
Importance of Landraces in Cereal Breeding for Stress Tolerance

 , , and
, , and
Abstract
:1. Introduction
2. Conservation ‘In Situ’ and ‘Ex Situ’ of the Main Cereal Collections and Availability of Databases
3. Exploration of Genetic Diversity and Population Structure
4. Landraces as a Genetic Resource to Identify the Genetic Factors Responsible for Resistance to Biotic and Abiotic Stresses
4.1. Abiotic Stresses and Traits of Agronomic Importance in Limiting Environments
4.2. Biotic Stresses
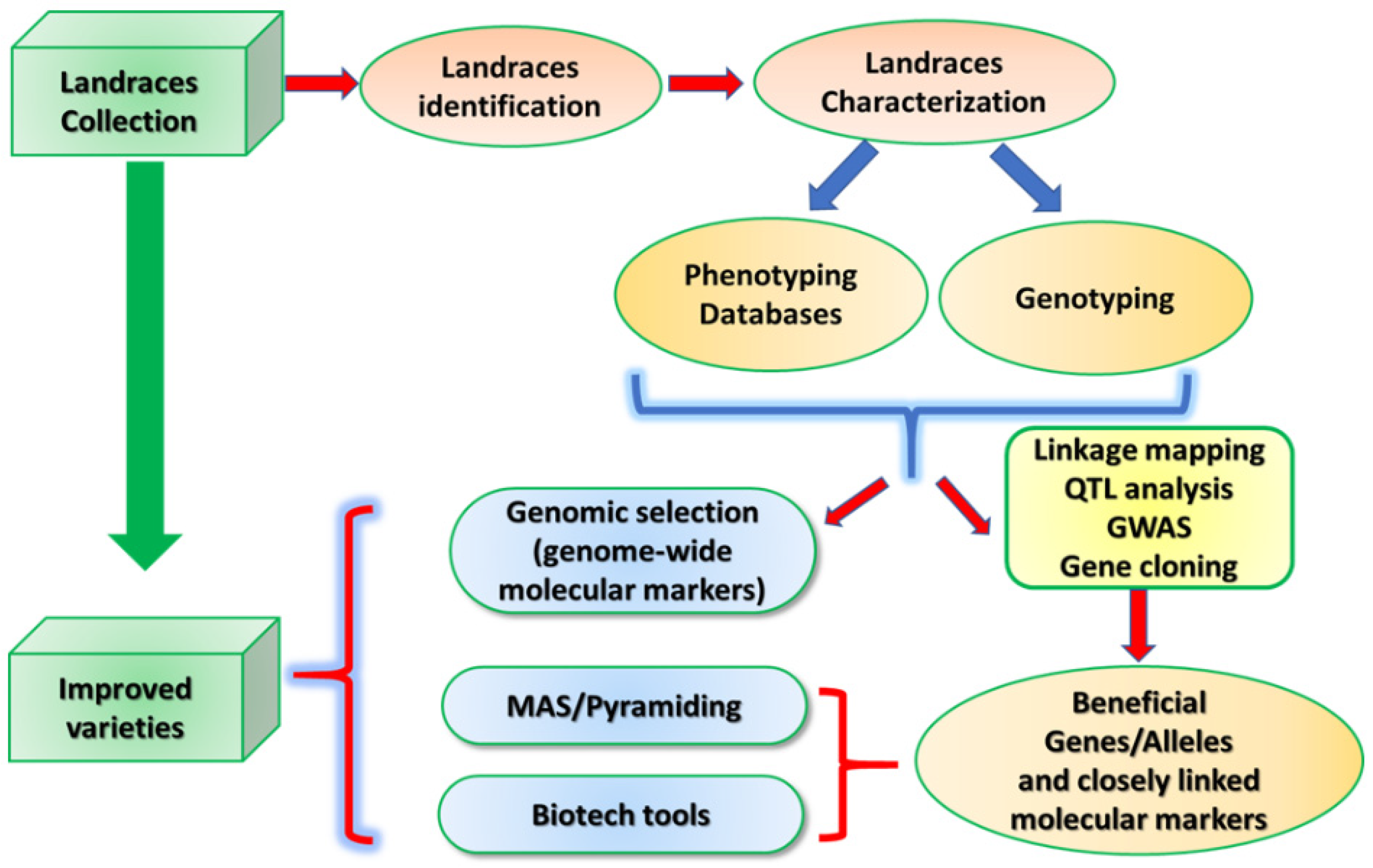
5. Use of Landraces in Cereal Breeding
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, A.H.D. Isozymes, plant population genetic structure and genetic conservation. Theor. Appl. Genet. 1978, 52, 145–157. [Google Scholar] [CrossRef]
- Harlan, J.R. Our vanishing genetic resources. Science 1975, 188, 618–621. [Google Scholar] [CrossRef]
- Hawkes, J.G. The Diversity of Crop Plants; Harvard University Press: Cambridge, MA, USA, 1983; p. 184. [Google Scholar]
- Camacho-Villa, T.C.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Res. 2005, 3, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace germplasm for improving yield and abiotic stress adaptation. Trends Plant Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Zeven, A.C. Landraces: A review of definitions and classifications. Euphytica 1998, 104, 127–139. [Google Scholar] [CrossRef]
- Bellon, M.R.; Brush, S. Keepers of maize in Chiapas, Mexico. Econ. Bot. 1994, 48, 196–209. [Google Scholar] [CrossRef]
- Wood, D.; Lenné, J.M. The conservation of agrobiodiversity on-farm: Questioning the emerging paradigm. Biodivers. Conserv. 1997, 6, 109–129. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.H.D. The genetic structure of crop landraces and the challenge to conserve them in situ on farms. In Genes in the Field: On-Farm Conservation of Crop Diversity, 1st ed.; Brush, S.B., Ed.; Lewis Publishers: Boca Raton, FL, USA, 2000; pp. 29–48. [Google Scholar]
- Ceccarelli, S.; Grando, S. Barley landraces from the Fertile Crescent: A lesson for plant breeders. In Genes in the Field: On-Farm Conservation of Crop Diversity, 1st ed.; Brush, S.B., Ed.; Lewis Publishers: Boca Raton, FL, USA, 2000; pp. 51–76. [Google Scholar]
- Jones, H.; Lister, D.L.; Bower, M.A.; Leigh, F.J.; Smith, L.; Jones, M. Approaches and constraints of using existing landraces and extant plant material to understand agricultural spread in prehistory. Plant Genet. Res. 2008, 6, 98–112. [Google Scholar] [CrossRef]
- Frankel, O.H.; Brown, A.H.D.; Burdon, J.J. The Conservation of Plant Biodiversity, 2nd ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 56–78. [Google Scholar]
- Zeven, A.C. The traditional inexplicable replacement of seed and seed ware of landraces and cultivars: A review. Euphytica 1999, 110, 181–191. [Google Scholar] [CrossRef]
- Singh, R.K.; Singh, A.; Pandey, C.B. Agro-biodiversity in rice–wheat-based agroecosystems of eastern Uttar Pradesh, India: Implications for conservation and sustainable management. Int. J. Sustain. Dev. World Ecol. 2014, 21, 46–59. [Google Scholar] [CrossRef]
- Negri, V. Landraces in central Italy: Where and why they are conserved and perspectives for their on-farm conservation. Genet. Resour. Crop. Evol. 2003, 50, 871–885. [Google Scholar] [CrossRef]
- Newton, A.C.; Akar, T.; Baresel, J.P.; Bebeli, P.J.; Bettencourt, E.; Bladenopoulos, K.V.; Czembor, J.H.; Fasoula, D.A.; Katsiotis, A.; Koutis, K.; et al. Cereal landraces for sustainable agriculture. A review. Agron. Sustain. Dev. 2010, 30, 237–269. [Google Scholar] [CrossRef] [Green Version]
- Bitocchi, E.; Nanni, L.; Rossi, M.; Rau, D.; Bellucci, E.; Giardini, A.; Buonamici, A.; Vendramin, G.G.; Papa, R. Introgression from modern hybrid varieties into landrace populations of maize (Zea mays ssp. mays L.) in central Italy. Mol. Ecol. 2009, 18, 603–621. [Google Scholar]
- Vigouroux, Y.; McMullen, M.; Hittinger, C.T.; Houchins, K.; Schulz, L.; Kresovich, S.; Matsuoka, Y.; Doebley, J. Identifying genes of agronomic importance in maize by screening microsatellites for evidence of selection during domestication. Proc. Natl. Acad. Sci. USA 2002, 99, 9650–9655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzucato, A.; Papa, R.; Bitocchi, E.; Mosconi, P.; Nanni, L.; Negri, V.; Picarella, M.E.; Siligato, F.; Soressi, G.P.; Tiranti, B.; et al. Genetic diversity, structure and marker-trait associations in a collection of Italian tomato (Solanum lycopersicum L.) landraces. Theor. Appl. Genet. 2008, 116, 657–669. [Google Scholar] [CrossRef]
- Comadran, J.; Kilian, B.; Russell, J.; Ramsay, L.; Stein, N.; Ganal, M.; Shaw, P.; Bayer, M.; Thomas, W.; Marshall, D.; et al. Natural variation in a homolog of Antirrhinum Centroradialis contributed to spring growth habit and environmental adaptation in cultivated barley. Nat. Genet. 2012, 44, 1388–1392. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro de Carvalho, M.A.A.; Bebeli, P.J.; Bettencourt, E.; Costa, G.; Dias, S.; Dos Santos, T.M.M.; Slaski, J.J. Cereal landraces genetic resources in worldwide GeneBanks. A review. Agron. Sustain. Dev. 2013, 33, 177–203. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, B.S.; Dua, R.P.; Brahmi, P.; Bisht, I.S. On farm conservation of plant genetic resources for food and agriculture. Curr. Sci. 2004, 87, 557–559. [Google Scholar]
- Brush, S.B. In Situ conservation of landraces in centers of crop diversity. Crop. Sci. 1995, 35, 346–354. [Google Scholar] [CrossRef]
- Hodgkin, T.; Rana, J.; Tuxill, J.; Balma, D.; Mar, I.; Karamura, D.; Valdivia, R.; Collado, L.; Latournerie, L.; Sadiki, M.; et al. Seed systems and crop genetic diversity in agroecosystems. In Managing Biodiversity in Agricultural Ecosystems; Jarvis, D., Padoch, C., Cooper, H.D., Eds.; CABI Publishing: Rome, Italy, 2007; pp. 77–116. [Google Scholar]
- Raggi, L.; Barata, A.M.; Heinonen, M.; Iriondo Alegría, J.M.; Kell, S.; Maxted, N.; Meierhofer, H.; Prohens, J.; Ralli, P.; Negri, V. In situ plant genetic resources in Europe: Landraces. In Farmer’s Pride: Networking, Partnerships and Tools to Enhance In Situ Conservation of European Plant Genetic Resources; The University of Birmingham: Birmingham, UK, 2020; pp. 1–26. [Google Scholar]
- Brush, S.B. The issues of in situ conservation of crop genetic resources. In Genes in the Field: On-Farm Conservation of Crop Diversity, 1st ed.; Brush, S.B., Ed.; Lewis Publishers: Boca Raton, FL, USA, 2000; pp. 1–28. [Google Scholar]
- Hawkes, J.G.; Maxted, N.; Floyd-Lloyd, B.V. The Ex Situ Conservation of Plant Genetic Resource; Kluwer: Amsterdam, The Netherlands, 2002; pp. 1–16. [Google Scholar]
- Mercer, K.L.; Perales, H.R. Evolutionary response of landraces to climate change in centers of crop diversity. Evol. Appl. 2010, 3, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.L.; Ogaya, J.R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O.; Peguero, G.; et al. Assessment of the impacts of climate change on Mediterranean terrestrial ecosystems based on data from field experiments and long-term monitored field gradients in Catalonia. Environ. Exp. Bot. 2018, 152, 49–59. [Google Scholar] [CrossRef]
- Khan, A.R.; Goldringer, I.; Thomas, M. Management practices and breeding history of varieties strongly determine the fine genetic structure of crop populations: A case study based on European wheat populations. Sustainability 2020, 12, 613. [Google Scholar] [CrossRef] [Green Version]
- Van Treuren, R.; Tchoudinova, I.; van Soest, L.J.M.; van Hintum, T.J.L. Marker assisted acquisition and core collection formation: A case study in barley using AFLPs and pedigree data. Genet. Resour. Crop. Evol. 2006, 53, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Reem, H.; Hodgkin, T. The use of wild relatives in crop improvement: A survey of developments over the last 20 years. Euphytica 2007, 156, 1–13. [Google Scholar]
- Jaradat, A.A. Wheat landraces: A mini review. Emir. J. Food Agric. 2013, 25, 20–29. [Google Scholar] [CrossRef]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Mazzucotelli, E.; Sciara, G.; Mastrangelo, A.M.; Desiderio, F.; Xu, S.S.; Faris, J.; Hayden, M.J.; Tricker, P.J.; Ozkan, H.; Echenique, V.; et al. The Global Durum Wheat Panel (GDP): An international platform to identify and exchange beneficial alleles. Front. Plant Sci. 2020, 11, 569905. [Google Scholar] [CrossRef]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Mangini, G.; Margiotta, B.; Marcotuli, I.; Signorile, M.A.; Gadaleta, A.; Blanco, A. Genetic diversity and phenetic analysis in wheat (Triticum turgidum subsp. durum and Triticum aestivum subsp. aestivum) landraces based on SNP markers. Genet. Resour. Crop. Evol. 2017, 64, 1269–1280. [Google Scholar]
- Alayachew, S.A.; Geletu, K.T. Genetic diversity of Ethiopian emmer wheat Triticum dicoccum Schrank landraces using seed storage proteins markers. Afr. J. Biotechnol. 2017, 16, 889–894. [Google Scholar]
- Negisho, K.; Shibru, S.; Pillen, K.; Ordon, F.; Wehner, G. Genetic diversity of Ethiopian durum wheat landraces. PLoS ONE 2021, 16, e0247016. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.C.; Mercati, F.; Spina, A.; Blangiforti, S.; Venora, G.; Dell’Acqua, M.; Lupini, A.; Preiti, G.; Monti, M.; Pè, M.E.; et al. High-throughput genotype, morphology, and quality traits evaluation for the assessment of genetic diversity of wheat landraces from Sicily. Plants 2019, 8, 116. [Google Scholar] [CrossRef] [Green Version]
- Sahri, A.; Chentoufi, L.; Arbaoui, M.; Ardisson, M.; Belqadi, L.; Birouk, A.; Roumet, P.; Muller, M.-H. Towards a comprehensive characterization of durum wheat landraces in Moroccan traditional agrosystems: Analysing genetic diversity in the light of geography, farmers’ taxonomy and tetraploid wheat domestication history. BMC Evol. Biol. 2014, 14, 264. [Google Scholar] [CrossRef] [Green Version]
- Alipour, H.; Bihamta, M.R.; Mohammadi, V.; Peyghambari, S.A.; Bai, G.; Zhang, G. Genotyping-by-Sequencing (GBS) revealed molecular genetic diversity of Iranian wheat landraces and cultivars. Front. Plant Sci. 2017, 8, 1293. [Google Scholar] [CrossRef]
- Frankin, S.; Kunta, S.; Abbo, S.; Sela, H.; Goldberg, B.Z.; Bonfil, D.J.; Levy, A.A.; Avivi-Ragolsky, N.; Nashef, K.; Roychowdhury, R.; et al. The Israeli–Palestinian wheat landraces collection: Restoration and characterization of lost genetic diversity. J. Sci. Food Agric. 2020, 100, 4083–4092. [Google Scholar] [CrossRef]
- Iqbal, N.; Tabasum, A.; Sayed, H.; Hameed, A. Evaluation of genetic diversity among bread wheat varieties and landraces of Pakistan by SSR markers. Cereal Res. Commun. 2009, 37, 489–498. [Google Scholar] [CrossRef]
- Karagoz, A. Wheat landraces of Turkey. Emir. J. Food Agric. 2014, 26, 149–156. [Google Scholar]
- Vikram, P.; Franco, J.; Burgueño-Ferreira, J.; Li, H.; Sehgal, D.; Saint Pierre, C.; Li, H.; Sehgal, D.; Saint Pierre, C.; Ortiz, C.; et al. Unlocking the genetic diversity of creole wheats. Nat. Sci. Rep. 2016, 6, 23092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, R.K.; Gautam, S.; Palikhey, E.; Joshi, B.K.; Ghimire, K.H.; Gurung, R.; Adhikari, A.R.; Pudasaini, N.; Dhakal, R. Agro-morphological diversity of Nepalese naked barley landraces. Agric. Food Secur. 2018, 7, 86. [Google Scholar] [CrossRef]
- Hagenblad, J.; Leino, M.W.; Afonso, G.H.; Morales, D.A. Morphological and genetic characterization of barley (Hordeum vulgare L.) landraces in the Canary Islands. Genet. Resour. Crop Evol. 2019, 66, 465–480. [Google Scholar] [CrossRef] [Green Version]
- Abdellaoui, R.; Kadir, K.; Naceur, M.B.; Kaab, L.B.B. Genetic diversity in some Tunisian barley landraces based on RAPD markers. Pak. J. Bot. 2010, 42, 3775–3782. [Google Scholar]
- Ben Romdhane, M.; Riahi, L.; Selmi, A.; Jardak, R.; Bouajila, A.; Ghorbel, A.; Zoghlami, N. Low genetic differentiation and evidence of gene flow among barley landrace populations in Tunisia. Crop. Sci. 2017, 57, 1585–1593. [Google Scholar] [CrossRef]
- Al-Abdallat, A.M.; Karadsheh, A.; Hadadd, N.I.; Akash, M.W.; Ceccarelli, S.; Baum, M.; Hasan, M.; Jighly, A.; Abu Elenein, J.M. Assessment of genetic diversity and yield performance in Jordanian barley (Hordeum vulgare L.) landraces grown under Rainfed conditions. BMC Plant Biol. 2017, 17, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boczkowska, M.; Tarczyk, E. Genetic diversity among Polish landraces of common oat (Avena sativa L.). Genet. Resour. Crop. Evol. 2013, 60, 2157–2169. [Google Scholar] [CrossRef] [Green Version]
- Montilla-Bascón, G.; Sánchez-Martín, J.; Rispail, N.; Rubiales, D.; Mur, L.; Langdon, T.; Griffiths, I.; Howarth, C.; Prats, E. Genetic diversity and population structure among oat cultivars and landraces. Plant Mol. Biol. Rep. 2013, 31, 1305–1314. [Google Scholar] [CrossRef] [Green Version]
- Pervaiz, Z.H.; Rabbani, M.A.; Khaliq, I.; Pearce, S.R.; Malik, S.A. Genetic diversity associated with agronomic traits using microsatellite markers in Pakistani rice landraces. Electr. J. Biotech. 2010, 13, 1–12. [Google Scholar]
- Ram, S.G.; Thiruvengadam, V.; Vinod, K.K. Genetic diversity among cultivars, landraces and wild relatives of rice as revealed by microsatellite markers. J. Appl. Genet. 2007, 48, 337–345. [Google Scholar] [CrossRef]
- Diack, O.; Kane, N.A.; Berthouly-Salazar, C.; Gueye, M.C.; Diop, B.M.; Fofana, A.; Sy, O.; Tall, H.; Zekraoui, L.; Piquet, M.; et al. New genetic insights into pearl millet diversity as revealed by characterization of early and late-flowering landraces from Senegal. Front. Plant Sci. 2017, 8, 818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Jia, G.; Zhi, H.; Niu, Z.; Chai, Y.; Li, W.; Wang, Y.; Li, H.; Lu, P.; Zhao, B.; et al. Genetic diversity and population structure of Chinese foxtail millet [Setaria italica (L.) Beauv.] landraces. G3 2012, 2, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Xu, Y.; He, J.; Zhang, S.; Wang, Y.; Lu, P. Genetic diversity and population structure of broomcorn millet (Panicum miliaceum L.) cultivars and landraces in China based on microsatellite markers. Int. J. Mol. Sci. 2016, 17, 370. [Google Scholar] [CrossRef]
- Raman, H.; Stodart, B.J.; Cavanagh, C.; Mackay, M.; Morell, M.; Milgate, A.; Martin, P. Molecular diversity and genetic structure of modern and traditional landrace cultivars of wheat (Triticum aestivum L.). Crop. Pasture Sci. 2010, 61, 222–229. [Google Scholar] [CrossRef]
- Kabbaj, H.; Sall, A.T.; Al-Abdallat, A.; Geleta, M.; Amri, A.; Filali-Maltouf, A.; Belkadi, B.; Ortiz, R.; Bassi, F.M. Genetic diversity within a Global Panel of Durum Wheat (Triticum durum) landraces and modern germplasm reveals the history of alleles exchange. Front. Plant Sci. 2017, 8, 1277. [Google Scholar] [CrossRef] [Green Version]
- Jilal, A.; Grando, S.; Henry, R.J.; Lee, L.S.; Rice, N.; Hill, H.; Baum, M.; Ceccarelli, S. Genetic diversity of ICARDA’s worldwide barley landrace collection. Genet. Resour. Crop. Evol. 2008, 55, 1221–1230. [Google Scholar] [CrossRef]
- Pasam, R.K.; Sharma, R.; Walther, A.; Özkan, H.; Graner, A.; Kilian, B. Genetic diversity and population structure in a legacy collection of spring barley landraces adapted to a wide range of climates. PLoS ONE 2014, 9, e116164. [Google Scholar] [CrossRef] [Green Version]
- Hagenblad, J.; Oliveira, H.R.; Forsberg, N.E.G.; Leino, M.W. Geographical distribution of genetic diversity in Secale landrace and wild accessions. BMC Plant Biol. 2016, 16, 23. [Google Scholar] [CrossRef] [Green Version]
- Mzid, R.; Chibani, F.; Ayed, R.B.; Hanana, M.; Breidi, J.; Kabalan, R.; El-Hajj, S.; Machlab, H.; Rebai, A.; Chalak, L. Genetic diversity in barley landraces (Hordeum vulgare L. subsp. vulgare) originated from Crescent Fertile region as detected by seed storage proteins. J. Genet. 2016, 95, 733–739. [Google Scholar]
- Melnikova, N.V.; Ganeva, G.D.; Popova, Z.G.; Landjeva, S.P.; Kudryavtsev, A.M. Gliadins of Bulgarian durum wheat (Triticum durum Desf.) landraces: Genetic diversity and geographical distribution. Genet. Resour. Crop. Evol. 2010, 57, 587–595. [Google Scholar] [CrossRef]
- Doležel, J.; Lucretti, S.; Molnár, I.; Cápal, P.; Giorgi, D. Chromosome analysis and sorting. Cytometry 2021, 99, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Zwyrtková, J.; Šimková, H.; Doležel, J. Chromosome genomics uncovers plant genome organization and function. Biotechnol. Adv. 2021, 46, 107659. [Google Scholar] [CrossRef] [PubMed]
- Kubaláková, M.; Vrána, J.; Cíhalíková, J.; Simková, H.; Doležel, J. Flow karyotyping and chromosome sorting in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2002, 104, 1362–1372. [Google Scholar] [CrossRef] [PubMed]
- Martis, M.M.; Zhou, R.; Haseneyer, G.; Schmutzer, T.; Vrána, J.; Kubaláková, M.; König, S.; Kugler, K.G.; Scholz, U.; Hackauf, B.; et al. Reticulate evolution of the rye genome. Plant Cell 2013, 25, 3685–3698. [Google Scholar] [CrossRef] [Green Version]
- Klaus, F.X.; Mayer, M.M.; Hedley, P.E.; Šimková, H.; Liu, H.; Morris, J.A.; Steuernagel, B.; Taudien, S.; Roessner, S.; Gundlach, H.; et al. Unlocking the barley genome by chromosomal and comparative genomics. Plant Cell 2011, 23, 1249–1263. [Google Scholar]
- International Wheat Genome Sequencing Consortium (IWGSC). A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar] [CrossRef]
- Robbana, C.; Kehel, Z.; Naceur, M.B.; Sansaloni, C.; Bassi, F.; Amri, A. Genome-Wide genetic diversity and population structure of Tunisian durum wheat landraces based on DArTseq technology. Int. J. Mol. Sci. 2019, 20, 1352. [Google Scholar] [CrossRef] [Green Version]
- Gaikpa, D.S.; Kessel, B.; Presterl, T.; Ouzunova, M.; Galiano-Carneiro, A.L.; Mayer, M.; Melchinger, A.E.; Schön, C.-C.; Miedaner, T. Exploiting genetic diversity in two European maize landraces for improving Gibberella ear rot resistance using genomic tools. Theor. Appl. Genet. 2021, 134, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, S. Landraces: Importance and use in breeding and environmentally friendly agronomic systems. In Agrobiodiversity Conservation: Securing the Diversity of Crop Wild Relatives and Landraces, 1st ed.; Maxted, N., Dulloo, E., Ford-Lloyd, B.V., Frese, L., Iriondo, J.M., Pinheiro de Carvalho, M.A.A., Eds.; CAB International: Wallingford, UK, 2012; pp. 103–117. [Google Scholar]
- Xu, K.; Mackill, D.J. A major locus for submergence tolerance mapped on rice chromosome 9. Mol. Breed. 1996, 2, 219–224. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Ronald, P.C.; Mackill, D.J. A high-resolution linkage map in the vicinity of the rice submergence tolerance locus Sub1. Mol. Gen. Genet. 2000, 263, 681. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Shabala, S.; Ma, Y.; Xu, R.; Zhou, M. Using QTL mapping to investigate the relationships between abiotic stress tolerance (drought and salinity) and agronomic and physiological traits. BMC Genomics 2015, 16, 43. [Google Scholar] [CrossRef] [Green Version]
- Reinert, S.; Kortz, A.; Léon, J.; Naz, A.A. Genome-wide association mapping in the global diversity set reveals new QTL controlling root system and related shoot variation in barley. Front. Plant Sci. 2016, 7, 1061. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Yi, X.; Tang, S.; Chen, W.; Wu, F.; Yang, X.; Jiang, X.; Shi, H.; Ma, J.; Chen, G.; et al. Dissection of phenotypic and genetic variation of drought-related traits in diverse Chinese wheat landraces. Plant Genome 2019, 12, 190025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.-L.; Huang, W.; Chao, D.-Y.; Zhu, M.-Z.; Wang, Z.-Y.; Luan, S.; Lin, H.-X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Puram, V.R.R.; Ontoy, J.; Subudhi, P.K. Identification of QTLs for salt tolerance traits and prebreeding lines with enhanced salt tolerance in an introgression line population of rice. Plant Mol. Biol. Rep. 2018, 36, 695–709. [Google Scholar] [CrossRef]
- Puram, V.R.R.; Ontoy, J.; Linscombe, S.; Subudhi, P.K. Genetic dissection of seedling stage salinity tolerance in rice using introgression lines of a salt tolerant landrace Nona Bokra. J. Hered. 2017, 108, 658–670. [Google Scholar] [CrossRef]
- Bonilla, P.S.; Dvorak, J.; Mackill, D.J.; Deal, K.; Gregorio, G.B. RFLP and SSLP mapping of salinity tolerance genes in chromosome 1 of rice (Oryza sativa L.) using recombinant inbred lines. Philipp. Agric. Sci. 2002, 85, 68–76. [Google Scholar]
- De Leon, T.B.; Linscombe, S.; Subudhi, P.K. Molecular dissection of seedling salinity tolerance in rice (Oryza sativa L.) using a high-density GBS-based SNP linkage map. Rice 2016, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Thomson, M.J.; de Ocampo, M.; Egdane, J.; Rahman, M.A.; Sajise, A.G.; Adorada, D.L.; Tumimbang-Raiz, E.; Blumwald, E.; Seraj, Z.I.; Singh, R.K.; et al. Characterizing the Saltol quantitative trait locus for salinity tolerance in rice. Rice 2010, 3, 148–160. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Singh, A.; Mithra, S.V.; Krishnamurthy, S.L.; Parida, S.K.; Jain, S.; Tiwari, K.K.; Kumar, P.; Rao, A.R.; Sharma, S.K.; et al. Genome-wide association mapping of salinity tolerance in rice (Oryza sativa). DNA Res. 2015, 22, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Wang, J.; Li, C.; Johnson, P.; Lu, C.; Zhou, M. A single locus is responsible for salinity tolerance in a Chinese landrace barley (Hordeum vulgare L.). PLoS ONE 2012, 7, e43079. [Google Scholar] [CrossRef]
- Rivandi, J.; Miyazaki, J.; Hrmova, M.; Pallotta, M.; Tester, M.; Collins, N.C. A SOS3 homologue maps to HvNax4, a barley locus controlling an environmentally sensitive Na+ exclusion trait. J. Exp. Bot. 2011, 62, 1201–1216. [Google Scholar] [CrossRef] [Green Version]
- Yau, S.K.; Ryan, J. Boron toxicity tolerance in crops: A viable alternative to soil amelioration. Crop. Sci. 2008, 48, 854–865. [Google Scholar] [CrossRef] [Green Version]
- Sutton, T.; Baumann, U.; Hayes, J.; Collins, N.C.; Shi, B.J.; Schnurbusch, T.; Hay, A.; Mayo, G.; Pallotta, M.; Tester, M.; et al. Boron-toxicity tolerance in barley arising from efflux transporter amplification. Science 2007, 318, 1446–1449. [Google Scholar] [CrossRef] [PubMed]
- Paull, J.G.; Nable, R.O.; Rathjen, A.J. Physiological and genetic control of the tolerance of wheat to high concentrations of boron and implications for plant breeding. Plant Soil 1992, 146, 251–260. [Google Scholar] [CrossRef]
- Hoang, G.T.; Gantet, P.; Nguyen, K.H.; Phung, N.T.P.; Ha, L.T.; Nguyen, T.T.; Lebrun, M.; Courtois, B.; Pham, X.H. Genome-wide association mapping of leaf mass traits in a Vietnamese rice landrace panel. PLoS ONE 2019, 14, e0219274. [Google Scholar] [CrossRef]
- Ta, K.N.; Khong, N.G.; Ha, T.L.; Nguyen, D.T.; Mai, D.C.; Hoang, T.G.; Phung, T.P.N.; Bourrie, I.; Courtois, B.; Tran, T.T.H.; et al. A genome-wide association study using a Vietnamese landrace panel of rice (Oryza sativa) reveals new QTLs controlling panicle morphological traits. BMC Plant Biol. 2018, 18, 282. [Google Scholar] [CrossRef]
- Huang, X.; Wei, X.; Sang, T.; Zhao, Q.; Feng, Q.; Zhao, Y.; Li, C.; Zhu, C.; Lu, T.; Zhang, Z.; et al. Genome-wide association studies of 14 agronomic traits in rice landraces. Nat. Genet. 2010, 42, 961–967. [Google Scholar] [CrossRef]
- Rahimi, Y.; Bihamta, M.R.; Taleei, A.; Alipour, H.; Ingvarsson, P.K. Genome-wide association study of agronomic traits in bread wheat reveals novel putative alleles for future breeding programs. BMC Plant Biol. 2019, 19, 541. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Dixit, S.; Ram, T.; Yadaw, R.B.; Mishra, K.K.; Mandal, N.P. Breeding high-yielding drought-tolerant rice: Genetic variations and conventional and molecular approaches. J. Exp. Bot. 2014, 65, 6265–6278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, I.S.; Neeraja, C.N.; Srikanth, B.; Subrahmanyam, D.; Swamy, K.N.; Rajesh, K.; Vijayalakshmi, P.; Kiran, T.V.; Sailaja, N.; Revathi, P.; et al. Identification of rice landraces with promising yield and the associated genomic regions under low nitrogen. Sci. Rep. 2018, 8, 9200. [Google Scholar] [CrossRef]
- Wissuwa, M.; Ae, N. Genotypic variation for tolerance to phosphorus deficiency in rice and the potential for exploitation in rice improvement. Plant Breed. 2001, 120, 43–48. [Google Scholar] [CrossRef]
- Wissuwa, M.; Wegner, J.; Ae, N.; Yano, M. Substitution mapping of the Pup1: A major QTL increasing phosphorus uptake of rice from a phosphorus deficient soil. Theor. Appl. Genet. 2002, 105, 890–897. [Google Scholar] [CrossRef]
- Chin, J.H.; Lu, X.; Haefele, S.M.; Gamuyao, R.; Ismail, A.; Wissuwa, M.; Heuer, S. Development and application of gene-based markers for the major rice QTL Phosphorus uptake 1. Theor. Appl. Genet. 2010, 120, 1073–1086. [Google Scholar] [CrossRef]
- Chin, J.H.; Gamuyao, R.; Dalid, C.; Bustamam, M.; Prasetiyono, J.; Moeljopawiro, S.; Wissuwa, M.; Heuer, S. Developing rice with high yield under phosphorus deficiency: Pup1 sequence to application. Plant Physiol. 2011, 156, 1202–1216. [Google Scholar] [CrossRef] [Green Version]
- Gamuyao, R.; Chin, J.H.; Pariasca-Tanaka, J.; Pesaresi, P.; Catausan, S.; Dalid, C.; Slamet-Loedin, I.; Tecson-Mendoza, E.M.; Wissuwa, M.; Heuer, S. The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency. Nature 2012, 488, 535–541. [Google Scholar] [CrossRef]
- Li, Y.; Huang, C.; Sui, X.; Fan, Q.; Li, G.; Chu, X. Genetic variation of wheat glutenin subunits between landraces and varieties and their contributions to wheat quality improvement in China. Euphytica 2009, 169, 159–168. [Google Scholar] [CrossRef]
- Sahu, P.K.; Mondal, S.; Sao, R.; Vishwakarma, G.; Kumar, V.; Das, B.K.; Sharma, D. Genome-wide association mapping revealed numerous novel genomic loci for grain nutritional and yield-related traits in rice (Oryza sativa L.) landraces. 3 Biotech 2020, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Cattivelli, L.; Rizza, F.; Badeck, F.-W.; Mazzucotelli, E.; Mastrangelo, A.M.; Francia, E.; Marè, C.; Tondelli, A.; Stanca, A.M. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crop. Res. 2008, 105, 1–14. [Google Scholar] [CrossRef]
- Gavhane, D.B.; Kulwal, P.L.; Kumbhar, S.D.; Jadhav, A.J.; Sarawate, C.D. Cataloguing of blast resistance genes in landraces and breeding lines of rice from India. J. Genet. 2019, 98, 106. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Kumar, S.; Gupta, S.; Gautam, N.; Singh, N.; Sharma, T. Functional complementation of rice blast resistance gene Pik h (Pi54) conferring resistance to diverse strains of Magnaporthe oryzae. J. Plant Biochem. Biotechnol. 2011, 20, 55–65. [Google Scholar] [CrossRef]
- Randhawa, M.; Bansal, U.; Valárik, M.; Klocová, B.; Doležel, J.; Bariana, H. Molecular mapping of stripe rust resistance gene Yr51 in chromosome 4AL of wheat. Theor. Appl. Genet. 2014, 127, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Babiker, E.M.; Gordon, T.C.; Chao, S.; Newcomb, M.; Rouse, M.N.; Jin, Y.; Wanyera, R.; Acevedo, M.; Brown-Guedira, G.; Williamson, S.; et al. Mapping resistance to the Ug99 race group of the stem rust pathogen in a spring wheat landrace. Theor. Appl. Genet. 2015, 128, 605–612. [Google Scholar] [CrossRef]
- Laidò, G.; Panio, G.; Marone, D.; Russo, M.A.; Ficco, D.B.M.; Giovanniello, V.; Cattivelli, L.; Steffenson, B.; De Vita, P.; Mastrangelo, A.M. Identification of new resistance loci to African stem rust race TTKSK in tetraploid wheats based on linkage and genome-wide association mapping. Front. Plant Sci. 2015, 6, 1033. [Google Scholar] [CrossRef] [Green Version]
- Babiker, E.M.; Gordon, T.C.; Chao, S.; Rouse, M.N.; Brown-Guedira, G.; Williamson, S.; Pretorius, Z.A.; Bonman, J.M. Rapid identification of resistance loci effective versus Puccinia graminis f. sp. tritici race TTKSK in 33 spring wheat landraces. Plant Dis. 2016, 100, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Muleta, K.T.; Rouse, M.N.; Rynearson, S.; Chen, X.; Buta, B.G.; Pumphrey, M.O. Characterization of molecular diversity and genome-wide mapping of loci associated with resistance to stripe rust and stem rust in Ethiopian bread wheat accessions. BMC Plant Biol. 2017, 17, 134. [Google Scholar] [CrossRef]
- Feng, J.Y.; Wang, M.N.; See, D.R.; Chao, S.M.; Zheng, Y.L.; Chen, X.M. Characterization of novel gene Yr79 and four additional QTL for all-stage and high-temperature adult-plant resistance to stripe rust in spring wheat PI 182103. Phytopathology 2018, 108, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Kolmer, J.A.; Garvin, D.F.; Hayden, M.; Spielmeyer, W. Adult plant leaf rust resistance derived from the wheat landrace cultivar Americano 44d is conditioned by interaction of three QTL. Euphytica 2018, 214, 59. [Google Scholar] [CrossRef]
- Saccomanno, A.; Matny, O.; Marone, D.; Laidò, G.; Petruzzino, G.; Mazzucotelli, E.; Desiderio, F.; Blanco, A.; Gadaleta, A.; Pecchioni, N.; et al. Genetic mapping of loci for resistance to stem rust in a tetraploid wheat collection. Int. J. Mol. Sci. 2018, 19, 3907. [Google Scholar] [CrossRef] [Green Version]
- Zurn, J.D.; Rouse, M.N.; Chao, S.; Aoun, M.; Macharia, G.; Hiebert, C.W.; Pretorius, Z.A.; Bonman, J.M.; Acevedo, M. Dissection of the multigenic wheat stem rust resistance present in the Montenegrin spring wheat accession PI 362698. BMC Genom. 2018, 19, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Lan, C.; Asad, M.A.; Gebrewahid, T.W.; Xia, X.; He, Z.; Li, Z.; Liu, D. QTL mapping of adult-plant resistance to leaf rust in the Chinese landraces Pingyuan 50/Mingxian 169 using the wheat 55K SNP array. Mol. Breed. 2019, 39, 98. [Google Scholar] [CrossRef]
- Saremirad, A.; Bihamta, M.R.; Malihipour, A.; Mostafavi, K.; Alipour, H. Genome-wide association study in diverse Iranian wheat germplasms detected several putative genomic regions associated with stem rust resistance. Food Sci. Nutr. 2020, 9, 1357–1374. [Google Scholar] [CrossRef]
- Tehseen, M.M.; Tonk, F.A.; Tosun, M.; Amri, A.; Sansaloni, C.P.; Kurtulus, E.; Yazbek, M.; Al-Sham’aa, K.; Ozseven, I.; Safdar, L.B.; et al. Genome wide association study of resistance to PstS2 and warrior races of stripe (yellow) rust in bread wheat landraces. Plant Genome 2020, 14, e20066. [Google Scholar] [CrossRef] [PubMed]
- Vikram, P.; Sehgal, D.; Sharma, A.; Bhavani, S.; Gupta, P.; Randhawa, M.; Pardo, N.; Basandra, D.; Srivastava, P.; Singh, S.; et al. Genome-wide association analysis of Mexican bread wheat landraces for resistance to yellow and stem rust. PLoS ONE 2021, 16, e0246015. [Google Scholar] [CrossRef] [PubMed]
- Aoun, M.; Kolmer, J.A.; Rouse, M.N.; Elias, E.M.; Breiland, M.; Bulbula, W.D.; Chao, S.; Acevedo, M. Mapping of novel leaf rust and stem rust resistance genes in the Portuguese durum wheat landrace PI 192051. G3 2019, 9, 2535–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, L.; Yao, F.; Yu, C.; Ye, X.; Cheng, Y.; Wang, Y.; Wu, Y.; Li, J.; Wang, J.; Jiang, Q.; et al. Genome-Wide association study for adult-plant resistance to stripe rust in Chinese wheat landraces (Triticum aestivum L.) from the Yellow and Huai River Valleys. Front. Plant Sci. 2019, 10, 256. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.W.; Herrera-Foessel, S.; Lan, C.; Schnippenkoetter, W.; Ayliffe, M.; Huerta-Espino, J.; Lillemo, M.; Viccars, L.; Milne, R.; Periyannan, S.; et al. A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 2015, 47.12, 1494–1498. [Google Scholar] [CrossRef]
- Cai, J.; Wang, S.; Li, T.; Zhang, G.; Bai, G. Multiple minor QTLs are responsible for Fusarium head blight resistance in Chinese wheat landrace Haiyanzhong. PLoS ONE 2016, 11, e0163292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, S.; Larkin, P.; Xu, R.; Hayden, M.; Forrest, K.; Meinke, H.; Hu, H.; Zhou, M.; Fan, Y. Genome wide association study reveals novel QTL for barley yellow dwarf virus resistance in wheat. BMC Genom. 2019, 20, 891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Li, H.; Zhang, D.; Guo, L.; Chen, J.; Chen, Y.; Wu, Q.; Xie, J.; Zhang, Y.; Sun, Q.; et al. Genetic and physical mapping of powdery mildew resistance gene MlHLT in Chinese wheat landrace Hulutou. Theor. Appl. Genet. 2015, 128, 365–373. [Google Scholar] [CrossRef]
- Sun, H.; Hu, J.; Song, W.; Qiu, D.; Cui, L.; Wu, P.; Zhang, H.; Liu, H.; Yang, L.; Qu, Y.; et al. Pm61: A recessive gene for resistance to powdery mildew in wheat landrace Xuxusanyuehuang identified by comparative genomics analysis. Theor. Appl. Genet. 2018, 131, 2085–2097. [Google Scholar] [CrossRef]
- Xue, S.; Lu, M.; Hu, S.; Xu, H.; Ma, Y.; Lu, N.; Bai, S.; Gu, A.; Wan, H.; Li, S. Characterization of PmHHXM, a new broad-spectrum powdery mildew resistance gene in Chinese wheat landrace Honghuaxiaomai. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Varella, A.C.; Zhang, H.; Weaver, D.K.; Cook, J.P.; Hofland, M.L.; Lamb, P.; Chao, S.; Martin, J.M.; Blake, N.K.; Talbert, L.E. A novel QTL in durum wheat for resistance to the wheat stem sawfly associated with early expression of stem solidness. G3 2019, 9, 1999–2006. [Google Scholar] [CrossRef] [Green Version]
- He, X.Y.; Singh, P.K.; Schlang, N.; Duveiller, E.; Dreisigacker, S.; Payne, T.; He, Z.H. Characterization of Chinese wheat germplasm for resistance to Fusarium head blight at CIMMYT, Mexico. Euphytica 2014, 195, 383–395. [Google Scholar] [CrossRef]
- Xue, S.L.; Li, G.Q.; Jia, H.Y.; Xu, F.; Lin, F.; Tang, M.Z.; Wang, Y.; An, X.; Xu, H.B.; Zhang, L.X.; et al. Fine mapping Fhb4 a major QTL conditioning resistance to Fusarium infection in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2010, 121, 147–156. [Google Scholar] [CrossRef]
- Xue, S.; Xu, F.; Tang, M.; Zhou, Y.; Li, G.; An, X.; Lin, F.; Xu, H.; Jia, H.; Zhang, L.; et al. Precise mapping Fhb5, a major QTL conditioning resistance to Fusarium infection in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2011, 123, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Bian, Q.; Gao, Y.; Ni, X.; Sun, Y.; Xuan, Y.; Cao, Y.; Li, T. Evaluation of resistance to powdery mildew and identification of resistance genes in wheat cultivars. PeerJ 2021, 9, e10425. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Guo, L.; Wang, Z.; Li, B.; Li, J.; Li, Y.; Qiu, D.; Shi, W.; Yang, L.; Wang, N.; et al. A rare gain of function mutation in a wheat tandem kinase confers resistance to powdery mildew. Nat. Commun. 2020, 11, 680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yahiaoui, N.; Srichumpa, P.; Dudler, R.; Keller, B. Genome analysis at different ploidy levels allows cloning of the powdery mildew resistance gene Pm3b from hexaploid wheat. Plant J. 2004, 37, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Silvar, C.; Perovic, D.; Nussbaumer, T.; Spannagl, M.; Usadel, B.; Casas, A.; Igartua, E.; Ordon, F. Towards positional isolation of three quantitative trait loci conferring resistance to powdery mildew in two Spanish barley landraces. PLoS ONE 2013, 8, e67336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goddard, R.; de Vos, S.; Steed, A.; Muhammed, A.; Thomas, K.; Griggs, D.; Ridout, C.; Nicholson, P. Mapping of agronomic traits, disease resistance and malting quality in a wide cross of two-row barley cultivars. PLoS ONE 2019, 14, e0219042. [Google Scholar] [CrossRef] [Green Version]
- Piechota, U.; Czembor, P.C.; Słowacki, P.; Czembor, J.H. Identifying a novel powdery mildew resistance gene in a barley landrace from Morocco. J. Appl. Genet. 2019, 60, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Kusch, S.; Panstruga, R. mlo-based resistance: An apparently universal “weapon” to defeat powdery mildew disease. MPMI 2017, 30, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Daba, S.; Horsley, R.; Brueggeman, R.; Chao, S.; Mohammadi, M. Genome-wide association studies and candidate gene identification for leaf scald and net blotch in barley (Hordeum vulgare L.). Plant Dis. 2019, 103. [Google Scholar] [CrossRef] [PubMed]
- Novakazi, F.; Afanasenko, O.; Anisimova, A.; Platz, G.J.; Snowdon, R.; Kovaleva, O.; Zubkovich, A.; Ordon, F. Genetic analysis of a worldwide barley collection for resistance to net form of net blotch disease (Pyrenophora teres f. teres). Theor. Appl. Genet. 2019, 132, 2633–2650. [Google Scholar] [CrossRef] [PubMed]
- Mamo, B.; Brueggeman, R.; Smith, K.; Steffenson, B. Genetic characterization of resistance to wheat stem rust race TTKSK in landrace and wild barley accessions identifies the rpg4/Rpg5 locus. Phytopathology 2015, 105, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Jiang, C.; He, Q.; Habekuß, A.; Ordon, F.; Luan, H.; Shen, H.; Liu, J.; Feng, Z.; Zhang, J.; et al. Bulked segregant RNA-sequencing (BSR-seq) identified a novel rare allele of eIF4E effective against multiple isolates of BaYMV/BaMMV. Theor. Appl. Genet. 2019, 132, 1777–1788. [Google Scholar] [CrossRef] [PubMed]
- Hurni, S.; Scheuermann, D.; Krattinger, S.G.; Kessel, B.; Wicker, T.; Herren, G.; Fitze, M.N.; Breen, J.; Presterl, T.; Ouzunova, M.; et al. The maize disease resistance gene Htn1 against northern corn leaf blight encodes a wall-associated receptor-like kinase. Proc. Natl. Acad. Sci. USA 2015, 112, 8780–8785. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yang, Q.; Dai, Z.; Wang, Y.; Zhang, Y.; Li, B.; Zhao, W.; Hao, J. Identification of QTLs for resistance to maize rough dwarf disease using two connected RIL populations in maize. PLoS ONE 2019, 14, e0226700. [Google Scholar] [CrossRef]
- Montilla-Bascón, G.; Rispail, N.; Sánchez-Martín, J.; Rubiales, D.; Mur, L.A.J.; Langdon, T.; Howarth, C.J.; Prats, E. Genome-wide association study for crown rust (Puccinia coronata f. sp. avenae) and powdery mildew (Blumeria graminis f. sp. avenae) resistance in an oat (Avena sativa) collection of commercial varieties and landraces. Front. Plant Sci. 2015, 6, 103. [Google Scholar]
- Yahiaoui, S.; Cuesta-Marcos, A.; Gracia, M.P.; Medina, B.; Lasa, J.M.; Casas, A.M.; Ciudad, F.J.; Montoya, J.L.; Moralejo, M.; Molina-Cano, J.L.; et al. Spanish barley landraces outperform modern cultivars at low-productivity sites. Plant Breed. 2014, 133, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Abtew, W.G.; Lakew, B.; Haussmann, B.I.G.; Schmid, K.J. Ethiopian barley landraces show higher yield stability and comparable yield to improved varieties in multi-environment field trials. J. Plant Breed. Crop. Sci. 2015, 7, 275–291. [Google Scholar]
- Ceccarelli, S.; Grando, S.; Impiglia, A. Choice of selection strategy in breeding barley for stress environments. Euphytica 1998, 103, 307–318. [Google Scholar] [CrossRef]
- Comadran, J.; Thomas, W.T.B.; Van Eeuwijk, F.A.; Ceccarelli, S.; Grando, S.; Stanca, A.M.; Pecchioni, N.; Akar, T.; Al-Yassin, A.; Benbelkacem, A.; et al. Patterns of genetic diversity and linkage disequilibrium in a highly structured Hordeum vulgare association-mapping population for the Mediterranean basin. Theor. Appl. Genet. 2009, 119, 175–187. [Google Scholar] [CrossRef]
- Dawson, I.K.; Russell, J.; Powell, W.; Steffenson, B.; Thomas, W.T.B.; Waugh, R. Barley: A translational model for adaptation to climate change. New Phytol. 2015, 206, 913–931. [Google Scholar] [CrossRef]
- Lopes, M.; Dreisigacker, S.; Peña, R.; Sukumaran, S.; Reynolds, M. Genetic characterization of the wheat association mapping initiative (WAMI) panel for dissection of complex traits in spring wheat. Theor. Appl. Genet. 2015, 128, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; et al. Exploiting genetic diversity from landraces in wheat breeding for adaptation to climate change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef] [PubMed]
- Hiebert, C.W.; Thomas, J.B.; McCallum, B.D.; Humphreys, D.G.; DePauw, R.M.; Hayden, M.J.; Mago, R.; Schnippenkoetter, W.; Spielmeyer, W. An introgression on wheat chromosome 4DL in RL6077 (Thatcher*6/PI 250413) confers adult plant resistance to stripe rust and leaf rust (Lr67). Theor. Appl. Genet. 2010, 121, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, G.; Srinivasarao, K.; Mohan, K.M.; Sudarshan, I.; Sivaranjani, A.K.P.; Gopalakrishna, K.; Neeraja, C.N.; Balachandran, S.M.; Sundaram, R.M.; Prasad, M.S.; et al. Development and validation of functional marker targeting an InDel in the major rice blast disease resistance gene Pi54 (Pik h). Mol. Breed. 2011, 27, 129–135. [Google Scholar] [CrossRef]
- Salgotra, R.K.; Stewart, C.N. Functional markers for precision plant breeding. Int. J. Mol. Sci. 2020, 21, 4792. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Serres, J.; Fukao, T.; Ronald, P.; Ismail, A.; Heuer, S.; Mackill, D. Submergence tolerant rice: SUB1’s journey from landrace to modern cultivar. Rice 2010, 3, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Dar, M.H.; de Janvry, A.; Emerick, K.; Raitzer, D.; Sadoulet, E. Flood tolerant rice reduces yield variability and raises expected yield, differentially benefitting socially disadvantaged groups. Sci. Rep. 2013, 3, 3315. [Google Scholar] [CrossRef] [Green Version]
- Mackill, D.J.; Ismail, A.M.; Singh, U.S.; Labios, R.V.; Paris, T.R. Chapter Six—Development and rapid adoption of submergence-tolerant (Sub1) rice varieties. Adv. Agron. 2012, 115, 299–352. [Google Scholar]
- Monteagudo, A.; Casas, A.M.; Cantalapiedra, C.P.; Contreras-Moreira, B.; Pilar Gracia, M.; Igartua, E. Harnessing novel diversity from landraces to improve an elite barley variety. Front. Plant Sci. 2019, 10, 434. [Google Scholar] [CrossRef]
- Prasanna, B.M. Diversity in global maize germplasm: Characterization and utilization. J. Biosci. 2012, 37, 843–855. [Google Scholar] [CrossRef]
- Meseka, S.; Fakorede, M.; Ajala, S.; Badu-Apraku, B.; Menkir, A. Introgression of alleles from maize landraces to improve drought tolerance in an unadapted germplasm. J. Crop. Improv. 2013, 27, 96–112. [Google Scholar] [CrossRef]
- Meseka, S.; Menkir, A.; Obeng-Antwi, K. Exploitation of beneficial alleles from maize (Zea mays L.) landraces to enhance performance of an elite variety in water stress environments. Euphytica 2015, 201, 149–160. [Google Scholar] [CrossRef]
- Rakshit, S.; Gomashe, S.S.; Ganapathy, K.N.; Elangovan, M.; Ratnavathi, C.V.; Seetharama, N.; Patil, J.V. Morphological and molecular diversity reveal wide variability among sorghum Maldandi landraces from India. J. Plant Biochem. Biotechnol. 2012, 21, 145–156. [Google Scholar] [CrossRef]
- Kemper, E.L.; Neto, G.C.; Papes, F.; Moraes, K.C.; Leite, A.; Arruda, P. The role of opaque2 in the control of lysine-degrading activities in developing maize endosperm. Plant Cell 1999, 11, 1981–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langridge, P.; Waugh, R. Harnessing the potential of germplasm collections. Nat. Genet. 2019, 51, 200–201. [Google Scholar] [CrossRef]
- Winfield, M.O.; Allen, A.M.; Wilkinson, P.A.; Burridge, A.J.; Barker, G.L.; Coghill, J.; Waterfall, C.; Wingen, L.U.; Griffiths, S.; Edwards, K.J. High-density genotyping of the AE Watkins collection of hexaploid landraces identifies a large molecular diversity compared to elite bread wheat. Plant Biotech. J. 2018, 16, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodhi, S.S.; Shafia, M.; Rafique, K.; Atif, S.; Yousaf, Z.A.; Talha, A.M.; Gul, A.; Amir, R. Overview of the prospective strategies for conservation of genomic diversity in wheat landraces. In Climate Change and Food Security with Emphasis on Wheat, 1st ed.; Munir-Ozturk, M., Gul, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; Chapter 21; pp. 293–309. [Google Scholar]


{kind=link}
| Species | Landrace (Origin) | Gene/QTL | Trait | Analysis Type | Refs. |
|---|---|---|---|---|---|
| Abiotic Stress | |||||
| Wheat | G61450 (Greece) | Bo4 | Boron tolerance | Molecular mapping | [92] |
| Rice | FR13A (India) | SUB1 | Submergence tolerance | Molecular mapping | [75] |
| Rice | Aus, Indica, and Basmati accessions (India) | 14 QTLs | Drought tolerance | Molecular mapping | [97] |
| Rice | Nona Bokra (India) | qSKC-1 | Salinity tolerance | Molecular mapping | [81] |
| Rice | Nona Bokra (India) | 16 QTLs | Salinity tolerance | Molecular mapping | [82,83] |
| Rice | Pokkali (India) | Saltol1 | Salinity tolerance | Molecular mapping | [84] |
| Rice | Pokkali (India) | 10 QTLs | Salinity tolerance | Molecular mapping | [85] |
| Rice | Kasalath (india) | PUP1 | Phosphorus uptake increase | Molecular mapping | [99,100] |
| Barley | TX9425 (China) | 2 QTLs | Drought tolerance | Molecular mapping | [78] |
| Barley | Accessions from diverse geographic areas | QRdw.5H | Drought tolerance | GWAS | [79] |
| Barley | Chinese accessions | 26 QTLs | Drought tolerance | GWAS | [80] |
| Barley | TX9425 (China) | 1 QTL | Salinity tolerance | Molecular mapping | [78] |
| Barley | TX9425 (China) | 1 QTL | Salinity and waterlogging tolerance | Molecular mapping | [88] |
| Barley | Sahara 3771 (Algeria) | HvNax4 | Salinity tolerance | Molecular mapping | [89] |
| Barley | Sahara 3771 (Algeria) | Bot1 | Boron tolerance | Molecular mapping | [91] |
| Biotic Stress | |||||
| Wheat | AUS27858 (Australia) | Yr51 | Stripe rust resistance | Molecular mapping | [109] |
| Wheat | CItr 4311 (Iran) | Sr9h | Stem rust resistance | Molecular mapping | [110] |
| Tetraploid Wheat | Accessions from diverse geographic areas | 35 QTLs | Stem rust resistance | GWAS | [111] |
| Wheat | Ethiopian accessions | 24 QTLs | Stem and stripe resistance | GWAS | [113] |
| Wheat | PI 182103 (Pakistan) | Yr79 | Stripe rust resistance | Molecular mapping | [114] |
| Wheat | Americano 44 (Uruguay) | 3 major QTLs | Leaf rust resistance | Molecular mapping | [115] |
| Tetraploid Wheat | Accessions from diverse geographic areas | 22 QTLs | Stem rust resistance | GWAS | [116] |
| Wheat | PI 362698 (Montenegro) | 5 QTLs | Stem rust resistance | Molecular mapping | [117] |
| Wheat | Pingyuan 50/Mingxian 169 (China) | 8 QTLs | Leaf rust resistance | Molecular mapping | [118] |
| Wheat | Iranian accessions | 54 QTLs | Stem rust resistance | GWAS | [119] |
| Wheat | ICARDA accessions | 19 QTLs | Stripe rust resistance | GWAS | [120] |
| Wheat | Mexican accessions | 17 QTLs | Stripe and stem rust resistance | GWAS | [121] |
| Durum Wheat | PI 192051 (Portugal) | 2 major and 3 minor QTLs | Leaf and stem rusts resistance | Molecular mapping | [122] |
| Wheat | Chinese accessions | 40 QTLs | Yellow rust resistance | GWAS | [123] |
| Wheat | PI 250413 (Pakistan) | Lr67 | Leaf, stem and stripe rust, and powdery mildew resistance | Mutagenesis and transformation | [124] |
| Wheat | Haiyanzhong (China) | 6 QTLs | Fusarium Head Blight resistance | Molecular mapping | [125] |
| Wheat | Chinese and Australian accessions | 36 QTLs | Barley yellow dwarf virus resistance | GWAS | [126] |
| Wheat | Hulutou (China) | MlHLT | Powdery mildew resistance | Molecular mapping | [127] |
| Wheat | Xuxusanyuehuang (China) | Pm61 | Powdery mildew Resistance | Molecular mapping | [128] |
| Wheat | Honghuaxiaomai (China) | PmHHXM | Powdery mildew resistance | Molecular mapping | [129] |
| Wheat | Duanganmang (China) | PmDGM | Powdery mildew resistance | Molecular mapping | [134] |
| Wheat | Chiyacao, Baihulu Hulutou (China) | Pm24, Pm24b, MIHLT | Powdery mildew resistance | Molecular mapping | [135] |
| Wheat | ChuI | Pm3b | Powdery mildew resistance | Positional cloning | [136] |
| Durum Wheat | PI 166471 (Turkey) | 9 QTLs | Stem sawfy resistance | Molecular mapping | [130] |
| Wheat | Wangshuibai (China) | 2 QTLs (Fhb4, Fhb5) | Fusarium Head Blight resistance | Molecular mapping | [132,133] |
| Rice | Tetep (Vietnma) | Pi54 | Blast resistance | Molecular mapping | [108] |
| Barley | SBCC097/SBCC145 (Spain) | 3 QTLs | Powdery mildew resistance | Molecular mapping and positional cloning | [137] |
| Barley | Chevallier (UK) | 5 QTLs | Powdery mildew resistance | Molecular mapping | [138] |
| Barley | Landrace 255 (ICARDA No. ICB 31956) (Morocco) | MlMor | Powdery mildew resistance | Molecular mapping | [139] |
| Barley | Ethiopian and American accessions | 51 QTLs | Leaf scald and net blotch | GWAS | [141] |
| Barley | Accessions from diverse geographic areas | 15 QTLs | Net blotch | GWAS | [142] |
| Barley | Hv501, Hv545, Hv602, Hv612 (Switzerland) | Rpg5 | Stem rust | BSA | [143] |
| Barley | HOR3298 (Iran) | eIF4E | Barley yellow mosaic virus | BSR-seq | [144] |
| Maize | Kemater Landmais Gelb/Petkuser Ferdinand Rot (Germany) | 8 QTLs | Gibberella ear rot | GWAS | [73] |
| Maize | Pepitilla (Mexico) | Htn1 | Northern Corn leaf blight | Map-based cloning | [145] |
| Maize | D863F/ZS301 (China) | 10 QTLs | Rough dwarf disease | Molecular mapping | [146] |
| Oat | Spanish accessions | 6 QTLs | Crown rust and powdery mildew | GWAS | [147] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marone, D.; Russo, M.A.; Mores, A.; Ficco, D.B.M.; Laidò, G.; Mastrangelo, A.M.; Borrelli, G.M. Importance of Landraces in Cereal Breeding for Stress Tolerance. Plants 2021, 10, 1267. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071267
Marone D, Russo MA, Mores A, Ficco DBM, Laidò G, Mastrangelo AM, Borrelli GM. Importance of Landraces in Cereal Breeding for Stress Tolerance. Plants. 2021; 10(7):1267. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071267
Chicago/Turabian StyleMarone, Daniela, Maria A. Russo, Antonia Mores, Donatella B. M. Ficco, Giovanni Laidò, Anna M. Mastrangelo, and Grazia M. Borrelli. 2021. "Importance of Landraces in Cereal Breeding for Stress Tolerance" Plants 10, no. 7: 1267. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071267