
Bioactive Compounds and Antioxidant Capacity in Anthocyanin-Rich Carrots: A Comparison between the Black Carrot and the Apulian Landrace “Polignano” Carrot

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
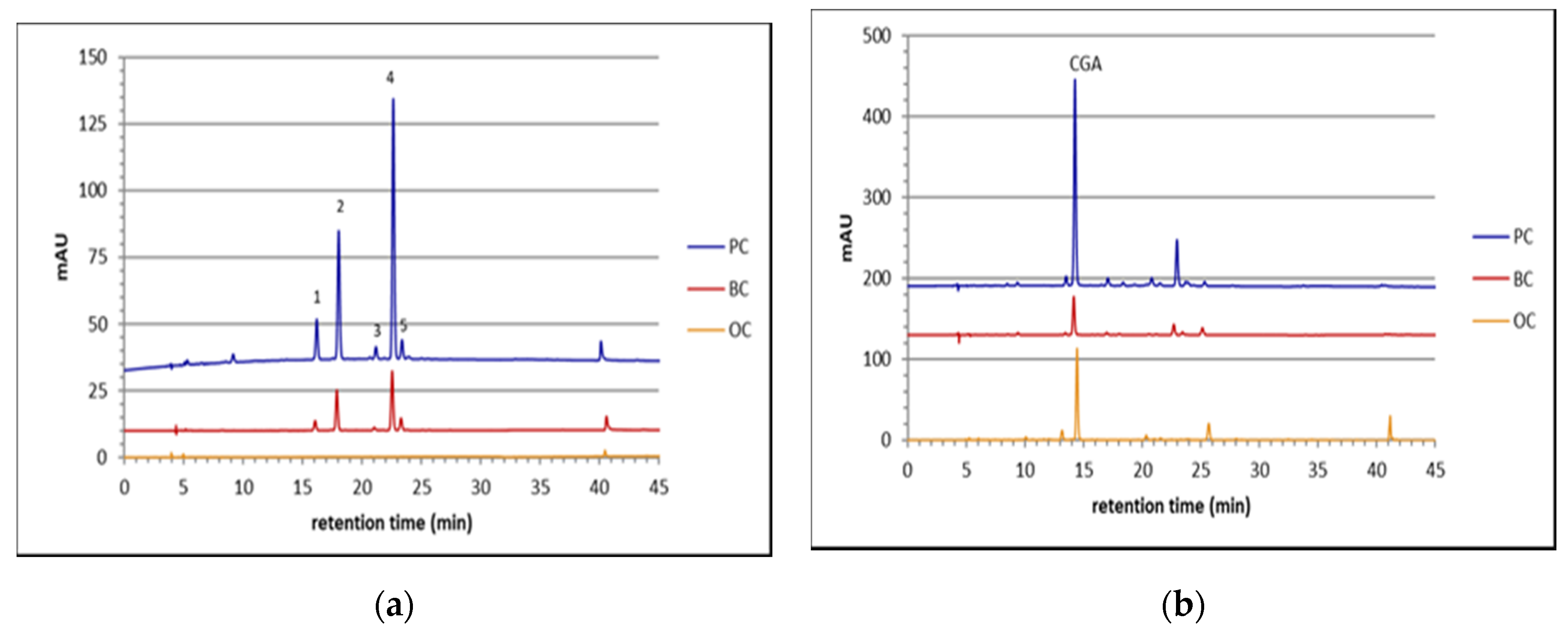
2.1. Anthocyanins
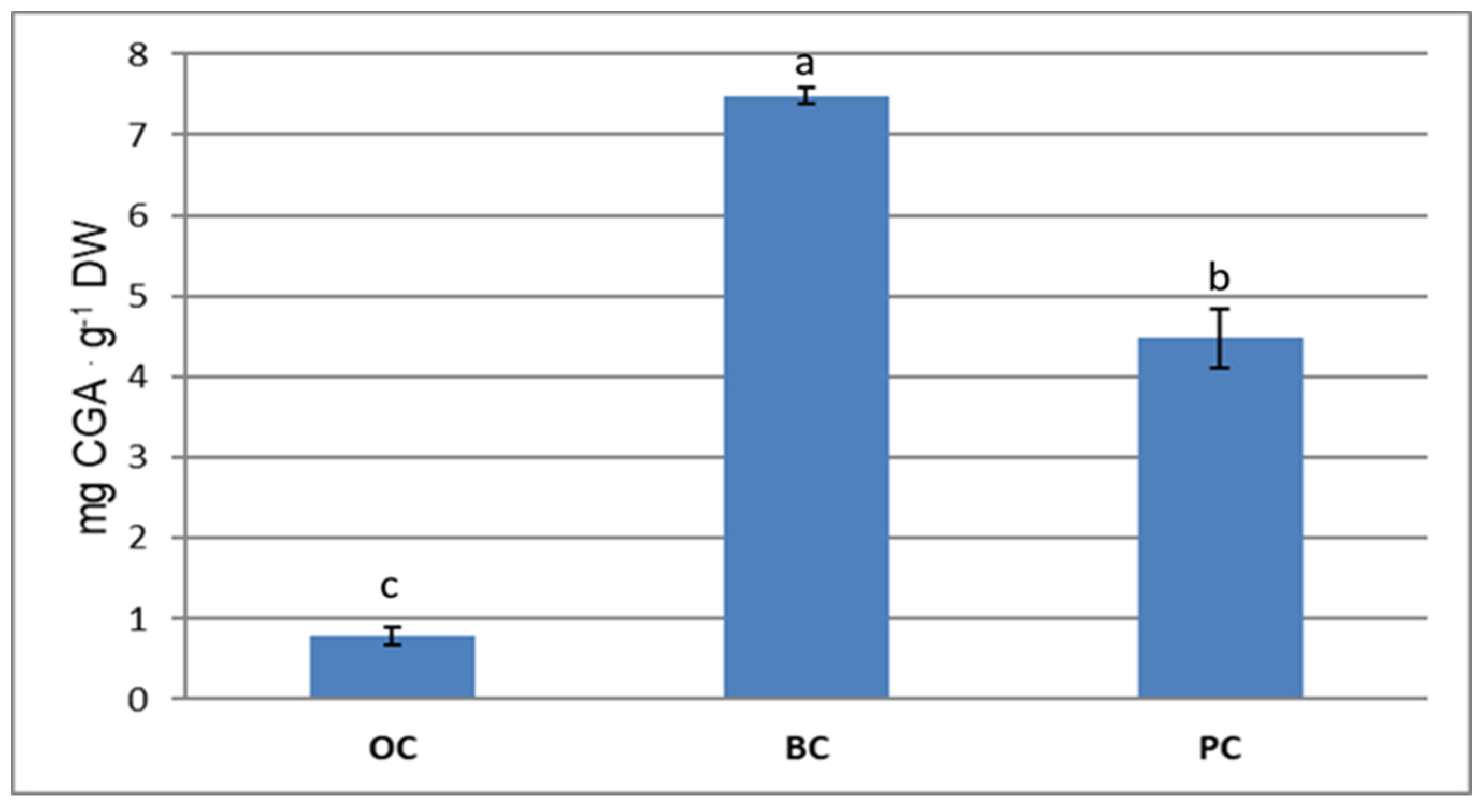
2.2. Phenolic Acids
2.3. Hydrophilic Antioxidant Capacity
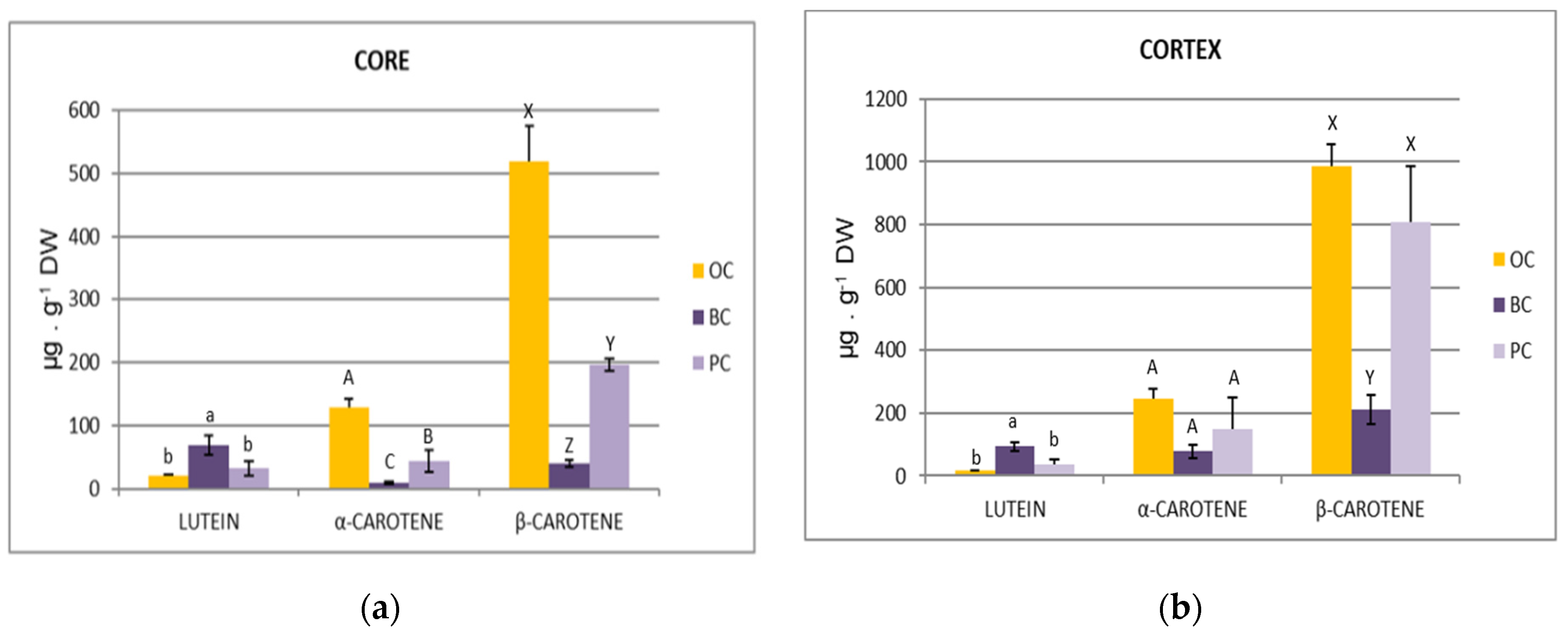
2.4. Carotenoids
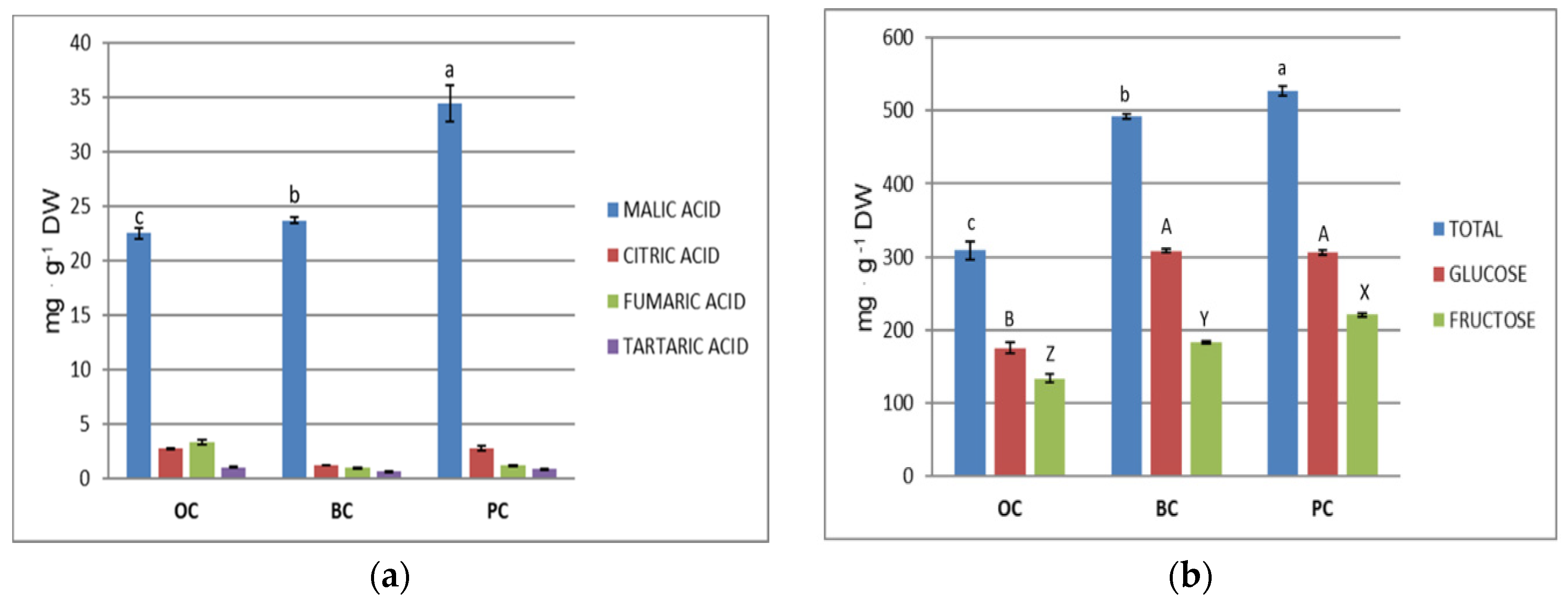
2.5. Organic Acids and Sugars
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Sample Preparation
3.4. Extraction and Analysis for Identification and Quantification of Anthocyanins and Phenolic Acids
3.5. Extraction and Analysis for Identification and Quantification of Carotenoids
3.6. Organic Acids and Sugars Extraction and Quantification
3.7. Hydrophilic Antioxidant Capacity
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Data. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 15 March 2021).
- Simon, P.W.; Wolff, X.Y. Carotenes in typical and dark orange carrots. J. Agric. Food Chem. 1987, 35, 1017–1022. [Google Scholar] [CrossRef]
- Arscott, S.A.; Tanumihardjo, S.A. Carrots of many colors provide basic nutrition and bioavailable phytochemicals acting as functional foods. Compr. Rev. Food Sci. Food Saf. 2010, 9, 223–239. [Google Scholar] [CrossRef]
- Stolarczyk, J.; Janick, J. Carrot: History and iconography. Chronica Hort. 2011, 51, 13–18. [Google Scholar]
- Iorizzo, M.; Senalik, D.A.; Ellison, S.L.; Grzebelus, D.; Cavagnaro, P.F.; Allender, C.; Brunet, J.; Spooner, D.M.; van Deynze, A.; Simon, P.W. Genetic structure and domestication of carrot (Daucus carota sbsp. sativus) (Apiaceae). Am. J. Bot. 2013, 100, 930–938. [Google Scholar] [CrossRef]
- Simon, P.W. Classical and Molecular Carrot Breeding. In The Carrot Genome; Simon, P., Iorizzo, M., Grzebelus, D., Baranski, R., Eds.; Compendium of Plant Genomes; Springer: Cham, Switzerland, 2019; pp. 137–147. [Google Scholar] [CrossRef]
- Iorizzo, M.; Curaba, J.; Pottor, M.; Ferruzzi, M.G.; Simon, P.W.; Cavagnaro, P.F. Carrot anthocyanins genetics and genomics: Status and perspectives to improve its application for the food colorant industry. Genes 2020, 11, 906. [Google Scholar] [CrossRef]
- Kammerer, D.; Carle, R.; Schieber, A. Quantification of anthocyanins in black carrot extracts (Daucus carota ssp. sativus var. atrorubens Alef.) and evaluation of their color properties. Eur. Food Res. Technol. 2004, 219, 479–486. [Google Scholar] [CrossRef]
- Espinosa-Acosta, G.; Ramos-Jacques, A.L.; Molina, G.A.; Maya-Cornejo, J.; Esparza, R.; Hernandez-Martinez, A.R.; Sánchez-González, I.; Estevez, M. Stability analysis of anthocyanins using alcoholic extracts from black carrot (Daucus carota ssp. sativus var. atrorubens Alef.). Molecules 2018, 23, 2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purkiewicz, A.; Ciborska, J.; Tańska, M.; Narwojsz, A.; Starowicz, M.; Przybyłowicz, K.E.; Sawicki, T. The impact of the method extraction and different carrot variety on the carotenoid profile, total phenolic content and antioxidant properties of juices. Plants 2020, 9, 1759. [Google Scholar] [CrossRef]
- Blando, F.; Calabriso, N.; Berland, H.; Maiorano, G.; Gerardi, C.; Carluccio, M.A.; Andersen, Ø.M. Radical scavenging and biological activities of representative anthocyanin groupings from pigment-rich fruits and vegetables. Int. J. Mol. Sci. 2018, 19, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, O.M.; Jordheim, M. Basic Anthocyanin Chemistry and Dietary Sources. In Anthocyanins in Health and Disease; Wallace, T.C., Giusti, M.M., Eds.; Taylor & Francis Inc.: Abingdon, UK; CRC Press: New York, NY, USA, 2013; pp. 13–90. [Google Scholar]
- Esatbeyoglu, T.; Rodriguez-Werner, M.; Schlösser, A.; Liehr, M.; Ipharraguerre, I.; Winterhalter, P.; Rimbach, G. Fractionation of plant bioactive from black carrots (Daucus carota subspecies sativus var. atrorubens Alef.) by adsorptive membrane chromatography and analysis of their potential anti-diabetic activity. J. Agric. Food Chem. 2016, 64, 5901–5908. [Google Scholar] [CrossRef]
- Bendokas, V.; Stanys, V.; Mažeikiene, I.; Trumbeckaite, S.; Baniene, R.; Liobikas, J. Anthocyanins: From the field to the antioxidants in the body. Antioxidants 2020, 9, 819. [Google Scholar] [CrossRef]
- Wallace, T.C.; Bailey, R.L.; Blumberg, J.B.; Burton-Freeman, B.; Chen, C.-Y.O.; Crowe-White, K.M.; Drewnowski, A.; Hooshmand, S.; Johnson, E.; Lewis, R.; et al. Fruits, vegetables, and health: A comprehensive narrative, umbrella review of the science and recommendations for enhanced public policy to improve intake. Crit. Rev. Food Sci. Nutr. 2020, 60, 2174–2211. [Google Scholar] [CrossRef] [Green Version]
- Giusti, M.M.; Wrolstad, R.E. Acylated anthocyanins from edible sources and their applications in food systems. Biochem. Eng. J. 2003, 14, 217–225. [Google Scholar] [CrossRef]
- Simon, P.W.; Rubatzky, V.E.; Basset, M.J.; Strandberg, J.O.; White, J.M. B7262, purple carrot inbred. HortScience 1997, 32, 146–147. [Google Scholar] [CrossRef] [Green Version]
- Simon, P.W.; Pollak, L.M.; Clevidence, B.A.; Holden, J.M.; Haytowitz, D.B. Plant breeding for human nutritional quality. In Plant Breeding Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; Volume 31, pp. 325–392. [Google Scholar]
- Renna, M.; Serio, F.; Signore, A.; Santamaria, P. The yellow–purple Polignano carrot (Daucus carota L.): A multicoloured landrace from the Puglia region (Southern Italy) at risk of genetic erosion. Genet. Resour. Crop Evol. 2014, 61, 1611–1619. [Google Scholar] [CrossRef]
- Renna, M.; Montesano, F.F.; Signore, A.; Gonnella, M.; Santamaria, P. BiodiverSO: A casa study of integrated project to preserve the biodiversity of vegetable crops in Puglia (Southern Italy). Agriculture 2018, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Scarano, A.; Gerardi, C.; D′Amico, L.; Accogli, R.; Santino, A. Phytochemical analysis and antioxidant properties in colored Tiggiano carrots. Agriculture 2018, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Signore, A.; Renna, M.; D′Imperio, M.; Serio, F.; Santamaria, P. Preliminary evidences of biofortification with iodine of “Carota di Polignano,” an Italian carrot landrace. Front. Plant Sci. 2018, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petropoulos, S.A.; Barros, L.; Ferreira, I.C.F.R. Rediscovering local landraces: Shaping horticulture for the future. Front. Plant Sci. 2019, 10, 126. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.; Rauf, A.; Imran, M.; Qamar, M.; Riaz, M.; Mubarak, M.S. Black carrot (Daucus carota L.), dietary and health promoting perspectives of its polyphenols: A review. Trend Food Sci. Technol. 2017, 66, 36–47. [Google Scholar] [CrossRef]
- Kammerer, D.D.; Carle, R.; Schieber, A. Characterization of phenolic acids in black carrots (Daucus carota ssp. sativus var. atrorubens Alef.) by high-performance liquid chromatography/electronspray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Montilla, E.; Arzaba, M.; Hillebrand, S.; Winterhalter, P. Anthocyanin composition of black carrot (Daucus carota ssp. sativus var. atrorubens Alef.) cultivars Antonina, Beta Sweet, Deep Purple, and Purple Haze. J. Agric. Food Chem. 2011, 59, 3385–3390. [Google Scholar] [CrossRef] [PubMed]
- Algarra, M.; Fernandes, A.; Mateus, N.; de Freitas, V.; Esteves da Silva, J.; Casado, J. Anthocyanin profile and antioxidant capacity of black carrots (Daucus carota L. ssp. sativus var. atrorubens Alef.) from Cuevas Bajas, Spain. J. Food Comp. Anal. 2014, 33, 71–76. [Google Scholar] [CrossRef]
- Yldiz, M.; Willis, D.K.; Cavagnaro, P.F.; Iorizzo, M.; Abak, K.; Simon, P.W. Expression and mapping of anthocyanin biosynthesis genes in carrot. Theor. Appl. Genet. 2013, 126, 1689–1702. [Google Scholar] [CrossRef] [PubMed]
- Mizgier, P.; Kucharska, A.Z.; Sokół-Łętowska, A.; Kolniak-Ostek, J.; Kidoń, M.; Fecka, I. Characterization of phenolic compounds and antioxidant and anti-inflammatory properties of red cabbage and purple carrot extracts. J. Funct. Food 2016, 21, 133–146. [Google Scholar] [CrossRef]
- Holme, I.B.; Dionisio, G.; Brinch-Pedersen, H. A roadmap to modulated anthocyanin compositions in carrots. Plants 2021, 10, 472. [Google Scholar] [CrossRef]
- Smeriglio, A.; Denaro, M.; Barreca, D.; D′Angelo, V.; Germanò, M.P.; Trombetta, D. Polyphenolic profile and biological activities of black carrot crude extract (Daucus carota L. ssp. sativus var. atrorubens Alef.). Fitoterapia 2018, 124, 49–57. [Google Scholar] [CrossRef]
- Alasalvar, C.; Al-Farsi, M.; Quantick, P.C.; Shahidi, F.; Wiktorowicz, R. Effect of chill storage and modified atmosphere packaging (MAP) on antioxidant activity, anthocyanins, carotenoids, phenolics and sensory quality of ready-to-eat shredded orange and purple carrots. Food Chem. 2005, 89, 69–76. [Google Scholar] [CrossRef]
- Leja, M.; Kamińska, I.; Kramer, M.; Maksylewicz-Kaul, A.; Kammerer, D.; Carle, R.; Baranski, R. The content of phenolic compounds and radical scavenging activity varies with carrot origin and root color. Plant Foods Hum. Nutr. 2013, 68, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Müller, R.; Acosta-Motos, J.R.; Großkinsky, D.K.; Hernández, J.A.; Lütken, H.; Barba-Espin, G. UV-B exposure of black carrot (Daucus carota ssp. sativus var. atrorubens) plants promotes growth, accumulation of anthocyanins, and phenolic compounds. Agronomy 2019, 9, 323. [Google Scholar] [CrossRef] [Green Version]
- Bannoud, F.; Ellison, S.; Paolinelli, M.; Horejsi, T.; Senalik, D.; Fanzone, M.; Iorizzo, M.; Simon, P.W.; Cavagnaro, P. Dissecting the genetic control of root and leaf tissue-specific anthocyanin pigmentation in carrot (Daucus carota L.). Theor. Appl. Genet. 2019, 132, 2485–2507. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Ellison, S.; Pottorff, M.; Cavagnaro, P.F. Carrot Molecular Genetics and Mapping. In The Carrot Genome; Simon, P., Iorizzo, M., Grzebelus, D., Baranski, R., Eds.; Compendium of Plant Genomes; Springer: Cham, Switzerland, 2019; pp. 101–117. [Google Scholar] [CrossRef]
- Alasalvar, C.; Grigor, J.M.; Zhang, D.; Quantick, P.C.; Shahidi, F. Comparison of volatiles, phenolics, sugars, antioxidant vitamins, and sensory quality of different colored carrot varieties. J. Agric. Food Chem. 2001, 49, 1410–1416. [Google Scholar] [CrossRef]
- Sun, T.; Simon, P.W.; Tanumihardjo, S. Antioxidant phytochemicals and antioxidant capacity of biofortified carrots (Daucus carota L.) of various colors. J. Agric. Food Chem. 2009, 57, 4142–4147. [Google Scholar] [CrossRef] [PubMed]
- Pace, B.; Capotorto, I.; Cefola, M.; Minasi, P.; Montemurro, N.; Carbone, V. Evaluation of quality, phenolic and carotenoid composition of fresh-cut purple Polignano carrots stored in modified atmosphere. J. Food Compos. Anal. 2020, 86, 103363. [Google Scholar] [CrossRef]
- Nicolle, C.; Simon, G.; Rock, E.; Amouroux, P.; Rémésy, C. Genetic variability influences carotenoid, vitamin, phenolic and mineral content in white, yellow, purple, orange and dark-orange carrot cultivars. J. Am. Soc. Hort. Sci. 2004, 129, 523–529. [Google Scholar] [CrossRef] [Green Version]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Kevers, C.; Falkowski, M.; Tabart, J.; Defraigne, J.-O.; Dommes, J.; Pincemail, J. Evolution of antioxidant capacity during storage of selected fruits and vegetables. J. Agric. Food Chem. 2007, 55, 8596–8603. [Google Scholar] [CrossRef]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing Oxygen Radical Absorbance Capacity (ORAC) and Ferric Reducing Antioxidant Power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef]
- Grassmann, J.; Schnitzler, W.H.; Habegger, R. Evaluation of different coloured carrot cultivars on antioxidative capacity based ontheir carotenoid and phenolic contents. Int. J. Food Sci. Nutr. 2007, 58, 603–611. [Google Scholar] [CrossRef]
- Cefola, M.; Pace, B.; Renna, M.; Santamaria, P.; Signore, A.; Serio, F. Compositional analysis and antioxidant profile of yellow, orange and purple Polignano carrots. Ital. J. Food Sci. 2012, 24, 284–291. [Google Scholar]
- Perrin, F.; Hartmann, L.; Dubois-Laurent, C.; Welsch, R.; Huet, S.; Hamama, L.; Briard, M.; Peltier, D.; Gagné, S.; Geoffriau, E. Carotenoid gene expression explains the difference of carotenoid accumulation in carrot root tissues. Planta 2017, 245, 737–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruhl, I.; Herrmann, K. Organic acids of vegetables. I. Brassica species, leaf and bulbous vegetables, carrots, celery. Z Lebensm Unters Forsch 1985, 180, 215–220. [Google Scholar] [CrossRef]
- Timpanaro, G.; Bellia, C.; Foti, V.T.; Scuderi, A. Consumer behaviour of purchasing biofortified food products. Sustainability 2020, 12, 6297. [Google Scholar] [CrossRef]
- Que, F.; Hou, X.-L.; Wang, G.-L.; Xu, Z.-S.; Tan, G.-F.; Li, T.; Wang, Y.-H.; Khadr, A.; Xiong, A.-S. Advances in research on the carrot, an important root vegetable in the Apiaceae family. Hortic. Res. 2019, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Holme, I.B.; Dionisio, G.; Kodama, M.; Dzhanfezova, T.; Joernsgaard, B.; Brinch-Pedersen, H. Cyanidin-based anthocyanin biosynthesis in orange carrot is restored by expression of AmRosea1 and AmDelila, MYB and bHLH transcription factors. Plant Mol. Biol. 2020, 103, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Yang, Q.; Feng, K.; Yu, X.; Xiong, A. DcMYB113, a root-specific R2R3-MYB, conditions anthocyanin biosynthesis and modification in carrot. Plant Biotechnol. J. 2020, 18, 1585–1597. [Google Scholar] [CrossRef] [Green Version]
- Gerardi, C.; Albano, C.; Calabriso, N.; Carluccio, M.A.; Durante, M.; Mita, G.; Renna, M.; Serio, F.; Blando, F. Techno-functional properties of tomato puree fortified with anthocyanin pigments. Food Chem. 2018, 240, 1184–1192. [Google Scholar] [CrossRef]
- Chandra, A.; Rana, J.; Li, Y. Separation, identification, quantification and method validation of anthocyanins in botanica row materials by HPLC and HPLC/MS. J. Agric. Food Chem. 2001, 49, 3515–3521. [Google Scholar] [CrossRef] [PubMed]
- Koch, T.C.; Goldman, I.L. Relationship of carotenoids and tocopherols in a sample of carrot root-color accessions and carrot germplasm carrying Rp and rp alleles. J. Agric. Food Chem. 2005, 53, 325–331. [Google Scholar] [CrossRef]
- Durante, M.; Lenucci, M.S.; Marrese, P.P.; Rizzi, V.; De Caroli, M.; Piro, G.; Fini, P.; Russo, G.L.; Mita, G. α-Cyclodextrin encapsulation of supercritical CO2 extracted oleoresins from different plant matrices: A stability study. Food Chem. 2016, 199, 684–693. [Google Scholar] [CrossRef]
- Blando, F.; Gerardi, C.; Renna, M.; Castellano, S.; Serio, F. Characterization of bioactive compounds in berries from plants grown under innovative photovoltaic greenhouses. J. Berry Res. 2018, 8, 55–69. [Google Scholar] [CrossRef]
- Blando, F.; Berland, H.; Maiorano, G.; Durante, M.; Mazzucato, A.; Picarella, M.E.; Nicoletti, I.; Gerardi, C.; Mita, G.; Andersen, O.M. Nutraceutical characterization of anthocyanin-rich fruits produced by “Sun Black′ tomato lines. Front. Nutr. 2019, 6, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Anth. 1 | Anth. 2 | Anth. 3 | Anth. 4 | Anth. 5 | Total |
|---|---|---|---|---|---|---|
| BC core | 0.25 ± 0.007 d | 1.14 ± 0.03 b | 0.33 ± 0.004 c | 5.39 ± 0.06 a | 0.39 ± 0.03 c | 7.49 ± 0.13 C |
| BC cortex | 1.11 ± 0.03 d | 5.92 ± 0.19 b | 0.50 ± 0.03 d | 9.66 ± 0.24 a | 2.26 ± 0.12 c | 19.45 ± 0.62 A |
| BC whole | 0.97 ± 0.07 cd | 3.28 ± 0.14 b | 0.37 ± 0.02 d | 7.69 ± 0.27 a | 1.52 ± 0.11 c | 13.84 ± 0.61 B |
| PC1 core | n.d. | n.d. | n.d. | 0.007 ± 0.0009 | n.d. | 0.007 ± 0.0009 F |
| PC1 cortex | 0.20 ± 0.002 c | 1.02 ± 0.007 b | 0.13 ± 0.001 c | 3.25 ± 0.03 a | 0.24 ± 0.002 c | 4.85 ± 0.05 D |
| PC1 whole | 0.39 ± 0.002 c | 1.05 ± 0.02 b | 0.12 ± 0.006 e | 3.28 ± 0.12 a | 0.21 ± 0.003 d | 5.06 ± 0.15 D |
| PC2 core | 0.02 ± 0.0004 b | 0.01 ± 0.003 c | 0.01 ± 0.0004 c | 0.43 ± 0.005 a | 0.006 ± 0.001 d | 0.49± 0.01 E |
| PC2 cortex | 0.27 ± 0.0001 c | 0.82 ± 0.0005 b | 0.09 ± 0 d | 6.82 ± 0.05 a | 0.18 ± 0.004 c | 8.18 ± 0.58 C |
| PC2 whole | 0.32 ± 0.01 c | 0.68 ± 0.01 b | 0.11 ± 0.007 e | 6.56 ± 0.25 a | 0.14 ± 0.002 d | 7.82 ± 0.28 C |
| FCR | TEAC | ORAC | |
|---|---|---|---|
| Sample | mg GAE.g−1 DW | μmol TE. g−1 DW | μmol TE. g−1 DW |
| C | 2.25 ± 0.31 c | 6.19 ± 0.84 c | 21.43 ± 3.06 c |
| BC | 16.57 ± 1.13 a | 76.67 ± 10.6 a | 159.93 ± 3.28 a |
| PC | 12.98 ± 0.89 b | 54.99 ± 5.53 b | 101.32 ± 14.17 b |
| Sample | Lutein | α-Carotene | β-Carotene | Total Carotenoids |
|---|---|---|---|---|
| OC | 17.2 ± 1.36 c | 255 ± 19.30 a | 1016.35 ± 68.5 a | 1288.55 ± 89.16 a |
| BC | 57.58 ± 10.60 a | 21.87 ± 3.85 b | 60.38 ± 10 c | 139.83 ± 24.45 c |
| PC | 37.42 ± 4.35 b | 47.26 ± 5.26 b | 247.45 ± 29 b | 332.13 ± 38.61 b |
| Lutein | α-Carotene | β-Carotene | Total Carotenoids | ||
|---|---|---|---|---|---|
| PC1 | core | 27.97 ± 0.93 b | n.d. | 4.23 ± 0.05 d | 32.20 ± 0.99 d |
| cortex | 48.82 ± 1.86 a | 37.73 ± 3.02 b | 98.95 ± 12.37 c | 185.50 ± 17.25 c | |
| PC2 | core | 28.35 ± 5.37 b | 37.52 ± 4.07 b | 206 ± 26.36 b | 271.87 ± 35.8 b |
| cortex | 13.21 ± 0.37 c | 369.65 ± 1.48 a | 899.49 ± 7.78 a | 1282.35 ± 9.63 a | |
| Sample | Anthocyanins | FCR | Carotenoids | TEAC Value | ORAC Value |
|---|---|---|---|---|---|
| mg K Eq.100–1 g FW | mg GAE. 100–1 g FW | mg.100–1 g FW | μmol TE.100–1 g FW | μmol TE.100–1 g FW | |
| OC | n.d. | 23.11 ± 3.24 c | 14.15 ± 0.81 a | 63.82 ± 8.24 c | 250.7 ± 35.81 c |
| BC | 186.85 ± 8.13 a | 222 ± 15.13 a | 1.82 ± 0.22 b | 1026.43 ± 142 a | 2159 ± 44.35 a |
| PC | 55.13± 23.12 b | 111.10 ± 7.61 b | 2.81 ± 0.76 b | 470.21 ± 47 b | 866.3 ± 121.12 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blando, F.; Marchello, S.; Maiorano, G.; Durante, M.; Signore, A.; Laus, M.N.; Soccio, M.; Mita, G. Bioactive Compounds and Antioxidant Capacity in Anthocyanin-Rich Carrots: A Comparison between the Black Carrot and the Apulian Landrace “Polignano” Carrot. Plants 2021, 10, 564. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030564
Blando F, Marchello S, Maiorano G, Durante M, Signore A, Laus MN, Soccio M, Mita G. Bioactive Compounds and Antioxidant Capacity in Anthocyanin-Rich Carrots: A Comparison between the Black Carrot and the Apulian Landrace “Polignano” Carrot. Plants. 2021; 10(3):564. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030564
Chicago/Turabian StyleBlando, Federica, Stefano Marchello, Gabriele Maiorano, Miriana Durante, Angelo Signore, Maura N. Laus, Mario Soccio, and Giovanni Mita. 2021. "Bioactive Compounds and Antioxidant Capacity in Anthocyanin-Rich Carrots: A Comparison between the Black Carrot and the Apulian Landrace “Polignano” Carrot" Plants 10, no. 3: 564. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10030564