
Genome-Wide Association Study Reveals the QTLs for Seed Storability in World Rice Core Collections


Abstract
:1. Introduction
2. Results
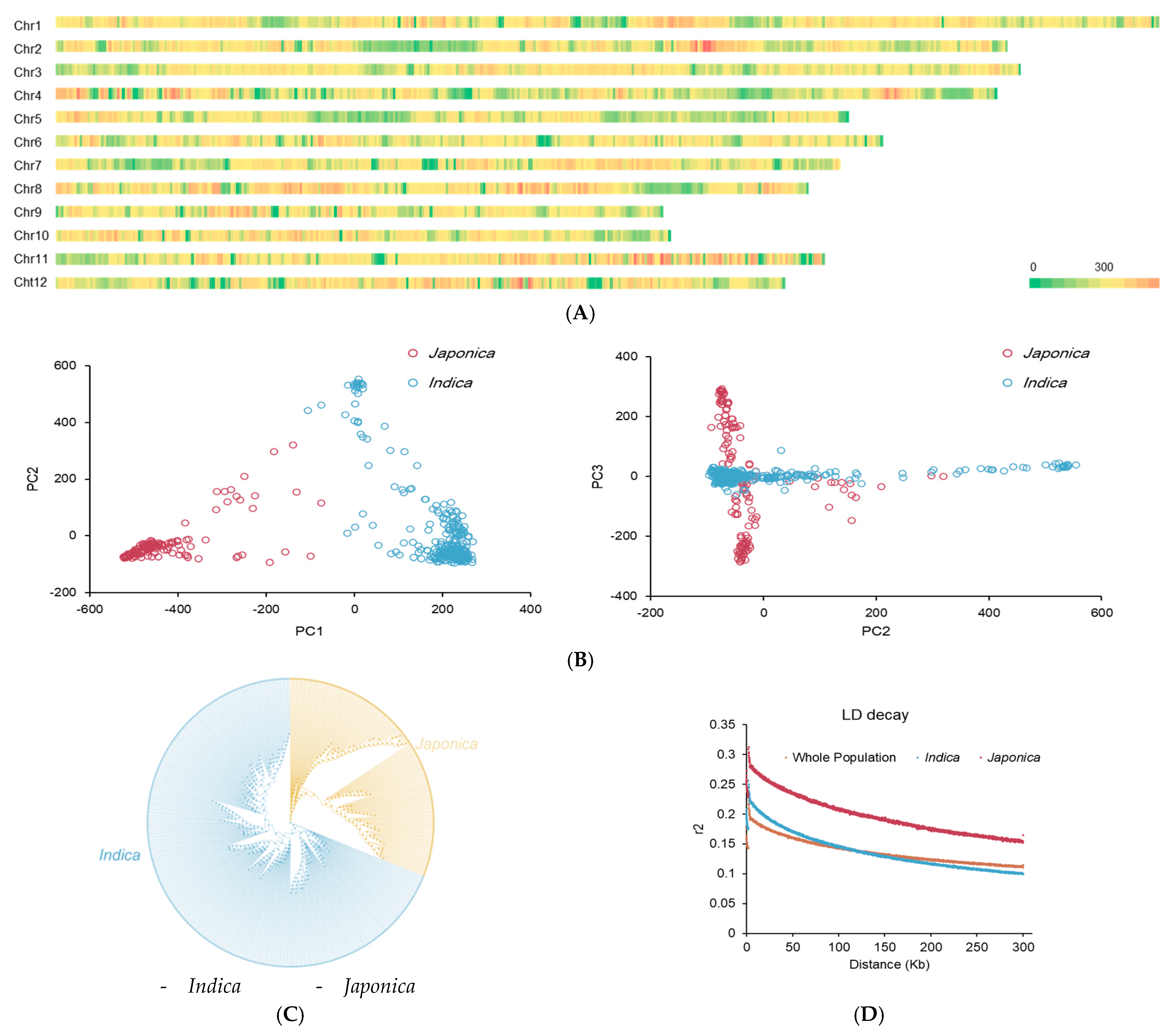
2.1. Population Structure and Phenotypic Evaluation of Seed Storability in Rice
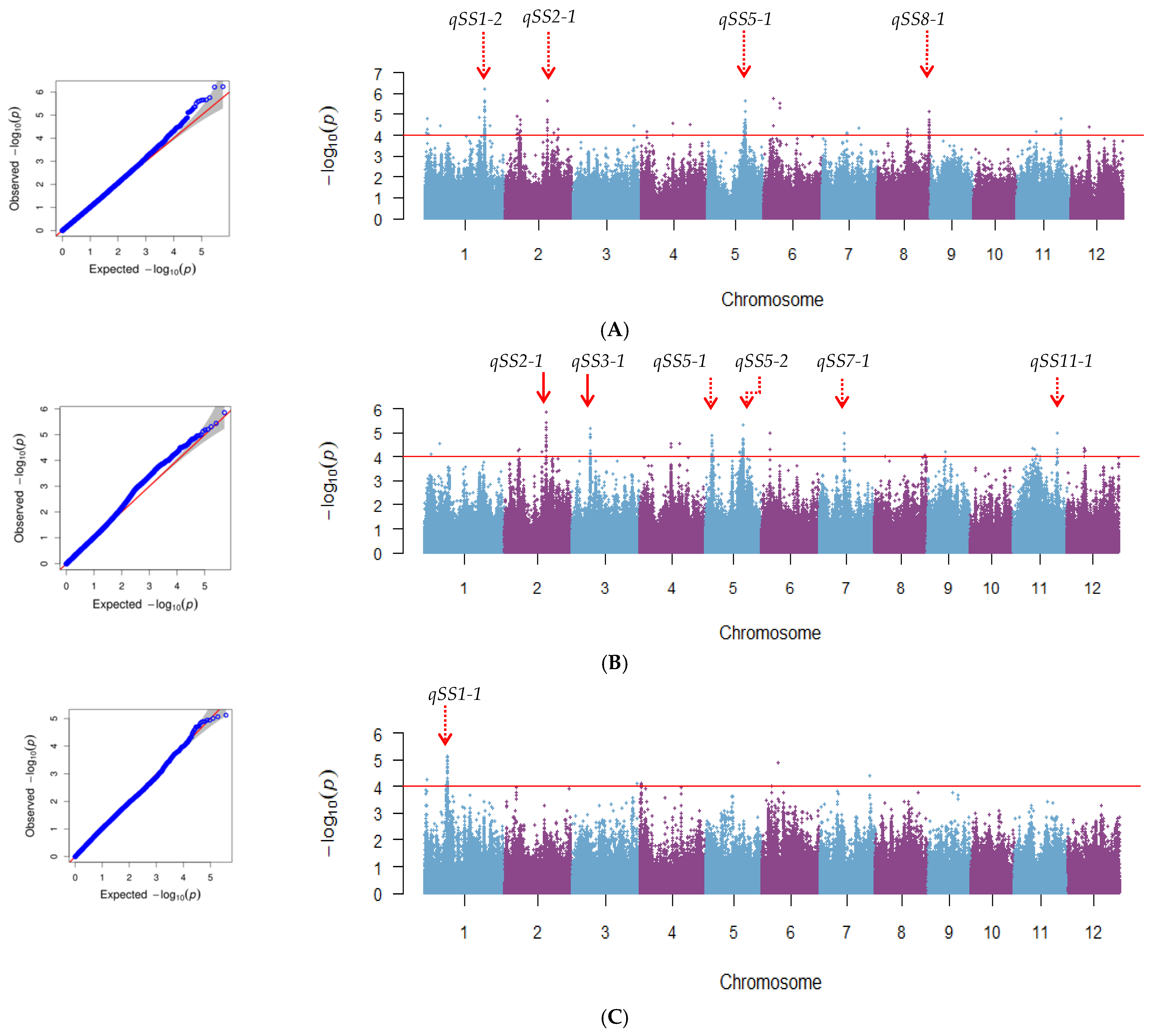
2.2. Identification of Nine New QTLs for Seed Storability by GWAS in Rice
2.3. Pyramiding Analysis of These QTLs for Rice Seed Storability
3. Discussion
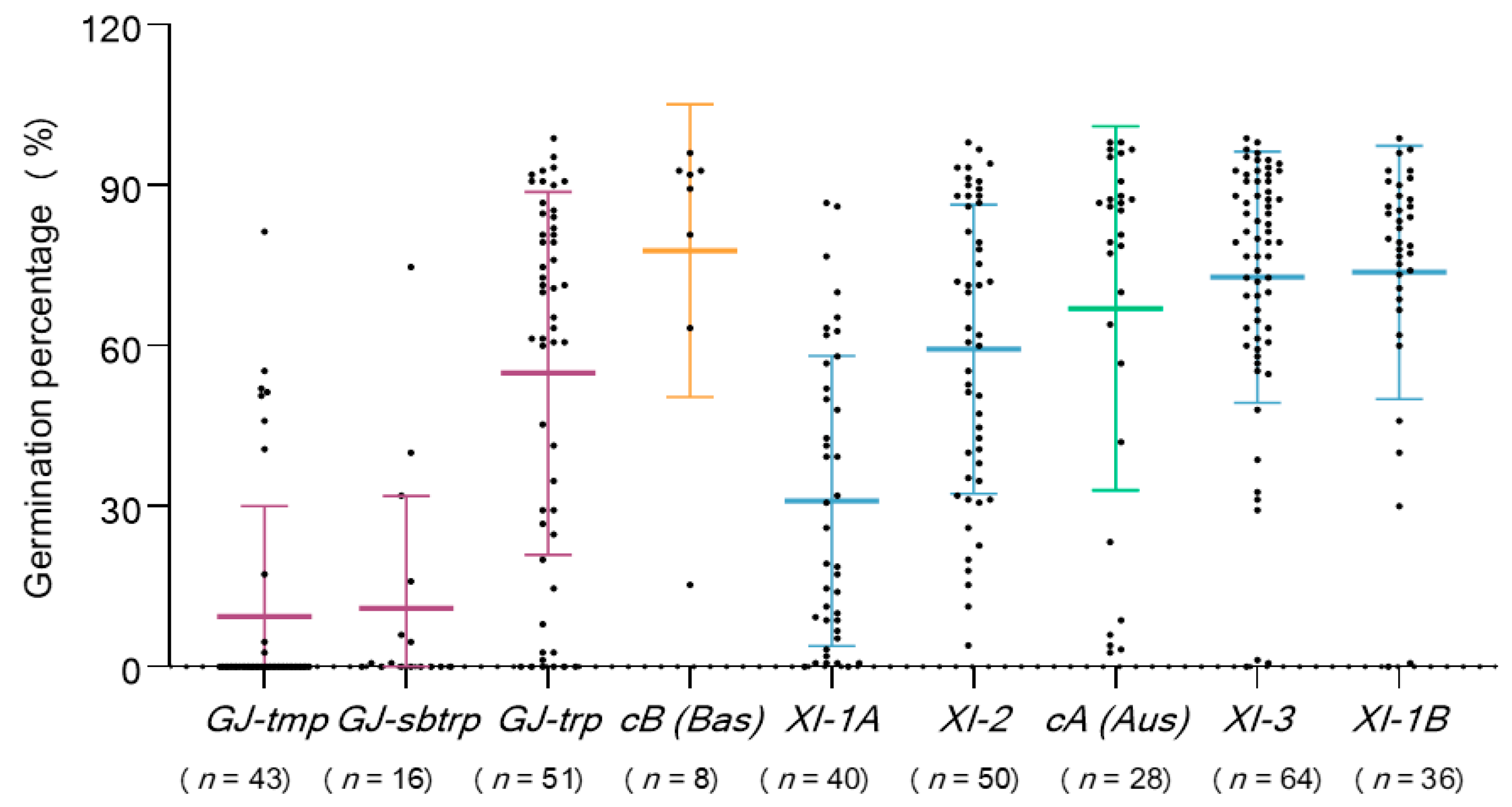
3.1. Seed Storability Variations between Indica and Japonica Subspecies in Rice
3.2. GWAS for Seed Storability by Using High-Throughput Genotyping in Rice
3.3. Exposing the Difference of the Two Subspecies for Rice Seed Storability by Analysis Superior Allele Frequency of the QTLs
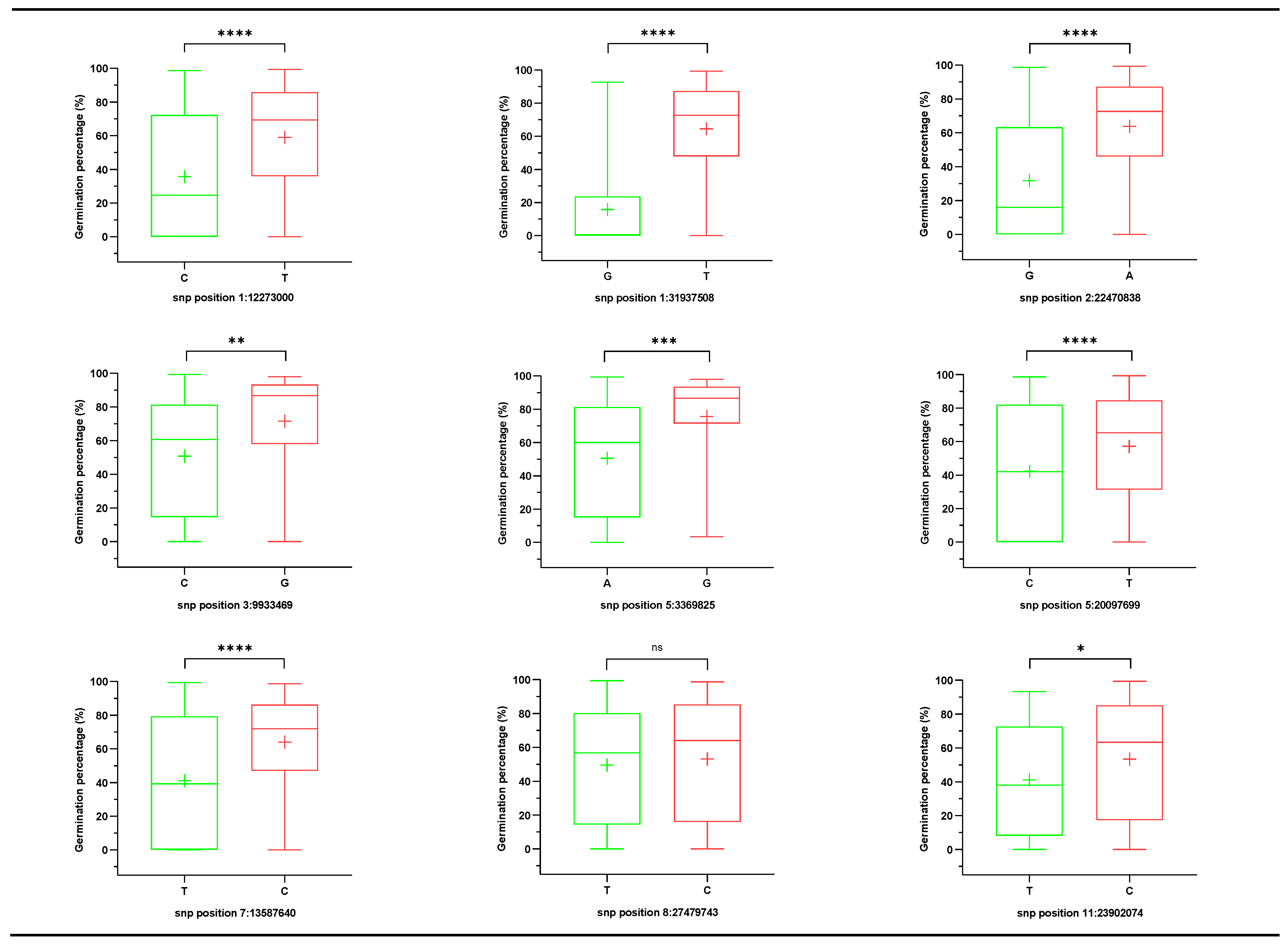
3.4. New SNPs Participate in the Seed Storability in Rice
4. Materials and Methods
4.1. Plant Materials
4.2. Artificial Aging Treatment
4.3. Seed Germination Test
4.4. Genotyping and GWAS for Rice Seed Storability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, H.; Wei, Y.; Zhu, Y.; Lian, L.; Xie, H.; Cai, Q.; Chen, Q.; Lin, Z.; Wang, Z.; Xie, H.; et al. Antisense suppression of LOX3 gene expression in rice endosperm enhances seed longevity. Plant Biotech. J. 2015, 13, 526–539. [Google Scholar] [CrossRef] [PubMed]
- Siddique, S.B.; Seshu, D.V.; Pardee, W.D. Rice cultivars variability in tolerance for accelerated aging of seed [Philippines]. IRRI Res. Pap. Ser. 1988, 131, 2–7. [Google Scholar]
- Rajjou, L.; Debeaujon, I. Seed longevity: Survival and maintenance of high germination ability of dry seeds. C. R. Biol. 2008, 331, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Hang, N.T.; Lin, Q.; Liu, L.; Liu, X.; Liu, S.; Wang, W.; Li, L.; He, N.; Liu, Z.; Jiang, L.; et al. Mapping QTLs related to rice seed storability under natural and artificial aging storage conditions. Euphytica 2015, 203, 673–681. [Google Scholar] [CrossRef]
- Rao, N.K.; Jackson, M.T. Variation in seed longevity of rice cultivars belonging to different isozyme groups. Genet. Resour. Crop Evol. 1997, 44, 159–164. [Google Scholar]
- Wu, F.; Zhu, Y.; Xie, H.; Zhang, J.; Xie, H. Preliminary study on storability of Chinese micro-core collections of rice. J. Chin. Cereals Oils Assoc. 2010, 25, 124–128. [Google Scholar]
- Lee, J.S.; Velasco-Punzalan, M.; Pacleb, M.; Valdez, R.; Kretzschmar, T.; McNally, K.L.; Ismail, A.M.; Cruz, P.C.S.; Sackville Hamilton, N.R. Variation in seed longevity among diverse indica rice varieties. Ann. Bot. 2019, 124, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Fukuta, Y.; Sato, T. Mapping of quantitative trait loci controlling seed longevity of rice (Oryza sativa L.) after various periods of seed storage. Plant Breed. 2005, 124, 361–366. [Google Scholar] [CrossRef]
- Jiang, W.; Lee, J.; Jin, Y.M.; Qiao, Y.; Piao, R.; Jang, S.M.; Woo, M.O.; Kwon, S.W.; Liu, X.; Pan, H.Y.; et al. Identification of QTLs for seed germination capability after various storage periods using two RIL, populations in rice. Mol. Cells 2011, 31, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lin, Q.; Liu, S.; Liu, X.; Wang, W.; Hang, N.; Liu, F.; Zhao, Z.; Jiang, L.; Wan, J. Identification of quantitative trait loci for seed storability in rice (Oryza sativa L.). Plant Breed. 2012, 131, 739–743. [Google Scholar] [CrossRef]
- Miura, K.; Lin, S.; Yano, M.; Nagamine, T. Mapping quantitative trait loci controlling seed longevity in rice (Oryza sativa L.). Theor. Appl. Genet. 2002, 104, 981–986. [Google Scholar] [CrossRef]
- Zeng, D.; Guo, L.; Xu, Y.; Yasukumi, K.; Zhu, L.; Qian, Q. QTL analysis of seed storability in rice. Plant Breed. 2006, 125, 57–60. [Google Scholar] [CrossRef]
- Xue, Y.; Zhang, S.; Yao, Q.; Peng, R.; Xiong, A.; Li, X.; Zhu, W.; Zhu, Y.; Zha, D. Identification of quantitative trait loci for seed storability in rice (Oryza sativa L.). Euphytica 2008, 164, 739–744. [Google Scholar] [CrossRef]
- Lin, Q.; Wang, W.; Ren, Y.; Jiang, Y.; Sun, A.; Qian, Y.; Zhang, Y.; He, N.; Hang, N.T.; Liu, Z.; et al. Genetic dissection of seed storability using two different populations with a same parent rice cultivar N22. Breed. Sci. 2015, 65, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Shao, G.; Wang, L.; Wang, Z.; Mao, Y.; Wang, X.; Zhang, X.; Liu, S.; Zhang, H. QTL identification and fine mapping for seed storability in rice (Oryza sativa L.). Euphytica 2017, 213, 12. [Google Scholar] [CrossRef]
- Yuan, Z.; Fan, K.; Xia, L.; Ding, X.; Tian, L.; Sun, W.; He, H.; Yu, S. Genetic dissection of seed storability and validation of candidate gene associated with antioxidant capability in rice (Oryza sativa L.). Int. J. Mol. Sci. 2019, 20, 4442. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Kim, S.R.; An, G. Rice aldehyde dehydrogenase7 is needed for seed maturation and viability. Plant Physiol. 2009, 149, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Cai, M.; Long, Q.; Liu, L.; Lin, Q.; Jiang, L.; Chen, S.; Wan, J. OsLOX2, a rice type I lipoxygenase, confers opposite effects on seed germination and longevity. Transgenic Res. 2014, 23, 643–655. [Google Scholar] [CrossRef]
- Wei, Y.; Xu, H.; Diao, L.; Zhu, Y.; Xie, H.; Cai, Q.; Wu, F.; Wang, Z.; Zhang, J.; Xie, H. Protein repair L-isoaspartylmethyltransferase 1 (PIMT1) in rice improves seed longevity by preserving embryo vigor and viability. Plant Mol. Biol. 2015, 89, 475–492. [Google Scholar] [CrossRef] [PubMed]
- Petla, B.P.; Kamble, N.U.; Kumar, M.; Verma, P.; Ghosh, S.; Singh, A.; Rao, V.; Salvi, P.; Kaur, H.; Saxena, S.C.; et al. Rice proteinl-isoaspartyl methyltransferase isoforms differentially accumulate during seed maturation to restrict deleterious isoAsp and reactive oxygen species accumulation and are implicated in seed vigor and longevity. New Phytol. 2016, 211, 627–645. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Chen, D.; Ren, Y.; Zhang, X.; Zhao, J. Characteristic and Expression Analysis of a Metallothionein Gene, OsMT2b, Down-Regulated by Cytokinin Suggests Functions in Root Development and Seed Embryo Germination of Rice. Plant Physiol. 2008, 146, 1637–1650. [Google Scholar] [CrossRef]
- Yuan, Z.; Fan, K.; Wang, Y.; Tian, L.; Zhang, C.; Sun, W.; He, H.; Yu, S. OsGRETCHENHAGEN3-2 modulates rice seed storability via accumulation of abscisic acid and protective substances. Plant Physiol. 2021. [Google Scholar] [CrossRef]
- Shakiba, E.; Edwards, J.D.; Jodari, F.; Duke, S.E.; Baldo, A.M.; Korniliev, P.; McCouch, S.R.; Eizenga, G.C. Genetic architecture of cold tolerance in rice (Oryza sativa) determined through high resolution genome-wide analysis. PLoS ONE 2017, 12, e0172133. [Google Scholar] [CrossRef]
- Wang, X.; Zou, B.; Shao, Q.; Cui, Y.; Lu, S.; Zhang, Y.; Huang, Q.; Huang, J.; Hua, J. Natural variation reveals that OsSAP16 controls low-Temperature germination in rice. J. Exp. Bot. 2018, 69, 413–421. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, W.; Zhang, S.; Yang, T.; Liu, Q.; Dong, J.; Fu, H.; Mao, X.; Liu, B. Genome-wide association study and candidate gene analysis of rice cadmium accumulation in grain in a diverse rice collection. Rice 2018, 11, 61. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, D.; Yan, S.; Liu, S.; Liu, B.; Kang, H.; Wang, G.L. Dissection of the genetic architecture of rice tillering using a genome-wide association study. Rice 2019, 43, 1–11. [Google Scholar] [CrossRef]
- Kameswara, N.; Jackson, M.T. Seed longevity of environment and storage longevity of japonica rices (Oryza sativa L.). Seed Sci. Res. 1996, 6, 17–21. [Google Scholar] [CrossRef]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F.; et al. Genomic variation in 3,010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef]
- Dong, X.; Fan, S.; Liu, J.; Wang, Q.; Li, M.; Jiang, X.; Liu, Z.; Yin, Y.; Wang, J. Identification of QTLs for seed storability in rice under natural aging conditions using two RILs with the same parent Shennong 265. J. Integr. Agric. 2017, 16, 1084–1092. [Google Scholar] [CrossRef]
- Liu, W.; Pan, X.; Li, Y.; Duan, Y.; Min, J.; Liu, S.; Sheng, X.; Li, X. Detection and validation of QTLs associated with seed longevity in rice (Oryza sativa L.). Plant Breed. 2018, 137, 546–552. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Vera Cruz, C.M.; McNally, K.L.; Virk, P.S.; Mackill, D.J. Rice molecular breeding laboratories in the genomics era: Current status and future considerations. Int. J. Plant Genom. 2008. [Google Scholar] [CrossRef]
- Famoso, A.N.; Zhao, K.; Clark, R.T.; Tung, C.W.; Wright, M.H.; Bustamante, C.; Kochian, L.V.; McCouch, S.R. Genetic architecture of aluminum tolerance in rice (Oryza Sativa) determined through genome-wide association analysis and QTL mapping. PLoS Genet. 2011, 7, e1002221. [Google Scholar] [CrossRef] [PubMed]
- Al-Tamimi, N.; Brien, C.; Oakey, H.; Berger, B.; Saade, S.; Ho, Y.S.; Schmockel, S.M.; Tester, M.; Negrao, S. Salinity tolerance loci revealed in rice using high-throughput non-invasive phenotyping. Nat. Commun. 2016, 7, 13342. [Google Scholar] [CrossRef]
- McCouch, S.R.; Wright, M.H.; Tung, C.W.; Maron, L.G.; McNally, K.L.; Fitzgerald, M.; Singh, N.; DeClerck, G.; Agosto-Perez, F.; Korniliev, P.; et al. Open access resources for genome-wide association mapping in rice. Nat. Commun. 2016, 7, 10532. [Google Scholar] [CrossRef]
- Zeng, D.; Qian, Q.; Yasukumi, K. Study on storability and morphological index in rice (Oryza sativa L.) under artificial ageing. Acta Agron. Sin. 2002, 28, 551–554. [Google Scholar]
- Alexandrov, N.; Tai, S.; Wang, W.; Mansueto, L.; Palis, K.; Fuentes, R.R.; Ulat, V.J.; Chebotarov, D.; Zhang, G.; Li, Z.; et al. SNP-seek database of SNPs derived from 3000 rice genomes. Nucleic Acids Res. 2015, 43, D1023. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. Plink: A tool set for whole-genome association and population based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Zhang, C.; Dong, S.; Xu, J.; He, W.; Yang, T. PopLDdecay: A fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 2019, 35, 1786–1788. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, X.; Wang, J.; Li, M.; Wang, Q.; Tian, F.; Su, Z.; Pan, Y.; Liu, D.; Lipka, A.E.; et al. GAPIT Version 2: An Enhanced Integrated Tool for Genomic Association and Prediction. Plant Genome 2016, 9, 1–9. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QTL Name | Population | Chr. | The Interval of the Significant SNP | Number of the Significant SNP | Lead SNP Position (bp) | -LOG(P) of Lead SNP | MAF (%) |
|---|---|---|---|---|---|---|---|
| qSS1-2 | Whole population | 1 | 31854852–32063190 | 26 | 31937508 | 6.22 | 0.27 |
| qSS2-1 | Whole population | 2 | 22405984–22470838 | 7 | 22470838 | 5.65 | 0.37 |
| qSS5-2 | Whole population | 5 | 19789530–20161102 | 26 | 20097699 | 5.66 | 0.36 |
| qSS8-1 | Whole population | 8 | 27476715–27509284 | 9 | 27479743 | 5.11 | 0.33 |
| qSS2-1 | Indica | 2 | 22405984–22590272 | 15 | 22470838 | 5.93 | 0.12 |
| qSS3-1 | Indica | 3 | 9908667–9943096 | 21 | 9933469 | 5.28 | 0.08 |
| qSS5-1 | Indica | 5 | 3248113–3450060 | 19 | 3369825 | 4.97 | 0.08 |
| qSS5-2 | Indica | 5 | 20007577–20170795 | 34 | 20097699 | 5.33 | 0.1 |
| qSS7-1 | Indica | 7 | 13568523–13590543 | 4 | 13587640 | 5.03 | 0.34 |
| qSS11-1 | Indica | 11 | 23865348–23902074 | 3 | 23902074 | 5.01 | 0.14 |
| qSS1-1 | Japonica | 1 | 12113558–12273000 | 15 | 12273000 | 4.8 | 0.14 |
| Chr. | QTL Name | Position of the Lead SNP | Effect Value (%) | ||
|---|---|---|---|---|---|
| Whole Population | Indica Subspecies | Japonica Subspecies | |||
| 1 | qSS1-1 | 12273000 | 6.36 | 0.46 | 22.97 |
| 1 | qSS1-2 | 31937508 | 12.19 | 9.72 | 15.30 |
| 2 | qSS2-1 | 22470838 | 11.43 | 12.99 | 5.62 |
| 3 | qSS3-1 | 9933469 | 10.25 | 32.48 | 15.70 |
| 5 | qSS5-1 | 3369825 | 17.51 | 24.38 | 1.71 |
| 5 | qSS5-2 | 20097699 | 16.45 | 17.98 | 14.31 |
| 7 | qSS7-1 | 13587640 | 7.87 | 8.60 | 2.05 |
| 8 | qSS8-1 | 27479743 | 7.91 | 6.80 | 15.61 |
| 11 | qSS11-1 | 23902074 | 8.37 | 10.72 | 0.33 |
| Groups | Subspecies | No. of Varieties | No. Superior SNPs | The Average Germination Percentage (%) | Genotype | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| qSS 1-1 | qSS 1-2 | qSS 2-1 | qSS 3-1 | qSS 5-1 | qSS 5-2 | qSS 7-1 | qSS 8-1 | qSS 11-1 | |||||
| 1 | Indica | 1 | 9 | 98 | + | + | + | + | + | + | + | + | + |
| 2 | Indica | 2 | 8 | 92 | + | + | + | + | + | - | + | + | + |
| 3 | Indica | 59 | 7 | 72.77 | + | + | + | - | - | + | + | + | + |
| 4 | Indica | 16 | 6 | 77.96 | + | + | + | - | - | + | - | + | + |
| 5 | Indica | 49 | 6 | 61.74 | + | + | + | - | - | + | + | - | + |
| 6 | Indica | 12 | 5 | 71.33 | - | + | + | - | - | + | + | - | + |
| 7 | Indica | 27 | 5 | 60.94 | + | + | + | - | - | + | - | - | + |
| 8 | Indica | 7 | 4 | 34.76 | + | + | + | - | - | + | - | - | - |
| 9 | Japonica | 2 | 5 | 91 | - | + | + | - | + | - | - | + | + |
| 10 | Japonica | 1 | 5 | 80.67 | + | + | + | - | - | - | - | + | + |
| 11 | Japonica | 10 | 4 | 39.67 | + | + | - | - | - | - | - | + | + |
| 12 | Japonica | 31 | 3 | 73.12 | - | + | - | - | - | - | - | + | + |
| 13 | Japonica | 70 | 2 | 15.58 | - | - | - | - | - | - | - | + | + |
| 14 | Japonica | 8 | 1 | 10.75 | - | - | - | - | - | - | - | - | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Luo, X.; Wang, L.; Wei, Y.; Li, J.; Xie, H.; Zhang, J.; Xie, G. Genome-Wide Association Study Reveals the QTLs for Seed Storability in World Rice Core Collections. Plants 2021, 10, 812. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040812
Wu F, Luo X, Wang L, Wei Y, Li J, Xie H, Zhang J, Xie G. Genome-Wide Association Study Reveals the QTLs for Seed Storability in World Rice Core Collections. Plants. 2021; 10(4):812. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040812
Chicago/Turabian StyleWu, Fangxi, Xi Luo, Lingqiang Wang, Yidong Wei, Jianguo Li, Huaan Xie, Jianfu Zhang, and Guosheng Xie. 2021. "Genome-Wide Association Study Reveals the QTLs for Seed Storability in World Rice Core Collections" Plants 10, no. 4: 812. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10040812