X.; He, B.; Shi, J.; Li, Q.; Zhu, H.-J. Comparative Proteomics Analysis of Human Liver Microsomes and S9 Fractions. Drug Metab. Dispos. 2020, 48, 31–40. [Google Scholar] [CrossRef]
- Avataneo, V.; de Nicolò, A.; Cusato, J.; Antonucci, M.; Manca, A.; Palermiti, A.; Waitt, C.; Walimbwa, S.; Lamorde, M.; di Perri, G.; et al. Development and validation of a UHPLC-MS/MS method for quantification of the prodrug remdesivir and its metabolite GS-441524: A tool for clinical pharmacokinetics of SARS-CoV-2/COVID-19 and Ebola virus disease. J. Antimicrob. Chemother. 2020, 75, 1772–1777. [Google Scholar] [CrossRef]
- Nie, X.; Qian, L.; Sun, R.; Huang, B.; Dong, X.; Xiao, Q.; Zhang, Q.; Lu, T.; Yue, L.; Chen, S.; et al. Multi-organ proteomic landscape of COVID-19 autopsies. Cell 2021, 184, 775–791.e14. [Google Scholar] [CrossRef]
- Foster, M.W.; Gwinn, W.M.; Kelly, F.L.; Brass, D.M.; Valente, A.M.; Moseley, M.A.; Thompson, J.W.; Morgan, D.L.; Palmer, S.M. Proteomic Analysis of Primary Human Airway Epithelial Cells Exposed to the Respiratory Toxicant Diacetyl. J. Proteome Res. 2017, 16, 538–549. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Vildhede, A.; Norén, A.; Artursson, P. In-depth quantitative analysis and comparison of the human hepatocyte and hepatoma cell line HepG2 proteomes. J. Proteom. 2016, 136, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Nusinow, D.P.; Szpyt, J.; Ghandi, M.; Rose, C.M.; McDonald, E.R.; Kalocsay, M.; Jané-Valbuena, J.; Gelfand, E.; Schweppe, D.K.; Jedrychowski, M.; et al. Quantitative Proteomics of the Cancer Cell Line Encyclopedia. Cell 2020, 180, 387–402.e16. [Google Scholar] [CrossRef] [PubMed]
- Satake, A.; Itoh, K.; Shimmoto, M.; Saido, T.C.; Sakuraba, H.; Suzuki, Y. Distribution of Lysosomal Protective Protein in Human Tissues. Biochem. Biophys. Res. Commun. 1994, 205, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Basit, A.; Neradugomma, N.K.; Wolford, C.; Fan, P.W.; Murray, B.; Takahashi, R.H.; Khojasteh, S.C.; Smith, B.J.; Heyward, S.; Totah, R.A.; et al. Characterization of Differential Tissue Abundance of Major Non-CYP Enzymes in Human. Mol. Pharm. 2020, 17, 4114–4124. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.-H.; Luo, T.; Shi, Y.; He, Z.-C.; Tang, R.; Zhang, P.-P.; Cai, J.; Zhou, X.-D.; Jiang, D.-P.; Fei, X.-C.; et al. A cohort autopsy study defines COVID-19 systemic pathogenesis. Cell Res. 2021, 31, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, N.; Jimmerson, L.C.; Bushman, L.R.; Kiser, J.J.; Aron, G.; Reusken, C.B.E.M.; Koopmans, M.P.G.; van Kampen, J.J.A. Cell-line dependent antiviral activity of sofosbuvir against Zika virus. Antivir. Res. 2017, 146, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Casey, J.R.; Grinstein, S.; Orlowski, J. Sensors and regulators of intracellular pH. Nat. Rev. Mol. Cell Biol. 2010, 11, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, X.; Nguyen, J.; Wu, A.H.; Bleske, B.E.; Zhu, H.-J. Sacubitril Is Selectively Activated by Carboxylesterase 1 (CES1) in the Liver and the Activation Is Affected by CES1 Genetic Variation. Drug Metab. Dispos. 2016, 44, 554–559. [Google Scholar] [CrossRef] [Green Version]
- Fujiyama, N.; Miura, M.; Kato, S.; Sone, T.; Isobe, M.; Satoh, S. Involvement of carboxylesterase 1 and 2 in the hydrolysis of mycophenolate mofetil. Drug Metab. Dispos. 2010, 38, 2210–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackman, R.L.; Ray, A.S.; Hui, H.C.; Zhang, L.; Birkus, G.; Boojamra, C.G.; Desai, M.C.; Douglas, J.L.; Gao, Y.; Grant, D.; et al. Discovery of GS-9131: Design, synthesis and optimization of amidate prodrugs of the novel nucleoside phosphonate HIV reverse transcriptase (RT) inhibitor GS-9148. Bioorg. Med. Chem. 2010, 18, 3606–3617. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.Y.; Wang, T.; Park, Y.; Babusis, D.; Birkus, G.; Xu, Y.; Voitenleitner, C.; Fenaux, M.; Yang, H.; Eng, S.; et al. Nucleotide Prodrug Containing a Nonproteinogenic Amino Acid To Improve Oral Delivery of a Hepatitis C Virus Treatment. Antimicrob. Agents Chemother. 2018, 62, e00620-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, A.; Selisko, B.; Le, N.-T.-T.; Huchting, J.; Touret, F.; Piorkowski, G.; Fattorini, V.; Ferron, F.; Decroly, E.; Meier, C.; et al. Rapid incorporation of Favipiravir by the fast and permissive viral RNA polymerase complex results in SARS-CoV-2 lethal mutagenesis. Nat. Commun. 2020, 11, 4682. [Google Scholar] [CrossRef]
- Abdelnabi, R.; Foo, C.S.; De Jonghe, S.; Maes, P.; Weynand, B.; Neyts, J. Molnupiravir Inhibits Replication of the Emerging SARS-CoV-2 Variants of Concern in a Hamster Infection Model. J. Infect. Dis. 2021, 224, 749–753. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme Resource | Study Reference | Sample Size | Assay Method and Quantification Type | Abundance | Abundance Ratio a CatA: CES1 | CatA Contribution to Hydrolysis b | |
|---|---|---|---|---|---|---|---|
| CatA | CES1 | ||||||
| Pooled HLungS9 | [16] | 4 donors, triplicate measurement | Label-free DIA MS-based proteomics, absolute quantification | 0.130 ± 0.006 µg/mg total protein | 1.702 ± 0.063 µg/mg total protein | 1:13 | 97% |
| Pooled HLS9 | 200 donors, triplicate measurement | 0.071 ± 0.004 µg/mg total protein | 19.244 ± 0.083 µg/mg total protein | 1:271 | 64% | ||
| HLung S9 | [13] | n = 3 | Western blot analysis | 0.220 ± 0.004 µg/mg total protein | 2.1 ± 0.9 µg/mg total protein | 1:10 | 98% |
| HLS9 | n = 3 | 0.23 ± 0.11 µg/mg total protein | 34 ± 1 µg/mg total protein | 1:148 | 76% | ||
| Primary human NHBE cells | n = 3 | 0.33 ± 0.10 µg/mg total protein | <0.1µg/mg total protein | >3.3:1 | >99% | ||
| Primary human TBEC cells | [27] | n = 4 | Label-free DDA MS-based proteomics, absolute quantification | 0.056 ± 0.013 µg/mg total protein | 0.019 ± 0.017 µg/mg total protein | 3:1 | >99% |
| Primary human hepatocytes | [28] | n = 7 | 0.059 ± 0.014 µg/mg total protein | 13.7 ± 3.0 µg/mg total protein | 1:232 | 67% | |
| Lung tissue autopsy from COVID-19 patients | [26] | n = 30 | Labeled DDA MS-based proteomics, relative quantification | 1.501 ± 0.878 | 0.249 ± 0.069 | - | - |
| Liver tissue autopsy from COVID-19 patients | n = 24 | 1.308 ± 0.300 | 3.465 ± 1.078 | - | - | ||
| A549 cells (human pneumocyte type II carcinoma cells) | [29] | n = 3 | Labeled DDA MS-based proteomics, relative quantification | 0.765 | 0.797 | - | - |
| Huh-7 cells (human hepatocellular carcinoma cells) | n = 3 | 1.11 | 13.96 | - | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Liu, S.; Shi, J.; Zhu, H.-J. Activation of Tenofovir Alafenamide and Sofosbuvir in the Human Lung and Its Implications in the Development of Nucleoside/Nucleotide Prodrugs for Treating SARS-CoV-2 Pulmonary Infection. Pharmaceutics 2021, 13, 1656. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13101656
Li J, Liu S, Shi J, Zhu H-J. Activation of Tenofovir Alafenamide and Sofosbuvir in the Human Lung and Its Implications in the Development of Nucleoside/Nucleotide Prodrugs for Treating SARS-CoV-2 Pulmonary Infection. Pharmaceutics. 2021; 13(10):1656. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13101656
Chicago/Turabian StyleLi, Jiapeng, Shuhan Liu, Jian Shi, and Hao-Jie Zhu. 2021. "Activation of Tenofovir Alafenamide and Sofosbuvir in the Human Lung and Its Implications in the Development of Nucleoside/Nucleotide Prodrugs for Treating SARS-CoV-2 Pulmonary Infection" Pharmaceutics 13, no. 10: 1656. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13101656