
Breeding of High Cooking and Eating Quality in Rice by Marker-Assisted Backcrossing (MABc) Using KASP Markers

 , and
, and
Abstract
:1. Introduction
2. Results
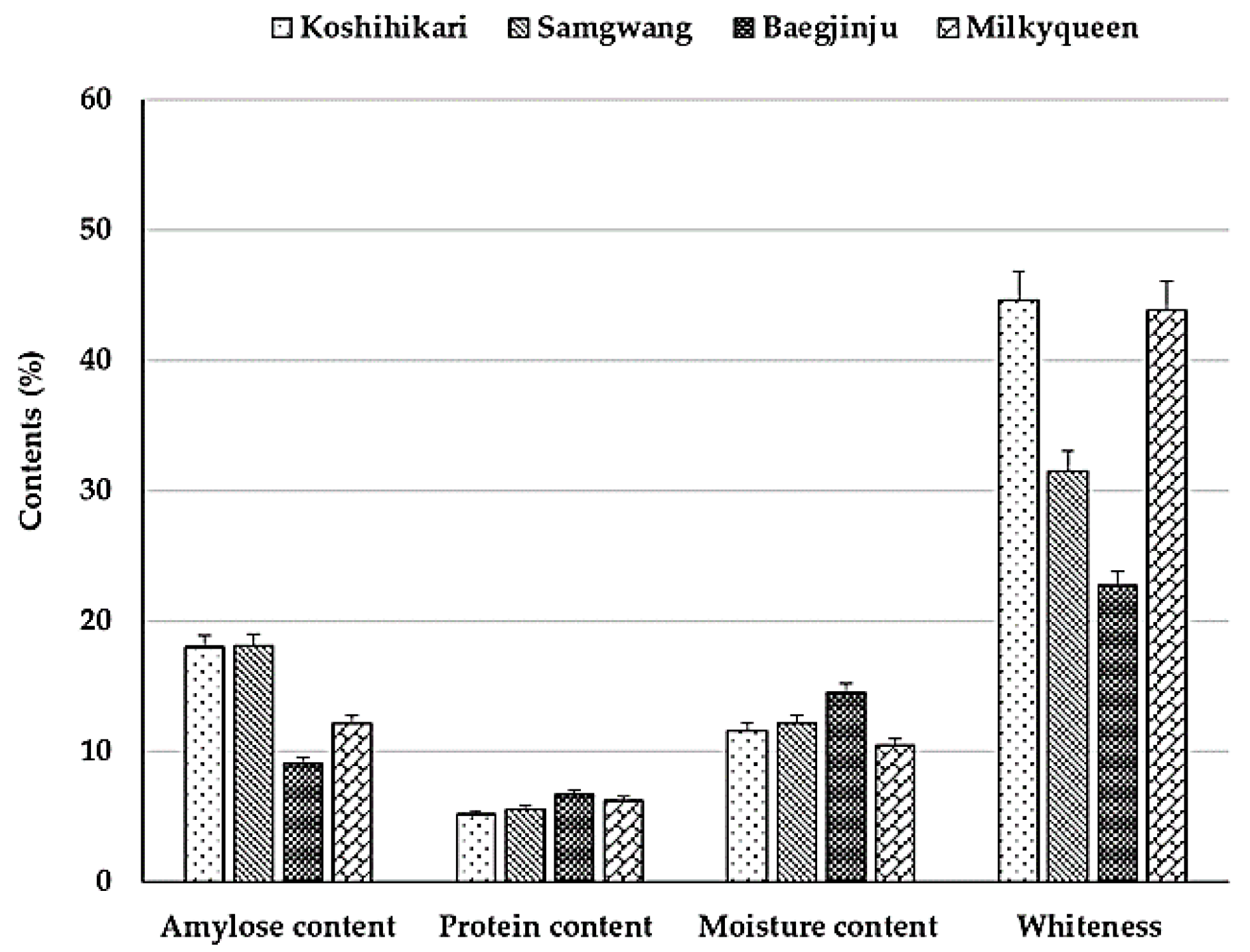
2.1. Evaluation of Characteristics Related to Cooking and Eating Quality in Parents
2.1.1. Characteristics Related to Cooking and Texture
2.1.2. Characteristics Related to Eating Quality
2.1.3. Viscosity Characteristics by RVA
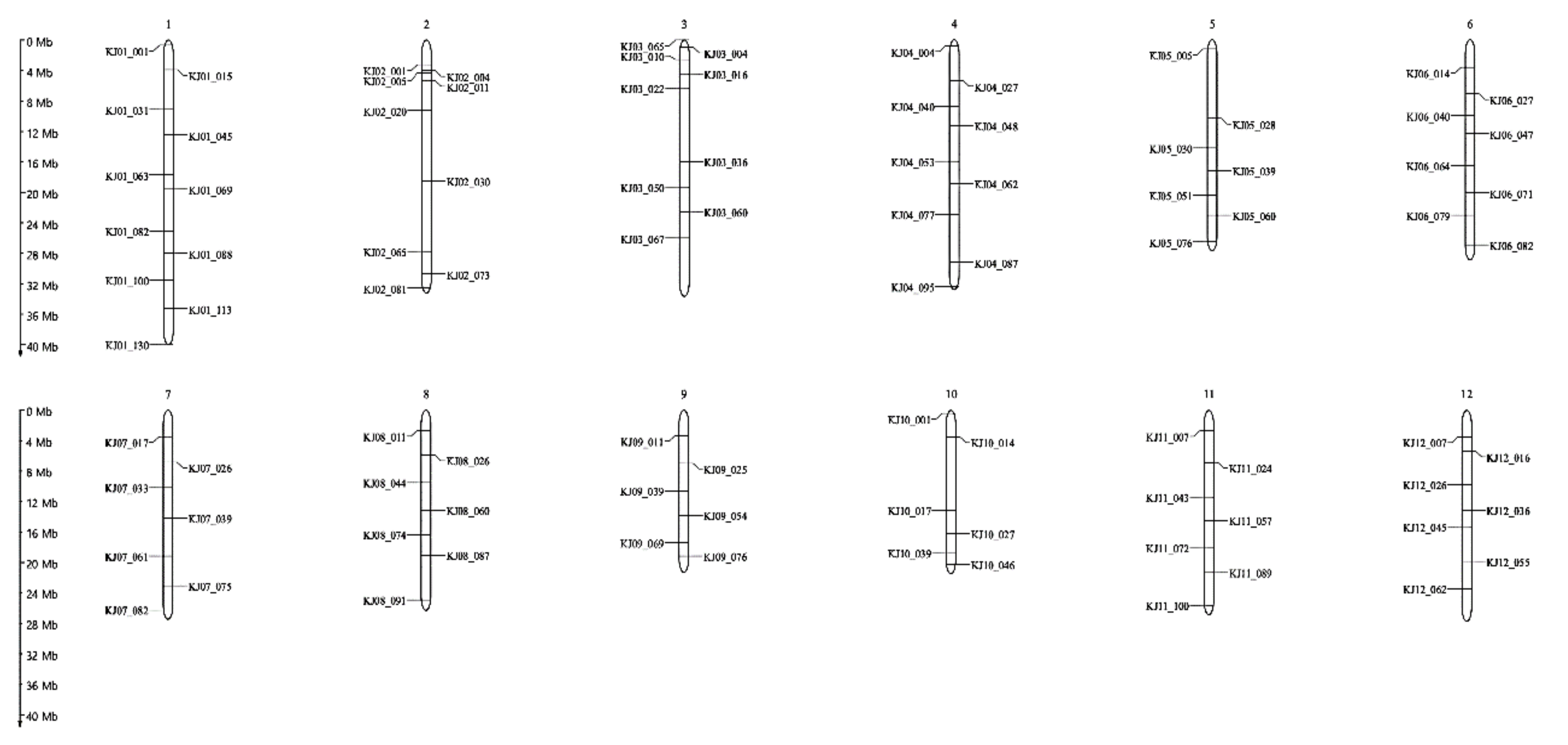
2.2. Whole-Genome Re-sequencing Analysis and Variant Discovery for Foreground Selection
2.2.1. Foreground Selection in BC1F1 and BC2F1
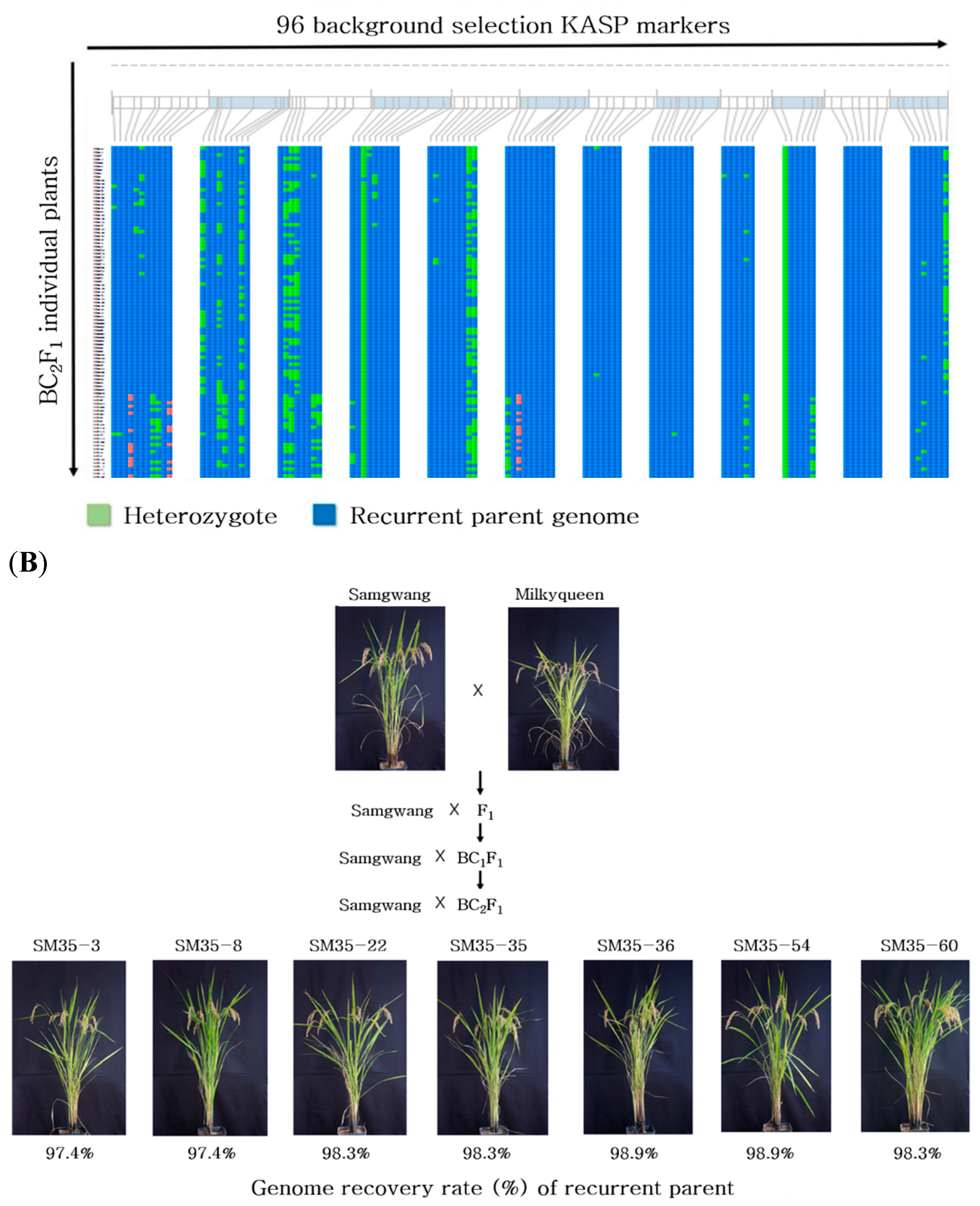
2.2.2. Background Selection in BC1F1 and BC2F1 Generation
2.3. Agronomic Traits of the Selected Lines in the BC2F2 Generation
2.4. Viscosity Properties of the Selected Lines in BC2F2 Generation
3. Discussion
4. Materials and Methods
Plant Materials
5. Cooking and Eating Quality Characteristics of Milled Rice
5.1. Analysis of Amylose and Protein Contents
5.2. Characterizations of Cooking and Eating Texture
5.3. Rapid Visco Analysis (RVA)
6. MABc Breeding Process
7. Molecular Marker Analysis
7.1. Semi-Nested PCR Analysis for Foreground Selection
7.2. KASP Marker Analysis for Background Selection and Investigation of Agronomic Traits
8. Statistical Analysis
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rohit, R.; Parmar, K. Unified approach in food quality evaluation using machine vision, Part III. Commun. Comput. Inf. Sci. 2011, 192, 239–248. [Google Scholar]
- Aluko, G.; Martinez, C.; Tohme, J.; Castano, C.; Bergman, C.; Oard, J.H. QTL mapping of grain quality traits from the interspecific cross Oryza sativa x O. glaberrima. Theor. Appl. Genet. 2004, 109, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Rabiei, B.; Valizadeh, M.; Ghareyazie, B.; Moghaddam, M.; Ali, A.J. Identification of QTLs for rice grain size and shape of Iranian cultivars using SSR markers. Euphytica 2004, 137, 325–332. [Google Scholar] [CrossRef]
- Sun, M.M.; Abdula, S.E.; Lee, H.J.; Cho, Y.C.; Han, L.Z.; Koh, H.J.; Cho, Y.G. Molecular Aspect of Good Eating Quality Formation in Japonica Rice. PLoS ONE 2011, 6, e18385. [Google Scholar] [CrossRef] [Green Version]
- Cornejo-Ramírez, Y.I.; Martínez-Cruz, O.; Toro-Sánchez, C.L.D.; Wong-Corral, F.J.; Borboa-Flores, J.; Cinco-Moroyoqui, F.J. The structural characteristics of starches and their functional properties, CyTA. J. Food 2018, 16, 1003–1017. [Google Scholar]
- Bao, J.; Shen, S.; Sun, M.; Corke, H. Analysis of genotypic diversity in the starch physicochemical properties of nonwaxy rice: Apparent amylose content, pasting viscosity and gel texture. Starch 2006, 58, 259–267. [Google Scholar] [CrossRef]
- Pang, Y.; Ali, J.; Wang, X.; Franj, N.J.; Revilleza, J.E.; Xu, J.; Li, Z. Relationship of Rice Grain Amylose, Gelatinization Temperature and Pasting Properties for Breeding Better Eating and Cooking Quality of Rice Varieties. PLoS ONE 2016, 11, e0168483. [Google Scholar] [CrossRef] [Green Version]
- Hanashiro, I.; Itoh, K.; Kuratomi, Y.; Yamazaki, M.; Igarashi, T.; Matsugasako, J.; Takeda, Y. Granule-bound starch synthase I is responsible for biosynthesis of extra-long unit chains of amylopectin in rice. Plant Cell Physiol. 2008, 49, 925–933. [Google Scholar] [CrossRef] [Green Version]
- Comparot-Moss, S.; Denyer, K. The evolution of the starch biosynthetic pathway in cereals and other grasses. J. Exp. Bot. 2009, 60, 2481–2492. [Google Scholar] [CrossRef] [Green Version]
- Hirose, T.; Terao, T. A comprehensive expression analysis of the starch synthase gene family in rice (Oryza sativa L.). Planta 2004, 220, 9–16. [Google Scholar] [CrossRef]
- Ohdan, T.; Francisco, P.B.; Sawada, T.; Hirose, T.; Terao, T.; Satoh, H.; Nakamura, Y. Expression profiling of genes involved in starch synthesis in sink and source organs of rice. J. Exp. Bot. 2005, 56, 3229–3244. [Google Scholar] [CrossRef] [Green Version]
- Fujita, N.; Yoshida, M.; Kondo, T.; Saito, K.; Utsumi, Y.; Tokunaga, T.; Nishi, A.; Park, J.H.; Jane, J.L.; Miyao, A.; et al. Characterization of SSIIIa-deficient mutants of rice: The function of SSIIIa and pleiotropic effects by SSIIIa deficiency in the rice endosperm. Plant Physiol. 2007, 144, 2009–2023. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Pang, Y.; Zhang, J.; Wu, Z.; Chen, K.; Ali, J.; Ye, G.; Xu, J.; Li, Z. Genome-wide and gene-based associated mapping for rice eating and cooking characteristics and protein content. Sci. Rep. 2017, 7, 17203. [Google Scholar] [CrossRef] [Green Version]
- Tian, Z.; Qian, Q.; Liu, Q.; Yan, M.; Liu, X.; Yan, C.; Liu, G.; Gao, Z.; Tang, S.; Zeng, D.; et al. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc. Natl. Acad. Sci. USA 2009, 106, 21760–21765. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Corke, H. Factor analysis of physicochemical properties of 63 rice varieties. J. Sci. Food Agric. 2002, 82, 745–752. [Google Scholar] [CrossRef]
- Sundaram, R.M.; Vishnupriya, M.R.; Biradar, S.K.; Laha, G.S.; Reddy, G.A.; Rani, G.N.; Sarma, N.P.; Sonti, R.V. Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphutica 2008, 160, 411–422. [Google Scholar] [CrossRef]
- Suh, J.P.; Jeung, J.U.; Noh, T.H.; Cho, Y.C.; Park, S.H.; Park, H.S.; Shin, M.S.; Kim, C.K.; Jena, K.K. Development of breeding lines with three pyramided resistance genes that confer broad-spectrum bacterial blight resistance and their molecular analysis in rice. Rice 2013, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Latif, M.A. Recurrent parent genome recovery analysis in a marker-assisted backcrossing program of rice (Oryza sativa L.). Comptes Rendus Biol. 2015, 338, 83–94. [Google Scholar] [CrossRef]
- Frisch, M.; Bohn, M.; Melchinger, A.E. Comparison of selection strategies for marker-assisted backcrossing of a gene. Crop Sci. 1999, 39, 1295–1301. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.M.; Rafii, M.Y.; Ismail, M.R.; Mahmood, M.; Rahim, H.A.; Alam, M.A.; Ashkani, S.; Malek, M.A.; Latif, M.A. Marker-assisted backcrossing: A useful method for rice improvement. Biotechnol. Biotechnol. Equip. 2015, 29, 237–254. [Google Scholar] [CrossRef] [Green Version]
- Chukwu, S.C.; Rafii, M.Y.; Ramlee, S.I.; Ismail, S.I.; Oladosu, Y.; Okporie, E.; Onyishi, G.; Utobo, E.; Ekwu, L.; Swaray, S.; et al. Marker-assisted selection and gene pyramiding for resistance to bacterial leaf blight disease of rice (Oryza sativa L.). Biotechnol. Biotechnol. Equip. 2019, 33, 440–455. [Google Scholar] [CrossRef] [Green Version]
- Poczai, P.; Varga, I.; Laos, M.; Cseh, A.; Bell, N.; Valkonen, J.P.; Hyvonen, J. Advances in plant gene-targeted and functional markers: A review. Plant Methods 2013, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single nucleotide polymorphism genotyping using kompetitive allele specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol. Breed. 2014, 33, 1–14. [Google Scholar] [CrossRef]
- Saxena, R.K.; Penmetsa, R.V.; Upadhyaya, H.D.; Kumar, A.; Carrasquilla-Garcia, N.; Schlueter, J.A.; Farmer, A.; Whaley, A.M.; Sarma, B.K.; May, G.D.; et al. Large-scale development of cost-effective single-nucleotide polymorphism marker assays for genetic mapping in pigeonpea and comparative mapping in legumes. DNA Res. 2012, 19, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Hiremath, P.J.; Kumar, A.; Penmetsa, R.V.; Farmer, A.; Schlueter, J.A.; Chamarthi, S.K.; Whaley, A.M.; Carrasquilla-Garcia, N.; Gaur, P.M.; Upadhyaya, H.D.; et al. Large-scale development of cost-effective SNP marker assays for diversity assessment and genetic mapping in chickpea and comparative mapping in legumes. Plant Biotechnol. J. 2012, 10, 716–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pariasca-Tanaka, J.; Lorieux, M.; He, C.; McCouch, S.; Thomson, M.J. Wissuwa M Development of a SNP genotyping panel for detecting polymorphisms in Oryza glaberrima/O. sativa interspecific crosses. Euphytica 2015, 201, 67–78. [Google Scholar] [CrossRef]
- Cheon, K.S.; Baek, J.H.; Cho, Y.I.; Jeong, Y.M.; Lee, Y.Y.; Jun, O.; Won, Y.J.; Kang, D.Y.; Oh, H.J.; Kim, S.L.; et al. Single Nucleotide Polymorphism (SNP) Discovery and Kompetitive Allele-Specific PCR (KASP) Marker Development with Korean Japonica Rice. Plant Breed. Biotech. 2018, 6, 391–403. [Google Scholar] [CrossRef]
- Cheon, K.S.; Jeong, Y.M.; Lee, Y.Y.; Oh, J.; Kang, D.Y.; Oh, H.J.; Kim, S.L.; Kim, N.H.; Lee, E.Y.; Baek, J.H.; et al. Kompetitive Allele-Specific PCR Marker Development and Quantitative Trait Locus Mapping for Bakanae Disease Resistance in Korean Japonica Rice Varieties. Plant Breed. Biotechnol. 2019, 7, 208–219. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kiyosumi, H.; Toshio, Y.; Masahiro, Y. Koshihikari: A premium short-grain rice cultivar—Its expansion and breeding in Japan. Rice 2018, 11, 15. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Cho, J.H.; Lee, J.Y.; Sohn, Y.B.; Han, S.I.; Shin, D.; Song, Y.C.; Park, D.-S.; Yeo, U.-S.; Oh, M.-G.; et al. Genetic Analysis and Molecular Mapping of Low Amylose Gene in Rice Cultivar ‘Baegjinju’ (Oryza sativa L.). Korean J. Breed. Sci. 2016, 48, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Correia, P.M.R.; Alves, M.; Lemos, D.; Guiné, R.P.F. Contribution for Physical Characterization of Carolino Rice. In Proceedings of the NAFI 2014—International Food Congress, Kusadasi, Turquia, 26–29 May 2014; p. 208. [Google Scholar]
- Choi, H.C. Current status and perspectives in varietal improvement of rice cultivars for high-quality and value-added products. Korean J. Crop Sci. 2002, 47, 15–32. [Google Scholar]
- Lin, L.; Lu, F.; Chang, Y. Prediction of protein content in rice using a near-infrared imaging system as diagnostic technique. Int. J. Agric. Biol. Eng. 2019, 12, 195–200. [Google Scholar] [CrossRef]
- Lestari, P.; Lee, G.; Ham, T.H.; Woo, M.O.; Piao, R.; Jiang, W.; Chu, S.H.; Lee, J.H.; Koh, H.J. Single Nucleotide Polymorphisms and Haplotype Diversity in Rice Sucrose Synthase 3. J. Hered. 2011, 102, 735–746. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.J.; Lee, S.Y.; Kim, Y.R.; Park, J.W.; Shim, J.Y. Effect of dry heating on the pasting/retrogradation and textural properties of starch-soy protein maxture. Korean J. Food Sci. Technol. 2004, 36, 568–573. [Google Scholar]
- Kwakt, T.S. Comparison of Amylogram Properties among Several Subspecies of Rice. Korean J. Crop Sci. 2005, 50, 186–190. [Google Scholar]
- Kim, W.S.; Shin, M. The properties of rice flours prepared by dry- and wet-milling of soaked glutinous and normal grains. Korean J. Food Cook. Sci. 2007, 23, 908. [Google Scholar]
- Lee, H.M.; Lee, Y.T. Pasting Properties of Corn, Potato, Sweet Potato Starches and Wheat Flours with Partial Rice Starch Substitution. Food Eng. Prog. 2013, 17, 238–244. [Google Scholar] [CrossRef]
- Kesarwani, A.; Chiang, P.Y.; Chen, S.S. Rapid Visco Analyzer Measurements of japonica Rice Cultivars to Study Interrelationship between Pasting Properties and Farming System. Int. J. Agronomy. 2016, 2016, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bashir, K.; Aggarwal, M. Physicochemical, structural and functional properties of native and irradiated starch: A review. J. Food Sci. Technol. 2019, 56, 513–523. [Google Scholar] [CrossRef]
- Chen, G.X.; Zhu, J.T.; Zhou, J.W.; Subburaj, S.; Zhang, M.; Han, C.X.; Hao, P.C.; Li, X.H.; Yan, Y.M. Dynamic development of starch granules and the regulation of starch biosynthesis in Brachypodium distachyon: Comparison with common wheat and Aegilops peregrina. BMC Plant Biol. 2014, 14, 198. [Google Scholar] [CrossRef] [Green Version]
- Crofts, N.; Abe, K.; Aihara, S.; Itoh, R.; Nakamura, Y.; Itoh, K.; Fujita, N. Lack of starch synthase IIIa and high expression of granule-bound starch synthase I synergistically increase the apparent amylose content in rice endosperm. Plant Sci. 2012, 193–194, 62–69. [Google Scholar] [CrossRef]
- Zhang, A.; Gao, Y.; Li, Y.; Ruan, B.; Yang, S.; Liu, C.; Zhang, B.; Jiang, H.; Fang, G.; Ding, S.; et al. Genetic Analysis for Cooking and Eating Quality of Super Rice and Fine Mapping of a Novel Locus qGC10 for Gel Consistency. Front. Plant Sci. 2020, 24, 342. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, S.; Dang, X.; Liu, E.; Hu, X.; Eltahawy, M.S.; Zaid, I.U.; Hong, D. Favorable alleles mining for gelatinization temperature, gel consistency and amylose content in Oryza sativa by association mapping. BMC Genet. 2019, 20, 34. [Google Scholar] [CrossRef]
- Yang, B.; Xu, S.; Xu, L.; You, H.; Xiang, X. Effects of Wx and Its Interaction With SSIII-2 on Rice Eating and Cooking Qualities. Front. Plant Sci. 2018, 9, 456. [Google Scholar] [CrossRef]
- Kharabian-Masouleh, A.; Waters, D.L.; Reinke, R.F.; Ward, R.; Henry, R.J. SNP in starch biosynthesis genes associated with nutritional and functional properties of rice. Sci. Rep. 2012, 2, 557. [Google Scholar] [CrossRef]
- Umemoto, T.; Yano, M.; Satoh, H.; Shomura, A.; Nakamura, Y. Mapping of a gene responsible for the difference in amylopectin structure between japonica-type and indica-type rice varieties. Theor. Appl. Genet. 2002, 104, 1–8. [Google Scholar] [CrossRef]
- Kawagoe, Y.; Kubo, A.; Satoh, H.; Takaiwa, F.; Nakamura, Y. Roles of isoamylase and ADP-glucose pyrophosphorylase in starch granule synthesis in rice endosperm. Plant J. 2005, 42, 164–174. [Google Scholar] [CrossRef]
- Edwards, D.; Batley, J. Plant genome sequencing: Application for crop improvement. Plant Biotechnol. J. 2010, 8, 2–9. [Google Scholar] [CrossRef]
- Neeraja, C.N.; Maghirang-Rodriguez, R.; Pamplona, A.; Heuer, S.; Collard, B.C.Y.; Septiningsih, E.M.; Vergara, G.; Sanchez, D.; Xu, K.; Ismail, A.M.; et al. A marker-assisted backcross approach for developing submergence-tolerant rice cultivars. Theor. Appl. Genet. 2007, 115, 757–776. [Google Scholar] [CrossRef] [PubMed]
- Neelam, K.; Brown-Guedira, G.; Huang, L. Development and validation of a breeder-friendly KASPar marker for wheat leaf rust resistance locus Lr21. Mol. Breed. 2013, 31, 233–237. [Google Scholar] [CrossRef]
- Kumpatla, S.P.; Buyyarapu, R.; Abdurakhmonov, I.Y.; Mammadov, J.A. Genomics-Assisted Plant Breeding in the 21st Century: Technological Advances and Progress. In Plant Breeding; Abdurakhmonov, I., Ed.; IntechOpen: London, UK, 2012; Available online: http://www.intechopen.com/books/plantbreeding/genomics-assisted-plant-breeding-in-the-21stcenturytechnological-advances-and-progress (accessed on 13 January 2021).
- Kumar, R.; Khatkar, B.S. Thermal, pasting and morphological properties of starch granules of wheat (Triticum aestivum L.) varieties. J. Food Sci. Technol. 2017, 54, 2403–2410. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Won, Y.; Cho, Y.; Lee, J.; Yang, C.; Kim, M.; Ahn, E.; Suh, J.; Lee, S.; Jeon, Y.; et al. ‘Samkwang1’, a Medium Maturing, Multiple Disease Resistant, and High-quality Rice. Korean J. Breed. Sci. 2018, 50, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Suzuki, Y.; Okuno, K.; Hirano, H.; Imbe, T. Genetic analysis of low-amylose content in a rice variety, ‘Milky Queen’. Breed. Res. 2001, 3, 13–19. [Google Scholar] [CrossRef]
- Rural Development Administration (RDA). Standard of Analysis and Survey for Agricultural Research; Rural Development Administration (RDA): Jeonju, Korea, 2012; pp. 315–338. [Google Scholar]
- Nogoy, F.M.; Jung, Y.J.; Kang, K.K.; Cho, Y.G. Characterization of ‘GolSam’ Lines Developed from the Cross between Samgwang and 5MT Resistant Lines in Rice. Plant Breed. Biotech. 2018, 6, 233–244. [Google Scholar] [CrossRef]
- Osborne, B.G. Near-infrared Spectroscopy in Food Analysis. Encycl. Anal. Chem. 2006, 1–14. [Google Scholar] [CrossRef]
- Kim, C.E.; Kang, M.Y.; Kim, M.H. Comparison of properties affecting the palatability of 33 commercial brands of rice. Korean J. Crop Sci. 2014, 57, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Chi, X.; Zhang, Y.; Xue, Z.; Feng, L.; Liu, H.; Wang, F.; Qi, X. Discovery of rare mutations in extensively pooled DNA samples using multiple target enrichment. Plant Biotechnol. J. 2014, 12, 709–717. [Google Scholar] [CrossRef]
- Park, H.S.; Lee, K.M.; Kim, K.Y.; Kim, J.J.; Shin, W.C.; Baek, M.K.; Kim, C.S.; Park, S.G.; Lee, C.M.; Suh, J.P.; et al. Development of near- Isogenic line of japonica rice cultivar Saenuri without lipoxygenase-3. Korean J. Breed. Sci. 2019, 51, 190–200. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Hardness | Adhesiveness | Springiness | Stickiness |
|---|---|---|---|---|
| Koshihikari | 31.75 ± 5.73 | 63.44 ± 2.37 | 34.56 ± 4.26 | 69.56 ± 3.49 |
| Samgwang | 36.03 ± 0.26 | 55.74 ± 2.47 | 30.92 ± 0.94 | 48.01 ± 3.42 |
| Baegjinju | 24.07 ± 3.68 | 60.94 ± 5.73 | 26.35 ± 2.95 | 76.10 ± 3.25 |
| Milkyqueen | 21.17 ± 4.36 | 57.90 ± 3.36 | 26.21 ± 1.82 | 60.03 ± 4.85 |
| F-value | 8.572 ** | 1.094 | 6.233 * | 3.099 |
| Variety | GT * (°C) | Viscosity (RVU) | ||||
|---|---|---|---|---|---|---|
| PV § | HPV ¶ | Breakdown | CPV † | Setback | ||
| Koshihikari | 72.78 ± 0.70 | 392.42 ± 9.90 | 175.79 ± 15.18 | 227.04 ± 11.46 | 265.92 ± 11.71 | −136.92 ± 9.44 |
| Samgwang | 72.24 ± 0.34 | 269.08 ± 4.92 | 168.14 ± 6.74 | 112.59 ± 18.86 | 265.98 ± 9.33 | −19.75 ± 13.73 |
| Baegjinju | 69.43 ± 0.45 | 172.78 ± 8.46 | 127.25 ± 5.78 | 127.25 ± 5.78 | 77.64 ± 4.64 | −95.14 ± 3.97 |
| Milkyqueen | 71.43 ± 0.11 | 299.15 ± 8.68 | 208.83 ± 8.68 | 90.32 ± 19.25 | 294.33 ± 7.95 | −14.42 ± 13.27 |
| F-value | 38.831 *** | 147–951 ** | 51.268 *** | 59.717 *** | 365.712 ** | 108.545 *** |
| Population. | Generation | Number of Plants | χ2 Value (1:1) | ||
|---|---|---|---|---|---|
| Total | Homo (G/G) | Hetero (G/A) | |||
| Samgwang × Milkyqueen | BC1F1 | 210 | 110 | 100 | 0.48 ns |
| BC2F1 | 196 | 94 | 102 | 0.33 ns | |
| Population | Line | Plant Height (cm) | Culm Length (cm) | Panicle Length (cm) | No. of Tillers |
|---|---|---|---|---|---|
| Parent | Samgwang | 95.2 ± 4.6 | 64.8 ± 6.0 | 19.7 ± 1.5 | 9 ± 3 |
| Milkyqueen | 92.6 ± 2.9 | 69.9 ± 5.4 | 18.4 ± 0.4 | 10 ± 2 | |
| BC1F1 | SM-30 | 95.8 | 71.0 | 22.2 | 9 |
| SM-43 | 93.8 | 72.4 | 18.4 | 7 | |
| SM-46 | 101.6 | 72.2 | 18.4 | 16 | |
| SM-79 | 90.2 | 64.0 | 20.4 | 6 | |
| SM-82 | 91.8 | 68.0 | 20.6 | 7 | |
| SM-85 | 91.4 | 71.6 | 21.8 | 9 | |
| SM-88 | 92.6 | 64.0 | 20.8 | 6 |
| Population | Line | Plant Height (cm) | Culm Length (cm) | Panicle Length (cm) | No. of Tillers | Amylose Content(%) |
|---|---|---|---|---|---|---|
| Parent | Samgwang | 112.3 ± 4.6 | 85.6 ± 2.4 | 21.9 ± 1.6 | 10.6 ± 1.8 | 18.5 ± 2.1 |
| Milkyqueen | 102.2 ± 3.0 | 74.6 ± 3.7 | 16.4 ± 1.1 | 8 ± 1.2 | 12.6 ± 1.3 | |
| BC2F2 | SM35-3 | 100.2 | 83.6 | 19 | 10 | 12.4 ± 5.2 |
| SM35-8 | 114.6 | 85.6 | 19.8 | 10 | 16.8 ± 5.2 | |
| SM35-22 | 114.2 | 86 | 17.6 | 11 | 14.2 ± 2.2 | |
| SM35-35 | 113.6 | 84.2 | 18 | 13 | 15.9 ± 7.5 | |
| SM35-36 | 116 | 82.9 | 17.4 | 13 | 15.1 ± 6.6 | |
| SM35-54 | 112.2 | 88.8 | 17.6 | 11 | 14.1 ± 4.7 | |
| SM35-60 | 115.2 | 85.6 | 18.2 | 14 | 14.2 ± 4.2 |
| GT* (°C) | Viscosity (RVU) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Population | PV § | HPV ¶ | Breakdown | CPV † | Setback | |||||||
| Mean ± SD | t | Mean ± SD | t | Mean ± SD | t | Mean ± SD | t | Mean ± SD | t | Mean ± SD | t | |
| Samgwang | 72.5 ± 0.4 | −6.22 ** | 228.4 ± 9.5 | 3.396 ** | 139.0 ± 9.7 | −1.114 ns | 94.2 ± 4.9 | 10.666 *** | 210.9 ± 5.7 | −5.399 *** | −17.4 ± 3.7 | −21.986 *** |
| Milkyqueen | 76.6 ± 1.04 | 205.9 ± 6.2 | 146.5 ± 6.1 | 59.4 ± 2.7 | 240.4 ± 7.4 | 34.4 ± 1.5 | ||||||
| SM35-02 | 70.80 | 210.25 | 124.50 | 85.75 | 210.42 | 0.17 | ||||||
| SM35-03 | 70.73 | 206.09 | 125.96 | 80.13 | 204.21 | −1.88 | ||||||
| SM35-05 | 71.25 | 246.42 | 135.17 | 111.25 | 216.63 | −29.79 | ||||||
| SM35-08 | 73.63 | 249.92 | 136.38 | 113.54 | 217.29 | −32.63 | ||||||
| SM35-20 | 72.83 | 245.88 | 137.17 | 108.71 | 222.04 | −23.84 | ||||||
| SM35-21 | 72.43 | 232.00 | 139.79 | 92.21 | 222.80 | −9.21 | ||||||
| SM35-31 | 72.15 | 241.38 | 136.50 | 104.88 | 215.75 | −25.63 | ||||||
| SM35-35 | 71.53 | 244.58 | 142.96 | 101.63 | 224.00 | −20.59 | ||||||
| SM35-36 | 72.20 | 245.25 | 144.04 | 101.21 | 224.00 | −21.25 | ||||||
| SM35-39 | 72.15 | 225.92 | 144.50 | 81.42 | 231.92 | 6.00 | ||||||
| SM35-40 | 72.10 | 237.25 | 155.67 | 81.58 | 238.17 | 0.92 | ||||||
| SM35-54 | 71.93 | 239.84 | 140.88 | 98.96 | 224.96 | −14.88 | ||||||
| SM35-55 | 72.08 | 226.50 | 126.25 | 100.25 | 205.04 | −21.46 | ||||||
| SM35-60 | 72.13 | 228.88 | 131.88 | 97.00 | 210.17 | −18.71 | ||||||
| SM35-62 | 71.40 | 232.17 | 139.50 | 92.67 | 221.08 | −11.08 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-S.; Yang, J.-Y.; Yu, J.-K.; Lee, Y.; Park, Y.-J.; Kang, K.-K.; Cho, Y.-G. Breeding of High Cooking and Eating Quality in Rice by Marker-Assisted Backcrossing (MABc) Using KASP Markers. Plants 2021, 10, 804. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040804
Kim M-S, Yang J-Y, Yu J-K, Lee Y, Park Y-J, Kang K-K, Cho Y-G. Breeding of High Cooking and Eating Quality in Rice by Marker-Assisted Backcrossing (MABc) Using KASP Markers. Plants. 2021; 10(4):804. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040804
Chicago/Turabian StyleKim, Me-Sun, Ju-Young Yang, Ju-Kyung Yu, Yi Lee, Yong-Jin Park, Kwon-Kyoo Kang, and Yong-Gu Cho. 2021. "Breeding of High Cooking and Eating Quality in Rice by Marker-Assisted Backcrossing (MABc) Using KASP Markers" Plants 10, no. 4: 804. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040804