High Intensity Violet Light (405 nm) Inactivates Coronaviruses in Phosphate Buffered Saline (PBS) and on Surfaces


1
Institute of Medical Engineering and Mechatronics, Ulm University of Applied Sciences, 89075 Ulm, Germany
2
MICROMUN, Privates Institut für Mikrobiologische Forschung GmbH, 17489 Greifswald, Germany
*
Author to whom correspondence should be addressed.
Photonics 2021, 8(10), 414; https://0-doi-org.brum.beds.ac.uk/10.3390/photonics8100414
Submission received: 7 September 2021
/
Revised: 24 September 2021
/
Accepted: 26 September 2021
/
Published: 28 September 2021
(This article belongs to the Special Issue Light-Based Technologies and Spectroscopic Techniques for Photo-Sensing and Photoinactivation of Microorganisms, Virus, and Cancer Cells)
Abstract
:It has been proven that visible light with a wavelength of about 405 nm exhibits an antimicrobial effect on bacteria and fungi if the irradiation doses are high enough. Hence, the question arises as to whether this violet light would also be suitable to inactivate SARS-CoV-2 coronaviruses. Therefore, a high-intensity light source was developed and applied to irradiate bovine coronaviruses (BCoV), which are employed as SARS-CoV-2 surrogates for safety reasons. Irradiation is performed in virus solutions diluted with phosphate buffered saline and on steel surfaces. Significant virus reduction by several log levels was observed both in the liquid and on the surface within half an hour with average log reduction doses of 57.5 and 96 J/cm2, respectively. Therefore, it can be concluded that 405 nm irradiation has an antiviral effect on coronaviruses, but special attention should be paid to the presence of photosensitizers in the virus environment in future experiments. Technically, visible violet radiation is therefore suitable for coronavirus reduction, but the required radiation doses are difficult to achieve rapidly.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The coronavirus SARS-CoV-2 that causes the new severe acute respiratory syndrome infection Covid-19 was first detected in December 2019 and has since spread around the world [1]. Nearly two years later, over 200 million infections have been diagnosed, resulting in approximately 4.5 million deaths to date [2]. Despite the availability of vaccines, the pandemic has not yet been ended. Therefore, personal protective equipment and disinfection measures should continue to stop or at least slow the further spread of the virus. The use of ultraviolet radiation, for example, is very effective against SARS-CoV-2 [3,4,5,6,7], but unfortunately this radiation can also harm human cells [8,9,10].
Therefore, a radiation that inactivates viruses but poses little danger to human cells is desirable. The recently often mentioned Far-UVC in the spectral range 200–230 nm could be a future solution [11,12], but suitable radiation sources are still relatively expensive and difficult to obtain, and the long-term effects on human health are still unknown.
Visible blue and violet light is generally harmless to humans, but capable of inactivating bacteria including the dreaded ESKAPE pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.), if irradiation doses are high enough [13,14,15]. The mechanism of action here was based on endogenous photosensitizers such as porphyrins and flavins, which absorb this light and subsequently generate intracellular reactive oxygen species (ROS) that attack membranes, DNA (deoxyribonucleic acid), and proteins inside the cell, destroying them [16,17,18,19,20,21,22].
There were little data on the effects of visible light on viruses prior to the coronavirus pandemic. Tomb et al. found that much higher irradiation doses were required to inactivate viruses in medium with violet light than for bacteria, and in doing so, the result was also highly dependent on the medium [14]. The required irradiation doses for a log-level reduction (90% reduction) are lower in an organically rich medium than in a minimal medium. This can be explained by the fact that organically rich media contain more ROS generating photosensitizers, which then provide external destruction of the viruses. This is supported by Kingsley et al., who observed a much stronger virus inactivation by violet light when riboflavin was added to the medium [23].
These relatively high required irradiation doses—compared to bacteria—are in agreement with our own studies on the enveloped RNA (ribonucleic acid) phage phi 6 [24,25]. Here, about 400 and 2100 J/cm2 were observed for a log reduction of phi6 in PBS (phosphate buffered saline) at 405 and 450 nm irradiation, respectively.
However, published reports exist [26,27] that SARS-CoV-2, in particular, is quite sensitive to visible violet light, with D90 doses (90% reduction dose) in the range of only a few J/cm2, which would correspond to a much higher sensitivity than that reported in bacteria. This would be highly encouraging for the containment of the coronavirus pandemic, but it must be ensured that these results do not depend on the media applied during the irradiation. Typical cell culture media do not contain porphyrins, which play a major role in the photoinactivation of bacteria by violet light, but among the many ingredients—even in MEM (minimal essential medium)—there is riboflavin, another photosensitizer, which absorbs violet and blue light and generates ROS. Therefore, viruses that are irradiated in fresh cell culture medium or insufficiently diluted cell culture medium, might be inactivated faster as in a photosensitizer-free solution.
Therefore, in the study presented here, coronavirus experiments are performed in PBS (phosphate buffered saline) and on the surfaces that are both mostly free of medium and external photosensitizers. For the irradiation, a strong 405 nm LED light source was developed, which allows high irradiation doses to be achieved in a relatively short time.
2. Materials and Methods
2.1. High Intensity Light Source
Irradiation should be performed by a strong 405 nm LED light source to allow for the application of high irradiation doses of up to 1000 J/cm2 within hours. Therefore, a device was designed for the irradiation of liquid and solid samples preferably kept in a Petri dish with a diameter of up to 80 mm. The Petri dish is located on a glass plate in the center of an approximately cubic chamber made of aluminum boards with two of them being removable doors for loading and cleaning (Figure 1). The bottom and the lid of the specimen chamber are each equipped with an array consisting of 8 × 8 LEDs (type NVSU119CT of Nichia, Tokushima, Japan). Thus, the specimen is irradiated from two sides, which is helpful for translucent (e.g., textile) or opaque specimens. Two glass plates in front of the LED arrays protect the LEDs from dirt and mechanical damage.
These LEDs emit violet light with a nominal peak wavelength of 405 nm and 12 nm spectral half width. The measured relative LED emission spectrum with an actual peak wavelength of 406 nm is given in Figure 2. At its nominal current of 0.7 A, it produces a radiant flux of about 1.4 W with a radiation angle of 144° at 50% radiant intensity. The LEDs were purchased on small metal core printed circuit boards (PCB) and these PCBs were grouped to form a 95.4 × 95.4 mm2 array. These were connected with copper straps type ZZZIP-band of Wieland (Voehringen, Germany) to form strings with eight LEDs and a 3.9 Ω balancing resistor each. With 16 LED strings in parallel, the LED arrays can be driven with a current of up to 11 A and 280 W electrical LED power, generating 180 W radiant flux. The irradiation device was operated by a laboratory power supply type NPS306W of Wanptek (Shenzhen, China) with a maximum current of 6 A. Under such conditions, thermal management of the whole device is critical. The waste heat of the LEDs is removed by cooling fins and blowers. PTC sensors and a modified cutout circuit of Pollin (Pfoerring, Germany) prevent overheating of the LEDs.
The irradiation time can be controlled by an electronic timer of KKmoon (Lakewood, CA, USA). This timer and the thermal protection circuit need a supply voltage of 12 V. As the LED string current starts at about 20 V string voltage, a DC/DC stepdown converter of Pollin was connected in parallel to the strings.
The irradiance of the specimen depends on the LED current, which was measured with a digital multimeter. This current also included the current of the electronics via the DC/DC converter, which decreases with increasing string voltage. This fact is incorporated in the radiometric calibration of the device. Additionally incorporated is the absorption of the glass plates in the specimen chamber and the bottom and lid of the Steriplan Petri dish of VWR/Avantor (Darmstadt, Germany).
The specimen plane was considered to be 2 mm above the bottom surface of the Petri dish. The Petri dish was located so that this specimen plane was 36 mm away from the lid and from the bottom of the specimen chamber. The irradiance in this specimen plane from the upper and the lower LED arrays was measured with a photodiode BPW21R. This photodiode was operated in reverse direction with a 9 V bias voltage and a 1.5 KΩ resistor. For absolute radiometric measurements, this arrangement was calibrated with a NVSU119CT array and an optical power meter type OPM150 UVS of Artifex (Emden, Germany) and the data fitted to a second order polynomial on the electrical current I with the resulting correlation irradiance = (0.1042 I2 + 67.496 I − 25.018 A) mW/cm2 (R2 = 1).
In the case of liquid or translucent specimens, the total irradiance is the sum of the top and the bottom irradiance. The irradiance from the bottom was about 8% lower than from the top, because the light has to cross one more glass plate. The maximum achievable 405 nm intensity of this setup is 350 mW/cm2 and is homogeneous within a few percent in an area of 80 × 80 mm2. However, to avoid heating of the liquid samples to temperatures above 40 °C, the doors have to be open and the current has to be limited to 3 A, resulting in an irradiance of 178.4 mW/cm2. Most experiments were performed at a current of only 2.5 A, which corresponds to 144.4 mW/cm2.
2.2. Virus Preparation
Due to laboratory safety restrictions, the experiment was not performed with the human coronavirus SARS-CoV-2, but a bovine coronavirus (BCoV), a betacoronavirus like SARS-COV-2, was used as a surrogate. The selected BCoV-strain was “S379 Riems” of the Friedrich–Loeffler–Institute (Greifswald, Germany) with the Madin–Darby bovine kidney cell (MDBK) strain “Riems 261”, also from the Friedrich–Loeffler–Institute as host cells. The cell culture medium was RPMI (Roswell Park Memorial Institute) 1640. With these hosts, bovine coronaviruses were proliferated to concentrations of about 4.5 log TCID50 (50% tissue culture infection dose).
For the experiments in solutions, the coronaviruses were first proliferated by the MDBK cells in fresh RPMI. At the end of cultivation, the medium and its components are largely consumed. The virus lysate clarified by centrifugation is then diluted 1:10 in PBS and 3 mL portions transferred to small transparent Petri dishes, leading to a liquid sample height of about 4 mm. A sensor was placed in 3 mL of pure PBS in an additional Petri dish to record the temperature in the samples during the irradiation, which was performed for up to 40 min with 144.4 mW/cm2. Unirradiated virus samples, which were kept at 43 °C for 60 min, served as the control for the temperature influence. Virus concentrations of the irradiated und unirradiated samples were determined by the standard plaque assay with the MDBK cells. All experiments were performed in triplicate.
The surface experiments were performed on steel coupons. A total of 100 µL virus suspension was dripped on to each of the three steel slides and dried. The loaded slides were either placed in a Petri dish with the loaded side up or down and irradiated for 30 min. As a control, three loaded virus carriers were kept in a Petri dish at 37 °C for 30 min. Subsequently, the viruses were detached from the carriers by placing them in a 10 mL cell culture medium and shaking briefly, and titrated onto the MDBK cells so that the concentration could be determined by a plaque assay. This procedure was performed twice but in each run with three samples facing upward and three downward. A sensor below a Petri dish recorded the temperature during these experiments.
3. Results
The recorded temperature in the virus samples increased to a maximum of 41 °C in the 40 min of irradiation. The liquid controls were not irradiated, but heated to a temperature of about 43 °C. A virus titer reduction was not observed in these references, even after 60 min of incubation. Therefore, it is assumed that even for the hottest temperature of 41 °C during irradiation, thermal virus inactivation can be neglected.
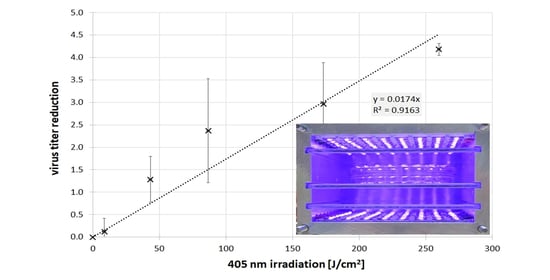
The results of the BCoV concentration reduction by the 405 nm irradiation are illustrated in Figure 3 with a logarithmic scale on the y-axis. The maximum irradiation on the x-axis was about 260 J/cm2, which corresponds to a 30 min irradiation, because there were no viruses left after 40 min. The data points are the averages of the single measurements and the error bars give the standard deviation of the individual measurements. A straight line with a slope of 0.174 log/(J/cm2) was fitted through the data. This is equivalent to an exponential BCoV concentration decrease in the solution with an average log reduction dose of 57.5 J/cm2.
For the unirradiated steel coupons, no virus reduction was detected after a period of 30 min. However, for the irradiated coupons, an average titer log reduction of 2.13 ± 0.8 was observed after the same period of time. These samples were only irradiated from one side, but if a reflection of about 57.5% at 405 nm is assumed [28], 30 min would mean an irradiation dose of about 205 J/cm2 for viruses on the steel surface. The resulting log reduction dose for this bovine coronavirus on steel is therefore approximately 96 J/cm2.
4. Discussion and Conclusions
For both media, in solutions and on surfaces, there is a clear coronavirus photoinactivation of several log levels, with a progression that appears to be approximately exponential, at least in the solution.
As for the required log reduction doses, the virus appears to be more sensitive in liquids than on surfaces. However, it cannot be ruled out that the effective irradiance on the steel coupons was assumed too high. Viruses in pores, scratches, or other irregularities might have been partially protected from the radiation, so that the effective irradiation dose was lower than expected.
For safety reasons, the described experiments were performed with bovine coronaviruses and not the more important SARS-CoV-2 because there have been several reported SARS-CoV accidents in medical laboratories in the past [29]. Nevertheless, though there seem to be large differences between cattle and humans, the differences between BCoV and SARS-CoV-2 appear to be rather small. Both are members of the genus betacoronavirus within the family Coronoviridae [30]. BCoV exhibits a similar size, RNA-strand length, and protein surface structure as SARS-CoV-2 [30,31,32,33] and it is even believed to be the ancestor of the human coronavirus HCoV-OC43 with whom it shares 95% of its RNA [34]. Additionally, the clinical respiratory tract infections show similarities [30]. Because of all these resemblances, BCoV has already been suggested and successfully employed as a SARS-CoV(-2) surrogate in many different applications [35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53].
As this is the first study of the effect of 405 nm irradiation on bovine coronaviruses, there is no evidence that BCoV and SARS-CoV-2 behave similarly in this regard. Nevertheless, because of the many resemblances and the same virus structure, we expect that both viruses are similarly sensitive to violet light and that the observed BCoV results reflect the SARS-CoV-2 behavior well.
The required log irradiation doses for this bovine coronavirus noted here were about an order of magnitude higher than results for SARS-CoV-2 [26,27] and about a factor of 2.5 higher than the results for feline infectious peritonitis virus, another animal coronavirus [54]. However, our determined log reduction doses seem to be roughly in the same order of magnitude than the recent results of Enwemeka et al., who irradiated the human coronaviruses HCoV-229E and HCoV-OC43 with pulsed violet and blue light and observed a 2.37 and 1.46 log reduction with pulsed 405 nm irradiation, respectively [55]. This would correspond to log reduction doses of 54.8 and 89.0 J/cm2.
All these 405 nm doses are quite low compared to the values for other viruses listed in [14] with log reduction doses between 84 and 1 020 J/cm2, depending on the applied medium, or compared to our own previous 405 nm results of 400 J/cm2 for the phage phi6, which is an enveloped RNA virus like the coronaviruses.
Especially in light of the results of Tomb et al. [14] and the observed influence of the medium, it cannot be excluded that the applied cell culture media, which all contain photosensitizers such as riboflavin might lead to seemingly reduced log reduction doses by generating reactive oxygen species that attack the coronavirus from the outside. The statements in the published reports regarding the medium content of the virus solution are not always clear. At least one author mentioned that their coronaviruses were actually irradiated in cell culture medium and another author claimed to have performed the irradiation in PBS, but according to the explicit experimental description, PBS and medium were mixed.
The irradiation in our own experiment was performed on viruses in used cell culture medium, in which the components including photosensitizers such as riboflavin should have been largely consumed and the solution was further diluted 1:10 in PBS prior to the irradiation. Therefore, we assumed the photosensitizer concentration to be negligible, but we cannot guarantee that it was totally free of external photosensitizers.
Different concentrations of external photosensitizers in the irradiated virus solution might at least partially cause the large variability of the coronavirus results and we suggest reducing the percentage of medium in the irradiated samples as much as possible, if the intention is to obtain “real world” log reduction doses in a photosensitizer free environment.
Potential “real world applications” of 405 nm irradiation are conceivable in health care or community environments (e.g., the decontamination of rooms, surfaces, liquids, and equipment). This is not as fast as it would be possible with 254 nm UVC irradiation, but in contrast to UVC, it offers the advantage of not being harmful to humans, and does not lead to an accelerated aging or bleaching of sensitive materials.
A 405 nm irradiation could be applied alone, as suggested by Maclean et al. [56] or it could be combined with a white illumination [57], even in the presence of people. In the presented experiments, a 4 log reduction was achieved within half an hour, but this is a high intensity irradiation, which is difficult to achieve for larger volumes or areas. A tenth of this irradiation intensity seems to be more realistic, though still ambitious, and this would lead to irradiation duration in the region of 1 h or even longer for a log reduction.
Therefore, 405 nm irradiation is capable of coronavirus inactivation, though it is not the fastest method. Nevertheless, with the much lower risk to humans and sensitive materials, it exhibits an important advantage compared to UVC irradiation.
Author Contributions
Conceptualization, B.L., D.B. and M.H.; Methodology, B.L., D.B. and M.H.; Validation, B.L., D.B. and M.H.; Investigation, D.B.; Resources, B.L.; Data curation, B.L., D.B. and M.H.; Writing—original draft preparation, B.L., D.B. and M.H.; Writing—review and editing, B.L., D.B. and M.H.; Project administration, B.L. and M.H.; Funding acquisition, B.L. All authors have read and agreed to the published version of the manuscript.
Funding
We are very grateful for the financial support by the Else-Kröner-Fresenius-Stiftung (2020_EKKP.140). The article processing charge was funded by the Baden–Württemberg Ministry of Science, Research, and Culture, and the Ulm University of Applied Sciences in the funding program Open Access Publishing.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The detailed data and the irradiation device can be obtained from the authors upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, L.; Wang, Y.; Ye, D.; Liu, Q. Review of the 2019 novel coronavirus (SARS-CoV-2) based on current evidence. Int. J. Antimicrob. Agents 2020, 55, 105948. [Google Scholar] [CrossRef]
- Coronavirus Resource Center. COVID-19 Dashboard: (Global Map). 2021. Available online: https://0-coronavirus-jhu-edu.brum.beds.ac.uk/map.html (accessed on 29 August 2021).
- Hessling, M.; Hönes, K.; Vatter, P.; Lingenfelder, C. Ultraviolet irradiation doses for coronavirus inactivation—review and analysis of coronavirus photoinactivation studies. GMS Hyg. Infect. Control 2020, 15. [Google Scholar] [CrossRef]
- Chiappa, F.; Frascella, B.; Vigezzi, G.P.; Moro, M.; Diamanti, L.; Gentile, L.; Lago, P.; Clementi, N.; Signorelli, C.; Mancini, N.; et al. The efficacy of ultraviolet light-emitting technology against coronaviruses: A systematic review. J. Hosp. Infect. 2021, 114, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, H.; Matsuda, J.; Iwasaki, T.; Hayasaka, D. Efficacy of 265-nm ultraviolet light in inactivating infectious SARS-CoV-2. J. Photochem. Photobiol. 2021, 7, 100050. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, H.; Saito, A.; Kaneko, C.; Sugiyama, H.; Okabayashi, T.; Fujimoto, S. Rapid Inactivation of SARS-CoV-2 Variants by Continuous and Intermittent Irradiation with a Deep-Ultraviolet Light-Emitting Diode (DUV-LED) Device. Pathogens 2021, 10, 754. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Linden, Y.S.; Gundy, P.M.; Gerba, C.P.; Sobsey, M.D.; Linden, K.G. Inactivation of Coronaviruses and Phage Phi6 from Irradiation across UVC Wavelengths. Environ. Sci. Technol. Lett. 2021, 8, 425–430. [Google Scholar] [CrossRef]
- Pfeifer, G.P.; You, Y.-H.; Besaratinia, A. Mutations induced by ultraviolet light. Mutat. Res. 2005, 571, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Delic, N.C.; Lyons, J.G.; Di Girolamo, N.; Halliday, G.M. Damaging Effects of Ultraviolet Radiation on the Cornea. Photochem. Photobiol. 2017, 93, 920–929. [Google Scholar] [CrossRef] [Green Version]
- Scientific Committee on Health, Environmental and Emerging Risks. Opinion on biological effects of UV-C radiation relevant to health with particular reference to UV-C lamps. In Proceedings of the Luxembourg: European Commission, Luxembourg, 3 February 2017. [Google Scholar]
- Hessling, M.; Haag, R.; Sieber, N.; Vatter, P. The impact of far-UVC radiation (200–230 nm) on pathogens, cells, skin, and eyes—A collection and analysis of a hundred years of data. GMS Hyg. Infect. Control 2021, 16. [Google Scholar] [CrossRef]
- International Ultraviolet Association. Far UV-C Radiation: Current State-of Knowledge (White Paper); Ational Ultraviolet Association: Chevy Chase, MD, USA, 2021. [Google Scholar]
- Hessling, M.; Spellerberg, B.; Hoenes, K. Photoinactivation of bacteria by endogenous photosensitizers and exposure to visible light of different wavelengths—A review on existing data. FEMS Microbiol. Lett. 2016, 364, fnw270. [Google Scholar] [CrossRef]
- Tomb, R.M.; White, T.A.; Coia, J.E.; Anderson, J.G.; MacGregor, S.J.; Maclean, M. Review of the Comparative Susceptibility of Microbial Species to Photoinactivation Using 380-480 nm Violet-Blue Light. Photochem. Photobiol. 2018, 94, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Hoenes, K.; Bauer, R.; Meurle, T.; Spellerberg, B.; Hessling, M. Inactivation Effect of Violet and Blue Light on ESKAPE Pathogens and Closely Related Non-pathogenic Bacterial Species—A Promising Tool Against Antibiotic-Sensitive and Antibiotic-Resistant Microorganisms. Front. Microbiol. 2020, 11, 612367. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, H.; Malik, Z.; Harth, Y.; Nitzan, Y. Eradication of Propionibacterium acnes by its endogenic porphyrins after illumination with high intensity blue light. FEMS Immunol. Med. Microbiol. 2003, 35, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guffey, J.S.; Wilborn, J. In vitro bactericidal effects of 405-nm and 470-nm blue light. Photomed. Laser Surg. 2006, 24, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Maclean, M.; MacGregor, S.J.; Anderson, J.G.; Woolsey, G. High-intensity narrow-spectrum light inactivation and wavelength sensitivity of Staphylococcus aureus. FEMS Microbiol. Lett. 2008, 285, 227–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feuerstein, O.; Ginsburg, I.; Dayan, E.; Veler, D.; Weiss, E.I. Mechanism of visible light phototoxicity on Porphyromonas gingivalis and Fusobacterium nucleatum. Photochem. Photobiol. 2005, 81, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Amin, R.M.; Bhayana, B.; Hamblin, M.R.; Dai, T. Antimicrobial blue light inactivation of Pseudomonas aeruginosa by photo-excitation of endogenous porphyrins: In vitro and in vivo studies. Lasers Surg. Med. 2016, 48, 562–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plavskii, V.Y.; Mikulich, A.V.; Tretyakova, A.I.; Leusenka, I.A.; Plavskaya, L.G.; Kazyuchits, O.A.; Dobysh, I.I.; Krasnenkova, T. Porphyrins and flavins as endogenous acceptors of optical radiation of blue spectral region determining photoinactivation of microbial cells. J. Photochem. Photobiol. B 2018, 183, 172–183. [Google Scholar] [CrossRef]
- Cieplik, F.; Spath, A.; Leibl, C.; Gollmer, A.; Regensburger, J.; Tabenski, L.; Hiller, K.; Maisch, T.; Schmalz, G. Blue light kills Aggregatibacter actinomycetemcomitans due to its endogenous photosensitizers. Clin. Oral Investig. 2014, 18, 1763–1769. [Google Scholar] [CrossRef]
- Kingsley, D.; Kuis, R.; Perez, R.; Basaldua, I.; Burkins, P.; Marcano, A.; Johnson, A. Oxygen-dependent laser inactivation of murine norovirus using visible light lasers. Virol. J. 2018, 15, 117. [Google Scholar] [CrossRef]
- Vatter, P.; Hoenes, K.; Hessling, M. Photoinactivation of the Coronavirus Surrogate phi6 by Visible Light. Photochem. Photobiol. 2021, 97, 122–125. [Google Scholar] [CrossRef]
- Vatter, P.; Hoenes, K.; Hessling, M. Blue light inactivation of the enveloped RNA virus Phi6. BMC Res. Notes 2021, 14, 187. [Google Scholar] [CrossRef] [PubMed]
- Santis, R.; de Luca, V.; Faggioni, G.; Fillo, S.; Stefanelli, P.; Rezza, G.; Lista, F. Rapid inactivation of SARS-CoV-2 with LED irradiation of visible spectrum wavelenghts. MedRxiv 2020. [Google Scholar] [CrossRef]
- Rathnasinghe, R.; Jangra, S.; Miorin, L.; Schotsasert, M.; Yahnke, C.; Garcίa-Sastre, A. Lighting a better future: The virucidal effects of 405 nm visible light on SARS-CoV-2 and influenza A virus. BioRxiv 2021. [Google Scholar] [CrossRef]
- Zwinkels, J.C.; Noël, M.; Dodd, C.X. Procedures and standards for accurate spectrophotometric measurements of specular reflectance. Appl. Opt. 1994, 33, 7933–7944. [Google Scholar] [CrossRef]
- Lim, W.; Ng, K.-C.; Tsang, D.N.C. Laboratory containment of SARS virus. Ann. Acad. Med. Singap. 2006, 35, 354–360. [Google Scholar] [PubMed]
- Saif, L.J.; Jung, K. Comparative Pathogenesis of Bovine and Porcine Respiratory Coronaviruses in the Animal Host Species and SARS-CoV-2 in Humans. J. Clin. Microbiol. 2020. [Google Scholar] [CrossRef]
- Llanes, A.; Restrepo, C.M.; Caballero, Z.; Rajeev, S.; Kennedy, M.A.; Lleonart, R. Betacoronavirus Genomes: How Genomic Information has been Used to Deal with Past Outbreaks and the COVID-19 Pandemic. IJMS 2020, 21, 4546. [Google Scholar] [CrossRef]
- Ghosh, S.; Malik, Y.S. Drawing Comparisons between SARS-CoV-2 and the Animal Coronaviruses. Microorganisms 2020, 8, 1840. [Google Scholar] [CrossRef]
- Alluwaimi, A.M.; Alshubaith, I.H.; Al-Ali, A.M.; Abohelaika, S. The Coronaviruses of Animals and Birds: Their Zoonosis, Vaccines, and Models for SARS-CoV and SARS-CoV2. Front. Vet. Sci. 2020, 7, 582287. [Google Scholar] [CrossRef]
- Vijgen, L.; Keyaerts, E.; Lemey, P.; Maes, P.; van Reeth, K.; Nauwynck, H.; Pensaert, M.; van Ranst, M. Evolutionary history of the closely related group 2 coronaviruses: Porcine hemagglutinating encephalomyelitis virus, bovine coronavirus, and human coronavirus OC43. J. Virol. 2006, 80, 7270–7274. [Google Scholar] [CrossRef] [Green Version]
- Franke, G.; Knobling, B.; Brill, F.H.; Becker, B.; Klupp, E.M.; Belmar Campos, C.B.; Pfefferle, S.; Lütgehetmann, M.; Knobloch, J.K. An automated room disinfection system using ozone is highly active against surrogates for SARS-CoV-2. J. Hosp. Infect. 2021, 112, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, S.A.; Zahran, E.M.; Abdel Fadeel, M.R.; Albohy, A.; Safwat, M.A. New Acaciin-Loaded Self-Assembled Nanofibers as MPro Inhibitors Against BCV as a Surrogate Model for SARS-CoV-2. Int. J. Nanomed. 2021, 16, 1789–1804. [Google Scholar] [CrossRef] [PubMed]
- Kenney, S.P.; Wang, Q.; Vlasova, A.; Jung, K.; Saif, L. Naturally Occurring Animal Coronaviruses as Models for Studying Highly Pathogenic Human Coronaviral Disease. Vet. Pathol. 2021, 58, 438–452. [Google Scholar] [CrossRef] [PubMed]
- Salvo, M.; Moller, A.; Alvareda, E.; Gamazo, P.; Colina, R.; Victoria, M. Evaluation of low-cost viral concentration methods in wastewaters: Implications for SARS-CoV-2 pandemic surveillances. J. Virol. Methods 2021, 297, 114249. [Google Scholar] [CrossRef]
- Barrios, M.E.; Díaz, S.M.; Torres, C.; Costamagna, D.M.; Blanco Fernández, M.D.; Mbayed, V.A. Dynamics of SARS-CoV-2 in wastewater in three districts of the Buenos Aires metropolitan region, Argentina, throughout nine months of surveillance: A pilot study. Sci. Total Environ. 2021, 800, 149578. [Google Scholar] [CrossRef]
- Todt, D.; Meister, T.L.; Tamele, B.; Howes, J.; Paulmann, D.; Becker, B.; Brill, F.H.; Wind, M.; Schijven, J.; Heinen, N.; et al. A realistic transfer method reveals low risk of SARS-CoV-2 transmission via contaminated euro coins and banknotes. IScience 2021, 24, 102908. [Google Scholar] [CrossRef]
- Yoshizawa, N.; Ishihara, R.; Omiya, D.; Ishitsuka, M.; Hirano, S.; Suzuki, T. Application of a Photocatalyst as an Inactivator of Bovine Coronavirus. Viruses 2020, 12, 1372. [Google Scholar] [CrossRef] [PubMed]
- Wensman, J.J.; Stokstad, M. Could Naturally Occurring Coronaviral Diseases in Animals Serve as Models for COVID-19? A Review Focusing on the Bovine Model. Pathogens 2020, 9, 991. [Google Scholar] [CrossRef]
- Lin, C.-H.; Yang, C.-Y.; Ou, S.-C.; Wang, M.; Lo, C.-Y.; Tsai, T.-L.; Wu, H.-Y. The Impacts of Antivirals on the Coronavirus Genome Structure and Subsequent Pathogenicity, Virus Fitness and Antiviral Design. Biomedicines 2020, 8, 376. [Google Scholar] [CrossRef]
- Ma, C.; Gong, C. ACE2 models of frequently contacted animals provide clues of their SARS-CoV-2 S protein affinity and viral susceptibility. J. Med. Virol. 2021, 93, 4469–4479. [Google Scholar] [CrossRef]
- Mullis, L.; Saif, L.J.; Zhang, Y.; Zhang, X.; Azevedo, M.S.P. Stability of bovine coronavirus on lettuce surfaces under household refrigeration conditions. Food Microbiol. 2012, 30, 180–186. [Google Scholar] [CrossRef]
- Kapil, S.; Oberst, R.; Bieker, J.; Tucker, M.; Souza, C.; Williams, C. Rapid Inactivation of SARS-Like Coronaviruses; Sandia National Laboratories: Albuquerque, NM, USA, 2004. [Google Scholar]
- Steinmann, J. Surrogate viruses for testing virucidal efficacy of chemical disinfectants. J. Hosp. Infection. 2004, 56, S49–S54. [Google Scholar] [CrossRef] [PubMed]
- Kratzel, A.; Todt, D.; V′kovski, P.; Steiner, S.; Gultom, M.; Thao, T.T.N.; Ebert, N.; Holwerda, M.; Steinmann, J.; Niemeyer, D.; et al. Inactivation of Severe Acute Respiratory Syndrome Coronavirus 2 by WHO-Recommended Hand Rub Formulations and Alcohols. Emerg. Infect. Dis. 2020, 26, 1592–1595. [Google Scholar] [CrossRef] [PubMed]
- Karthigeyan, K.P.; Flanigan, C.; Machado, D.J.; Kiziltas, A.A.; Janies, D.A.; Chen, J.; Cooke, D.; Lee, M.V.; Saif, L.J.; Henegar, S.; et al. Heat efficiently inactivates coronaviruses inside vehicles. BioRxiv 2021. [Google Scholar] [CrossRef]
- Graham, K.E.; Loeb, S.K.; Wolfe, M.K.; Catoe, D.; Sinnott-Armstrong, N.; Kim, S.; Yamahara, K.M.; Sassoubre, L.M.; Grijalva, L.M.M.; Roldan-Hernandez, L.; et al. SARS-CoV-2 RNA in Wastewater Settled Solids Is Associated with COVID-19 Cases in a Large Urban Sewershed. Env. Sci. Technol. 2021, 55, 488–498. [Google Scholar] [CrossRef]
- Gonzalez, R.; Curtis, K.; Bivins, A.; Bibby, K.; Weir, M.H.; Yetka, K.; Thompson, H.; Keeling, D.; Mitchell, J.; Gonzalez, D. COVID-19 surveillance in Southeastern Virginia using wastewater-based epidemiology. Water Res. 2020, 186, 116296. [Google Scholar] [CrossRef] [PubMed]
- Kwiek, J.J.; Pickett, C.R.; Flanigan, C.A.; Lee, M.V.; Saif, L.J.; Jahnes, J.; Blonder, G. A practical PPE decontamination method using warm air and ambient humidity. BioRxiv 2020. [Google Scholar] [CrossRef]
- Lucassen, R.; Weide, M.; Bockmühl, D. Virucidal Efficacy of Household Dishwashers. Microbiol. Res. 2021, 12, 395–402. [Google Scholar] [CrossRef]
- Gardner, A.; Ghosh, S.; Dunowska, M.; Brightwell, G. Virucidal Efficacy of Blue LED and Far-UVC Light Disinfection against Feline Infectious Peritonitis Virus as a Model for SARS-CoV-2. Viruses 2021, 13, 1436. [Google Scholar] [CrossRef]
- Enwemeka, C.S.; Bumah, V.V.; Mokili, J.L. Pulsed blue light inactivates two strains of human coronavirus. J. Photochem. Photobiol. B 2021, 222, 112282. [Google Scholar] [CrossRef] [PubMed]
- Maclean, M.; Macgregor, S.J.; Anderson, J.G.; Woolsey, G.A.; Coia, J.E.; Hamilton, K.; Taggart, I.; Watson, S.B.; Thakker, B.; Gettinby, G. Environmental decontamination of a hospital isolation room using high-intensity narrow-spectrum light. J. Hosp. Infect. 2010, 76, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Buehler, J.; Sommerfeld, F.; Meurle, T.; Hoenes, K.; Hessling, M. Disinfection Properties of Conventional White LED Illumination and Their Potential Increase by Violet LEDs for Applications in Medical and Domestic Environments. Adv. Sci. Technol. Res. J. 2021, 15, 169–175. [Google Scholar] [CrossRef]
Figure 1.
Setup of the high intensity 405 nm irradiation device.

Figure 2.
Relative emission spectrum of the employed Nichia LED at a total current of 2.5 A or 156 mA per LED with a measured peak wavelength of 406 nm.
Figure 2.
Relative emission spectrum of the employed Nichia LED at a total current of 2.5 A or 156 mA per LED with a measured peak wavelength of 406 nm.

Figure 3.
Log reduction of BCoV titer in PBS during 405 nm irradiation. The error bars represent the standard deviation of the individual measurements.
Figure 3.
Log reduction of BCoV titer in PBS during 405 nm irradiation. The error bars represent the standard deviation of the individual measurements.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lau, B.; Becher, D.; Hessling, M. High Intensity Violet Light (405 nm) Inactivates Coronaviruses in Phosphate Buffered Saline (PBS) and on Surfaces. Photonics 2021, 8, 414. https://0-doi-org.brum.beds.ac.uk/10.3390/photonics8100414
AMA Style
Lau B, Becher D, Hessling M. High Intensity Violet Light (405 nm) Inactivates Coronaviruses in Phosphate Buffered Saline (PBS) and on Surfaces. Photonics. 2021; 8(10):414. https://0-doi-org.brum.beds.ac.uk/10.3390/photonics8100414
Chicago/Turabian StyleLau, Bernhard, Dietmar Becher, and Martin Hessling. 2021. "High Intensity Violet Light (405 nm) Inactivates Coronaviruses in Phosphate Buffered Saline (PBS) and on Surfaces" Photonics 8, no. 10: 414. https://0-doi-org.brum.beds.ac.uk/10.3390/photonics8100414
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.