
Potential Plant–Plant Communication Induced by Infochemical Methyl Jasmonate in Sorghum (Sorghum bicolor)

 , , , and
, , , and 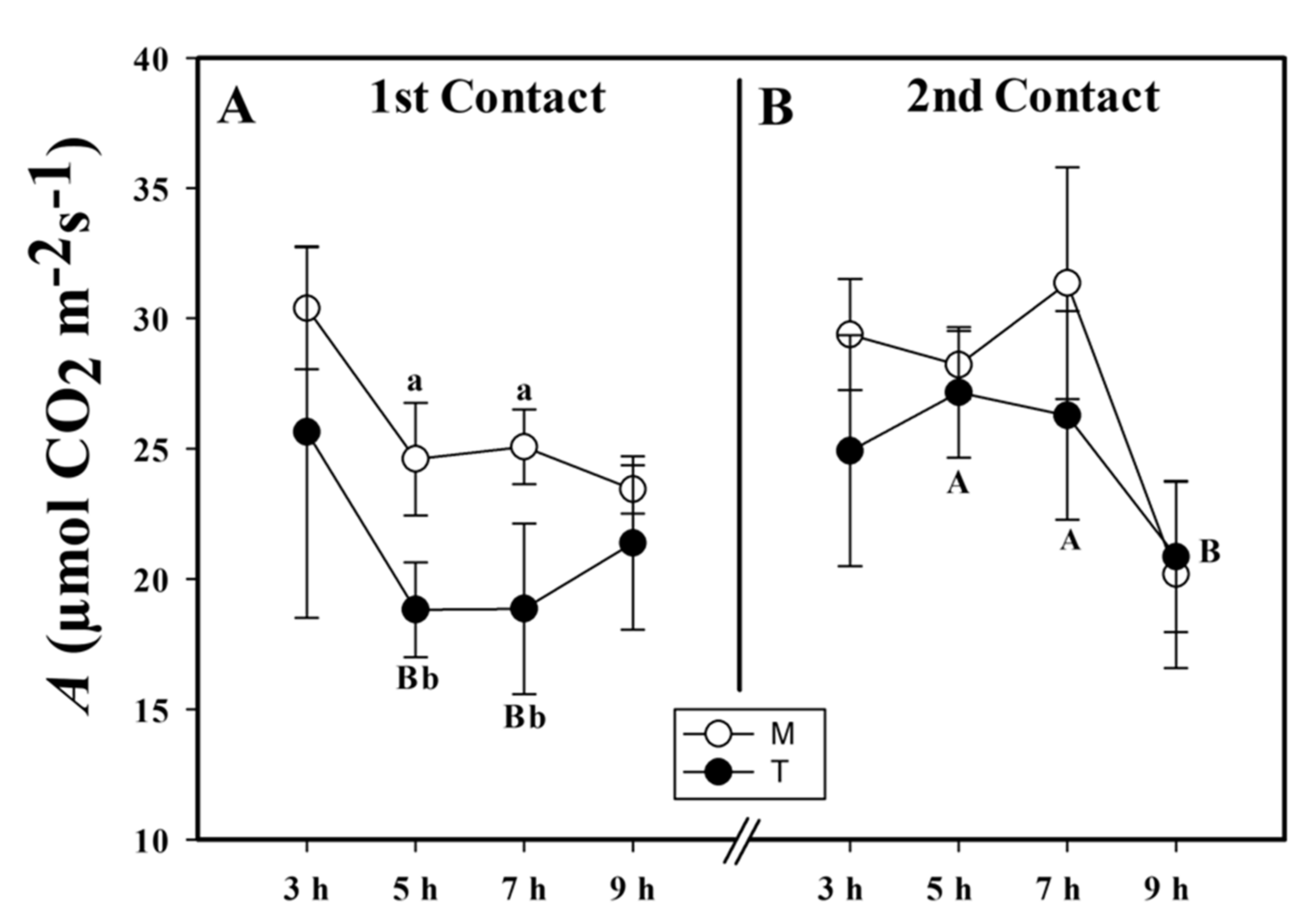{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Assimilation Rate
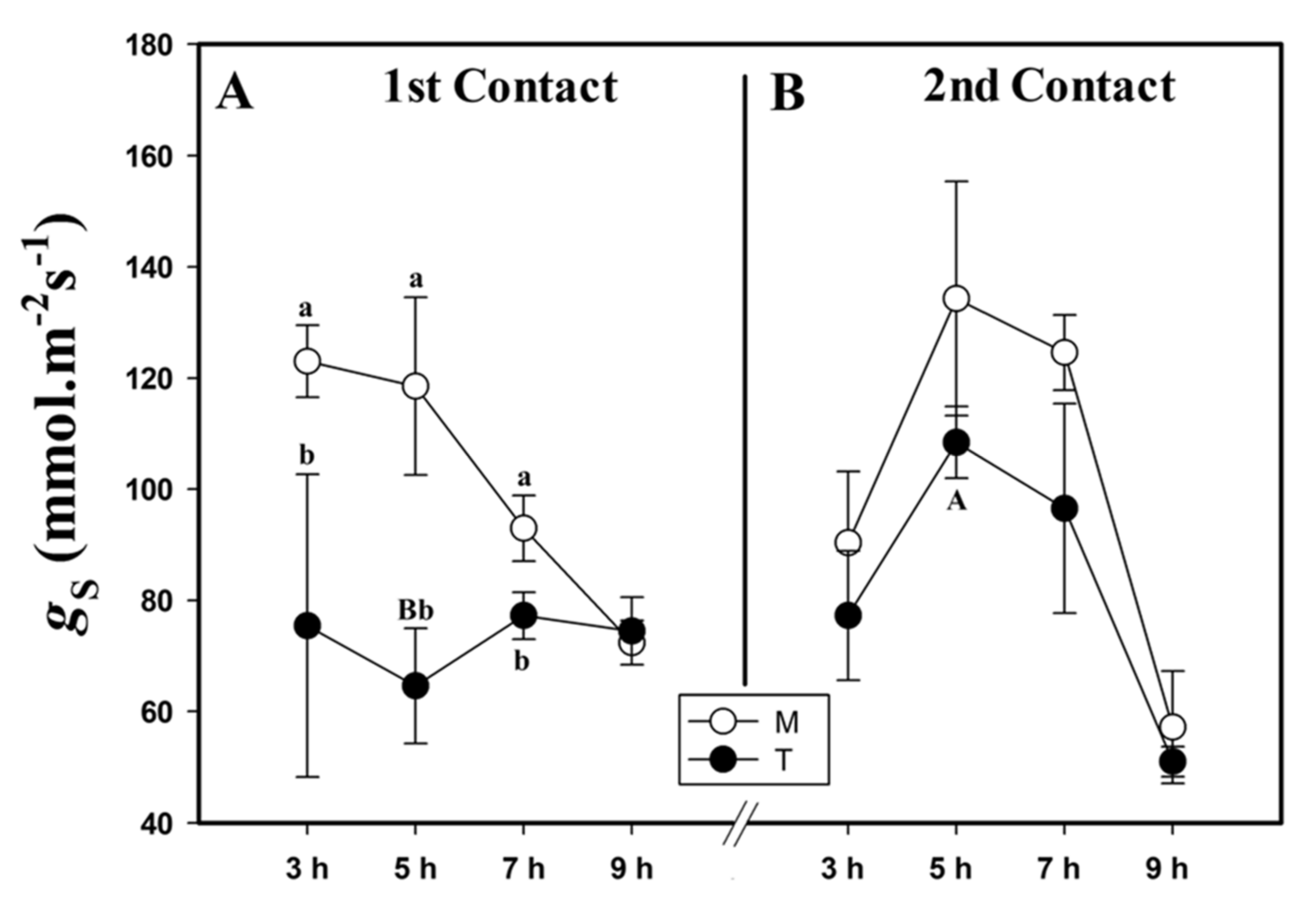
2.2. Stomatal Conductance
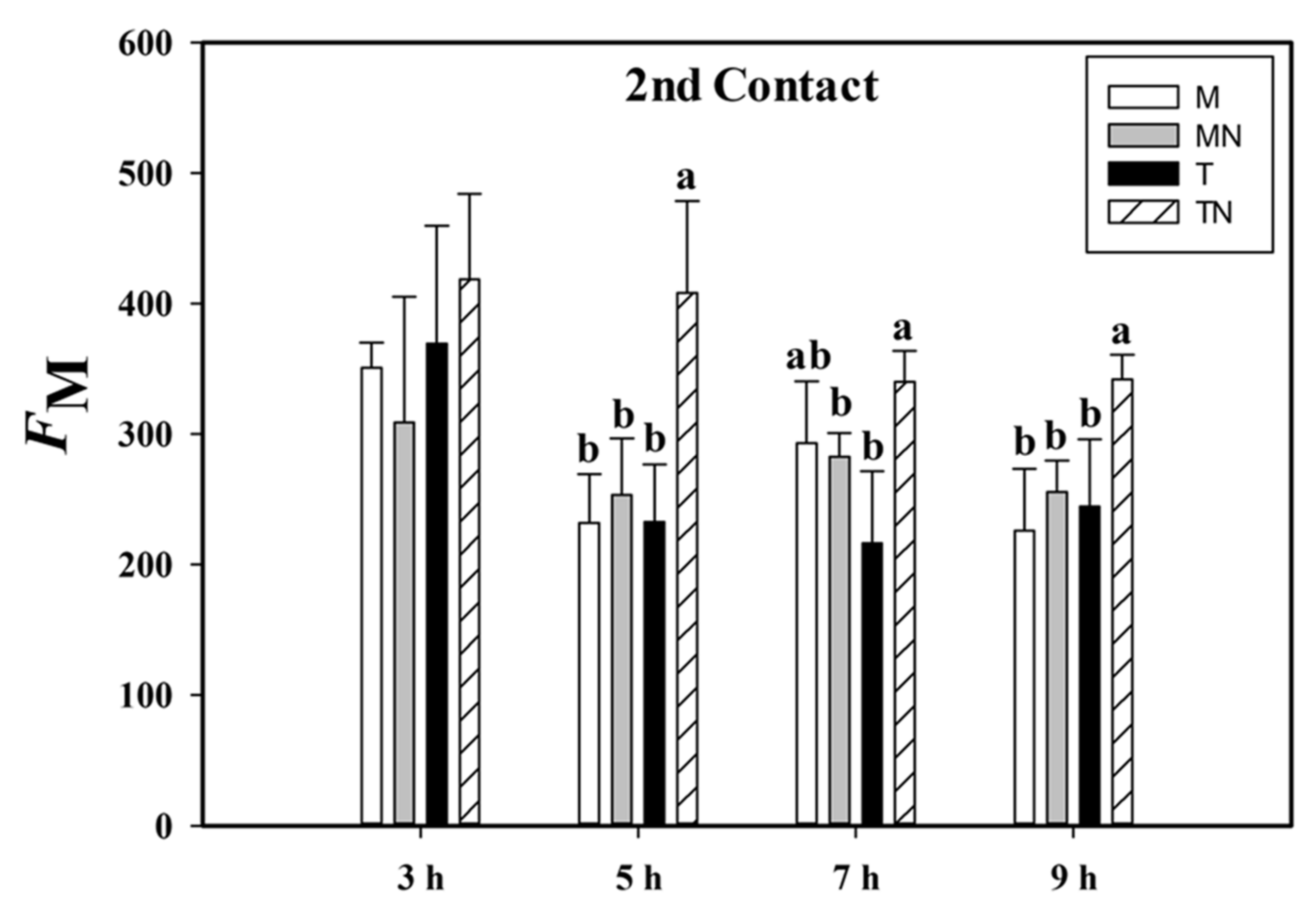
2.3. Maximum Fluorescence
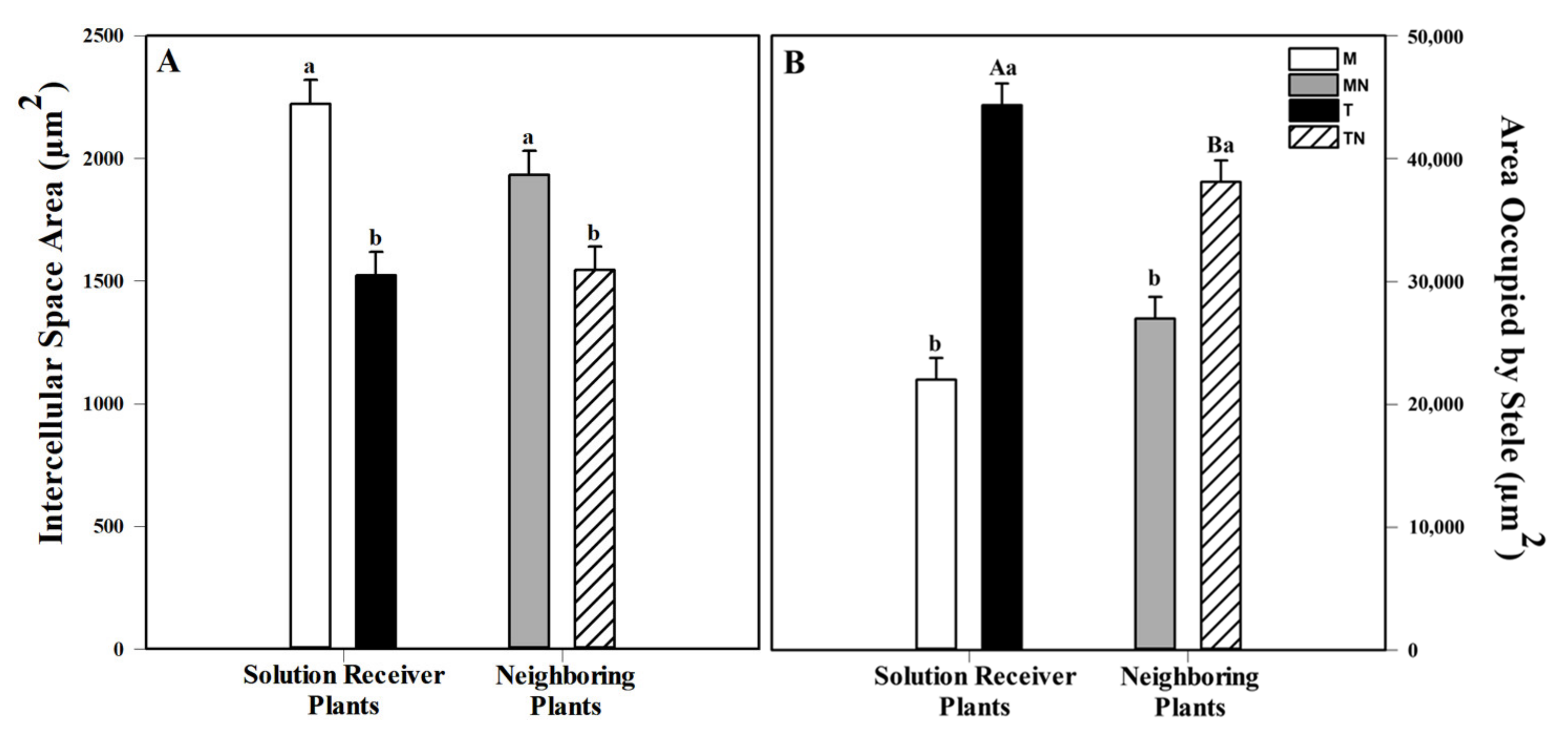
2.4. Anatomical Analyses of Adventitious Roots
3. Discussion
4. Materials and Methods
4.1. Plants and Growth Conditions
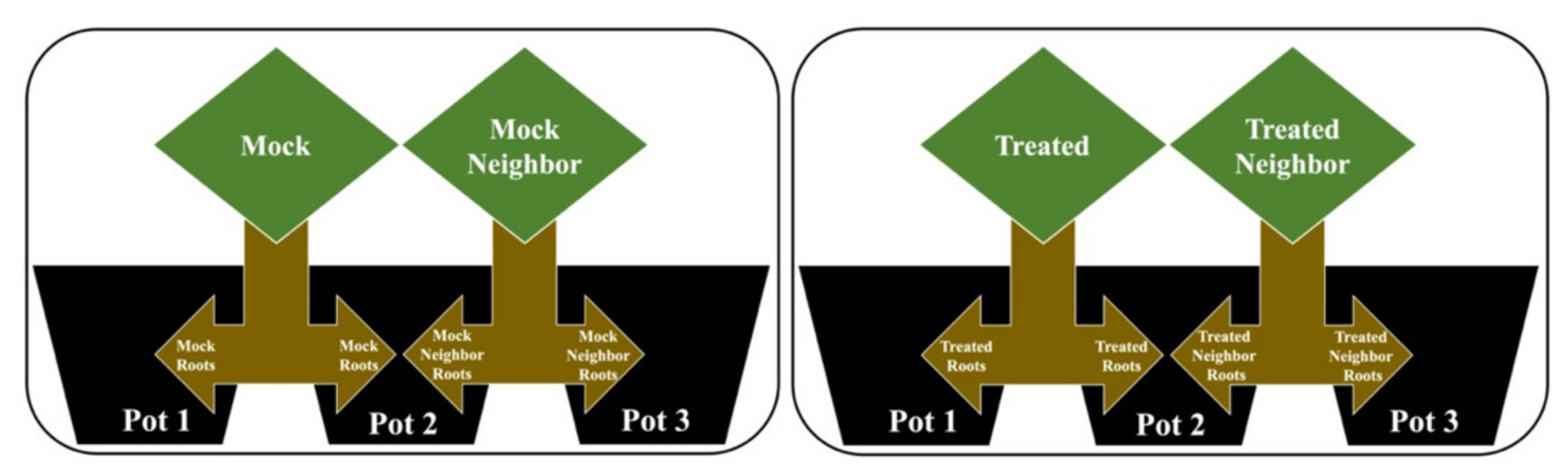
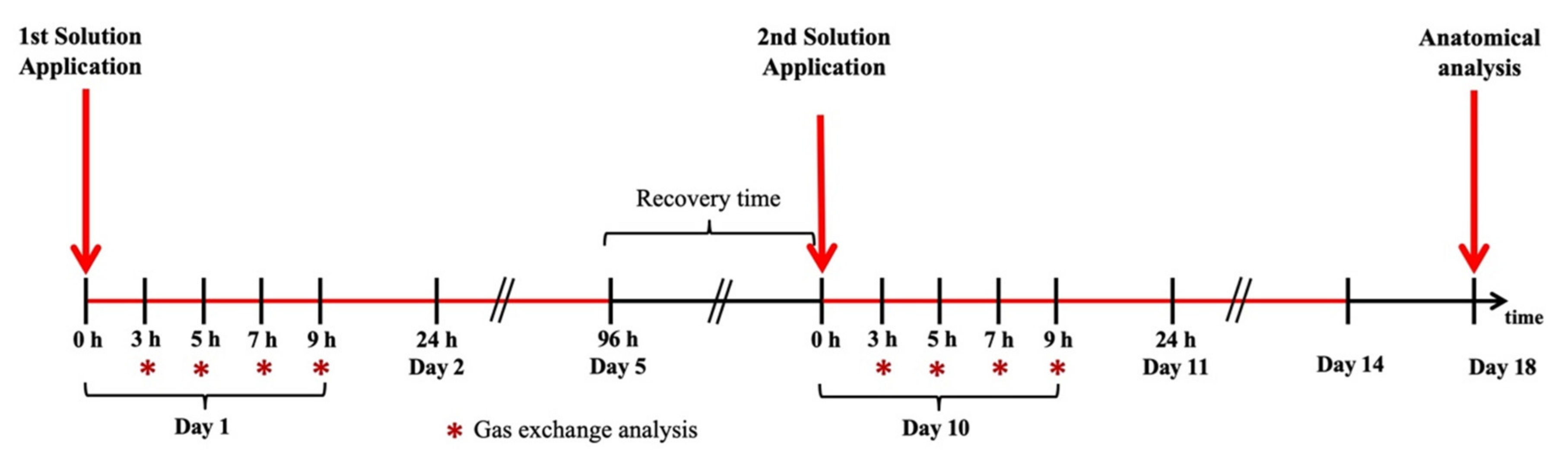
4.2. Plant–Plant Communication Experimental Design
- (1)
- Mock (M): Contact with the mock solution (without the addition of MeJa), with its root separated into two parts, where half remained in pot 1 and the second half was allocated to pot 2.
- (2)
- Mock neighbor (MN): Without contact with any solution, with its root also separated into two parts, where the first half was in pot 2, allowing direct contact with the roots of the mock group, while the second half was in pot 3.
- (3)
- Treated (T): Contact with the MeJa solution, with its root separated into two parts, where half remained in pot 1 and the second half was allocated to pot 2.
- (4)
- Treated neighbor (TN): Without contact with the MeJa solution, with its root also separated into two parts, where half was in pot 2, allowing direct contact with the roots of the treated group, while the second half was in pot 3.
4.3. Physiological Analysis
4.4. Morphological Analysis
4.5. Data analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MeJa | Methyl jasmonate |
| ROS | Reactive oxygen species |
| A | CO2 net assimilation |
| T | Treated group |
| M | Mock group |
| HAA | Hours after application |
| gS | Stomatal conductance |
| TN | Treated neighbor group |
| MN | Mock neighbor group |
| FM | Maximum fluorescence adapted to light |
| VPD | Vapor pressure deficit |
| FAA 50 | Formaldehyde, acetic acid, 50% ethyl alcohol |
| HPLC–MS/MS | High-performance liquid chromatography-mass spectrometry |
References
- Lyon, P. The biogenic approach to cognition. Cogn. Process. 2006, 7, 11–29. [Google Scholar] [CrossRef]
- Baluška, F.; Gagliano, M.; Witzany, G. Memory and Learning in Plants, 1st ed.; Baluska, F., Gagliano, M., Witzany, G., Eds.; Signaling and Communication in Plants; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-75595-3. [Google Scholar]
- Baluška, F.; Levin, M. On having no head: Cognition throughout biological systems. Front. Psychol. 2016, 7, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiler, E.W. Sensory principles of higher plants. Angew. Chem. Int. Ed. 2003, 42, 392–411. [Google Scholar] [CrossRef] [PubMed]
- Mescher, M.C.; De Moraes, C.M. Role of plant sensory perception in plant-animal interactions. J. Exp. Bot. 2015, 66, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Baluška, F.; Mancuso, S. Plant cognition and behavior: From environmental awareness to synaptic circuits navigating root apices. In Memory and Learning in Plants; Baluška, F., Gagliano, M., Witzany, G., Eds.; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-75595-3. [Google Scholar]
- Karban, R. Plant behaviour and communication. Ecol. Lett. 2008, 11, 727–739. [Google Scholar] [CrossRef]
- Kumar, A.; Memo, M.; Mastinu, A. Plant behaviour: An evolutionary response to the environment? Plant Biol. 2020, 22, 961–970. [Google Scholar] [CrossRef]
- Zimmermann, M.R.; Mithöfer, A. Long-Distance Systemic Signaling and Communication in Plants; Baluška, F., Ed.; Signaling and Communication in Plants; Springer: Berlin/Heidelberg, Germany, 2013; Volume 19, ISBN 978-3-642-36469-3. [Google Scholar]
- Landi, M. Airborne signals and abiotic factors: The neglected side of the plant communication. Commun. Integr. Biol. 2020, 13, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M. Plants talk, but are they deaf? Trends Plant Sci. 2003, 8, 403–405. [Google Scholar] [CrossRef]
- Meents, A.K.; Mithöfer, A. Plant–plant communication: Is there a role for volatile damage-associated molecular patterns? Front. Plant Sci. 2020, 11, 583275. [Google Scholar] [CrossRef]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proc. Natl. Acad. Sci. USA 2004, 101, 1781–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, J.J.; Choi, Y. Do Methyl jasmonate as a vital substance in plants. Trends Genet. 2003, 19, 409–413. [Google Scholar] [CrossRef]
- Okada, K.; Abe, H.; Arimura, G. Jasmonates induce both defense responses and communication in monocotyledonous and dicotyledonous plants. Plant Cell Physiol. 2015, 56, 16–27. [Google Scholar] [CrossRef]
- Huang, H.; Liu, B.; Liu, L.; Song, S. Jasmonate action in plant growth and development. J. Exp. Bot. 2017, 68, 1349–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attaran, E.; Major, I.T.; Cruz, J.A.; Rosa, B.A.; Koo, A.J.K.; Chen, J.; Kramer, D.M.; He, S.Y.; Howe, G.A. Temporal dynamics of growth and photosynthesis suppression in response to jasmonate signaling. Plant Physiol. 2014, 165, 1302–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Ye, J.; Rasulov, B.; Niinemets, Ü. Role of stomatal conductance in modifying the dose response of stress-volatile emissions in methyl jasmonate treated leaves of cucumber (Cucumis Sativa). Int. J. Mol. Sci. 2020, 21, 1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Carvalhais, L.C.; Schenk, P.M.; Dennis, P.G. Effects of jasmonic acid signalling on the wheat microbiome differ between body sites. Sci. Rep. 2017, 7, 41766. [Google Scholar] [CrossRef] [PubMed]
- Rakwal, R.; Komatsu, S. Jasmonic acid-induced necrosis and drastic decreases in ribulose-1,5-bisphosphate carboxylase/oxygenase in rice seedlings under light involves reactive oxygen species. J. Plant Physiol. 2001, 158, 679–688. [Google Scholar] [CrossRef]
- Zhang, L.; Xing, D. Methyl jasmonate induces production of reactive oxygen species and alterations in mitochondrial dynamics that precede photosynthetic dysfunction and subsequent cell death. Plant Cell Physiol. 2008, 49, 1092–1111. [Google Scholar] [CrossRef] [Green Version]
- Howe, G.A.; Major, I.T.; Koo, A.J. Modularity in jasmonate signaling for multistress resilience. Annu. Rev. Plant Biol. 2018, 69. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Bae, D.W.; Ryu, C.M. Aboveground whitefly infestation modulates transcriptional levels of anthocyanin biosynthesis and jasmonic acid signaling-related genes and augments the cope with drought stress of maize. PLoS ONE 2015, 10, e0143879. [Google Scholar] [CrossRef] [Green Version]
- Elhakeem, A.; Markovic, D.; Broberg, A.; Anten, N.P.R.; Ninkovic, V. Aboveground mechanical stimuli affect belowground plant-plant communication. PLoS ONE 2018, 13, e0195646. [Google Scholar] [CrossRef] [Green Version]
- Song, G.C.; Sim, H.J.; Kim, S.G.; Ryu, C.M. Root-mediated signal transmission of systemic acquired resistance against above-ground and below-ground pathogens. Ann. Bot. 2016, 118, 821–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Hu, C.; Zhou, J.; Liu, Y.; Cai, J.; Pan, C.; Wang, Y.; Wu, X.; Shi, K.; Xia, X.; et al. Systemic root-shoot signaling drives jasmonate-based root defense against nematodes. Curr. Biol. 2019, 29, 3430–3438.e4. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Sugimoto, K.; Ramadan, A.; Arimura, G.I. Memory of plant communications for priming anti-herbivore responses. Sci. Rep. 2013, 3, srep01872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagliano, M.; Vyazovskiy, V.V.; Borbély, A.A.; Grimonprez, M.; Depczynski, M. Learning by association in plants. Sci. Rep. 2016, 6, 38427. [Google Scholar] [CrossRef] [Green Version]
- Song, G.C.; Ryu, C.M. Evidence for volatile memory in plants: Boosting defence priming through the recurrent application of plant volatiles. Mol. Cells 2018, 41, 724–732. [Google Scholar] [CrossRef]
- Walter, J.; Jentsch, A.; Beierkuhnlein, C.; Kreyling, J. Ecological stress memory and cross stress tolerance in plants in the face of climate extremes. Environ. Exp. Bot. 2013, 94, 3–8. [Google Scholar] [CrossRef]
- Kosová, K.; Vitámvás, P.; Prásil, I.T.; Renault, J. Plant proteome changes under abiotic stress—Contribution of proteomics studies to understanding plant stress response. J. Proteom. 2011, 74, 1301–1322. [Google Scholar] [CrossRef]
- Crisp, P.A.; Ganguly, D.; Eichten, S.R.; Borevitz, J.O.; Pogson, B.J. Reconsidering plant memory: Intersections between stress recovery, RNA turnover, and epigenetics. Sci. Adv. 2016, 2, e1501340. [Google Scholar] [CrossRef] [Green Version]
- Gagliano, M.; Renton, M.; Depczynski, M.; Mancuso, S. Experience teaches plants to learn faster and forget slower in environments where it matters. Oecologia 2014, 175, 63–72. [Google Scholar] [CrossRef]
- Toledo, G.R.A. Transmissão de Sinal de Falta de Água Entre Plantas: Efeitos Sobre a Temperatura e a Pressão de Turgor Foliar. Master’s Thesis, Western São Paulo University, Presidente Prudente, Brazil, 2015. [Google Scholar]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates: Sunderland, MA, USA, 2015; ISBN 978-85-8271-366-2. [Google Scholar]
- Aliche, E.B.; Prusova-Bourke, A.; Ruiz-Sanchez, M.; Oortwijn, M.; Gerkema, E.; Van As, H.; Visser, R.G.F.; van der Linden, C.G. Morphological and physiological responses of the potato stem transport tissues to dehydration stress. Planta 2020, 251, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witzany, G. Memory and learning as key competences of living organisms. In Memory and Learning in Plants; Baluska, F., Gagliano, M., Witzany, G., Eds.; Springer International Publishing: Cham, Switzerland, 2018; p. 223. [Google Scholar]
- Crespo-Salvador, Ó.; Escamilla-Aguilar, M.; López-Cruz, J.; López-Rodas, G.; González-Bosch, C. Determination of histone epigenetic marks in Arabidopsis and tomato genes in the early response to Botrytis cinerea. Plant Cell Rep. 2018, 37, 153–166. [Google Scholar] [CrossRef]
- Walter, J.; Nagy, L.; Hein, R.; Rascher, U.; Beierkuhnlein, C.; Willner, E.; Jentsch, A. Do plants remember drought? Hints towards a drought-memory in grasses. Environ. Exp. Bot. 2011, 71, 34–40. [Google Scholar] [CrossRef]
- Valladares, F.; Wright, S.J.; Lasso, E.; Kitajima, K.; Pearcy, R.W. Plastic phenotypic response to light of 16 congeneric shrubs from a panamanian rainforest. Ecology 2000, 81, 1925–1936. [Google Scholar] [CrossRef]
- Suhita, D.; Raghavendra, A.S.; Kwak, J.M.; Vavasseur, A. Cytoplasmic alkalization precedes reactive oxygen species production during methyl jasmonate- and abscisic acid- induced stomatal closure. Plant Physiol. 2004, 134, 1536–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyystjärvi, E. Photoinhibition of Photosystem II. In International Review of Cell and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 300, pp. 243–303. ISBN 9780124052109. [Google Scholar]
- Costa, A.C.; Rezende-Silva, S.L.; Megguer, C.A.; Moura, L.M.F.; Rosa, M.; Silva, A.A. The effect of irradiance and water restriction on photosynthesis in young jatoba-do-cerrado (Hymenaea stigonocarpa) plants. Photosynthetica 2015, 53, 118–127. [Google Scholar] [CrossRef]
- Franco, A.C.; Matsubara, S.; Orthen, B. Photoinhibition, carotenoid composition and the co-regulation of photochemical and non-photochemical quenching in neotropical savanna trees. Tree Physiol. 2007, 27, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R.; Oxborough, K.; Lawson, T.; Morison, J.I.L. High resolution imaging of photosynthetic activities of tissues, cells and chloroplasts in leaves. J. Exp. Bot. 2001, 52, 615–621. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Fu, B.; Kang, H.; Song, Z.; Jia, M.; Cao, D.; Wei, A. Response of gas-exchange characteristics and chlorophyll fluorescence to acute sulfur dioxide exposure in landscape plants. Ecotoxicol. Environ. Saf. 2019, 171, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Gasperini, D.; Chételat, A.; Acosta, I.F.; Goossens, J.; Pauwels, L.; Goossens, A.; Dreos, R.; Alfonso, E.; Farmer, E.E. Multilayered organization of jasmonate signalling in the regulation of root growth. PLoS Genet. 2015, 11, e1005300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, A.; Zimmer, M.; Mielke, S.; Stellmach, H.; Melnyk, C.W.; Hause, B.; Gasperini, D. Wound-induced shoot-to-root relocation of JA-Ile precursors coordinates arabidopsis growth. Mol. Plant 2019, 12, 1383–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Moerkercke, A.; Duncan, O.; Zander, M.; Šimura, J.; Broda, M.; Van den Bossche, R.; Lewsey, M.G.; Lama, S.; Singh, K.B.; Ljung, K.; et al. A MYC2/MYC3/MYC4-dependent transcription factor network regulates water spray-responsive gene expression and jasmonate levels. Proc. Natl. Acad. Sci. USA 2019, 116, 23345–23356. [Google Scholar] [CrossRef]
- Herbert, G. Beitrage zur Anatomie adulter Gramineenwurzeln im Hinblick auf taxonomische Verwendbarkeit. Beitrage zur Biol. der Pflanz. 1977, 53, 217–307. [Google Scholar]
- Ort, D.R.; Baker, N.R. A photoprotective role for O2 as an alternative electron sink in photosynthesis? Curr. Opin. Plant Biol. 2002, 5, 193–198. [Google Scholar] [CrossRef]
- Wilson, A.J.; Robards, A.W.; Goss, M.J. Effects of mechanical impedance on root growth in barley, Hordeum vulgare L. J. Exp. Bot. 1977, 28, 1216–1227. [Google Scholar] [CrossRef]
- Atwell, B.J. Physiological responses of lupin roots to soil compaction. In Structural and Functional Aspects of Transport in Roots; Springer: Berlin/Heidelberg, Germany, 1988; pp. 277–281. [Google Scholar]
- Colombi, T.; Kirchgessner, N.; Walter, A.; Keller, T. Root tip shape governs root elongation rate under increased soil strength. Plant Physiol. 2017, 174, 2289–2301. [Google Scholar] [CrossRef] [Green Version]
- Potocka, I.; Szymanowska-Pulka, J. Morphological responses of plant roots to mechanical stress. Ann. Bot. 2018, 122, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.S.; Kim, Y.S.; Redillas, M.C.F.R.; Jang, G.; Jung, H.; Bang, S.W.; Choi, Y.D.; Ha, S.-H.; Reuzeau, C.; Kim, J.-K. OsNAC5 overexpression enlarges root diameter in rice plants leading to enhanced drought tolerance and increased grain yield in the field. Plant Biotechnol. J. 2013, 11, 101–114. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Hsu, P.-K.; Tsay, Y.-F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Regulation of root water uptake under abiotic stress conditions. J. Exp. Bot. 2012, 63, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Franco, D.M. Comunicação Radicular Induzida por Diferentes Tipos de Substâncias Químicas. Ph.D. Thesis, São Paulo State University, Botucatu, Brazil, 2017. [Google Scholar]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill: London, UK, 1940. [Google Scholar]
- Gerrits, P.O.; Horobin, R.W. The Application of Glycol Methacrylate in Histotechnology; Some Fundamental Principles; Faculteit der Geneeskunde, Rijksuniversiteit Groningen: Groningen, The Netherlands, 1991. [Google Scholar]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- Markovic, D.; Colzi, I.; Taiti, C.; Ray, S.; Scalone, R.; Ali, J.G.; Mancuso, S.; Ninkovic, V. Airborne signals synchronize the defenses of neighboring plants in response to touch. J. Exp. Bot. 2019, 70, 691–700. [Google Scholar] [CrossRef]
- Ballaré, C.L. Jasmonate-induced defenses: A tale of intelligence, collaborators and rascals. Trends Plant Sci. 2011, 16, 249–257. [Google Scholar] [CrossRef]
- Kriegs, B.; Jansen, M.; Hahn, K.; Peisker, H.; Šamajová, O.; Beck, M.; Braun, S.; Ulbrich, A.; Baluška, F.; Schulz, M. Cyclic monoterpene mediated modulations of Arabidopsis thaliana phenotype. Plant Signal. Behav. 2010, 5, 832–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlovič, A.; Libiaková, M.; Bokor, B.; Jakšová, J.; Petřík, I.; Novák, O.; Baluška, F. Anaesthesia with diethyl ether impairs jasmonate signalling in the carnivorous plant Venus flytrap (Dionaea muscipula). Ann. Bot. 2019, 5, 1–31. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamashita, F.; Rodrigues, A.L.; Rodrigues, T.M.; Palermo, F.H.; Baluška, F.; Almeida, L.F.R.d. Potential Plant–Plant Communication Induced by Infochemical Methyl Jasmonate in Sorghum (Sorghum bicolor). Plants 2021, 10, 485. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030485
Yamashita F, Rodrigues AL, Rodrigues TM, Palermo FH, Baluška F, Almeida LFRd. Potential Plant–Plant Communication Induced by Infochemical Methyl Jasmonate in Sorghum (Sorghum bicolor). Plants. 2021; 10(3):485. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030485
Chicago/Turabian StyleYamashita, Felipe, Angélica Lino Rodrigues, Tatiane Maria Rodrigues, Fernanda Helena Palermo, František Baluška, and Luiz Fernando Rolim de Almeida. 2021. "Potential Plant–Plant Communication Induced by Infochemical Methyl Jasmonate in Sorghum (Sorghum bicolor)" Plants 10, no. 3: 485. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030485