
Comparison of Genomic Prediction Methods for Yellow, Stem, and Leaf Rust Resistance in Wheat Landraces from Afghanistan

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
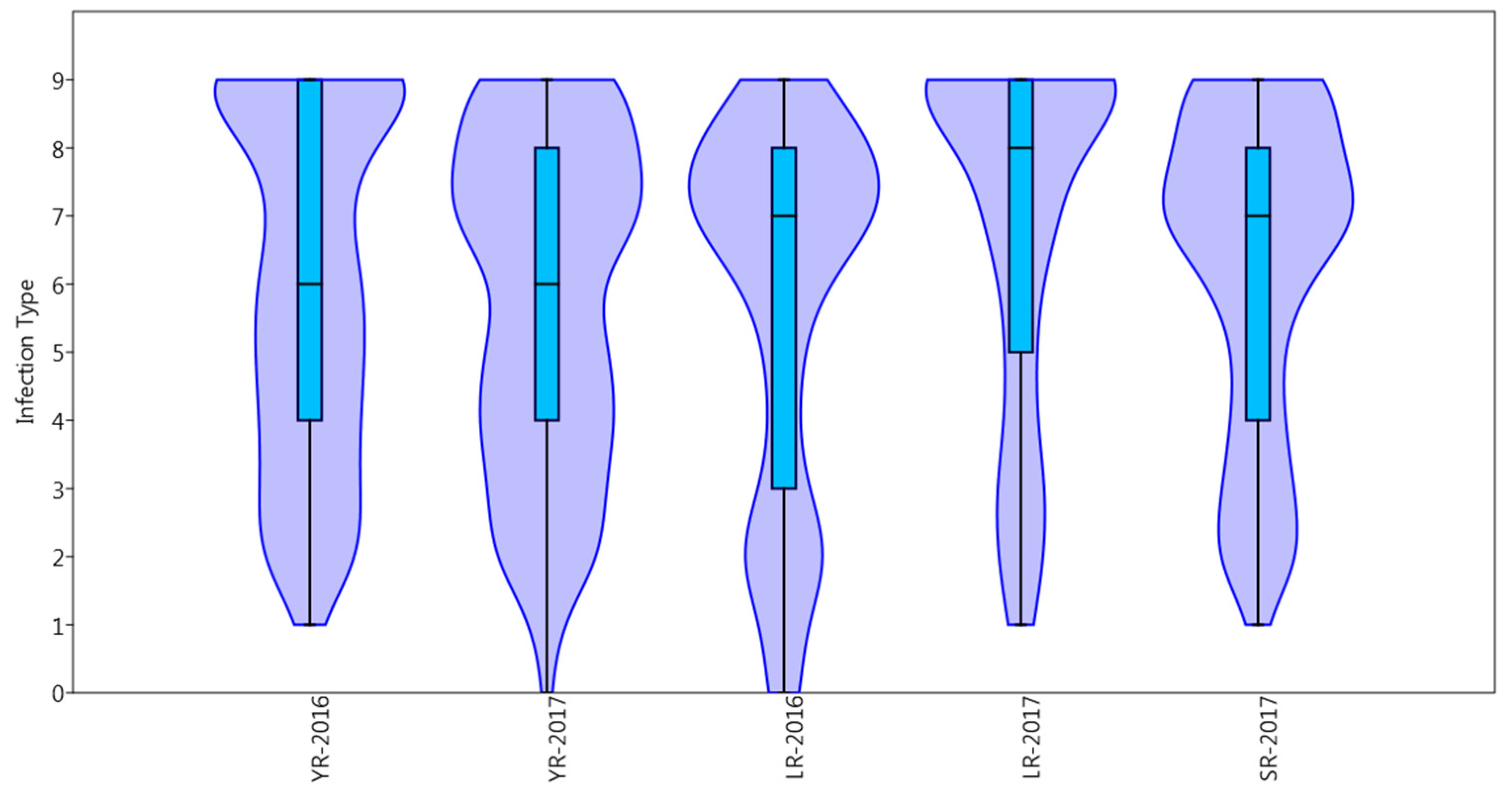
2.1. Phenotypic Data and Heritability
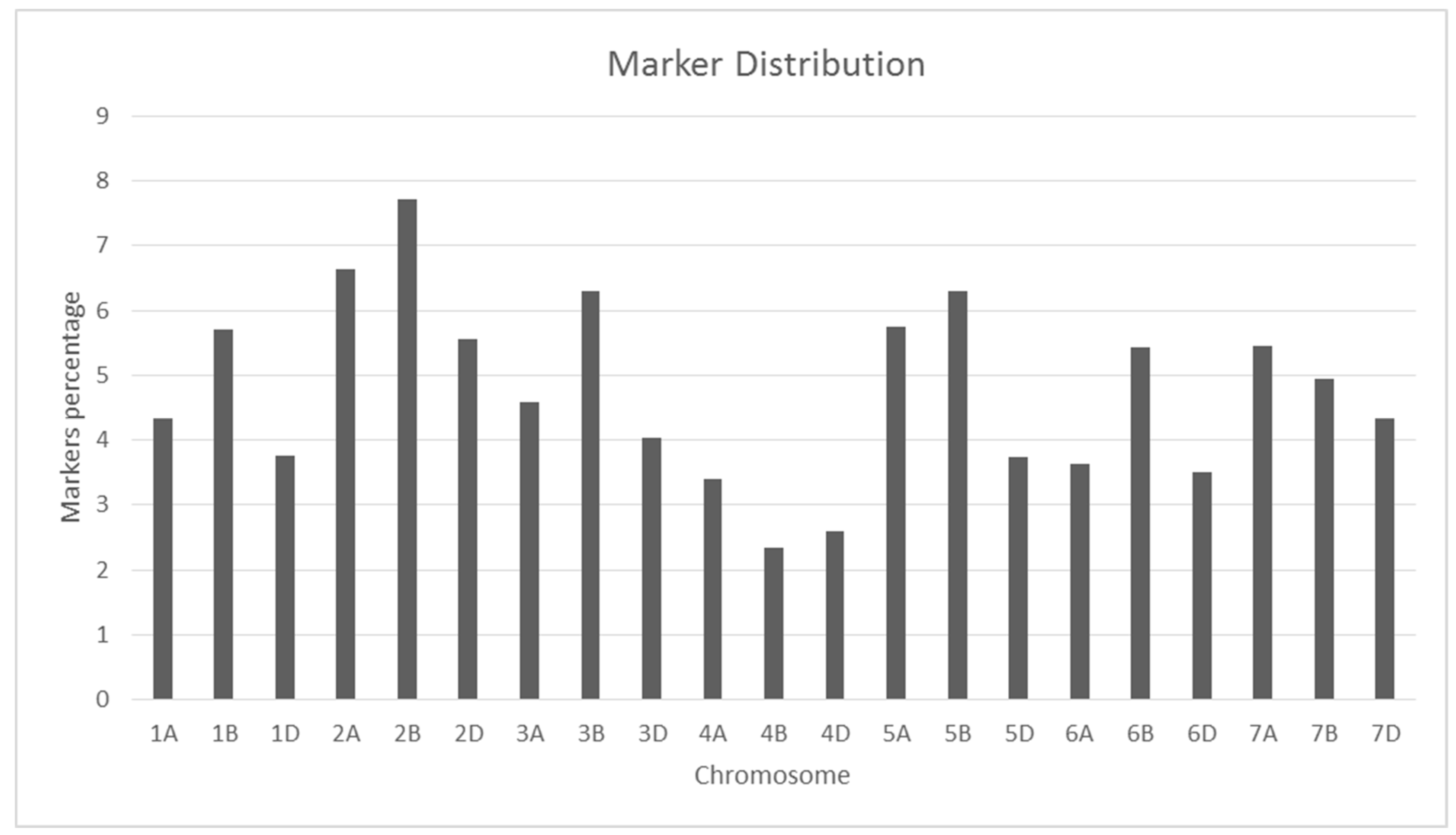
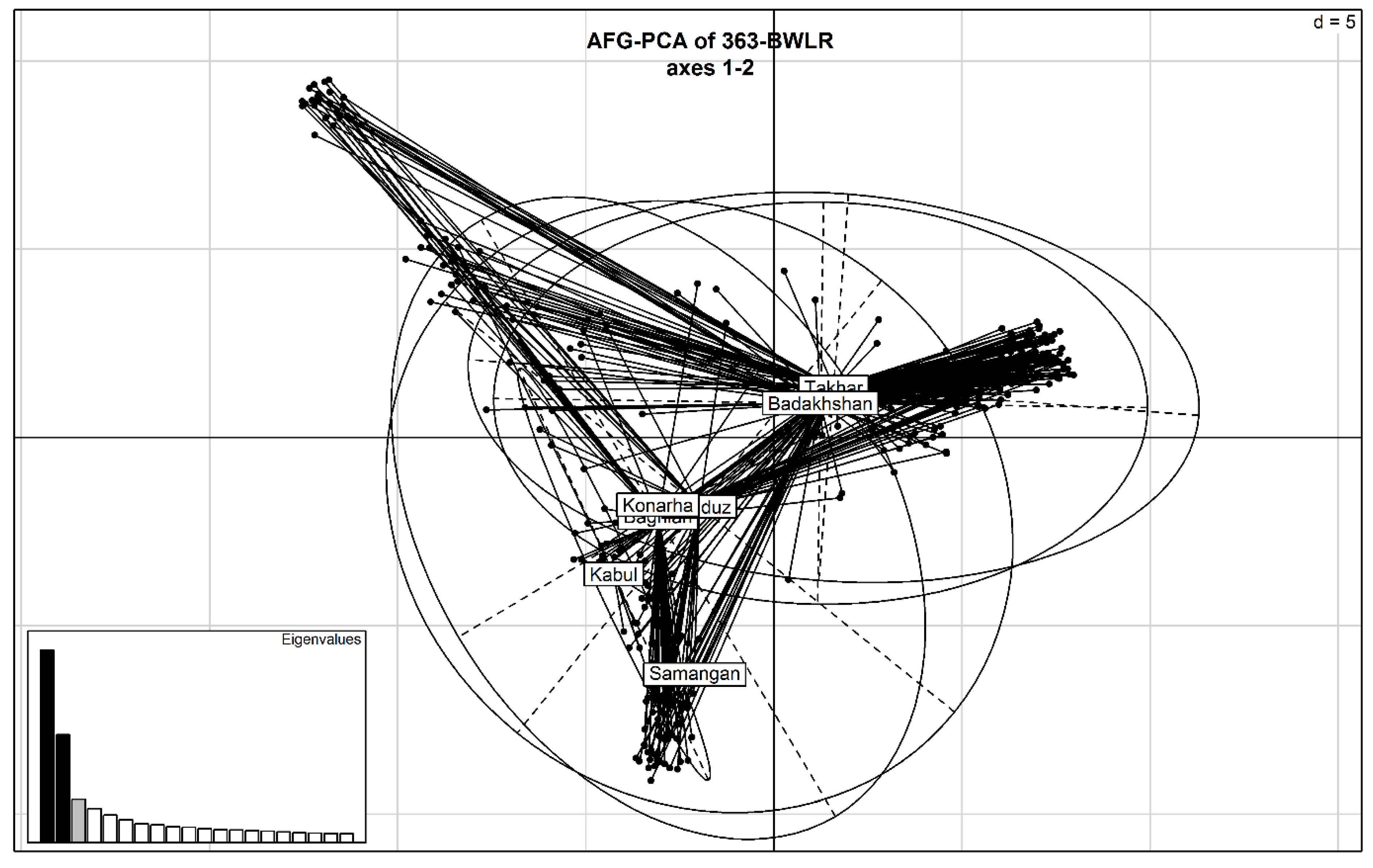
2.2. Marker Distribution and Genetic Relationship Matrix
2.3. Genomic Predictive Abilities of the Nine Methods for Yellow Rust
2.4. Predictive Abilities of the Different Methods for Leaf Rust
2.5. Comparison between the Models
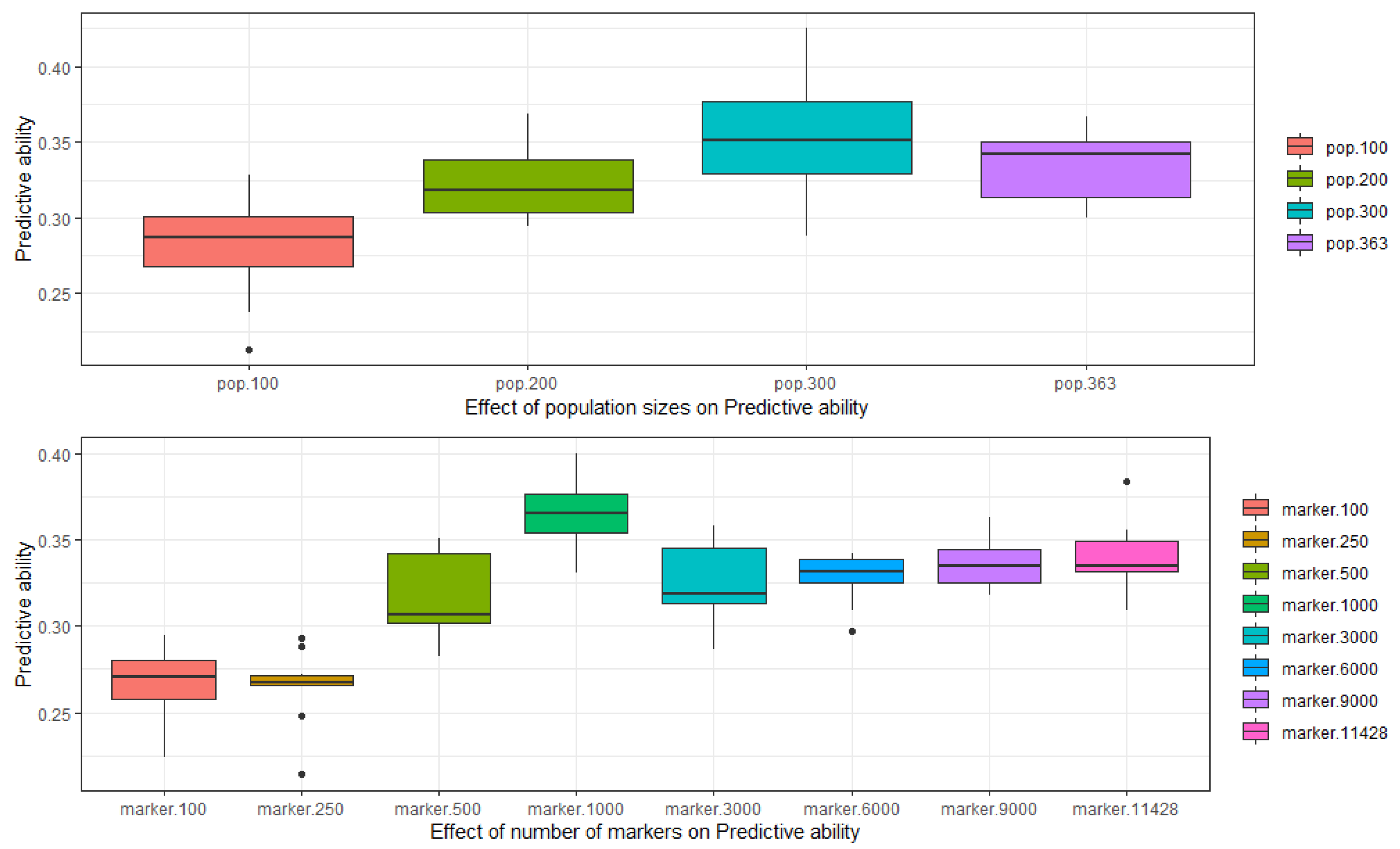
2.6. Simulation Analysis
3. Discussion
4. Materials and Methods
4.1. Adult Plant Evaluation and Phenotypic Data
4.1.1. Adult Plant Resistance for Yellow Rust
4.1.2. Adult Plant Resistance for Leaf Rust
4.1.3. Adult Plant Resistance to Stem Rust
4.2. Correlation, Heritability, and Relationship Matrix
4.3. Genotyping
4.4. Genomic Prediction Methods
4.4.1. GBLUP and RR-BLUP
4.4.2. LASSO and Elastic Net
4.4.3. Bayesian Models
Bayesian Ridge Regression
Bayes A
Bayes B
Bayes C
4.4.4. Reproducing Kernel Hilbert Spaces (RKHS)
4.5. Simulation Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rajaram, S. Challenges in Wheat Research and Development. In The International Dimension of the American Society of Agronomy: Past and Future; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 39–47. [Google Scholar]
- Tehseen, M.M.; Tonk, F.A.; Tosun, M.; Amri, A.; Sansaloni, C.P.; Kurtulus, E.; Yazbek, M.; Al-Sham’aa, K.; Ozseven, I.; Safdar, L.B.; et al. Genome-Wide Association Study of Resistance to PstS2 and Warrior Races of Puccinia Striiformis f. Sp. Tritici (Stripe Rust) in Bread Wheat Landraces. Plant Genome 2021, 14, e20066. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Chen, X.; Jin, Y. Diseases Which Challenge Global Wheat Production–The Cereal Rusts; Wiley Press: Hoboken, NJ, USA, 2009. [Google Scholar]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, J.W.; Morris, C.F.; Appels, R.; Xia, X.C. Catalogue of Gene Symbols for Wheat: 2013-14 Supplement; Annual Wheat Newsletter; GrainGenes: Yokohama, Japan, 2014; Volume 58. [Google Scholar]
- Ali, S.; Gladieux, P.; Leconte, M.; Gautier, A.; Justesen, A.F.; Hovmøller, M.S.; Enjalbert, J.; de Vallavieille-Pope, C. Origin, Migration Routes and Worldwide Population Genetic Structure of the Wheat Yellow Rust Pathogen Puccinia Striiformis f.Sp. Tritici. Plos Pathog 2014, 10, e1003903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figlan, S.; Ntushelo, K.; Mwadzingeni, L.; Terefe, T.; Tsilo, T.J.; Shimelis, H. Breeding Wheat for Durable Leaf Rust Resistance in Southern Africa: Variability, Distribution, Current Control Strategies, Challenges and Future Prospects. Front. Plant Sci. 2020, 11, 549. [Google Scholar] [CrossRef] [PubMed]
- Zeven, A.C. Landraces: A Review of Definitions and Classifications. Euphytica 1998, 104, 127–139. [Google Scholar] [CrossRef]
- Daetwyler, H.D.; Bansal, U.K.; Bariana, H.S.; Hayden, M.J.; Hayes, B.J. Genomic Prediction for Rust Resistance in Diverse Wheat Landraces. Appl. Genet. 2014, 127, 1795–1803. [Google Scholar] [CrossRef]
- Gorjanc, G.; Jenko, J.; Hearne, S.J.; Hickey, J.M. Initiating Maize Pre-Breeding Programs Using Genomic Selection to Harness Polygenic Variation from Landrace Populations. BMC Genom. 2016, 17, 30. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Morris, C.; Xia, X.C. Catalogue of Gene Symbols for Wheat: 2017 Supplement; GrainGenes: Albany, CA, USA, 2010; Volume 20. [Google Scholar]
- Mallard, S.; Gaudet, D.; Aldeia, A.; Abelard, C.; Besnard, A.-L.; Sourdille, P.; Dedryver, F. Genetic Analysis of Durable Resistance to Yellow Rust in Bread Wheat. Appl. Genet. 2005, 110, 1401–1409. [Google Scholar] [CrossRef]
- Ornella, L.; Singh, S.; Perez, P.; Burgueño, J.; Singh, R.; Tapia, E.; Bhavani, S.; Dreisigacker, S.; Braun, H.-J.; Mathews, K. Genomic Prediction of Genetic Values for Resistance to Wheat Rusts. Plant Genome 2012, 5, 136–148. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, A.J.; Smith, K.P.; Jannink, J.-L. Potential and Optimization of Genomic Selection for Fusarium Head Blight Resistance in Six-Row Barley. Crop Sci. 2012, 52, 1609–1621. [Google Scholar] [CrossRef]
- Bassi, F.M.; Bentley, A.R.; Charmet, G.; Ortiz, R.; Crossa, J. Breeding Schemes for the Implementation of Genomic Selection in Wheat (Triticum Spp.). Plant Sci. 2016, 242, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Juliana, P.; Singh, R.P.; Singh, P.K.; Crossa, J.; Rutkoski, J.E.; Poland, J.A.; Bergstrom, G.C.; Sorrells, M.E. Comparison of Models and Whole-Genome Profiling Approaches for Genomic-Enabled Prediction of Septoria Tritici Blotch, Stagonospora Nodorum Blotch, and Tan Spot Resistance in Wheat. Plant Genome 2017, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crossa, J.; de Los Campos, G.; Pérez, P.; Gianola, D.; Burgueño, J.; Araus, J.L.; Makumbi, D.; Singh, R.P.; Dreisigacker, S.; Yan, J. Prediction of Genetic Values of Quantitative Traits in Plant Breeding Using Pedigree and Molecular Markers. Genetics 2010, 186, 713–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heffner, E.L.; Sorrells, M.E.; Jannink, J.-L. Genomic Selection for Crop Improvement. Crop Sci. 2009, 49, 1–12. [Google Scholar] [CrossRef]
- Heffner, E.L.; Jannink, J.-L.; Sorrells, M.E. Genomic Selection Accuracy Using Multifamily Prediction Models in a Wheat Breeding Program. Plant Genome 2011, 4, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Crossa, J.; Jarquín, D.; Franco, J.; Pérez-Rodríguez, P.; Burgueño, J.; Saint-Pierre, C.; Vikram, P.; Sansaloni, C.; Petroli, C.; Akdemir, D. Genomic Prediction of Gene Bank Wheat Landraces. G3 Genes Genomes Genet. 2016, 6, 1819–1834. [Google Scholar] [CrossRef] [Green Version]
- Manickavelu, A.; Hattori, T.; Yamaoka, S.; Yoshimura, K.; Kondou, Y.; Onogi, A.; Matsui, M.; Iwata, H.; Ban, T. Genetic Nature of Elemental Contents in Wheat Grains and Its Genomic Prediction: Toward the Effective Use of Wheat Landraces from Afghanistan. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Rutkoski, J.E.; Poland, J.A.; Singh, R.P.; Huerta-Espino, J.; Bhavani, S.; Barbier, H.; Rouse, M.N.; Jannink, J.-L.; Sorrells, M.E. Genomic Selection for Quantitative Adult Plant Stem Rust Resistance in Wheat. Plant Genome 2014, 7. [Google Scholar] [CrossRef] [Green Version]
- Lantican, M.A.; Dubin, H.J.; Morris, M.L. Impacts of International Wheat Breeding Research in the Developing World, 1988–2002; Cimmyt: near Texcoco, Mexico, 2005. [Google Scholar]
- Muqaddasi, Q.H.; Zhao, Y.; Rodemann, B.; Plieske, J.; Ganal, M.W.; Röder, M.S. Genome-Wide Association Mapping and Prediction of Adult Stage Septoria Tritici Blotch Infection in European Winter Wheat via High-Density Marker Arrays. Plant Genome 2019, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, S.A.; Hickey, J.M.; Van der Werf, J.H. Different Models of Genetic Variation and Their Effect on Genomic Evaluation. Genet. Sel. Evol. 2011, 43, 18. [Google Scholar] [CrossRef] [Green Version]
- Onogi, A.; Ideta, O.; Inoshita, Y.; Ebana, K.; Yoshioka, T.; Yamasaki, M.; Iwata, H. Exploring the Areas of Applicability of Whole-Genome Prediction Methods for Asian Rice (Oryza Sativa L.). Theor. Appl. Genet. 2015, 128, 41–53. [Google Scholar] [CrossRef]
- Moser, G.; Tier, B.; Crump, R.E.; Khatkar, M.S.; Raadsma, H.W. A Comparison of Five Methods to Predict Genomic Breeding Values of Dairy Bulls from Genome-Wide SNP Markers. Genet. Sel. Evol. 2009, 41, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asoro, F.G.; Newell, M.A.; Beavis, W.D.; Scott, M.P.; Jannink, J.-L. Accuracy and Training Population Design for Genomic Selection on Quantitative Traits in Elite North American Oats. Plant Genome 2011, 4, 132–144. [Google Scholar] [CrossRef] [Green Version]
- Habier, D.; Fernando, R.L.; Kizilkaya, K.; Garrick, D.J. Extension of the Bayesian Alphabet for Genomic Selection. BMC Bioinform. 2011, 12, 186. [Google Scholar] [CrossRef] [Green Version]
- Resende, M.F.; Muñoz, P.; Resende, M.D.; Garrick, D.J.; Fernando, R.L.; Davis, J.M.; Jokela, E.J.; Martin, T.A.; Peter, G.F.; Kirst, M. Accuracy of Genomic Selection Methods in a Standard Data Set of Loblolly Pine (Pinus Taeda L.). Genetics 2012, 190, 1503–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of Total Genetic Value Using Genome-Wide Dense Marker Maps. Genetics 2001, 157, 1819–1829. [Google Scholar]
- Daetwyler, H.D.; Pong-Wong, R.; Villanueva, B.; Woolliams, J.A. The Impact of Genetic Architecture on Genome-Wide Evaluation Methods. Genetics 2010, 185, 1021–1031. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Dekkers, J.C.; Fernando, R.L.; Jannink, J.-L. Factors Affecting Accuracy from Genomic Selection in Populations Derived from Multiple Inbred Lines: A Barley Case Study. Genetics 2009, 182, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Howard, R.; Carriquiry, A.L.; Beavis, W.D. Parametric and Nonparametric Statistical Methods for Genomic Selection of Traits with Additive and Epistatic Genetic Architectures. G3: GenesGenomesGenet. 2014, 4, 1027–1046. [Google Scholar] [CrossRef] [Green Version]
- Crossa, J.; Beyene, Y.; Kassa, S.; Pérez, P.; Hickey, J.M.; Chen, C.; de los Campos, G.; Burgueño, J.; Windhausen, V.S.; Buckler, E. Genomic Prediction in Maize Breeding Populations with Genotyping-by-Sequencing. G3: GenesGenomesGenet. 2013, 3, 1903–1926. [Google Scholar] [CrossRef] [Green Version]
- Pérez, P.; de los Campos, G.; Crossa, J.; Gianola, D. Genomic-Enabled Prediction Based on Molecular Markers and Pedigree Using the Bayesian Linear Regression Package in R. Plant Genome 2010, 3, 106–116. [Google Scholar] [CrossRef]
- Onogi, A.; Watanabe, M.; Mochizuki, T.; Hayashi, T.; Nakagawa, H.; Hasegawa, T.; Iwata, H. Toward Integration of Genomic Selection with Crop Modelling: The Development of an Integrated Approach to Predicting Rice Heading Dates. Theor. Appl. Genet. 2016, 129, 805–817. [Google Scholar] [CrossRef]
- Jia, Y.; Jannink, J.-L. Multiple-Trait Genomic Selection Methods Increase Genetic Value Prediction Accuracy. Genetics 2012, 192, 1513–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Zhao, Y.; Rodemann, B.; Plieske, J.; Kollers, S.; Korzun, V.; Ebmeyer, E.; Argillier, O.; Hinze, M.; Ling, J. Potential and Limits to Unravel the Genetic Architecture and Predict the Variation of Fusarium Head Blight Resistance in European Winter Wheat (Triticum Aestivum L.). Heredity 2015, 114, 318–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, R.F.; Campbell, A.B.; Hannah, A.E. A Diagrammatic Scale for Estimating Rust Intensity on Leaves and Stems of Cereals. Can. J. Res. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Line, R.F.; Qayoum, A. Virulence, Aggressiveness, Evolution, and Distribution of Races of Puccinia Striiformis (the Cause of Stripe Rust of Wheat) in North America, 1968-87; Technical bulletin-United States Department of Agriculture: Washington, DC, USA, 1992. [Google Scholar]
- Jin, Y.; Szabo, L.J.; Pretorius, Z.A.; Singh, R.P.; Ward, R.; Fetch, T., Jr. Detection of Virulence to Resistance Gene Sr24 within Race TTKS of Puccinia Graminis f. Sp. Tritici. Plant Dis. 2008, 92, 923–926. [Google Scholar] [CrossRef] [Green Version]
- Luu, K.; Bazin, E.; Blum, M.G. Pcadapt: An R Package to Perform Genome Scans for Selection Based on Principal Component Analysis. Mol. Ecol. Resour. 2017, 17, 67–77. [Google Scholar] [CrossRef]
- Sansaloni, C.; Petroli, C.; Jaccoud, D.; Carling, J.; Detering, F.; Grattapaglia, D.; Kilian, A. Diversity Arrays Technology (DArT) and next-Generation Sequencing Combined: Genome-Wide, High Throughput, Highly Informative Genotyping for Molecular Breeding of Eucalyptus. In Proceedings of the IUFRO Tree Biotechnology Conference 2011: From Genomes to Integration and Delivery, Arraial d’Ajuda, Bahia, Brazil, 26 June–2 July 2011; 2011; Volume 5, p. 54. [Google Scholar]
- Charmet, G.; Tran, L.-G.; Auzanneau, J.; Rincent, R.; Bouchet, S. BWGS: AR Package for Genomic Selection and Its Application to a Wheat Breeding Programme. PLoS ONE 2020, 15, e0222733. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, R. Prediction of Maize Single-Cross Performance Using RFLPs and Information from Related Hybrids. Crop Sci. 1994, 34, 20–25. [Google Scholar] [CrossRef]
- Habier, D.; Fernando, R.L.; Garrick, D.J. Genomic BLUP Decoded: A Look into the Black Box of Genomic Prediction. Genetics 2013, 194, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.; Hastie, T. Regularization and Variable Selection via the Elastic Net. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2005, 67, 301–320. [Google Scholar] [CrossRef] [Green Version]
- Gianola, D. Priors in Whole-Genome Regression: The Bayesian Alphabet Returns. Genetics 2013, 194, 573–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, P.; de Los Campos, G. Genome-Wide Regression and Prediction with the BGLR Statistical Package. Genetics 2014, 198, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, A.J.; Chao, S.; Asoro, F.G.; Heffner, E.L.; Hayashi, T.; Iwata, H. Advances in Agronomy; Elseiver: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Gianola, D.; Van Kaam, J.B. Reproducing Kernel Hilbert Spaces Regression Methods for Genomic Assisted Prediction of Quantitative Traits. Genetics 2008, 178, 2289–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease-Year | Rust Infection Types | Heritability | |||
|---|---|---|---|---|---|
| R (%) | MR (%) | MS (%) | S (%) | ||
| YR-2016 | 12.4 | 19.5 | 22.0 | 46.0 | 0.97 |
| YR-2017 | 13.0 | 21.9 | 18.2 | 46.9 | 0.97 |
| LR-2016 | 21.5 | 7.2 | 13.1 | 58.0 | 0.98 |
| LR-2017 | 9.0 | 11.4 | 8.1 | 71.2 | 0.97 |
| SR-2017 | 10.1 | 13.5 | 14.7 | 61.5 | 0.97 |
| YR-2016 | YR-2017 | LR-2016 | LR-2017 | SR-2017 | |
|---|---|---|---|---|---|
| YR-2016 | 0.70* | −0.22 | −0.26 * | −0.09 | |
| YR-2017 | 0.70 * | −0.26 * | −0.11 | −0.14 | |
| LR-2016 | −0.22 | −0.26* | 0.41 * | 0.48 * | |
| LR-2017 | −0.26 * | −0.11 | 0.41 * | 0.51 * | |
| SR-2017 | −0.09 | −0.14 | 0.48 * | 0.51 * |
| Model | Disease-Year of Field Experiments | ||||
|---|---|---|---|---|---|
| YR-2016 | YR-2017 | LR-2016 | LR-2017 | SR-2017 | |
| GBLUP | 0.32 ± 0.02 | 0.30 ± 0.01 | 0.38 ± 0.01 | −0.003 ± 0.05 | 0.30 ± 0.01 |
| RR | 0.32 ± 0.01 | 0.30 ± 0.01 | 0.38 ± 0.01 | 0.03 ± 0.04 | 0.302 ± 0.02 |
| LASSO | 0.31 ± 0.03 | 0.26 ± 0.02 | 0.36 ± 0.02 | −0.03 ± 0.05 | 0.33 ± 0.02 |
| EN | 0.31 ± 0.02 | 0.28 ± 0.02 | 0.36 ± 0.02 | −0.04 ± 0.05 | 0.33 ± 0.02 |
| BRR | 0.32 ± 0.02 | 0.30 ± 0.01 | 0.38 ± 0.01 | 0.04 ± 0.04 | 0.31 ± 0.02 |
| BA | 0.32 ± 0.02 | 0.30 ± 0.01 | 0.37 ± 0.01 | 0.09 ± 0.04 | 0.30 ± 0.02 |
| BB | 0.32 ± 0.02 | 0.30 ± 0.01 | 0.38 ± 0.01 | 0.04 ± 0.04 | 0.31 ± 0.02 |
| BC | 0.32 ± 0.01 | 0.30 ± 0.02 | 0.37 ± 0.01 | 0.04 ± 0.03 | 0.30 ± 0.02 |
| RKHS | 0.33 ± 0.01 | 0.29 ± 0.01 | 0.37 ± 0.01 | 0.05 ± 0.03 | 0.29 ± 0.01 |
| GBLUP | RR | LASSO | EN | BRR | BA | BB | BC | RKHS | |
|---|---|---|---|---|---|---|---|---|---|
| GBLUP | 0.97 | 0.82 | 0.82 | 0.97 | 1 | 0.97 | 1 | 1 | |
| RR | 0.97 | 0.9 | 0.9 | 1 | 0.97 | 1 | 0.97 | 0.97 | |
| LASSO | 0.82 | 0.9 | 1 | 0.9 | 0.82 | 0.9 | 0.82 | 0.82 | |
| EN | 0.82 | 0.9 | 1 | 0.9 | 0.82 | 0.9 | 0.82 | 0.82 | |
| BRR | 0.97 | 1 | 0.9 | 0.9 | 0.97 | 1 | 0.97 | 0.97 | |
| BA | 1 | 0.97 | 0.82 | 0.82 | 0.97 | 0.97 | 1 | 1 | |
| BB | 0.97 | 1 | 0.9 | 0.9 | 1 | 0.97 | 0.97 | 0.97 | |
| BC | 1 | 0.97 | 0.82 | 0.82 | 0.97 | 1 | 0.97 | 1 | |
| RKHS | 1 | 0.97 | 0.82 | 0.82 | 0.97 | 1 | 0.97 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tehseen, M.M.; Kehel, Z.; Sansaloni, C.P.; Lopes, M.d.S.; Amri, A.; Kurtulus, E.; Nazari, K. Comparison of Genomic Prediction Methods for Yellow, Stem, and Leaf Rust Resistance in Wheat Landraces from Afghanistan. Plants 2021, 10, 558. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030558
Tehseen MM, Kehel Z, Sansaloni CP, Lopes MdS, Amri A, Kurtulus E, Nazari K. Comparison of Genomic Prediction Methods for Yellow, Stem, and Leaf Rust Resistance in Wheat Landraces from Afghanistan. Plants. 2021; 10(3):558. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030558
Chicago/Turabian StyleTehseen, Muhammad Massub, Zakaria Kehel, Carolina P. Sansaloni, Marta da Silva Lopes, Ahmed Amri, Ezgi Kurtulus, and Kumarse Nazari. 2021. "Comparison of Genomic Prediction Methods for Yellow, Stem, and Leaf Rust Resistance in Wheat Landraces from Afghanistan" Plants 10, no. 3: 558. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030558