
From Genetic Maps to QTL Cloning: An Overview for Durum Wheat




1
Department of Agricultural and Environmental Science, University of Bari ‘Aldo Moro’, Via G. Amendola 165/A, 70126 Bari, Italy
2
Sustainable Field Crops Programme, IRTA (Institute for Food and Agricultural Research and Technology), 25198 Lleida, Spain
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Plants 2021, 10(2), 315; https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020315
Submission received: 11 January 2021
/
Revised: 26 January 2021
/
Accepted: 2 February 2021
/
Published: 6 February 2021
(This article belongs to the Special Issue Genetic Improvement of Cereal Crops for Resistance to Biotic and Abiotic Stresses)
Abstract
:Durum wheat is one of the most important cultivated cereal crops, providing nutrients to humans and domestic animals. Durum breeding programs prioritize the improvement of its main agronomic traits; however, the majority of these traits involve complex characteristics with a quantitative inheritance (quantitative trait loci, QTL). This can be solved with the use of genetic maps, new molecular markers, phenotyping data of segregating populations, and increased accessibility to sequences from next-generation sequencing (NGS) technologies. This allows for high-density genetic maps to be developed for localizing candidate loci within a few Kb in a complex genome, such as durum wheat. Here, we review the identified QTL, fine mapping, and cloning of QTL or candidate genes involved in the main traits regarding the quality and biotic and abiotic stresses of durum wheat. The current knowledge on the used molecular markers, sequence data, and how they changed the development of genetic maps and the characterization of QTL is summarized. A deeper understanding of the trait architecture useful in accelerating durum wheat breeding programs is envisioned.
Keywords:
durum wheat; genetic mapping; QTL; GWAS; fine mapping; positional cloning; quality; abiotic stress; biotic stress1. Introduction
The United Nations expects the world’s population to grow from seven to nine billion by 2050 (https://www.un.org/development/desa/en/news/population/world-population-prospects-2019.html; accessed on 25 January 2021). Crop production needs to satisfy future demands while facing extreme climate changes and hurdles in natural resources (such as water and soil) and disease management (https://www.fao.org; accessed on 24 July 2020). The main challenge faced by plant scientists in the 21st century is to increase the crop productivity per unit area and, at the same time, enhance sustainability and preserve biodiversity (European Plant Science Organization, https://epsoweb.org/; accessed on 24 July 2020). These have important implications on the breeding efforts and strategies for cereals, particularly wheat, which is a main source for food consumption worldwide.
Although common wheat (Triticum aestivum L.) is cultivated globally, durum wheat (Triticum durum Desf.) represents about 5% of the total wheat production, with almost 17 million ha worldwide and a global production of 38 million tons in 2019 (https://www.agr.gc.ca/; accessed on 25 January 2021), and is primarily cultivated in three different regions: the Mediterranean basin, the Northern Plains between the United States and Canada, and the desert areas of the south west of the United States and Mexico [1], characterized by a mix of variable and harsh environmental conditions (rainy weather, dry winters, and hot summers). Durum semolina is used for many products, including pasta, couscous, bread, and bulgur. Understanding the genetic basis of important agronomic traits in durum wheat is key for breeding programs. A significant yield increase was achieved with the introduction of semi-dwarf cultivars. Durum wheat germplasm developed by international Consultative Group on International Agricultural Research (CGIAR) centers (CIMMYT and ICARDA) were the most widely used by national programs worldwide. Besides these institutions, breeding programs in Italy were also very relevant from the beginning of the 20th century. According to Royo et al. [2] studying Spanish and Italian durum wheat cultivars from different periods, grain yield improvement was based on increasing the number of grains per unit area and harvest index, whereas grain weight did not change with the breeding process. In a similar work studying pasta quality, Subirà et al. [3] found that although protein content per grain was reduced, the yield increase produced by the new cultivars augmented the protein yield per ha. Other quality traits that increased significantly were the yellow index and gluten strength, obtaining very favorable allele combinations for high and low molecular weight glutenin subunits (HMW and LMW). The most important agronomic traits of cultivated plants are quantitative traits controlled by several genes across the genome and often influenced by the environmental conditions. The identification of major regions in the genome controlling these traits, quantitative trait loci (QTL), offers the opportunity to track them with molecular markers through molecular-assisted selection. Thus far, genetic maps and phenotypic data from segregating populations have allowed researchers to localize and map important genes, and to identify closely associated markers for marker-assisted selection (MAS) and, eventually, positional cloning [4,5]. However, the selection steps have been labor intensive and sometimes elusive, hindered by the development of extremely large mapping populations, non-specific molecular markers, large wheat genome size, and the lack of sequence information.
The identification of durum wheat QTL focuses on its main agronomic traits, including the protein grain content [6], high grain yield [7], disease resistance [8,9], and quality traits [10]. The studies used molecular markers such as simple sequence repeats (SSRs), expressed sequence tags (ESTs), and restriction fragment length polymorphism (RFLP) markers [11]. Standard mapping procedure through these DNA markers allowed for the identification of regions of several centiMorgans (cM) on the genetic maps, indicating the involvement of main genes. This non-specificity makes it difficult to find the key locus responsible for a specific trait, and the bystander effect of unwanted genomic regions is also difficult to control.
In the last decade, DNA sequencing technologies and applications including the discovery of new types of molecular markers have significantly improved plant breeding and assisted fine mapping procedures. With the completion of the Durum Wheat Genome Project [12], the scientific community examined the whole-genome sequences and all the data available from the studies regarding genes and QTL locations, the common variations found in genetic polymorphism sites, and other information that provides a large number of single nucleotide polymorphisms (SNPs) in the whole genome. Coupled with the rapid development of high-throughput genotypic technology, intense fine mapping analysis has been conducted in both a time- and cost-effective manner.
In the present review, we summarize the recent studies regarding the fine-map QTL, cloning QTL/genes, and identification of candidate genes for the main quality traits as well as biotic and abiotic stress resistance in recent years in durum wheat. The results presented can be very useful to program future studies and to identify major and stable QTL that can be considered to be cloned or further investigated by researchers for future applications in plant breeding programs.
In recent years, durum wheat research has undergone considerable expansion due to various agronomic, genetic, and commercial factors. The importance of durum wheat end-products in the food chain makes it crucial to maintain or increase durum wheat production under disease pressure and adverse climatic conditions but while preserving the grain quality. Thus, the grain quality and agronomic traits are also equally significant in determining the quality and yields of the processed products.
Here, we report the research into the improvement of wheat traits focused on the grain/flour quality of the breeding lines, environmental effects, disease resistance, development of evaluation methods, and processing for end uses [13].
2. Genotyping Tools
In the early 1990s, many agronomic complex traits were analyzed through polymorphic markers based on a hybridization method, such as restriction fragment length polymorphism (RFLP) markers [14], and polymerase chain reaction (PCR) techniques [15], such as random amplified polymorphic DNA (RAPD) [16], SSR [17,18], and amplified fragment length polymorphism (AFLP) markers [19].
The application of PCR led to the explosion of ESTs available in several plant databases, which have revolutionized current molecular biology and genetic approaches. In this revolutionary phase, DNA-based markers represent solid bases in plant breeding programs for their high rate of polymorphisms, numerous alleles for each locus, genome-wide distribution, and compliance to automation. However, the advances in high-throughput sequencing technologies provide new solutions for the development of high-density genetic maps, such as diversity array technology (DArT) [20], Genotyping-By-Sequencing (GBS) [21], and SNPs [22], and they were slowly replaced. These new technologies allow the identification of candidate genes within a few Kb in complex genomes. All of these sequencing applications have been a source of intense research. Here, we summarize some of the most recent methods and opportunities.
Due to the high number of SNPs in genomes compared with other DNA-based markers, high-throughput SNP discovery technology has been used more commonly for fine mapping studies. In particular, high-throughput sequencing technologies gave rise to millions of SNP markers [23], allowing the creation of DNA arrays, including those used for durum wheat such as the wheat 9K iSelect SNP array [24], the Illumina Wheat 90K iSelect SNP genotyping array [25], the Wheat 15K SNP array [26], the Axiom® Wheat 660K SNP array, the Wheat 55K SNP array, the Axiom® HD Wheat genotyping (820K) array [27], the Wheat Breeders’ 35K Axiom array [28], and the Wheat 50K Triticum TraitBreed array [29]. These arrays are efficient in the detection of polymorphisms in landraces and germplasms, increasing the availability of SNP markers [30], in particular the 820K array, derived from the exome capture technique of 43 wheat and wild species accessions, including elite cultivars, landraces, synthetic hexaploids, and wheat relatives.
Among these arrays, the high-density (90K) wheat SNP array developed by Wang et al. [25] represents the most used tool, including more than 8000 SNPs from durum cultivars [31]. The 90K SNP array was characterized by sequence information derived from 19 bread and 18 durum wheat lines, in addition to sequences from 28 [24], 24 [25], 8 [32], and 23 [33] wheat genotypes.
SNPs were widely used for the development of highly saturated genetic maps with 100,000 loci [5,34] for genetic diversity studies [35] and several SNP-based consensus wheat maps [31]. This intense application of SNP markers was mainly due to the dramatic reduction in sequencing costs, encouraging researchers to have a deep view of the target genomic regions by sequencing entire mapping populations [36]. Although the amount of SSR markers is the lowest compared to other SNP arrays, their combination has allowed analysis of the diversity in wheat, as demonstrated by Sajjad et al. [37], and fine mapping [38].
3. Linkage and Consensus Maps
Molecular markers have been widely used for the construction of linkage maps, representing the position of molecular markers along the chromosomes measured in genetic distance, based on the recombination events between individuals. The different applications of linkage maps in plant breeding are crucial for the identification of the associations of molecular markers with traits of interest for gene discovery, comparative genomics between different species, and physical mapping to facilitate genome assembly. Linkage maps have been developed for most crop species, from fruit trees to vegetables and cereals.
The first linkage map in durum wheat was developed by Blanco et al. [39]. This map was mainly constructed using restriction fragment length polymorphism (RFLP) markers. Since 2000, throughout the decade, other linkage maps, including inter-specific maps resulting from the cross between durum wheat and wild emmer, were developed. They were constructed mainly with SSR or the newly developed diversity arrays technology (DArT) markers developed by Diversity Arrays Technology Pty Ltd. (Canberra, Australia) with the microarray technology platform [20]. Through hybridization-based methods, diversity arrays detect single base pair changes (SNPs) [40]. DArTs became highly used markers as they provide extensive genome coverage, and they are obtained through a low-cost marker system. Table 1 summarizes the main durum wheat linkage maps reported in the literature.
Based on the high coverage reported utilizing the newly developed DArT markers, new consensus maps of durum wheat were developed by Marone et al. [41] and Maccaferri et al. [4]. These consensus maps integrate different linkage maps from several mapping populations containing common markers that act as anchor markers between the maps. These maps provide higher marker coverage and serve to correct incongruences between individual maps [42], thus improving the fine mapping of genes of interest. Recently, advances in next-generation sequencing (NGS) technologies have reduced the costs of DNA sequencing, making it feasible to genotype base on sequence data. These advances fostered the development of high-throughput SNPs platforms [25,43] and the development of DArTseq technology based on genotyping by sequencing (GBS) (https://www.diversityarrays.com/; accessed on 24 July 2020). With these high-throughput platforms, Maccaferri et al. [31] constructed a second-generation consensus map for durum wheat.
Three consensus maps have been developed for durum wheat in the last decade. The consensus map developed by Marone et al. [41] integrated six linkage maps, with a marker coverage of 0.15 to 0.46 markers/cM, containing a total of 1898 loci, mainly DArTs (1185), covering a total distance of 3059 cM with a mean coverage of 0.62 markers/cM. A total of 650 markers were shared by at least two individual linkage maps. Maccaferri et al. [4] also developed a consensus map integrating six linkage maps as a framework, including 598 markers, mainly SSRs (295) and DArTs (281), and 1977 markers were interpolated from eight other mapping populations. The consensus map contained a total of 2575 markers covering 2463 cM, which represents a coverage of 1.04 markers/cM and is a significant improvement from the original maps, which ranged from 0.08 to 0.48 markers/cM. More recently, with the development of SNP arrays, Maccaferri et al. [31] constructed a new consensus map integrating this technology. A total of 13 biparental mapping populations were used, and the final map included 30,144 markers, mainly SNPs, from the array developed by Wang et al. [25] spanning 2631 cM. The marker coverage increased from 4.83 markers/cM in the most saturated linkage map (Svevo × Zavitan) to 11.45 markers/cM in the consensus map.
4. Traits and QTL Analysis
4.1. Quality
Durum grains, a storehouse of nutritional elements for the human diet, contain starch (70.2%), proteins (12.2%), lipids (1.9%), fiber (1.6%), and minerals (1.6%), with varying water content [79]. In addition, the kernels comprise high carbohydrate and antioxidant (such as carotenoid pigments) contents, together with high vitamin, potassium, calcium, sodium, and magnesium levels.
The kernel quality is of paramount importance in the end-product quality of commercial wheat varieties, determining the type of products that can be produced. Thus, it is possible to distinguish a commercial value in the beginning of wheat production and a technological value linked to the worldwide market requirements for end-product uses.
For the commercial value, the wheat quality is primarily evaluated through the milling rate, which consists of the quantity of wheat flour derived from 100 kg of seeds. This rate aims to obtain the maximum quantity of flour, analyzing the seed size, thousand-kernel weight, ash mass (or mineral content), and the percentage of seed defects, such as pre-germination, small or white seeds, and pathologic darkness of the grain. For the technological value, wheat quality is evaluated for the strength of the flour protein when mixed with water to make dough (e.g., hardness, quantity and quality of the proteins, and rheological parameters), the quantity of water required for workable dough, and the flour color (e.g., yellow color). The protein quantity and quality influence the characteristics of the end wheat products: for example, a high protein content brings major water absorbance, increasing the productivity rate and the shelf life of the final products. Several investigations [54,80,81,82,83,84,85] indicated that factors influencing the protein concentration in cultivated and wild wheat include quantitative trait loci (QTL) located almost on all chromosomes.
Carotenoid pigments have an enormous importance for the nutritional value for human health and affect the wheat flour color. The antioxidant activity of carotenoids, together with protein, increases the nutritional and technological characteristics of flour [86,87]. An example of flour with a high carotenoid content is whole wheat flour, which also contains high levels of fiber (β-glucan and arabinoxylan), vitamins, and antioxidant molecules such as tocopherols and flavonoids, which are also considered to be kernel components that are important for quality.
Another important trait related to wheat quality is the starch content (75% of the weight of the mature grain), which consists of two types: amylose and amylopectin. In fact, the water absorption of the dough is connected to the starch content [88,89]. High starch content changes the functional properties of the flour, such as the gelatinization, pasta characteristics, and baking applications [90]. This aspect is important in the production of baked foods, such as cakes and some types of breads, providing a tender status to the final products [79].
4.2. Biotic Stress
Major breeding programs are also focused on improving biotic stress resistance because of the severe damage to production worldwide. These kinds of stresses are generated by living organisms (fungi, bacteria, insects, etc.) causing diseases such as rust, powdery mildew, Fusarium graminearum, and various viruses.
Amongst the pathogens damaging durum wheat crops at the leaf and stem levels, rust pathogens are the most prevalent. This includes leaf rust (Puccinia triticina), stem rust (P. graminis tritici), and stripe rust (P. striiformis), causing losses of 15–20% worldwide [91]. To date, over 77 genes conferring resistance to leaf rust (Lr) have been characterized and localized to specific wheat chromosomes [92]. Durum wheat has been historically more resistant to leaf rust compared with bread wheat; however, this kind of resistance could evolve into a rapid breakdown due to the birth of new virulent races.
Powdery mildew (Blumeria graminis f. sp. tritici) is recognized as a disease of wheat of high economic importance, especially in warm climate areas where the productivity is high. Therefore, one of the most important purposes of breeding programs is to make resistant durum wheat. To achieve this goal, several studies, including a careful analysis of the environmental conditions that can influence the host–pathogen interactions, a study of the genetic and molecular interactions between the host and pathogen, and the search for new sources of resistance to be transferred in varieties, are required [93].
Fusarium head blight (FHB) is one of the most common and harmful diseases of durum wheat in the world. The incidence and severity of FHB depends on the weather, the areas, and the varieties used. This pathogen has caused consequential production losses together with damage to the quality of grain and the presence of mycotoxins. The presence of Fusarium toxins (deoxynivalenol, DON) in wheat represents a serious hygienic–sanitary problem [30,94].
4.3. Abiotic Stress
In the Mediterranean Basin, durum wheat is cultivated under variable environmental conditions. In rainfed agricultural environments, drought stress critically constrains the crop yield. It is particularly challenging for breeders under the current unpredictable climate change to stretch the adaptability and performance stability of their cultivars. In the Mediterranean, the environment is responsible for as much as 98% of bread and durum wheat yield variations [95].
Wheat yield is dependent on the grain number per unit area and grain weight. With climate change, yield reduction is led by a significant decrease in one or both yield components. The grain number, in turn, may be split into spikes per unit area and grains per spike. The yield components are sequentially determined and are counter-dependent. Reductions in the grain number per unit land area due to an increase in temperature have been widely reported, as has a reduction in the grain weight, which depends on the environmental conditions before flowering and during grain filling [96,97]. Although drought stress in the pre-flowering period can influence the grain weight [98], the grain filling period is considered critical for the final grain weight [99]. In Mediterranean environments, water becomes mostly limiting after anthesis, typically being accompanied by high temperatures, which causes a reduction in the yield potential of approximately 50% [100]. The main impact of drought stress after anthesis is to reduce the grain setting, size, and weight [101].
Breeding for adaptation to drought is extremely challenging due to the complexity of the target environments as well as the stress-adaptive mechanisms adopted by plants to withstand and mitigate the negative effects of water deficit [102]. The incomplete knowledge of the physiological and genetic basis of drought resistance [103] as well as insufficient consideration of drought environments when defining target traits for stress resistance [104] may explain the low yield improvements observed in wheat grown in dry regions [105].
Understanding the physiological mechanisms associated with drought resistance and the genetics underlying them may provide new strategies for engineering varieties resilient to drought, which is essential for wheat breeding [106,107]. The crop traits to be considered as selection targets under drought conditions must be genetically correlated with the yield and should have a greater heritability than the yield itself [99,108]. Among them, early vigor, leaf area duration, crop water status, radiation use efficiency, and root architecture have been identified as being associated with the yield under rainfed conditions (reviewed in [109]).
Another important constraint in crop production is heat stress. This is tightly associated with drought in the Mediterranean region as it is of particular importance when it occurs between heading and maturity, affecting the flowering and grain filling, thus reducing the grain yield [110]. Heat stress in plants produces a wide range of effects, affecting their physiology and altering their gene regulation, causing a decrease in the synthesis of proteins [111]. The ability of plants to cope with high temperatures is called thermotolerance and can be measured through the increase in the fluidity of the cellular membrane, causing the escape of electrolytes to the extracellular medium [112].
Soil salinity is considered an important limiting factor for crop production in arid and semi-arid regions. Although related to drought stress, salinity affects plant growth during the whole cycle of plant development [113]. The last factor to be considered as abiotic stress is cold and/or frost tolerance, although growing winter wheat in regions such as Central Europe or North America substantially increases the yield potential [114].
4.4. QTL Mapping
Classically, QTL mapping has been performed predominantly in populations derived from two parents (biparental) with different phenotypic performances. The success in detecting QTL depends on the marker density, population size, and the heritability of the trait. With durum wheat, a large number of QTL studies have been performed to investigate yield performance, biotic and abiotic stresses, phenology, and quality.
The grain yield and phenology are traits much studied in wheat, as reviewed in Soriano et al. [115]; thus, the present review focuses on the traits involved in abiotic and biotic stress resistance and grain quality. To synthesize all the QTL in the same linkage map to obtain an overview of the QTL distribution, the durum wheat consensus map developed by Maccaferri et al. [31] was used for QTL projection based on the homothetic approach described by Chardon et al. [116]. A total of 45 QTL studies—six corresponding to abiotic stress tolerance, 24 to biotic stress resistance, and 15 to quality traits—were revised (Table 2 and Table S1), comprising 368 QTL, from which 127 corresponded to traits related to abiotic stress, 71 to biotic stresses, and 171 to quality-related traits (Figure 1).
4.5. Genome-Wide Association Studies
Association mapping or genome-wide association study (GWAS) is a complementary approach to dissect the genetic basis of complex traits, providing broader allelic coverage and offering higher mapping resolution. This is based on linkage disequilibrium (LD), defined as the non-random association of alleles at different loci, and is used to detect the relationship between phenotypic variation and genetic polymorphisms [117]. It is important, however, to differentiate the LD due to physical linkages from LD due to the population structure, which can be caused by selection, genetic drift, and species-dependent characteristics, such as the mating system. Germplasm collections characterized by medium to high LD levels are suitable for the identification of chromosome regions harboring genes/QTL controlling agronomic traits in wheat [118]. The main differences between biparental QTL mapping and GWAS are shown in Table 3. Many studies have been conducted in durum wheat to investigate the genetic basis of yield and yield components [115,119,120], crop phenology [115], or biomass [115]. Here, we summarize a total of 19 GWAS studies that identified marker–trait associations (MTAs) involved in abiotic (4) and biotic (8) stress and grain quality (7) (Table 2).
A summary of the total number of QTL and MTAs reported in durum wheat with the analyzed traits is shown in Table 4. Our review included ten traits related to abiotic stress, nine related to biotic stress, and nineteen related to quality traits.
4.6. QTL Meta-Analysis
As reported in Table 2, hundreds of QTL have been reported for the different traits considered. One way to synthesize this information is the QTL meta-analysis developed by Goffinet and Gerber [154]. The aim of QTL meta-analysis is the identification of genome regions repeatedly involved in trait variation to narrow down the QTL supporting intervals and make them useful for breeding and enabling the identification of candidate genes. The process involves several steps: the construction of a consensus map integrating different types of markers or the use of a reference map for the species; the projection of the initial QTL on the consensus maps; or the estimation of consensus regions harboring different QTL or meta-QTL.
To select the most appropriate meta-QTL for breeding purposes or candidate gene isolation, Löffler et al. [155] established three criteria that the meta-QTL must meet: (1) a small supporting interval, (2) integrating a high number of original QTL, and (3) a high effect of the phenotypic variance explained by the original QTL.
QTL meta-analysis has been performed in both bread wheat and durum wheat for different traits such as grain yield [112], crop phenology [112,156], disease resistance [155,157,158,159], plant height [160], grain-related traits [161,162], root-related traits [118,163,164], and sprouting tolerance and dormancy [165].
The use of consensus maps integrating the most common molecular markers and the identification of consensus QTL regions among different mapping populations from different parental sources will help breeders to select the most appropriate plant material for the development of new cultivars. Redefining (and shortening) QTL regions by QTL meta-analysis will help with the identification of candidate genes for accelerating the breeding process. The use of the recent release genome sequence of durum wheat [12] will allow the identification of the physical regions of QTL of interest to abord map-based cloning strategies.
5. Innovative Experimental Designs for Enhanced Gene Discovery
5.1. Bulked Segregant Analysis by Sequencing (BSAseq)
The analysis of transcriptome through SNP arrays allowed the combination of bulked segregant analysis (BSA) and RNA sequencing to fine-map small proportions of a wheat genome involved in the traits of interest [61]. This strategy has been successfully used for identifying candidate genes and gene cloning purposes in maize [166] and bread wheat [167,168,169]. In durum wheat, BSAseq was used to refine the species cytoplasm-specific (scs) locus [170]. The limitation of BSAseq in tetraploid wheat could be due to the presence of co-expressed homoeologous genes and the absence of a good reference sequence for mapping, which was completed last year [12].
5.2. Development of New Populations
As described above, the main limits of biparental QTL analysis rely on the low number of recombinant events, the limited number of polymorphic markers, and the reduced availability of genetic diversity. To overcome these drawbacks, in recent years, new breeding programs and new plant populations have been proposed, increasing the statistical power and the association between markers and traits and allowing the identification of key genes involved in the phenotypic variation [171]. These include the multiparent advanced generation intercross (MAGIC) and the nested association mapping (NAM) populations, both based on multiple founders.
In particular, the MAGIC populations are generated through inter-crossing several parental lines (eight or more) through two-way, four-way, and eight-way crosses and subsequent self-fertilization to generate recombinant inbred lines (RILs). Compared to traditional biparental populations, the use of more founders in the initial crosses allowed an increase in the recombination, consequently improving the mapping resolution and allowing greater allelic diversity. The MAGIC design was successfully realized in several crops for fine genetic mapping; however, a slow uptake has been observed in durum wheat, where a four-parent durum wheat population was produced [172]. A successful four-way durum wheat MAGIC population was developed by Milner et al. [173] for the first time by crossing four elite cultivars from different origins. The population was used to study important quantitative traits such as heading and maturity date, plant height, and grain yield. The results reported based on the analysis based on founder haplotype probabilities proved to be more efficient in QTL mapping in this multiparent population as compared to a conventional bi-allelic assay. In the present review, we reported studies on 40 traits that can help breeders and researchers in the choice of the most useful genotypes carrying major QTL for the production of new MAGIC populations in durum wheat.
The NAM populations were developed through crossing several founders with the same reference line to generate a series of “interconnected” segregating inbred families. The advantage of this population is represented by the possibility of incorporating a high number of alleles in one species gene pool, accurate QTL effects and positions, and overcoming the limits of linkage analysis and association mapping approaches [174,175].
The NAM design for QTL mapping was developed in maize [176,177] and later applied to other crops including soybean [178], sorghum [179], barley [180], bread wheat [181], and durum wheat [182,183,184].
Germplasm resources as landrace collections could be of interest for the development of new mapping populations following the approaches of MAGIC and NAM to incorporate allelic diversity. As reported by Soriano et al. [185], Mediterranean landraces represent an important group of genetic resources due to their good adaptation, huge genetic diversity, resilience to abiotic and abiotic stresses, and their differences in yield formation strategies [186].
5.3. Candidate Gene: A New Approach for Studying Quantitative Trait Loci
One of the main objectives of molecular genetics is to identify and isolate genes controlling important traits. Three main approaches lead to the cloning of genes of interest: positional cloning [187], insertional mutagenesis [188], and candidate genes (CGs) [189,190]. The last strategy has been successfully used after the assumptions regarding the biological function of the gene of interest. It started from sequenced genes of known function that could correspond to major loci (Mendelian trait loci, MTLs, or quantitative trait loci, QTL). CGs may be structural genes or genes involved in the regulation of a metabolic pathway [189]. The hypothesis is that a molecular gene polymorphism is related to the phenotypic variation.
The CG approach is based on three chronological steps: (1) choice of CGs, established through molecular and functional studies or based on linkage data; (2) molecular polymorphism in the CG to localize it on a genetic map or to calculate statistical correlations between CG polymorphisms and phenotypic variation in a set of unrelated individuals; and (3) complementary experiments (if map co-segregation and/or statistical correlation were found) to confirm the real association of the CG in the trait variation [191]. The CG approach is more adapted to QTL characterization than positional cloning or insertional mutagenesis due to the multiplicity of genes involved in the trait, their partial effect, and indefinite genetic map positions.
The CG approach has been used with success in durum wheat for many traits such as disease resistance [34,192], carotenoid content (as reported in [87]), protein content [85], fiber (as reported in [79,193]), and salinity stress [194].
Thanks to the recent release of durum wheat genome [12] and gene expression analysis databases as the wheat expression browser [195] (http://www.wheat-expression.com/; accessed on 25 January 2021), new advancements in CG identifications can be obtained through the application of new computational methods developed to prioritize CGs in QTL before functional studies. The identification of the gene TdHMA3-B1 responsible for variation in cadmium accumulation in the grain is a successful example of the use of the genome sequence data together with the genetic variation reported in germplasm collections as the global durum panel, as reported in [12].
Moyers [196] developed a computational approach, “Camoco”, that integrates loci identified by GWAS with functional information derived from gene co-expression networks. More recently, Lin et al. [197] developed an algorithm, “QTG-Finder2”, for prioritizing QTL causal genes in plants and validated Sorghum bicolor and Setaria viridis models.
6. Gene Editing in Durum Wheat
Genome editing is a new biotechnology allowing specific manipulation of a target genomic sequence, which has the potential to improve wheat performance and supersede traditional methods in plant breeding.
This technology in wheat is made more problematic due to the complexity of the wheat genome and the difficulty in genetic transformation. Despite this, recent progress in plant genome editing has been reported, above all, for bread wheat. One of the first papers using the CRISPR/Cas9 system in wheat was reported in 2014 by [198], who reported a mutation of three homoeoalleles TaMLO in hexaploid wheat for a gene that confers heritable resistance to powdery mildew.
The scientific community have concentrated their efforts on resistance to pathogens such as powdery mildew [198]; leaf rust; the analysis of a pathogenesis-related protein 1 Gene, TcLr19PR1, involved in resistance against leaf rust as reported by [199]; stripe rust with the resistance gene Yr10, which encodes an evolutionarily conserved and unique CC-NBS-LRR sequence in wheat [200]; sharp eyespot disease [201]; and resistance to Bipolaris sorokiniana with an R2R3 MYB transcription factor in wheat, TaPIMP1 [202].
In addition, yield was the second most studied trait; the genes TaGASR7, TaGW2, and TaLOX2 in hexaploid wheat and only TdGASR7 in tetraploid durum wheat were edited because they were found to be associated with grain yield components [203,204,205]. However, the research in this field is rapidly increasing. Recently, two independent studies [206,207] reported CRISPR/Cas9 editing of the α-Amylase/Trypsin inhibitor (ATI) genes, reducing the allergen proteins in durum wheat. The ATI subunits WTAI-CM3 and WTAI-CM16 from the durum wheat cultivar Svevo were edited to produce lines with reduced allergens. The authors obtained durum wheat lines with stable and heritable mutations in these genes. These lines will be of special interest for durum wheat breeding programs for the introgression of the alleles in other elite cultivars.
The recent advance in high-quality reference genomes for durum wheat along with new efficient genetic transformation strategies will accelerate the application of genome editing technology.
7. From Classical Breeding to Genomic Selection
The selection methods used in wheat breeding programs have deeply evolved with time. Classical breeding based on the phenotypic selection of a trait was the main approach used by breeders to increase crop productivity during the 20th century [208]. This technique implies the selection of varieties carrying the desired characteristics for the target trait, usually morphological or visual features such as the yield, yield components, or disease resistance. Despite the large improvements in genetic gains for yield and quality obtained in bread [209,210] and durum wheat [3,211], this process takes several years to finally obtain a commercial variety and has some limitations, especially when target traits are highly dependent on the environment due to low heritability [212].
The development of molecular biology allowed the use of markers based on the sequence or polymorphisms in DNA for the identification of traits enhancing agronomic performance in the earlier stages of development. Marker-assisted selection (MAS) resulted in an important advantage integrated with traditional breeding methods to enhance the efficiency of cultivar development. However, MAS depends on the genetic linkage of traits with markers, and typically, only genetic loci with major effects are exploitable in this way [213]. Since many agronomic traits present a multigenic quantitative nature and the effect of the environment on them needs to be assessed, MAS cannot replace traditional breeding methods for these traits, particularly in later-generation screening and cultivar evaluation.
The availability of high-density, low-cost marker genotyping platforms has enabled a change in plant breeding by making genomic prediction and selection feasible. Genomic selection (GS) refers to the selection of genotypes using genomic information on a genome-wide scale to make selections [214]. Genomic selection uses genome-wide markers to estimate the effects of all genes or chromosome positions simultaneously to predict the breeding values of progeny, which are used for the selection of individuals without costly phenotyping, saving money and time and increasing the accuracy of selection [214].
Different studies in durum wheat applied this technology for predicting the grain yield, quality traits, and disease resistance. Fiedler et al. [215] found prediction accuracies from 0.27 to 0.66 for different grain and semolina quality traits using 1184 breeding lines from the North Dakota State University (NDSU) durum wheat breeding program. Haile et al. [216] studied the potential of single- and multi-trait genomic prediction models on grain yield and quality traits using a breeding panel comprising 170 cultivars and advanced lines as well as a doubled haploid population.
The accuracy found by these authors ranged from 0.5 to 0.8 for single traits, increasing in multi-traits for grain yield. Zaim et al. [217] used four populations to develop genomic prediction models for grain yield under drought conditions. These authors found that including QTL information from the populations increased the prediction accuracies by 0.06 to 0.12 points. In addition to grain yield and quality traits, disease resistance is another target for GS. In this sense, Moreno-Amores et al. [218] assessed the prediction ability for fusarium head blight (FHB) using plant height and heading date as covariates as they are influenced negatively by FHB.
8. Speed Breeding
The development of speed breeding protocols can be considered as a useful approach to help in the development of new mapping populations and to advance the first generations in the breeding programs, which, being assisted by molecular markers, will save time for breeders, reducing the length of the breeding cycles and selecting the best genotypes. The technique implies the use of extended photoperiods in a controlled environment. For durum wheat, up to six generations in a year have been achieved [219]. These protocols have been adapted for multi-trait phenotyping in durum wheat, as described by Alahmad et al. [220] for the rapid selection of generations and the characterization of breeding lines anytime during the year.
9. Future Prospects
To meet the food needs for the future, farmers must increase crop yields considering the climate and environment changes. Information gained from sequenced genomes in related species and in durum wheat, together with studies of fine mapping and QTL cloning, allows the identification of a high number of molecular markers, key genes, quantitative trait loci, and networks, which will lead to higher yielding crops.
The decreasing cost of NGS technologies and the huge availability of sequence data on web databases allow for drastic reductions in the time required to identify candidate genes. Thus, projects that previously required more than 10 years for fine mapping, QTL cloning, and candidate gene identification could now be completed in 1–2 years for more simple traits.
Supplementary Materials
Supplementary materials can be found at https://0-www-mdpi-com.brum.beds.ac.uk/2223-7747/10/2/315/s1. Table S1: QTL database.
Author Contributions
Conceptualization, A.G and J.M.S.; methodology, P.C., I.M., A.G. and J.M.S.; formal analysis, P.C., I.M., A.G. and J.M.S.; writing—original draft preparation, P.C., I.M., A.G. and J.M.S.; writing—review and editing, P.C., I.M., A.G. and J.M.S.; visualization, P.C., I.M., A.G. and J.M.S.; funding acquisition, A.G. and J.M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the projects PRIMA 2019 “CEREALMED” (Italy), PON-AIM project AIM1812334 (Ministero dell’Istruzione, dell’Università e della Ricerca, Italy) and PID2019-109089RB-C31 (Ministerio de Ciencia e Innovación, Spain).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data present in this study is available within the manuscript and its supplementary information.
Acknowledgments
J.M.S. acknowledges the contribution of the CERCA Program (Generalitat de Catalunya).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 78–202. [Google Scholar] [CrossRef]
- Royo, C.; Álvaro, F.; Martos, V.; Ramdani, A.; Isidro, J.; Villegas, D.; García del Moral, L.F. Genetic changes in durum wheat yield components and associated traits in Italian and Spanish varieties during the 20th century. Euphytica 2007, 155, 259–270. [Google Scholar] [CrossRef]
- Subira, J.; Peña, R.J.; Álvaro, F.; Ammar, K.; Ramdani, A.; Royo, C. Breeding progress in the pasta-making quality of durum wheat cultivars released in Italy and Spain during the 20th Century. Crop Pasture Sci. 2014, 65, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Maccaferri, M.; Cané, M.A.; Sanguineti, M.C.; Salvi, S.; Colalongo, M.C.; Massi, A.; Clarke, F.; Knox, R.; Pozniak, C.J.; Clarke, J.M.; et al. A consensus framework map of durum wheat (Triticum durum Desf.) suitable for linkage disequilibrium analysis and genome-wide association mapping. BMC Genom. 2014, 15, 873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colasuonno, P.; Gadaleta, A.; Giancaspro, A.; Nigro, D.; Give, S.; Incerti, O.; Mangini, G.; Signorile, A.; Simeone, R.; Blanco, A. Development of a high-density SNP-based linkage map and detection of yellow pigment content QTLs in durum wheat. Mol. Breed. 2014, 34, 1563–1578. [Google Scholar] [CrossRef]
- Kumar, S.; Goyal, A.; Mohan, A.; Balyan, H.S.; Gupta, P.K. An integrated physical map of simple sequence repeats in bread wheat. Aust. J. Crop Sci. 2013, 7, 460–468. [Google Scholar]
- Blanco, A.; Mangini, G.; Giancaspro, A.; Giove, S.; Colasuonno, P.; Simeone, R.; Signorile, A.; de Vita, P.; Mastrangelo, A.M.; Cattivelli, L.; et al. Relationships between grain protein content and grain yield components through quantitative trait locus analyses in a recombinant inbred line population derived from two elite durum wheat cultivars. Mol. Breed. 2012, 30, 79–92. [Google Scholar] [CrossRef]
- Maccaferri, M.; Mantovani, P.; Tuberosa, R.; DeAmbrogio, E.; Giuliani, S.; Demontis, A.; Massi, A.; Sanguineti, M.C. A major QTL for durable leaf rust resistance widely exploited in durum wheat breeding programs maps on the distal region of chromosome arm 7BL. Theor. Appl. Genet. 2008, 117, 1225–1240. [Google Scholar] [CrossRef]
- Blanco, A.; Gadaleta, A.; Cenci, A.; Carluccio, A.V.; Abdelbacki, A.M.M.; Simeone, R. Molecular mapping of the novel powdery mildew resistance gene Pm36 introgressed from Triticum turgidum var dicoccoides in durum wheat. Theor. Appl. Genet. 2008, 117, 135–142. [Google Scholar] [CrossRef]
- Blanco, A.; Colasuonno, P.; Gadaleta, A.; Mangini, G.; Schiavulli, A.; Simeone, R.; Digesù, A.M.; de Vita, P.; Mastrangelo, A.M.; Cattivelli, L. Quantitative trait loci for yellow pigment concentration and individual carotenoid compounds in durum wheat. J. Cereal Sci. 2011, 54, 255–264. [Google Scholar] [CrossRef]
- Gadaleta, A.; Giancaspro, A.; Give, S.L.; Zacheo, S.; Mangini, G.; Simeone, R.; Signorile, A.; Blanco, A. Genetic and physical mapping of new EST-derived SSRs on the A and B genome chromosomes of wheat. Theor. Appl. Genet. 2009, 118, 1015–1025. [Google Scholar] [CrossRef]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome reveals past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Nam, J.H.; Song, H.S.; Suh, S.J.; Heo, H.Y.; Park, M.W.; Baek, S.B.; Park, H.H. Achievement and prospect of good quality breeding in winter cereals. Korean J. Crop Sci. 1998, 43, 11–15. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Mullis, K.B.; Faloona, F. Specific synthesis of DNA in vitro via a polymerase catalysed chain reaction. Meth. Enzymol. 1987, 155, 335–350. [Google Scholar]
- Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [Green Version]
- Tautz, D.; Trick, M.; Dover, G. Cryptic simplicity in DNA is a major source of genetic variation. Nature 1986, 322, 652–656. [Google Scholar] [CrossRef]
- Tautz, D. Hypervariability of simple sequence as a general source for polymorphic DNA markers. Nucleic Acids Res. 1989, 17, 6463–6471. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity arrays: A solid state technology for sequence information independent genotyping. Nucleic Acid Res. 2001, 29, E25. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wang, Y.; Chen, M.; Edae, E.; Poland, J.; Akhunov, E.; Chao, S.; Bai, G.; Carver, B.F.; Yan, L. Precisely mapping a major gene conferring resistance to Hessian fly in bread wheat using genotyping-by-sequencing. BMC Genom. 2015, 16, 108. [Google Scholar] [CrossRef] [Green Version]
- Cabral, A.L.; Jordan, M.C.; McCartney, C.A.; You, F.M.; Humphreys, D.G.; MacLachlan, R.; Pozniac, C.J. Identification of candidate genes.; regions and markers for pre-harvest sprouting resistance in wheat (Triticum aestivum L.). BMC Plant Biol. 2014, 14, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Priya, P.; Misra, G.; Yadav, G. Structural and biochemical perspectives in plant isoprenoid biosynthesis. Phytochem. Rev. 2013, 12, 255–291. [Google Scholar] [CrossRef]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using the high-density 90,000 SNP array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Boeven, P.H.; Longin, C.F.; Leiser, W.L.; Kollers, S.; Ebmeyer, E.; Würschum, T. Genetic architecture of male floral traits required for hybrid wheat breeding. Theor. Appl. Genet. 2016, 129, 2343–2357. [Google Scholar] [CrossRef]
- Winfield, M.O.; Allen, A.M.; Burridge, A.J.; Barker, G.L.A.; Benbow, H.R.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; Scopes, G. High-density SNP genotyping array for hexaploid wheat and its secondary and tertiary gene pool. Plant Biotechnol. J. 2016, 14, 1195–1206. [Google Scholar] [CrossRef]
- Allen, A.M.; Winfield, M.O.; Burridge, A.J.; Downie, R.C.; Benbow, H.R.; Barker, G.L.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A. Characterization of a Wheat Breeders Array suitable for high-throughput SNP genotyping of global accessions of hexaploid bread wheat (Triticum aestivum). Plant Biotechnol. J. 2017, 15, 390–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasheed, A.; Xia, X. From markers to genome-based breeding in wheat. Theor. Appl. Genet. 2019, 132, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Dong, Z.; Zhao, L.; Ren, Y.; Zhang, N.; Chen, F. The Wheat 660K SNP array demonstrates great potential for marker-assisted selection in polyploid wheat. Plant Biotechnol. J. 2020, 18, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Maccaferri, M.; Ricci, A.; Salvi, S.; Milner, S.G.; Noli, E.; Martelli, P.L.; Casadio, R.; Akhunov, E.; Scalabrin, S.; Vendramin, V.; et al. A high-density SNP-based consensus map of tetraploid wheat as a bridge to integrate durum and bread wheat genomics and breeding. Plant Biotechnol. J. 2015, 13, 5. [Google Scholar] [CrossRef]
- Pont, C.; Murat, F.; Guizard, S.; Flores, R.; Foucrier, S.; Bidet, Y.; Quraishi, U.M.; Alaux, M.; Doležel, J.; Fahima, T.; et al. Wheat syntenome unveils new evidences of contrasted evolutionary plasticity between paleo- and neoduplicated subgenomes. Plant J. 2013, 76, 1030–1044. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.M.; Barker, G.L.A.; Berry, S.T.; Coghill, J.A.; Gwilliam, R.; Kirby, S.; Robinson, P.; Brenchley, R.C.; D’Amore, R.; McKenzie, N.; et al. Transcript-specific.; single-nucleotide polymorphism discovery and linkage analysis in hexaploid bread wheat (Triticum aestivum L.). Plant Biotechnol. J. 2011, 9, 1086–1099. [Google Scholar] [CrossRef] [Green Version]
- Giancaspro, A.; Give, S.L.; Zito, D.; Blanco, A.; Gadaleta, A. Mapping QTL for Fusarium head blight resistance in an interspecific wheat population. Front. Plant Sci. 2016, 7, 1381. [Google Scholar] [CrossRef]
- Marcotuli, I.; Houston, K.; Schwerdt, J.G.; Waugh, R.; Fincher, G.B.; Burton, R.A.; Blanco, A.; Gadaleta, A. Genetic diversity and genome wide association study of β-glucan content in tetraploid wheat grains. PLoS ONE 2016, 11, e0152590. [Google Scholar] [CrossRef] [Green Version]
- Jaganathan, D.; Bohra, A.; Thudi, M.; Varshney, R.K. Fine mapping and gene cloning in the post-NGS era: Advances and prospects. Theor. Appl. Genet. 2020, 133, 1791–1810. [Google Scholar] [CrossRef] [Green Version]
- Sajjad, M.; Khan, S.H.; Shahzad, M. Patterns of allelic diversity in spring wheat populations by SSR-markers. Cytol. Genet. 2018, 52, 155–160. [Google Scholar] [CrossRef]
- Gill, H.S.; Li, C.; Sidhu, J.S.; Liu, W.; Wilson, D.; Bai, G.; Gill, B.S.; Sehgal, S.K. Fine Mapping of the Wheat Leaf Rust Resistance Gene Lr42. Int. J. Mol. Sci. 2019, 20, 2445. [Google Scholar] [CrossRef] [Green Version]
- Blanco, A.; De Giovanni, C.; Bellomo, M.P.; Cenci, A.; D’Ovidio, R.; Iacono, E.; Laddomada, B.; Pagnotta, M.A.; Porcedu, E.; Sciancalepore, A.; et al. A genetic linkage map of durum wheat. Theor. Appl. Genet. 1998, 97, 721–728. [Google Scholar] [CrossRef]
- Wenzl, P.; Carling, J.; Kudrna, D.; Jaccoud, D.; Huttner, E.; Kleinhofs, A.; Kilian, A. Diversity Arrays Technology (DArT) for whole-genome profiling of barley. Proc. Natl. Acad. Sci. USA 2004, 101, 9915–9920. [Google Scholar] [CrossRef] [Green Version]
- Marone, D.; Laidò, G.; Gadaleta, A.; Colasuonno, P.; Ficco, D.B.; Giancaspro, A.; Giove, S.; Panio, G.; Russo, M.A.; De Vita, P.; et al. A high-density consensus map of A and B wheat genomes. Theor. Appl. Genet. 2012, 125, 1619–1638. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Han, Y.; Zhao, Y.F.; Troggio, M.; Korban, S.S. A multi-population consensus genetic map reveals inconsistent marker order among maps likely attributed to structural variations in the apple genome. PLoS ONE 2012, 7, e47864. [Google Scholar] [CrossRef] [Green Version]
- Korzun, V.; Röder, M.S.; Wendekake, K.; Pasqualone, A.; Lotti, C.; Ganal, M.W.; Blanco, A. Integration of dinucleotide microsatellites from hexaploid bread wheat into a genetic linkage map of durum wheat. Theor. Appl. Genet. 1999, 98, 1202–1207. [Google Scholar] [CrossRef]
- Lotti, C.; Salvi, S.; Pasqualone, A.; Tuberosa, R.; Blanco, A. Integration of AFLP markers into an RFLP-based map of durum wheat. Plant Breed. 2000, 119, 393–401. [Google Scholar] [CrossRef]
- Nachit, M.; Elouafi, I.; Pagnotta, M.A.; El Saleh, A.; Iacono, E.; Labhilili, M.; Asbati, A.; Azrak, M.; Hazzam, H.; Benscher, D.; et al. Molecular linkage map for an intraspecific recombinant inbred population of durum wheat (Triticum turgidum L. var durum). Theor. Appl. Genet. 2001, 102, 177–186. [Google Scholar] [CrossRef]
- Blanco, A.; Simeone, R.; Cenci, A.; Gadaleta, A.; Tanzarella, O.A.; Porceddu, E.; Salvi, S.; Tuberosa, R.; Figliuolo, G.; Spagnoletti, P.; et al. Extension of the Messapia X dicoccoides linkage map of Triticum turgidum (L.) Thell. Cell. Mol. Biol. Lett. 2004, 9, 529–541. [Google Scholar]
- Elouafi, I.; Nachit, M.M. A genetic linkage map of the Durum x Triticum dicoccoides backcross population based on SSRs and AFLP markers.; and QTL analysis for milling traits. Theor. Appl. Genet. 2004, 108, 401–413. [Google Scholar] [CrossRef]
- Somers, D.J.; Fedak, G.; Clarke, J.; Cao, W. Mapping of FHB resistance QTLs in tetraploid wheat. Genome 2006, 49, 1586–1593. [Google Scholar] [CrossRef]
- Pozniak, C.J.; Knox, R.E.; Clarke, F.R.; Clarke, J.M. Identification of QTL and association of a phytoene synthase gene with endosperm colour in durum wheat. Theor. Appl. Genet. 2007, 114, 525–537. [Google Scholar] [CrossRef]
- Mantovani, P.; Maccaferri, M.; Sanguineti, M.C.; Tuberosa, R.; Catizone, I.; Wenzl, P.; Thompson, B.; Carling, J.; Huttner, E.; DeAmbrogio, E.; et al. An integrated DArT-SSR linkage map of durum wheat. Mol. Breed. 2008, 22, 629–648. [Google Scholar] [CrossRef]
- Peleg, Z.; Saranga, Y.; Suprunova, T.; Ronin, Y.; Röder, M.; Kilian, A.; Korol, A.B.; Fahima, T. High-density genetic map of durum wheat × wild emmer wheat based on SSR and DArT markers. Theor. Appl. Genet. 2008, 117, 103–115. [Google Scholar] [CrossRef]
- Zhang, W.; Dubcovsky, J. Association between allelic variation at the Phytoene synthase 1 gene and yellow pigment content in the wheat grain. Theor. Appl. Genet. 2008, 116, 635–645. [Google Scholar] [CrossRef] [Green Version]
- Marone, D.; Del Olmo, A.I.; Laido, G.; Sillero, J.C.; Emeran, A.A.; Russo, M.A.; Ferragonio, P.; Giovaniello, V.; Mazzucotelli, E.; De Leonardis, A.M. Genetic analysis of durable resistance against leaf rust in durum wheat. Mol. Breed. 2009, 24, 25–39. [Google Scholar] [CrossRef]
- Suprayogi, Y.; Pozniak, C.J.; Clarke, F.R.; Clarke, J.M.; Knox, R.E.; Singh, A.K. Identification and validation of quantitative trait loci for grain protein concentration in adapted Canadian durum wheat populations. Theor. Appl. Genet. 2009, 119, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.A.; Ficco, D.B.M.; Marone, D.; De Vita, P.; Vallega, V.; Rubies-Autonell, C.; Rati, C.; Ferragonio, P.; Giovaniello, V.; Pecchioni, N. A major QTL for resistance to soil-borne cereal mosaic virus derived from an old Italian durum wheat cultivar. J. Plant Interact. 2012, 7, 290–300. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Huber, K.; Heckmann, J.; Steiner, B.; Nelson, J.; Buerstmayr, H. Mapping of QTL for Fusarium head blight resistance and morphological and developmental traits in three backcross populations derived from Triticum dicoccum × Triticum durum. Theor. Appl. Genet. 2012, 125, 1751–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.; Comeau, A.; Langevin, F.; Hucl, P.; Clarke, J.; Brule-Babel, A.; Pozniak, C. Identification of novel QTL for resistance to Fusarium head blight in a tetraploid wheat population. Genome 2012, 55, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.; Tamhankar, S.; Oak, M.; Raut, A.; Honrao, B.; Rao, V.S.; Misra, S.C. Mapping of QTL for agronomic traits and kernel characters in durum wheat (Triticum durum Desf). Euphytica 2013, 190, 117–129. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Alimari, A.; Steiner, B.; Buerstmayr, H. Genetic mapping of QTL for resistance to Fusarium head blight spread (type 2 resistance) in a Triticum dicoccoides × Triticum durum backcross-derived population. Theor. Appl. Genet. 2013, 126, 2825–2834. [Google Scholar] [CrossRef]
- Bansal, U.K.; Kazi, A.G.; Singh, B.; Hare, R.A.; Bariana, H.S. Mapping of durable stripe rust resistance in a durum wheat cultivar Wollaroi. Mol. Breed. 2013, 33, 51–59. [Google Scholar] [CrossRef]
- Faris, J.D.; Zhang, Q.; Chao, S.; Zhang, Z.; Xu, S.S. Analysis of agronomic and domestication traits in a durum × cultivated emmer wheat population using a high-density single nucleotide polymorphism-based linkage map. Theor. Appl. Genet. 2014, 127, 2333–2348. [Google Scholar] [CrossRef]
- Russo, M.A.; Ficco, D.B.M.; Laidò, G.; Marone, D.; Papa, R.; Blanco, A.; Gadaleta, A.; De Vita, P.; Mastrangelo, A.M. A dense durum wheat × T dicoccum linkage map based on SNP markers for the study of seed morphology. Mol. Breed. 2014, 34, 1579–1597. [Google Scholar] [CrossRef]
- Desiderio, F.; Guerra, D.; Rubiales, D.; Piarulli, L.; Pasquini, M.; Mastrangelo, A.M.; Simeone, R.; Blanco, A.; Cattivelli, L.; Valé, G. Identification and mapping of quantitative trait loci for leaf rust resistance derived from a tetraploid wheat Triticum dicoccum accession. Mol. Breed. 2014, 34, 1659–1675. [Google Scholar] [CrossRef]
- Alsaleh, A.; Baloch, F.S.; Derya, M.; Azrak, M.; Kilian, B.; Nachit, M.M. Genetic linkage map of Anatolian durum wheat derived from a cross of Kunduru- 1149 × Cham1. Plant Mol. Biol. Rep. 2015, 33, 209–220. [Google Scholar] [CrossRef]
- Prat, N.; Guilbert, C.; Prah, U.; Wachter, E.; Steiner, B.; Langin, T.; Robert, O.; Buerstmayr, H. QTL mapping of Fusarium head blight resistance in three related durum wheat populations. Theor. Appl. Genet. 2017, 130, 13–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, C.; Basnet, B.R.; Singh, R.P.; Huerta-Espino, J.; Herrera-Foessel, S.A.; Ren, Y.; Randhawa, M.S. Genetic analysis and mapping of adult plant resistance loci to leaf rust in durum wheat cultivar Bairds. Theor. Appl. Genet. 2017, 130, 609–619. [Google Scholar] [CrossRef]
- Nilsen, K.T.; N’Diaye, A.; MacLachlan, P.R.; Clarke, J.M.; Ruan, Y.; Cuthbert, R.D.; Knox, R.E.; Wiebe, K.; Cory, A.T.; Walkowiak, S.; et al. High density mapping and haplotype analysis of the major stem-solidness locus SSt1 in durum and common wheat. PLoS ONE 2017, 12, e0175285. [Google Scholar] [CrossRef]
- Marcotuli, I.; Gadaleta, A.; Mangini, G.; Signorile, A.M.; Zacheo, S.A.; Blanco, A.; Simeone, R.; Colasuonno, P. Development of a high-density SNP-based linkage map and detection of QTL for beta-glucans.; protein content.; grain yield per spike and heading time in durum wheat. Int. J. Mol. Sci. 2017, 18, 1329. [Google Scholar] [CrossRef]
- Giunta, F.; De Vita, P.; Mastrangelo, A.M.; Sanna, G.; Motzo, R. Environmental and genetic variation for yield-related traits of durum wheat as affected by development. Front. Plant Sci. 2018, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Leng, Y.; Chao, S.; Xu, S.S.; Zhong, S. Molecular mapping of QTL for Fusarium head blight resistance introgressed into durum wheat. Theor. Appl. Genet. 2018, 131, 1939–1951. [Google Scholar] [CrossRef]
- Sari, E.; Berraies, S.; Knox, R.E.; Singh, A.K.; Ruan, Y.; Cuthbert, R.D.; Pozniak, C.J.; Henriquez, M.A.; Kumar, S.; Burt, A.J.; et al. High density genetic mapping of Fusarium head blight resistance QTL in tetraploid wheat. PLoS ONE 2018, 13, e0204362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatiukha, A.; Filler, N.; Lupo, I.; Lidzbarsky, G.; Klymiuk, V.; Korol, A.B.; Pozniak, C.; Tzion, F.; Krugmar, T. Grain protein content and thousand kernel weight QTLs identified in a durum × wild emmer wheat mapping population tested in five environments. Theor. Appl. Genet. 2020, 133, 119–131. [Google Scholar] [CrossRef]
- Bassi, F.M.; Brahmi, H.; Sabraoui, A.; Amri, A.; Nsarellah, N.; Nachit, M.M.; Al-Abdallat, A.; Chen, M.S.; Lazraq, A.; El Bouhssini, E. Genetic identification of loci for Hessian fly resistance in durum wheat. Mol. Breed. 2019, 39, 24. [Google Scholar] [CrossRef] [Green Version]
- Pirseyedi, S.M.; Kumar, A.; Ghavami, F.; Hegstad, J.B.; Mergoum, M.; Mazaheri, M.; Elias, E.M. Mapping QTL for Fusarium head blight resistance in a Tunisian-derived durum wheat population. Cereal Res. Commun. 2019, 47, 78–87. [Google Scholar] [CrossRef]
- Desiderio, F.; Zarei, L.; Licciardello, S.; Cheghamirza, K.; Farshadfar, E.; Virzi, N.; Sciacca, F.; Bagnaresi, P.; Battaglia, R.; Guerra, D.; et al. Genomic regions from an Iranian landrace increase kernel size in durum wheat. Front. Plant Sci. 2019, 10, 448. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.S.; Katherine, L.D.; Running, K.L.D.; Xu, S.S.; Zhang, Q.; Peters Haugrud, A.R.; Sharma, S.; McClean, P.E.; Faris, J.D. Genetic analysis of threshability and other spike traits in the evolution of cultivated emmer to fully domesticated durum wheat. Mol. Genet. Genom. 2019, 294, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Aoun, M.; Kolmer, J.A.; Rouse, M.N.; Elias, E.M.; Breiland, M.; Bulbula, W.D.; Chao, S.; Acevedo, M. Mapping of novel leaf rust and stem rust resistance genes in the Portuguese durum wheat landrace PI 192051. G3 2019, 9, 2535–2547. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Salsman, E.; Wang, R.; Galagedara, N.; Zhang, Q.; Fiedler, J.D.; Liu, Z.; Xu, S.; Faris, J.D.; Li, X. Meta-QTL analysis of tan spot resistance in wheat. Theor. Appl. Genet. 2020, 133, 2363–2375. [Google Scholar] [CrossRef]
- Marcotuli, I.; Colasuonno, P.; Hsieh, Y.S.Y.; Fincher, G.B.; Gadaleta, A. Non-Starch Polysaccharides in Durum Wheat: A Review. Int. J. Mol. Sci. 2020, 21, 2933. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.C.; Andreescu, C.; Breseghello, F.; Finney, P.L.; Gualberto, D.G.; Bergman, C.J.; Peña, R.J.; Perretant, M.R.; Leroy, P.; Qualset, C.O.; et al. Quantitative trait locus analysis of wheat quality traits. Euphytica 2006, 149, 145–159. [Google Scholar] [CrossRef]
- Zhang, W.; Chao, S.; Manthey, F.; Chicaiza, O.; Brevis, J.C.; Echenique, V.; Dubcovsky, J. QTL analysis of pasta quality using a composite microsatellite and SNP map of durum wheat. Theor. Appl. Genet. 2008, 117, 1361–1377. [Google Scholar] [CrossRef]
- Mann, G.; Diffey, S.; Cullis, B.; Azanza, F.; Martin, D.; Kelly, A.; McIntyre, L.; Schmidt, A.; Ma, W.; Nath, Z.; et al. Genetic control of wheat quality: Interactions between chromosomal regions determining protein content and composition.; dough rheology.; and sponge and dough baking properties. Theor. Appl. Genet. 2009, 118, 1519–1537. [Google Scholar] [CrossRef]
- Raman, R.; Allen, H.; Raman, H.; Diffey, S.; Martin, P.; McKelvie, K. Localisation of quantitative trait loci for quality attributes in a doubled haploid population of wheat (Triticum aestivum L.). Genome 2009, 52, 701–715. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, J.; Crouch, J.H.; Xu, Y. Efficiency of selective genotyping for genetic analysis of complex traits and potential applications in crop improvement. Mol. Breed. 2010, 26, 493–511. [Google Scholar] [CrossRef]
- Nigro, D.; Gadaleta, A.; Mangini, G.; Colasuonno, P.; Marcotuli, I.; Giancaspro, A.; Giove, S.L.; Simeone, R.; Blanco, A. Candidate genes and genome-wide association study of grain protein content and protein deviation in durum wheat. Planta 2019, 249, 1157–1175. [Google Scholar] [CrossRef]
- Colasuonno, P.; Lozito, M.L.; Marcotuli, I.; Nigro, D.; Giancaspro, A.; Mangini, G.; De Vita, P.; Mastrangelo, A.M.; Pecchioni, N.; Houston, K.; et al. The carotenoid biosynthetic and catabolic genes in wheat and their association with yellow pigments. BMC Genom. 2017, 18, 122. [Google Scholar] [CrossRef] [Green Version]
- Colasuonno, P.; Marcotuli, I.; Blanco, A.; Maccaferri, M.; Condorelli, G.E.; Tuberosa, R.; Parada, R.; de Camargo, A.C.; Schwember, A.R.; Gadaleta, A. Carotenoid pigment contentin durum wheat (Triticum turgidum l var durum), an overview of quantitative trait loci and candidate genes. Front. Plant Sci. 2019, 10, 1347. [Google Scholar] [CrossRef] [Green Version]
- Marcotuli, I.; Colasuonno, P.; Cutillo, S.; Simeone, R.; Blanco, A.; Gadaleta, A. β-glucan content in a panel of Triticum and Aegilops genotypes. Genet. Resour. Crop Evol. 2019, 66, 897–907. [Google Scholar] [CrossRef]
- Colasuonno, P.; Marcotuli, I.; Cutillo, S.; Simeone, R.; Blanco, A.; Gadaleta, A. Effect of barley chromosomes on the β-glucan content of wheat. Genet. Resour. Crop Evol. 2020, 67, 561–567. [Google Scholar] [CrossRef]
- Seib, P.A. Wheat starch: Isolation, structure and properties. J. Appl. Glycoscience 1994, 41, 49–69. [Google Scholar]
- Limbalkar, O.M.; Sharma, J.B.; Jha, S.K.; Mallick, N.; Niranjana, M.; Vinod, N. Mapping of genes for leaf and stem rust resistance in bread wheat genotype selection 212. Indian J. Genet. Plant Breed. 2019, 79, 1–8. [Google Scholar] [CrossRef]
- Kthiri, D.; Loladze, A.; N’Diaye, A.; Nilsen, K.T.; Walkowiak, S.; Dreisigacker, S.; Ammar, K.; Pozniak, C.J. Mapping of genetic loci conferring resistance to leaf rust from three globally resistant durum wheat sources. Front. Plant Sci. 2019, 10, 1247. [Google Scholar] [CrossRef]
- Kang, Y.; Zhou, M.; Merry, A.; Barry, K. Mechanisms of powdery mildew resistance of wheat—A review of molecular breeding. Plant Pathol. 2020, 69, 601–617. [Google Scholar] [CrossRef]
- Gadaleta, A.; Colasuonno, P.; Give, S.L.; Blanco, A.; Giancaspro, A. Map-based cloning of QFhbmgb-2A identifies a WAK2 gene responsible for Fusarium Head Blight resistance in wheat. Sci. Rep. 2019, 9, 6929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royo, C.; Maccaferri, M.; Álvaro, F.; Moragues, M.; Sanguineti, M.C.; Tuberosa, R.; Maalouf, F.; García del Moral, L.F.; Demontis, A.; Rhouma, S.; et al. Understanding the relationships between genetic and phenotypic structures of a collection of elite durum wheat accessions. Field Crops Res. 2010, 119, 91–105. [Google Scholar] [CrossRef]
- Bergkamp, B.; Impa, S.; Asebedo, A.; Fritz, A.; Jagadish, S.K. Prominent winter wheat varieties response to post-flowering heat stress under controlled chambers and field based heat tents. Field Crops Res. 2018, 222, 143–152. [Google Scholar] [CrossRef]
- Hlaváčová, M.; Klem, K.; Rapantová, B.; Novotná, K.; Urban, O.; Hlavinka, P.; Smutna, P.; Horakova, V.; Skarpa, P.; Pohankova, E.; et al. Interactive effects of high temperature and drought stress during stem elongation.; anthesis and early grain filling on the yield formation and photosynthesis of winter wheat. Field Crops Res. 2018, 221, 182–195. [Google Scholar] [CrossRef]
- Ugarte, C.; Calderini, D.F.; Slafer, G.A. Grain weight and grain number responsiveness to pre-anthesis temperature in wheat, barley and triticale. Field Crops Res. 2007, 100, 240–248. [Google Scholar] [CrossRef]
- Royo, C.; Villegas, D.; Rharrabti, Y.; Blanco, R.; Martos, V.; Garcia del Moral, L.F. Grain growth and yield formation of durum wheat grown at contracting latitudes and water regimes in a Mediterranean environment. Cereal Res. Commun. 2006, 34, 1021–1028. [Google Scholar] [CrossRef]
- Altenbach, S.B. New insights into the effects of high temperature.; drought and post-anthesis fertilizer on wheat grain development. J. Cereal Sci. 2012, 56, 39–50. [Google Scholar] [CrossRef]
- Dolferus, R. To grow or not to grow, A stressful decision for plants. Plant Sci. 2014, 229, 247–261. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Mujeeb-Kazi, A.; Sawkins, M. Prospects for utilizing plant-adaptive mechanisms to improve wheat and other crops in drought- and salinity-prone environments. Ann. Appl. Biol. 2005, 146, 239–259. [Google Scholar] [CrossRef]
- Salekdeh, G.H.; Reynolds, M.; Bennett, J.; Boyer, J. Conceptual framework for drought phenotyping during molecular breeding. Trends Plant Sci. 2009, 14, 488–496. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Chenu, K.; Biddulph, B.; Moeller, C.; Deery, D.M.; Rattey, A.R.; Bennett, D.; Barrett-Lennard, E.G.; Mayer, J.E. A multisite managed environment facility for targeted trait and germplasm phenotyping. Funct. Plant Biol. 2013, 40, 1–13. [Google Scholar] [CrossRef]
- Graybosch, R.A.; Peterson, C.J. Genetic improvement in winter wheat yields in the Great Plains of North America, 1959–2008. Crop Sci. 2010, 50, 1882–1890. [Google Scholar] [CrossRef]
- Bechtold, N.; Bouchez, D. Planta Agrobacterium-Mediated Transformation of Adult Arabidopsis thaliana Plants by Vacuum Infiltration. In Gene Transfer to Plants Springer Lab Manual; Potrykus, I., Spangenberg, I.G., Eds.; Springer: Berlin, Germany, 1995; pp. 19–23. [Google Scholar]
- Zhang, Q.; Bartels, D. Molecular responses to dehydration and desiccation in desiccation-tolerant angiosperm plants. J. Exp. Bot. 2018, 69, 3211–3222. [Google Scholar] [CrossRef] [PubMed]
- Royo, C.; Villegas, D. Field measurements of canopy spectra for biomass assessment of small-grain cereals. In Biomass-Detection, Production and Usage; Matovic, D., Ed.; InTech: Rijeka, Croatia, 2007; ISBN 978-953-307-492-4. [Google Scholar]
- Tuberosa, R. Phenotyping for drought tolerance of crops in the genomics era. Front. Physiol. 2012, 3, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Hassouni, K.; Belkadi, B.; Filali-Maltouf, A.; Tidiane-Sall, A.; Al-Abdallat, A.; Nachit, M.; Bassi, F.M. Loci controlling adaptation to heat stress occurring at the reproductive stage in durum wheat. Agromomy 2019, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Bento, M.; Pereira, S.G.; Viegas, W.; Silva, M. Durum wheat diversity for heat stress tolerance during inflorescence emergence is correlated to TdHSP101C expression in early developmental stages. PLoS ONE 2017, 28, e0190085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Kouadria, R.; Bouzouina, M.; Azzouz, R.; Lotmani, B. Salinity stress resistance of durum wheat (Triticum durum) enhanced by fungi. Int. J. Biosci. 2018, 12, 70–77. [Google Scholar]
- Longin, F.; Sieber, A.N.; Reif, J.C. Combining frost tolerance, high grain yield and good pasta quality in durum wheat. Plant Breed. 2013, 132, 353–358. [Google Scholar] [CrossRef]
- Soriano, J.M.; Malosetti, M.; Roselló, M.; Sorrells, M.E.; Royo, C. Dissecting the old Mediterranean durum wheat genetic architecture for phenology.; biomass and yield formation by association mapping and QTL meta-analysis. PLoS ONE 2017, 12, e0178290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chardon, F.; Virlon, B.; Moreau, L.; Falque, M.; Joet, J.; Decousset, L.; Murigneux, A.; Charcosset, A. Genetic architecture of flowering time in maize as inferred from quantitative trait loci meta-analysis and synteny conservation with the rice genome. Genetics 2004, 168, 2169–2185. [Google Scholar] [CrossRef] [Green Version]
- Flint-Garcia, S.A.; Thornsberry, J.M.; Buckler, E.S. Structure of linkage disequilibrium in plants. Ann. Rev. Plant Biol. 2003, 54, 357–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breseghello, F.; Sorrells, M.S. Association mapping of kernel size and milling quality in wheat (Triticum aestivum L.) cultivars. Genetics 2006, 172, 1165–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccaferri, M.; Sanguineti, M.C.; Demontis, A.; El-Ahmed, A.; Garcia del Moral, L.; Maalouf, F.; Nachit, M.; Nserallah, N.; Ouabbou, H.; Rhouma, S.; et al. Association mapping in durum wheat grown across a broad range of water regimes. J. Exp. Bot. 2011, 62, 409–438. [Google Scholar] [CrossRef] [Green Version]
- Laido, G.; Marone, D.; Russo, M.A.; Colecchia, S.A.; Mastrangelo, A.M.; De Vita, P.; Papa, R. Linkage disequilibrium and genome-wide association mapping in tetraploid wheat (Triticum turgidum L.). PLoS ONE 2014, 9, e95211. [Google Scholar] [CrossRef] [Green Version]
- Peleg, Z.; Cakmak, I.; Ozturk, L.; Yazici, A.; Jun, Y.; Budak, H.; Korol, A.B.; Fahima, T.; Saranga, Y. Quantitative trait loci conferring grain mineral nutrient concentrations in durum wheat × wild emmer wheat RIL population. Theor. Appl. Genet. 2009, 119, 353–369. [Google Scholar] [CrossRef] [Green Version]
- Graziani, M.; Maccaferri, M.; Royo, C.; Salvatorelli, F.; Tuberosa, R. QTL dissection of yield components and morpho-physiological traits in a durum wheat elite population tested in contrasting thermo-pluviometric conditions. Crop Pasture Sci. 2014, 65, 80–95. [Google Scholar] [CrossRef]
- Nagel, M.; Navakode, S.; Scheibal, V.; Baum, M.; Nachit, M.; Röder, M.S.; Börner, A. The genetic basis of durum wheat germination and seedling growth under osmotic stress. Biol. Plant 2014, 58, 681–688. [Google Scholar] [CrossRef]
- Maccaferri, M.; El-Feki, W.; Nazemi, G.; Salvi, S.; Cané, M.A.; Colalongo, C.; Stefanelli, S.; Tuberosa, R. Prioritizing quantitative trait loci for root system architecture in tetraploid wheat. J. Exp. Bot. 2016, 67, 1161–1178. [Google Scholar] [CrossRef]
- Iannucci, A.; Marone, D.; Russo, M.A.; De Vita, P.; Miullo, V.; Ferragonio, P.; Blanco, A.; Gadaleta, A.; Mastrangelo, A.M. Mapping QTL for Root and shoot morphological traits in a durum wheat × T dicoccum segregating population at seedling stage. Int. J. Genom. 2017, 6876393. [Google Scholar] [CrossRef] [Green Version]
- Sanguineti, M.C.; Li, S.; Maccaferri, M.; Corneti, S.; Rotondo, F.; Chiari, T.; Tuberosa, R. Genetic dissection of seminal root architecture in elite durum wheat germplasm. Ann. Appl. Biol. 2007, 151, 291–305. [Google Scholar] [CrossRef]
- Canè, M.A.; Maccaferri, M.; Nazemi, G.; Salvi, S.; Francia, R.; Colalongo, C.; Tuberosa, R. Association mapping for root architectural traits in durum wheat seedlings as related to agronomic performance. Mol. Breed. 2014, 34, 1629–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condorelli, G.E.; Maccaferri, M.; Newcomb, M.; Andrade-Sanchez, P.; White, J.W.; French, A.N.; Sciara, G.; Ward, R.; Tuberosa, R. Comparative aerial and ground based high throughput phenotyping for the genetic dissection of NDVI as a proxy for drought adaptive traits in durum wheat. Front. Plant Sci. 2018, 9, 893. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stack, R.W.; Friesen, T.L.; Faris, J.D. Identification of a novel fusarium head blight resistance quantitative trait locus on chromosome 7A in tetraploid wheat. Phytopathol. 2007, 97, 592–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haile, J.K.; Nachit, M.M.; Hammer, K.; Badebo, A.; Röder, M.S. QTL mapping of resistance to race Ug99 of Puccinia graminis f.sp. tritici in durum wheat (Triticum durum Desf.). Mol. Breed. 2012, 30, 1479–1493. [Google Scholar]
- Maccaferri, M.; Francia, R.; Ratti, C.; Rubies-Autonell, C.; Colalongo, C.; Ferrazzano, G.; Tubersoa, R.; Sanguineti, M.C. Genetic analysis of Soil-Borne Cereal Mosaic Virus response in durum wheat: Evidence for the role of the major quantitative trait locus QSbm.ubo-2BS and of minor quantitative trait loci. Mol. Breed. 2012, 29, 973–988. [Google Scholar] [CrossRef]
- Ben-David, R.; Peleg, Z.; Dinoor, A.; Saranga, Y.; Korol, A.B.; Fahima, T. Genetic dissection of quantitative powdery mildew resistance loci in tetraploid wheat. Mol. Breed. 2014, 34, 1647–1658. [Google Scholar] [CrossRef]
- Zhang, Q.; Axtman, J.E.; Faris, J.D.; Shiaoman, C.; Zengcui, Z.; Timothy, L.F.; Zhong, S.; Cai, X.; Elias, E.M.; Xu, S.S. Identification and molecular mapping of quantitative trait loci for Fusarium head blight resistance in emmer and durum wheat using a single nucleotide polymorphism-based linkage map. Mol. Breed. 2014, 34, 1677–1687. [Google Scholar] [CrossRef]
- Kumar, S.; Knox, R.E.; Singh, A.K.; DePaw, R.M.; Campbell, H.R.; Isidro-Sanchez, J.; Clarke, F.R.; Pozniak, C.J.; N’Daye, A.; Meyer, B.; et al. High-density genetic mapping of a major QTL for resistance to multiple races of loose smut in a tetraploid wheat cross. PLoS ONE 2018, 13, e0192261. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; N’Diaye, A.; Walkowiak, S.; Nilsen, K.T.; Cory, A.T.; Haile, J.; Kutcher, H.R.; Ammar, K.; Loladze, A.; Huerta-Espino, J.; et al. Genetic analysis of resistance to stripe rust in durum wheat (Triticum turgidum L. var. durum). PLoS ONE 2018, 13, e0203283. [Google Scholar] [CrossRef]
- Gordon, A.; McCartney, C.; Knox, R.E.; Ereful, N.; Hiebert, C.W.; Konkin, D.J.; Hsueh, Y.C.; Bhadauria, V.; Sgroi, M.; O’Sullivan, D.M.; et al. Genetic and transcriptional dissection of resistance to Claviceps purpurea in the durum wheat cultivar Greenshank. Theor. Appl. Genet. 2020, 133, 1873–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghavami, F.; Elias, E.M.; Mamidi, S.; Ansari, O.; Sargolzaei, M.; Adhikari, T.; Mergoum, M.; Kianian, S.F. Mixed model association mapping for fusarium head blight resistance in Tunisian-derived durum wheat populations. G3 2011, 1, 209–218. [Google Scholar] [CrossRef]
- Letta, T.; Olivera, P.; Maccaferri, M.; Jin, Y.; Ammar, K.; Badebo, A.; Salvi, S.; Noli, E.; Crossa, J.; Tuberosa, R. Association mapping reveals novel stem rust resistance loci in Durum wheat at the seedling stage. Plant Genome 2014, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Aoun, M.; Breiland, M.; Kathryn Turner, M.; Loladze, A.; Chao, S.; Xu, S.S.; Ammar, K.; Anderson, J.A.; Kolmer, J.A.; Acevedo, M. Genome-wide association mapping of leaf rust response in a durum wheat worldwide germplasm collection. Plant Genome 2016, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidane, Y.G.; Hailemariam, B.N.; Mengistu, D.K.; Fadda, C.; Pè, M.E.; Dell’Acqua, M. Genome-Wide Association Study of Septoria tritici Blotch Resistance in Ethiopian Durum Wheat Landraces. Front. Plant Sci. 2017, 8, 1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Maccaferri, M.; Bulli, P.; Rynearson, S.; Tuberosa, R.; Chen, X.; Pumphrey, M. Genome-wide association mapping for seedling and field resistance to Puccinia striiformis f. sp. tritici in elite durum wheat. Theor. Appl. Genet. 2017, 130, 649–667. [Google Scholar] [CrossRef]
- Saccomanno, A.; Matny, O.; Marone, D.; Laidò, G.; Petruzzino, G.; Mazzucotelli, E.; Desiderio, F.; Blanco, A.; Gadaleta, A.; Pecchioni, N.; et al. Genetic mapping of loci for resistance to stem rust in a tetraploid wheat collection. Int. J. Mol. Sci. 2018, 19, 3907. [Google Scholar] [CrossRef] [Green Version]
- Steiner, B.; Buerstmayr, M.; Wagner, C.; Danler, A.; Eshonkulov, B.; Ehn, M.; Buerstmayr, H. Fine-mapping of the Fusarium head blight resistance QTL Qfhs.ifa-5A identifies two resistance QTL associated with anther extrusion. Theor. Appl. Genet. 2019, 132, 2039–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, V.; Roncallo, P.F.; Beaufort, V.; Cervigni, G.L.; Miranda, R.; Jensen, C.A.; Echenique, V.C. Mapping of main and epistatic effect QTLs associated with grain protein and gluten strength using a RIL population of durum wheat. J. Appl. Genet. 2011, 52, 287–298. [Google Scholar] [CrossRef]
- Roncallo, P.F.; Cervigni, G.L.; Jensen, C.; Miranda, R.; Carrera, A.D.; Helguera, M.; Echenique, V. QTL analysis of main and epistatic effects for flour color traits in durum wheat. Euphytica 2012, 185, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Xue, W.T.; Yang, R.Z.; Qin, H.B.; Zhao, G.; Tzion, F.; Cheng, J.P. Quantitative trait loci conferring grain selenium nutrient in durum wheat×wild emmer wheat RIL population. Czech. J. Genet. Plant Breed. 2018, 54, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Giancaspro, A.; Give, S.L.; Zacheo, S.A.; Blanco, A.; Gadaleta, A. Genetic Variation for Protein Content and Yield-Related Traits in a Durum Population Derived from an Inter-Specific Cross Between Hexaploid and Tetraploid Wheat Cultivars. Front. Plant Sci. 2019, 10, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.; Yu, B.; Knox, R.E.; Singh, A.K.; DePauw, R.; Cuthbert, R.; Zhang, W.; Piche, I.; Gao, P.; Sharpe, A.; et al. High density mapping of quantitative trait loci conferring gluten strength in canadian durum wheat. Front. Plant Sci. 2020, 11, 170. [Google Scholar] [CrossRef] [Green Version]
- Reimer, S.; Pozniak, C.J.; Clarke, F.R.; Clarke, J.M.; Somers, D.J.; Knox, R.E.; Singh, A.K. Association mapping of yellow pigment in an elite collection of durum wheat cultivars and breeding lines. Genome 2008, 51, 1016–1025. [Google Scholar] [CrossRef]
- Marcotuli, I.; Houston, K.; Waugh, R.; Fincher, G.B.; Burton, R.A.; Blanco, A.; Gadaleta, A. Genome wide association mapping for arabinoxylan content in a collection of tetraploid wheats. PLoS ONE 2015, 10, e0132787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- N’Diaye, A.; Haile, J.K.; Cory, A.T.; Clarke, F.R.; Clarke, J.M.; Knox, R.E.; Pozniak, C.J. Single marker and haplotype-based association analysis of semolina and pasta colour in elite durum wheat breeding lines using a high-density consensus map. PLoS ONE 2017, 12, e0170941. [Google Scholar]
- N’Diaye, A.; Haile, J.K.; Nilsen, K.T.; Walkowiak, S.; Ruan, Y.; Singh, A.K.; Clarke, F.R.; Clarke, J.M.; Pozniak, C.J. Haplotype loci under selection in Canadian durum wheat germplasm over 60 years of breeding: Association with grain yield, quality traits, protein loss, and plant height. Front. Plant Sci. 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Rosello, M.; Royo, C.; Alvaro, F.; Villegas, D.; Nazco, R.; Soriano, J.M. Pasta-making quality QTLome from Mediterranean durum wheat landraces. Front. Plant Sci. 2018, 9, 1512. [Google Scholar] [CrossRef]
- Goffinet, B.; Gerber, S. Quantitative Trait Loci: A metaanalysis. Genetics 2000, 155, 463–473. [Google Scholar]
- Löffler, M.; Schön, C.; Miedaner, T. Revealing the genetic architecture of FHB resistance in hexaploid wheat (Triticum aestivum L.) by QTL meta-analysis. Mol. Breed. 2009, 23, 473–488. [Google Scholar] [CrossRef]
- Hanocq, E.; Laperche, A.; Jaminon, O. Most significant genome regions involved in the control of earliness traits in bread wheat.; as revealed by QTL meta-analysis. Theor. Appl. Genet. 2007, 114, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Goudemand, E.; Laurent, V.; Duchalais, L.; Ghaffary, S.M.; Kema, G.H.; Lonnet, P.; Margalé, E.; Robert, O. Association mapping and meta-analysis: Two complementary approaches for the detection of reliable Septoria tritici blotch quantitative resistance in bread wheat (Triticum aestivum L.). Mol. Breed. 2013, 32, 563–584. [Google Scholar] [CrossRef]
- Marone, D.; Russo, M.A.; Laidò, G.; De Vita, P.; Papa, R.; Blanco, A.; Gadaleta, A.; Rubiales, D.; Mastrangelo, A.M. Genetic basis of qualitative and quantitative resistance to powdery mildew in wheat: From consensus regions to candidate genes. BMC Genom. 2013, 14, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano, J.M.; Royo, C. Dissecting the genetic architecture of leaf rust resistance in wheat by QTL meta-analysis. Phytopathology 2015, 105, 1585–1593. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, S.; Simmonds, J.; Leverington, M.; Wang, Y.; Fish, L.J.; Sayers, L.; Alibert, L.; Orford, S.; Wingen, L.; Snape, J. Meta-QTL analysis of the genetic control of crop height in elite European winter wheat germplasm. Mol. Breed. 2012, 29, 159–171. [Google Scholar] [CrossRef]
- Quraishi, U.M.; Murat, F.; Abrouk, M.; Pont, C.; Confolent, C.; Oury, F.X.; Ward, J.; Boros, D.; Gebruers, K.; Delcour, J.A.; et al. Combined meta-genomics analyses unravel candidate genes for the grain dietary fiber content in bread wheat (Triticum aestivum L.). Funct. Integr. Genom. 2011, 11, 71–83. [Google Scholar] [CrossRef]
- Tyagi, S.; Mir, R.R.; Balyan, H.S.; Gupta, P.K. Interval mapping and meta-QTL analysis of grain traits in common wheat (Triticum aestivum L.). Euphytica 2015, 201, 367–380. [Google Scholar] [CrossRef]
- Darzi-Ramandi, H.; Shariati, J.V.; Tavakol, E.; Najafi-Zarini, H.; Bilgrami, S.S.; Razavi, K. Detection of consensus genomic regions associated with root architecture of bread wheat on groups 2 and 3 chromosomes using QTL meta-analysis. Aust. J. Crop Sci. 2017, 11, 777. [Google Scholar] [CrossRef]
- Soriano, J.M.; Alvaro, F. Discovering consensus genomic regions in wheat for root-related traits by QTL meta-analysis. Sci. Rep. 2019, 9, 10537. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, S.; Gupta, P.K. Meta-analysis of QTLs involved in pre-harvest sprouting tolerance and dormancy in bread wheat. Triticeae Genom. Genet. 2012, 3, 9–24. [Google Scholar] [CrossRef]
- Liu, R.; Jia, H.; Cao, X.; Huang, J.; Li, F.; Tao, Y.; Qiu, F.; Zheng, Y.; Zhang, Z. Fine mapping and candidate gene prediction of a pleiotropic quantitative trait locus for yield-related trait in Zea mays. PLoS ONE 2012, 7, e49836. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Gonzalez, R.H.; Segovia, V.; Bird, N.; Fenwick, P.; Holdgate, S.; Berry, S.; Jack, P.; Caccamo, M.; Uauy, C. RNA-Seq bulked segregant analysis enables the identification of high-resolution genetic markers for breeding in hexaploid wheat. Plant Biotechnol. 2015, 13, 613–624. [Google Scholar] [CrossRef]
- Zou, C.; Wang, P.; Xu, Y. Bulked sample analysis in genetics.; genomics and crop improvement. Plant Biotechnol. 2016, 10, 1941–1955. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xie, J.; Zhang, H.; Guo, B.; Ning, S.; Chen, Y.; Lu, P.; Wu, Q.; Li, M.; Zhang, D.; et al. Mapping stripe rust resistance gene YrZH22 in Chinese wheat cultivar Zhoumai 22 by bulked segregant RNA-Seq (BSR-Seq) and comparative genomics analyses. Theor. Appl. Genet. 2017, 130, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Bassi, F.M.; Ghavami, F.; Hayden, M.J.; Wang, Y.; Forrest, K.L.; Kong, S.; Dizon, R.; Michalak de Jimenez, M.K.; Meinhardt, S.W.; Mergoum, M.; et al. Fast-forward genetics by radiation hybrids to saturate the locus regulating nuclear-cytoplasmic compatibility in Triticum. Plant Biotechnol. J. 2016, 14, 1716–1726. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Kulwal, P.L.; Mir, R.R. QTL mapping, methodology and applications in cereal breeding. In Cereal Genomics II; Gupta, P.K., Varshney, R.K., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 275–318. [Google Scholar]
- Huang, B.E.; Verbyla, K.L.; Verbyla, A.P. MAGIC populations in crops: Current status and future prospects. Theor. Appl. Genet. 2015, 128, 999–1017. [Google Scholar] [CrossRef] [PubMed]
- Milner, S.G.; Maccaferri, M.; Huang, B.E.; Mantovani, P.; Massi, A.; Frascaroli, E.; Tuberosa, R.; Salvi, S. A multiparental cross population for mapping QTL for agronomic traits in durum wheat (Triticum turgidum ssp. durum). Plant Biotechnol. J. 2015, 14, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Vatter, T.; Maurer, A.; Kopahnke, D.; Perovic, D.; Ordon, F.; Pillen, K. A nested association mapping population identifies multiple small effect QTL conferring resistance against net blotch (Pyrenophora teres f teres) in wild barley. PLoS ONE 2017, 12, 0186803. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Fang, X.; Jiang, P.; Zhang, G.; Hu, J.; Wang, X.; Meng, Q.; Cui, W.; Lan, S.; Ma, X.; et al. Genetic architecture of nitrogen-deficiency tolerance in wheat seedlings based on a nested association mapping (NAM) population. Front. Plant Sci. 2018, 9, 845. [Google Scholar] [CrossRef]
- McMullen, M.D.; Kresovich, S.; Villeda, H.S.; Bradbury, P.; Li, H.; Sun, Q.; Flint-Garcia, S.; Thornsberry, J.; Acharya, C.; Bottoms, C.; et al. Genetic properties of the maize nested association mapping population. Science 2009, 325, 737–740. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Holland, J.B.; McMullen, M.D.; Buckler, E.S. Genetic design and statistical power of nested association mapping in maize. Genetics 2008, 178, 539–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Cao, Y.; He, J.; Zhao, T.; Gai, J. Detecting the QTL-allele system conferring flowering date in a nested association mapping population of soybean using a novel procedure. Theor. Appl. Genet. 2017, 130, 2297–2314. [Google Scholar] [CrossRef]
- Bouchet, S.; Olatoye, M.O.; Marla, S.R.; Perumal, R.; Tesso, T.; Yu, J.; Tuinstra, M.; Morris, G.P. Increased power to dissect adaptive traits in global sorghum diversity using a nested association mapping population. Genetics 2017, 206, 573–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nice, L.M.; Steffenson, B.J.; Blake, T.K.; Horsley, R.D.; Smith, K.P.; Muehlbauer, G.J. Mapping agronomic traits in a wild barley advanced backcross-nested association mapping population. Crop Sci. 2017, 57, 1199–2210. [Google Scholar] [CrossRef]
- Bajgain, P.; Rouse, M.N.; Tsilo, T.J.; Macharia, G.K.; Bhavani, S.; Jin, Y.; Anderson, J.A. Nested association mapping of stem rust resistance in wheat using genotyping by sequencing. PLoS ONE 2016, 11, e0155760. [Google Scholar] [CrossRef]
- Alahmad, S.; El Hassouni, K.; Bassi, F.M.; Dinglasan, E.; Youssef, C.; Quarry, G.; Aksoy, A.; Mazzucotelli, E.; Juhász, A.; Able, J.A.; et al. A Major Root Architecture QTL Responding to Water Limitation in Durum Wheat. Front. Plant Sci. 2019, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Kidane, Y.G.; Gesesse, C.A.; Hailemariam, B.N.; Desta, E.A.; Mengistu, D.K.; Fadda, C.; Pè, M.E.; Dell’Acqua, M. A large nested association mapping population for breeding and quantitative trait locus mapping in Ethiopian durum wheat. Plant Biotechnol. J. 2019, 17, 1380–1393. [Google Scholar] [CrossRef]
- Alahmad, S.; Kang, Y.; Dinglasan, E.; Mazzucotelli, E.; Voss-Fels, K.P.; Able, J.A.; Christopher, J.; Bassi, F.M.; Hickey, L.T. Adaptive Traits to Improve Durum Wheat Yield in Drought and Crown Rot Environments. Int. J. Mol. Sci. 2020, 21, 5260. [Google Scholar] [CrossRef]
- Soriano, J.; Villegas, D.; Aranzana, M.; García del Moral, L.; Royo, C. Genetic structure of modern durum wheat cultivars and mediterranean landraces matches with their agronomic performance. PLoS ONE 2016, 11, e0160983. [Google Scholar] [CrossRef] [Green Version]
- Soriano, J.M.; Villegas, D.; Sorrells, M.E.; Royo, C. Durum Wheat Landraces from East and West Regions of the Mediterranean Basin Are Genetically Distinct for Yield Components and Phenology. Front. Plant Sci. 2018, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Rommens, J.M.; Iannuzzi, M.C.; Kerem, B.; Drumm, M.L.; Melmer, G.; Dean, M.; Rozmahel, R.; Cole, J.L.; Kennedy, D.; Hidaka, N.; et al. Identification of the cystic fibrosis gene: Chromosome walking and jumping. Science 1989, 245, 1059–1065. [Google Scholar] [CrossRef]
- Bechtold, U.; Ferguson, J.N.; Mullineaux, P.M. To defend or to grow, lessons from Arabidopsis C24. J. Exp. Bot. 2018, 69, 2809–2821. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.F.; McMullen, M.D. Defining genes for agricultural traits, QTL analysis and the candidate gene approach. Probe 1996, 7, 24–27. [Google Scholar]
- De Vienne, D. La génomique pour l’étude des gènes contrôlant la variation des caractères d’intérêt agronomique. Oléagineux Corps Gras Lipides 1999, 6, 143–148. [Google Scholar]
- Pflieger, S.; Lefebvre, V.; Causse, M. The candidate gene approach in plant genetics: A review. Mol. Breed. 2001, 7, 275–291. [Google Scholar] [CrossRef]
- Faris, J.D.; Li, W.L.; Liu, D.J.; Chen, P.D.; Gill, B.S. Candidate gene analysis of quantitative disease resistance in wheat. Theor. Appl. Genet. 1999, 98, 219–225. [Google Scholar] [CrossRef]
- Marcotuli, I.; Colasuonno, P.; Blanco, A.; Gadaleta, A. Expression analysis of cellulose synthase-like genes in durum wheat. Sci. Rep. 2018, 8, 15675. [Google Scholar] [CrossRef] [PubMed]
- James, R.A.; von Caemmerer, S.; Condon, A.G.; Zwart, A.B.; Munns, R. Genetic variation in tolerance to the osmotic stress component of salinity stress in durum wheat. Funct. Plant Biol. 2008, 35, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-González, R.H.; Borrill, P.; Lang, D.; Harrington, S.A.; Brinton, J.; Venturini, L.; Davey, M.; Jacobs, J.; van Ex, F.; Pasha, A.; et al. The transcriptional landscape of polyploid wheat. Science 2018, 361, eaar6089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyers, B.T. Camoco: A net for the sea of candidate genes. Plant Cell 2018, 30, 2889. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Lazarus, E.Z.; Rhee, S.Y. QTG-Finder2: A Generalized Machine-Learning Algorithm for Prioritizing QTL Causal Genes in Plants. G3 2020, 10, 2411–2421. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Gao, L.; Wang, S.; Li, X.Y.; Wei, X.J.; Zhang, Y.J.; Wang, H.Y.; Liu, D.Q. Expression and functional analysis of a pathogenesis-related protein 1 gene, TcLr19PR1, involved in wheat resistance against leaf rust fungus. Plant Mol. Biol. Rep. 2015, 33, 797–805. [Google Scholar] [CrossRef]
- Liu, W.; Frick, M.; Huel, R.; Nykiforuk, C.L.; Wang, X.; Gaudet, D.A.; Eudes, F.; Conner, R.L.; Kuzyk, A.; Chen, Q.; et al. The stripe rust resistance gene Yr10 encodes an evolutionary-conserved and unique CC-NBS-LRR sequence in wheat. Mol. Plant 2014, 7, 1740–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.N.; Shen, F.D.; Hong, Y.T.; Rong, W.; Du, L.P.; Liu, X.; Xu, H.; Ma, L.; Zhang, Z. The wheat calcium-dependent protein kinase TaCPK7-D positively regulates host resistance to sharp eyespot disease. Mol. Plant Pathol. 2016, 17, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, X.; Wang, X.; Zhou, M.; Zhou, X.; Ye, X.; Wei, X. An R2R3 MYB transcription factor in wheat, TaPIMP1, mediates host resistance to Bipolaris sorokiniana and drought stresses through regulation of defense- and stress-related genes. New Phytol. 2012, 196, 1155–1170. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.P.; Liu, J.X.; Chen, K.L.; Qiu, J.L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 12617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.L.; Wang, D.; Gao, C. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef]
- Camerlengo, F.; Frittelli, A.; Sparks, C.; Doherty, A.; Martignago, D.; Larré, C.; Lupi, R.; Sestili, F.; Masci, F. CRISPR-Cas9 multiplex editing of the α-amylase/trypsin inhibitor genes to reduce allergen proteins in durum wheat. Front. Sustain. Food Syst. 2020, 4, 104. [Google Scholar] [CrossRef]
- Kalunke, R.M.; Tundo, S.; Sestili, F.; Camerlengo, F.; Lafiandra, D.; Lupi, R.; Larré, C.; Denery-Papini, S.; Islam, S.; Ma, W.; et al. Reduction of allergenic potential in bread wheat rnai transgenic lines silenced for cm3, cm16 and 0.28 ati genes. Int. J. Mol. Sci. 2020, 21, 5817. [Google Scholar] [CrossRef]
- Loss, S.P.; Siddique, K.H.M. Morphological and Physiological Traits Associated with Wheat Yield Increases in Mediterranean Environments. Adv. Agron. 1994, 52, 229–276. [Google Scholar]
- Sanchez-Garcia, M.; Royo, C.; Aparicio, N.; Martín-Sánchez, J.A.; Alvaro, F. Genetic improvement of bread wheat yield and associated traits in Spain during the 20th century. J. Agric. Sci. 2013, 151, 105–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Garcia, M.; Álvaro, F.; Peremarti, A.; Martín-Sánchez, J.A.; Royo, C. Changes in bread-making quality attributes of bread wheat varieties cultivated in Spain during the 20th century. Eur. J. Agron. 2015, 63, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Royo, C.; Martos, V.; Ramdani, A.; Villegas, D.; Rharrabti, Y.; García Del Moral, L.F. Changes in yield and carbon isotope discrimination of Italian and Spanish durum wheat during the 20th century. Agron. J. 2008, 100, 352–360. [Google Scholar] [CrossRef]
- Jackson, P.; Robertson, M.; Cooper, M.; Hammer, G. The role of physiological understanding in plant breeding; from a breeding perspective. Field Crops Res. 1996, 49, 1–37. [Google Scholar] [CrossRef]
- Goddard, M.; Hayes, B. Mapping genes for complex traits in domestic animals and their use in breeding programmes. Nat. Rev. Genet. 2009, 10, 381–391. [Google Scholar] [CrossRef]
- Isidro, J.; Akdemir, D.; Burke, J. Genomic selection. In The World Wheat Book: A History of Wheat Breeding; William, A., Alain, B., Maarten, V.G., Eds.; Lavoisier: Paris, France, 2016; pp. 1001–1023. [Google Scholar]
- Fiedler, J.D.; Salsman, E.; Liu, Y.; Michalak de Jiménez, M.; Hegstad, J.B.; Chen, B.; Manthey, F.A.; Chao, S.; Xu, S.; Elias, E.M.; et al. Genome-Wide Association and Prediction of Grain and Semolina Quality Traits in Durum Wheat Breeding Populations. Plant Genome 2017, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Haile, J.K.; N’Diaye, A.; Clarke, F.; Clarke, J.; Knox, R.; Rutkosky, J.; Bassi, F.M.; Pozniak, C.J. Genomic selection for grain yield and quality traits in durum wheat. Mol. Breed. 2018, 38, 75. [Google Scholar] [CrossRef]
- Zaïm, M.; Kabbaj, H.; Kehel, Z.; Gorjanc, G.; Filali-Maltouf, A.; Belkadi, B.; Nachit, M.M.; Bassi, F.M. Combining QTL Analysis and Genomic Predictions for Four Durum Wheat Populations Under Drought Conditions. Front. Genet. 2020, 11, 316. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Amores, J.; Michel, S.; Miedaner, T.; Longin, C.F.H.; Buerstmayr, H. Genomic predictions for Fusarium head blight resistance in a diverse durum wheat panel: An effective incorporation of plant height and heading date as covariates. Euphytica 2020, 216, 22. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.; Ghosh, S.; Williams, M.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alahmad, S.; Dinglasan, E.; Leung, K.M.; Riaz, A.; Derbal, N.; Voss-Fels, K.P.; Hickey, L.T. Speed breeding for multiple quantitative traits in durum wheat. Plant Methods 2018, 14, 36. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Quantitative trait loci (QTL) projection onto the consensus map developed by Maccaferri et al. [31]. Color code: green, abiotic QTL; orange, biotic QTL; blue, quality QTL.
Figure 1.
Quantitative trait loci (QTL) projection onto the consensus map developed by Maccaferri et al. [31]. Color code: green, abiotic QTL; orange, biotic QTL; blue, quality QTL.


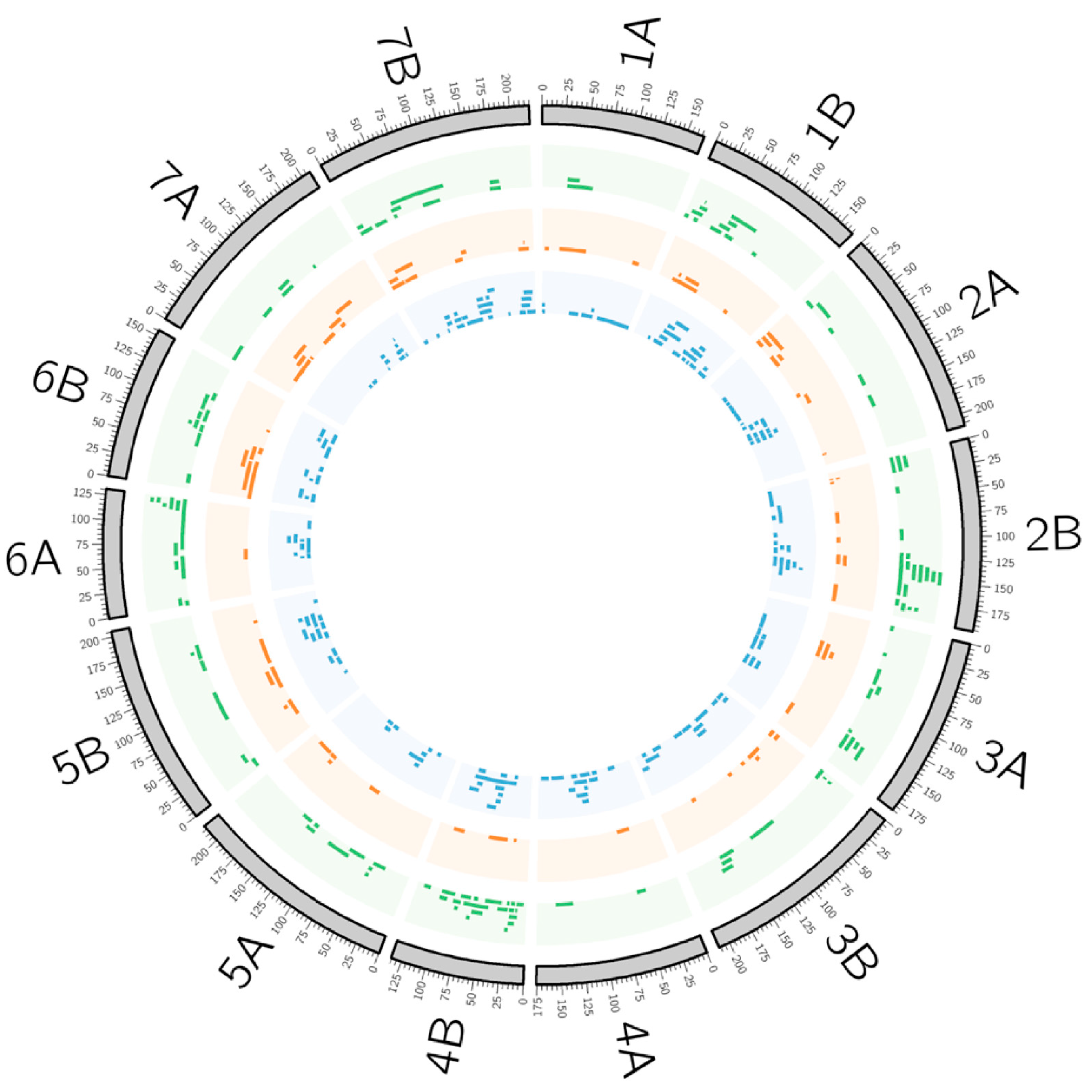{kind=link}
Table 1.
Durum wheat linkage maps.
| Cross | Type | N Progenies | N Markers | Marker Type | Distance (cM) | Reference |
|---|---|---|---|---|---|---|
| Messapia × MG4343 | RIL | 65 | 213 | RFLP, biochemical | 1352 | [39] |
| Messapia × MG4343 | RIL | 65 | 292 | SSR, RFLP, biochemical | 2034 | [43] |
| Messapia × MG4343 | RIL | 65 | 293 | AFLP, RFLP, biochemical | 2063 | [44] |
| Jennah Khetifa × Cham 1 | RIL | 110 | 306 | AFLP, SSP, SSR | 3598 | [45] |
| Messapia × MG4343 | RIL | 65 | 458 | AFLP, RFLP, SSR, biochemical, morphological | 1352 | [46] |
| Omrabi 5 × 600545 | RIL | 114 | 295 | AFLP, SSP, SSR | 2289 | [47] |
| Strongfield × Blackbird | DH | 85 | 424 | SSR | 2052 | [48] |
| W9262-260D3 × Kofa | DH | 155 | 194 | DArT, SSR | 1400 | [49] |
| Colosseo × Lloyd | RIL | 176 | 554 | DArT, SSR | 2022 | [50] |
| Langdon × G1816 | RIL | 152 | 669 | DArT, SSR | 2317 | [51] |
| Kofa × UC1113 | RIL | 93 | 269 | SNP, SSR | 2140 | [52] |
| Svevo × Ciccio | RIL | 120 | 522 | SSR | 1605 | [11] |
| Creso × Pedroso | RIL | 123 | 456 | DArT, SSR | 1800 | [53] |
| DT695 × Strongfield | DH | 185 | 345 | DArT, SSR | 1474 | [54] |
| Latino × Primadur | RIL | 121 | 454 | DArT, SSR | 1172 | [10] |
| Neodur × Cirillo | RIL | 146 | 414 | DArT, SSR, STS | 1917 | [55] |
| Svevo × Ciccio | RIL | 120 | 833 | DArT, SSR, morphological | 1716 | [7] |
| DS × Td161 | BC | 134 | 368 | AFLP, SSR | 1921 | [56] |
| Floradur × Td161 | BC | 129 | 248 | AFLP, SSR | 1414 | [56] |
| Helidur × Td161 | BC | 126 | 239 | AFLP, SSR | 1515 | [56] |
| BGRC3487/2 × DT735 | RIL | 160 | 948 | DArT, SSR | 1089 | [57] |
| Rugby × Maier | DH | 105 | 228 | DArT, SSR | 801 | [6] |
| PDW233 × Bhalegaon 4 | RIL | 140 | 245 | SSR, ISSR, TRAP, SCAR, STS, biochemical | 2773 | [58] |
| Gerizim #36 × Helidur | RIL | 103 | 421 | AFLP, SSR | 1808 | [59] |
| Wollaroi × Bansi | RIL | 92 | 799 | DArT, SSR | 3859 | [60] |
| Svevo × Ciccio | RIL | 120 | 5670 | DArT, SNP, SSR | 1774 | [5] |
| Ben × PI41025 | RIL | 200 | 2593 | SNP, SSR, STS | 2444 | [61] |
| Simeto × Molise Colli | RIL | 136 | 9040 | SNP | 2879 | [62] |
| Latino × MG5323 | RIL | 110 | 10840 | SNP, SSR | 2363 | [63] |
| Kofa × Svevo | RIL | 249 | 311 | SNP, SSR | 1259 | [4] |
| Gallareta × Demetra | DH | 127 | 147 | DArT, SSR | 1017 | [4] |
| DT707 × DT696 | DH | 127 | 136 | DArT, SSR | 861 | [4] |
| DT712 × Blackbird | DH | 89 | 392 | SSR | 1848 | [4] |
| Lebsock × PI94749 | DH | 146 | 240 | SSR, biochemical | 1463 | [4] |
| PDW1216 × MvTD10-98 | RIL | 182 | 440 | SSR | 984 | [4] |
| W9292-260D3 × Kofa | DH | 155 | 3712 | SNP, SSR | 1685 | [31] |
| Svevo × Zavitan | RIL | 140 | 10911 | SNP | 2258 | [31] |
| Simeto × Levante | RIL | 180 | 5978 | DArT, SNP, SSR | 2185 | [31] |
| Mohawk × Cocorit69 | RIL | 81 | 5554 | SNP | 2013 | [31] |
| Meridiano × Claudio | RIL | 180 | 5970 | DArT, SNP, SSR | 2239 | [31] |
| Colosseo × Lloyd | RIL | 176 | 7946 | DArT, SNP, SSR | 2064 | [31] |
| Kunduru-1149 × Cham 1 | RIL | 141 | 395 | AFLP, SSP, SSR | 4854 | [64] |
| 02-5B-318 × Saragolla | RIL | 135 | 4366 | SNP | 4227 | [30] |
| Karur × DBC-480 | RIL | 110 | 1609 | DArTseq, SSR | 2806 | [65] |
| Durobonus × DBC-480 | RIL | 100 | 1052 | DArTseq, SSR | 1781 | [65] |
| SZD1029K × DBC-480 | RIL | 100 | 1006 | DArTseq, SSR | 2219 | [65] |
| Bairds × Atred#1 | RIL | 137 | 1150 | SNP, SSR | 2639 | [66] |
| W9262-260D3 × Kofa | DH | 155 | 4227 | SNP | 2282 | [67] |
| Duilio × Avonlea | RIL | 134 | 5444 | SNP | 1962 | [68] |
| Ofanto × Cappelli | RIL | 98 | 9267 | DArT, DArTseq, SSR | 2119 | [69] |
| Joppa × 10Ae564 | RIL | 205 | 5216 | SNP | 3428 | [70] |
| DT707 × DT696 | DH | 127 | 2943 | SNP | 1808 | [71] |
| Strongfield × Blackbird | DH | 85 | 9568 | SNP | 2763 | [71] |
| Svevo × Y12-3 | RIL | 208 | 4166 | SNP | 2169 | [72] |
| Faraj × Gidara 2 | RIL | 143 | 2067 | SNP | 2578 | [73] |
| Tunisian 108 × Ben | BIL | 178 | 329 | DArT, SSR | 1888 | [74] |
| Zardak × Iran249 | RIL | 118 | 6195 | SNP, SSR | 2884 | [75] |
| Rusty × PI193883 | RIL | 190 | 9346 | SNP, SSR | 2440 | [76] |
| Rusty × PI192051-1 | RIL | 180 | 1138 | SNP | 1436 | [77] |
| Rusty × PI 387336 | RIL | 200 | 2894 | SNP | 2858 | [78] |
| Rusty × PI 387696 | RIL | 200 | 2059 | SNP | 2724 | [78] |
| Rusty × PI 466979 | RIL | 200 | 3692 | SNP | 2776 | [78] |
| Rusty × Iumillo | RIL | 200 | 2911 | SNP | 2968 | [78] |
BC: backcross; DH: double haploid; RIL: recombinant inbred line; AFLP: amplified fragment length polymorphism; DArT: diversity array technology; ISSR: inter simple sequence repeat; RFLP: restriction fragment length polymorphism; SCAR: sequence characterized amplified region; SNP: single nucleotide polymorphism; SSR: simple sequence repeat; STS: sequence-tagged sites; TRAP: target region amplification polymorphism.
Table 2.
Summary of quantitative trait loci (QTL) reviewed in the present work.
| Reference | Cross | Type | N Genotypes | N QTL | Traits |
|---|---|---|---|---|---|
| Abiotic stress | |||||
| [121] | Langdon × G18-16 | RIL | 156 | 31 | CIR, OP, CC, FLRI |
| [122] | Kofa × Svevo | RIL | 247 | 12 | PDL, SPAD, NDVI |
| [123] | Omrabi5 × Belikh2 | RIL | 114 | 6 | CL, RRT |
| [124] | Colosseo × Lloyd | RIL | 176 | 28 | RRT |
| [124] | Meridiano × Caludio | RIL | 181 | 32 | RRT |
| [125] | Simeto × Mollise Colli | RIL | 136 | 18 | RRT |
| [126] | Elite cultivars | GWAS | 57 | 4 | RRT |
| [127] | Elite cultivars | GWAS | 183 | 2 | RRT |
| [124] | Elite cultivars | GWAS | 183 | 31 | RRT |
| [128] | UNIBO-DP | GWAS | 248 | 73 | DB, NDVI, SPAD |
| Biotic stress | |||||
| [48] | Strongfield × Blackbird | DH | 85 | 2 | FHB |
| [129] | LDN × LDN-Dic7A | RIL | 118 | 1 | FHB |
| [8] | Colosseo × Lloyd | RIL | 176 | 1 | LR |
| [119] | Meridiano × Claudio | RIL | 181 | 1 | SBCMV |
| [56] | DS × Td161 | BC | 134 | 1 | FHB |
| [56] | Floradur × Td161 | BC | 129 | 3 | FHB |
| [56] | Helidur × Td161 | BC | 126 | 1 | FHB |
| [130] | Kristal × Sebatel | RIL | 85 | 7 | SR |
| [131] | Simeto × Levante | RIL | 180 | 7 | SBCMV |
| [57] | BGRC3487 × 2 * DT735 | RIL | 160 | 2 | FHB |
| [55] | Neodur × Cirillo | RIL | 146 | 2 | SBCMV |
| [60] | Wollaroi × Bansi | RIL | 92 | 2 | YR |
| [59] | Gerizim × Helidur | RIL | 103 | 1 | FHB |
| [132] | Langdon × G18-16 | RIL | 157 | 4 | PM |
| [63] | Latino × MG5323 | RIL | 110 | 3 | LR |
| [133] | Ben × PI41025 | RIL | 200 | 3 | FHB |
| [94] | Sumai-3 × Saragolla | RIL | 135 | 11 | FHB |
| [65] | Karur × DBC-480 | RIL | 111 | 1 | FHB |
| [134] | Strongfield × Blackbird | DH | 90 | 2 | LS |
| [135] | Kofa × W9262-260D3 | DH | 155 | 1 | YR |
| [70] | Joppa × 10Ae564 | RIL | 205 | 3 | FHB |
| [77] | Rusty × PI 192051-1 | RIL | 180 | 5 | LR |
| [74] | Ben × Tunisian 108 | BIL | 171 | 3 | FHB |
| [136] | Greenshank × AC Avonlea | DH | 132 | 4 | CP |
| [137] | Different sources | GWAS | 323 | 3 | FHB |
| [138] | Elite cultivars | GWAS | 183 | 8 | SR |
| [139] | Worldwide collection | GWAS | 496 | 50 | LR |
| [140] | Ethiopian landraces | GWAS | 318 | 20 | STB |
| [141] | Elite cultivars | GWAS | 250 | 16 | YR |
| [135] | Elite cultivars | GWAS | 92 | 1 | YR |
| [142] | Tetraploid panel | GWAS | 230 | 37 | SR |
| [143] | Spring lines | GWAS | 228 | 7 | FHB |
| Quality | |||||
| [81] | UC1113 × Kofa | BP | 93 | 5 | YPC |
| [121] | Langdon × G18-16 | RIL | 152 | 55 | GCaC, GCuC, GFeC, GKC, GMgC, GMnC, GPC, GSC, GZnC, PGC |
| [54] | DT695 × Strongfield | DH | 185 | 6 | GPC |
| [10] | Latino × Primadur | BP | 121 | 4 | YPC |
| [144] | UC1113 × Kofa | RIL | 93 | 18 | GPC, SV |
| [145] | UC1113 × Kofa | BP | 93 | 13 | F, YPC |
| [5] | Svevo × Ciccio | BP | 120 | 7 | YPC |
| [68] | Duilio × Avonlea | RIL | 134 | 2 | BG |
| [146] | Langdon × G18-16 | RIL | 152 | 15 | GSeC, GSeY |
| [87] | Colosseo × Lloyd | BP | 176 | 12 | YPC |
| [87] | Kofa × Svevo | BP | 249 | 4 | YPC |
| [87] | Meridiano × Claudio | BP | 181 | 6 | YPC |
| [72] | Svevo × Y12-3 | RIL | 208 | 9 | GPC |
| [147] | Saragolla × 02-5B-318 | RIL | 135 | 9 | GPC |
| [148] | Pelissier × Strongfield | DH | 162 | 6 | SV |
| [149] | Worldwide elite collection | GWAS | 93 | 20 | YPC |
| [150] | Agrogen | GWAS | 104 | 19 | AX |
| [35] | Agrogen | GWAS | 230 | 7 | BG |
| [86] | Durum collection | GWAS | 124 | 6 | YPC |
| [151] | Canadian durum wheats | GWAS | 169 | 6 | YPC |
| [152] | Canadian durum lines | GWAS | 192 | 28 | YPC |
| [153] | Mediterranean landraces | GWAS | 172 | 14 | GPC, GS, TW, YPC |
Abiotic stress: CC: Chlorophyll content; CIR: carbon isotope ratio; CL: coleoptile length; DB: dry biomass; FLRI: flag leaf rolling index; NDVI: normalized difference vegetation index; OP: osmotic potential; PDL: length of the ear peduncle; RRT: root-related trait; SPAD: Chlorophyll concentration measure. Biotic stress: CP: Claviceps purpurea; FHB: Fusarium head blight; LR: leaf rust; LS: loose smut; PM: powdery mildew; SBCMV: soil-borne cereal mosaic virus; SR: stem rust; STB: Septoria tritici blotch; YR: yellow rust. Quality: AX: Arabinoxylan; BG: β-glucan; Fb: flour yellow color; GCaC: grain calcium concentration; GCdC: grain cadmium concentration; GCuC: grain copper concentration; GFeC: grain iron concentration; GKC: grain potassium concentration; GMgC: grain magnesium concentration; GMnC: grain manganese concentration; GPC: grain protein content; GS: gluten strength; GSC: grain sulfur concentration; GSeC: grain selenium concentration; GSeY: grain selenium yield; GZnC: grain zinc concentration; PGC: phosphorus grain concentration; SV: SDS-sedimentation volume; TW: test weight; YPC: yellow pigment content.
Table 3.
Comparison between biparental QTL mapping and genome-wide association study (GWAS).
| Linkage QTL Mapping | GWAS |
|---|---|
| Two known ancestors (parents) | Multiple (unknown) ancestors |
| Short known recombination history | Long (unknown) recombination history |
| Simple population structure | Complex population structure |
| LD caused by linkage | LD caused by different genetic events |
| Requires construction of specific maps | Existing maps can be used |
| Contrasting genetic background | Diverse genetic background |
| Phenotyping is required for new populations | Phenotyping data might already be available |
Table 4.
The number of QTL per single trait and chromosome.
| Trait | 1A | 1B | 2A | 2B | 3A | 3B | 4A | 4B | 5A | 5B | 6A | 6B | 7A | 7B | Total | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Abiotic | CC | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 7 | |||||||
| CIR | 1 | 1 | 2 | 1 | 1 | 6 | ||||||||||
| CL | 1 | 1 | 2 | 1 | 5 | |||||||||||
| DB | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | |||||||
| FLRI | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 9 | |||||||
| NDVI | 1 | 3 | 3 | 6 | 1 | 6 | 2 | 3 | 3 | 3 | 4 | 3 | 4 | 5 | 47 | |
| OP | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | ||||||
| PDL | 1 | 1 | 1 | 1 | 4 | |||||||||||
| RRT | 10 | 0 | 6 | 17 | 14 | 4 | 17 | 7 | 3 | 14 | 9 | 7 | 8 | 116 | ||
| SPAD | 1 | 2 | 2 | 2 | 1 | 5 | 2 | 3 | 3 | 1 | 1 | 2 | 25 | |||
| Total | 15 | 7 | 13 | 32 | 19 | 18 | 6 | 28 | 15 | 15 | 21 | 16 | 13 | 19 | 237 | |
| Biotic | CP | 1 | 1 | 1 | 1 | 4 | ||||||||||
| FHB | 1 | 2 | 7 | 3 | 3 | 5 | 2 | 2 | 4 | 2 | 4 | 4 | 3 | 42 | ||
| LR | 6 | 1 | 7 | 9 | 4 | 6 | 4 | 3 | 5 | 1 | 4 | 2 | 4 | 3 | 59 | |
| LS | 1 | 1 | 2 | |||||||||||||
| PM | 1 | 1 | 1 | 1 | 4 | |||||||||||
| SBCMV | 1 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 10 | |||||||
| SR | 1 | 3 | 2 | 6 | 1 | 4 | 10 | 0 | 1 | 3 | 6 | 3 | 4 | 8 | 52 | |
| STB | 7 | 1 | 10 | 1 | 1 | 20 | ||||||||||
| YR | 2 | 3 | 1 | 2 | 1 | 1 | 2 | 1 | 4 | 3 | 20 | |||||
| Total | 16 | 9 | 21 | 24 | 19 | 18 | 16 | 7 | 11 | 12 | 12 | 13 | 17 | 18 | 213 | |
| Quality | AX | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 3 | 1 | 1 | 3 | 1 | 19 | ||
| BG | 1 | 3 | 2 | 1 | 2 | 9 | ||||||||||
| Fb | 2 | 1 | 1 | 1 | 2 | 7 | ||||||||||
| GCaC | 1 | 4 | 5 | |||||||||||||
| GCuC | 2 | 5 | 3 | 10 | ||||||||||||
| GFeC | 1 | 5 | 3 | 1 | 10 | |||||||||||
| GKC | 8 | 8 | ||||||||||||||
| GMgC | 2 | 2 | ||||||||||||||
| GMnC | 2 | 2 | ||||||||||||||
| GPC | 2 | 4 | 6 | 4 | 1 | 2 | 4 | 3 | 2 | 3 | 1 | 1 | 1 | 5 | 39 | |
| GS | 1 | 1 | 1 | 3 | ||||||||||||
| GSC | 1 | 4 | 5 | |||||||||||||
| GSeC | 2 | 2 | 1 | 4 | 9 | |||||||||||
| GseY | 1 | 2 | 1 | 1 | 1 | 6 | ||||||||||
| GZnC | 6 | 6 | ||||||||||||||
| PGC | 2 | 1 | 3 | |||||||||||||
| SV | 2 | 2 | 2 | 2 | 3 | 1 | 1 | 2 | 1 | 16 | ||||||
| TW | 1 | 1 | 1 | 3 | ||||||||||||
| YPC | 7 | 11 | 5 | 1 | 10 | 6 | 7 | 7 | 6 | 10 | 3 | 22 | 13 | 108 | ||
| Total | 15 | 22 | 23 | 18 | 10 | 20 | 18 | 19 | 15 | 17 | 15 | 16 | 31 | 31 | 270 | |
| 46 | 38 | 57 | 74 | 48 | 56 | 40 | 54 | 41 | 44 | 48 | 45 | 61 | 68 | 720 |
Color gradient for the number of traits per chromosomes: from green (lower number) to red (higher number). Color gradient for the number of QTL per trait: low intensity (lower number) to high intensity (higher number). Abiotic stress (green): CC: Chlorophyll content; CIR: carbon isotope ratio; CL: coleoptile length; DB: dry biomass; FLRI: plag leaf rolling index; NDVI: normalized difference vegetation index; OP: osmotic potential; PDL: length of the ear peduncle; RRT: root-related trait; SPAD: Chlorophyll concentration measure. Biotic stress (red): CP: Claviceps purpurea; FHB: Fusarium head blight; LR: leaf rust; LS: loose smut; PM: powdery mildew; SBCMV: soil-borne cereal mosaic virus; SR: stem rust; STB: Septoria tritici blotch; YR: yellow rust. Quality (blue): AX: Arabinoxylan; BG: β-glucan; Fb: flour yellow color; GCaC: grain calcium concentration; GCdC: grain cadmium concentration; GCuC: grain copper concentration; GFeC: grain iron concentration; GKC: grain potassium concentration; GMgC: grain magnesium concentration; GMnC: grain manganese concentration; GPC: grain protein content; GS: gluten strength; GSC: grain sulfur concentration; GSeC: grain selenium concentration; GSeY: grain selenium yield; GZnC: grain zinc concentration; PGC: phosphorus grain concentration; SV: SDS-sedimentation volume; TW: test weight; YPC: yellow pigment content.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Colasuonno, P.; Marcotuli, I.; Gadaleta, A.; Soriano, J.M. From Genetic Maps to QTL Cloning: An Overview for Durum Wheat. Plants 2021, 10, 315. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020315
AMA Style
Colasuonno P, Marcotuli I, Gadaleta A, Soriano JM. From Genetic Maps to QTL Cloning: An Overview for Durum Wheat. Plants. 2021; 10(2):315. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020315
Chicago/Turabian StyleColasuonno, Pasqualina, Ilaria Marcotuli, Agata Gadaleta, and Jose Miguel Soriano. 2021. "From Genetic Maps to QTL Cloning: An Overview for Durum Wheat" Plants 10, no. 2: 315. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10020315
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.