
The Combination of Mild Salinity Conditions and Exogenously Applied Phenolics Modulates Functional Traits in Lettuce

 , , , , ,
, , , , ,  , and
, and 
Abstract
:1. Introduction
2. Results
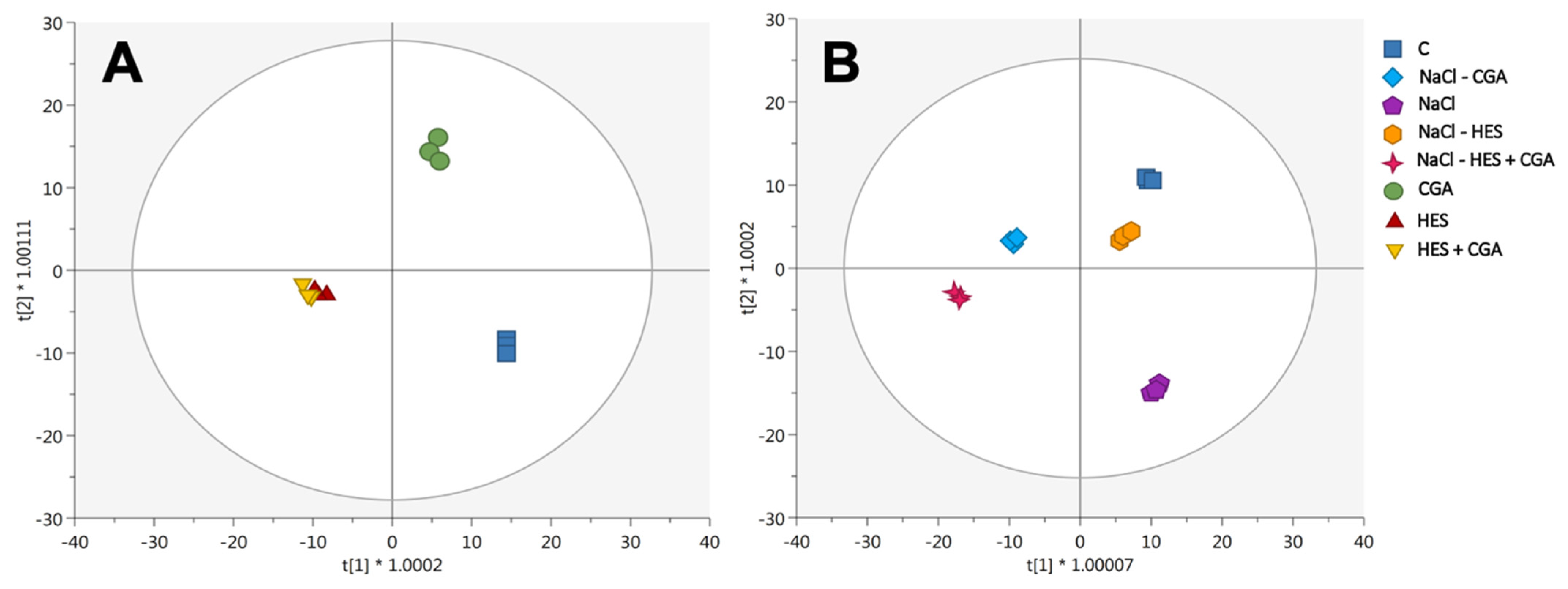
2.1. Metabolomic Profiling of Lettuce Treated with Hesperidin and Chlorogenic Acid
2.2. Multivariate Discrimination Analysis Phenolic Profile Following the Different Treatments Applied under Salinity and Non-Salinity Conditions
2.3. Antioxidant Capacity
2.4. Enzyme Inhibition Activity
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions, and Treatment
4.2. Untargeted Phenolic Compounds Profiling by UHPLC-QTOF Mass Spectrometry
4.3. In Vitro Antioxidant Assays
4.4. In Vitro Enzyme-Inhibitory Assays
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gong, X.; Li, X.; Xia, Y.; Xu, J.; Li, Q.; Zhang, C.; Li, M. Effects of phytochemicals from plant-based functional foods on hyperlipidemia and their underpinning mechanisms. Trends Food Sci. Technol. 2020, 103, 304–320. [Google Scholar] [CrossRef]
- Robson, A.A. Chapter 25–Preventing the Epidemic of Non-Communicable Diseases: An Overview. In Bioactive Food as Dietary Interventions for Liver and Gastrointestinal Disease; Watson, R.R., Preedy, V.R., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 383–400. [Google Scholar]
- Mei, Y.; Sun, H.; Du, G.; Wang, X.; Lyu, D. Exogenous chlorogenic acid alleviates oxidative stress in apple leaves by enhancing antioxidant capacity. Sci. Hortic. 2020, 274, 109676. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Dhewa, T.; Moreno-Rojas, J.M. Potential Health Benefits of Plant Food-Derived Bioactive Components: An Overview. Foods 2021, 10, 839. [Google Scholar] [CrossRef] [PubMed]
- House, N.C.; Puthenparampil, D.; Malayil, D.; Narayanankutty, A. Variation in the polyphenol composition, antioxidant, and anticancer activity among different Amaranthus species. South Afr. J. Bot. 2020, 135, 408–412. [Google Scholar] [CrossRef]
- Yu, M.; Gouvinhas, I.; Rocha, J.; Barros, A. Phytochemical and antioxidant analysis of medicinal and food plants towards bioactive food and pharmaceutical resources. Sci Rep. 2021, 11, 10041. [Google Scholar] [CrossRef]
- Mpofu, A.; Sapirstein, H.D.; Beta, T. Genotype and environmental variation in phenolic content, phenolic acid composition, and antioxidant activity of hard spring wheat. J. Agric. Food Chem. 2006, 54, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Samec, D.; Karalija, E.; Sola, I.; Vujcic Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Grzeszczuk, M.; Salachna, P.; Meller, E. Changes in Photosynthetic Pigments, Total Phenolic Content, and Antioxidant Activity of Salvia coccinea Buc’hoz Ex Etl. Induced by Exogenous Salicylic Acid and Soil Salinity. Molecules 2018, 23, 1296. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Cheng, D.M.; Pogrebnyak, N.; Kuhn, P.; Poulev, A.; Waterman, C.; Rojas-Silva, P.; Johnson, W.D.; Raskin, I. Polyphenol-rich Rutgers Scarlet Lettuce improves glucose metabolism and liver lipid accumulation in diet-induced obese C57BL/6 mice. Nutrition 2014, 30 (Suppl. 7–8), S52–S58. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Liang, X.; Li, K.; Dai, P.; Li, J.; Liang, B.; Sun, C.; Lin, X. Metabolomics analysis reveals potential mechanisms of phenolic accumulation in lettuce (Lactuca sativa L.) induced by low nitrogen supply. Plant Physiol Biochem. 2021, 158, 446–453. [Google Scholar] [CrossRef]
- Malarz, J.; Michalska, K.; Stojakowska, A. Stem Lettuce and Its Metabolites: Does the Variety Make Any Difference? Foods 2020, 10, 59. [Google Scholar] [CrossRef]
- Pannico, A.; Graziani, G.; El-Nakhel, C.; Giordano, M.; Ritieni, A.; Kyriacou, M.; Rouphael, Y. Nutritional stress suppresses nitrate content and positively impacts ascorbic acid concentration and phenolic acids profile of lettuce microgreens. Italus Hortus 2020, 27, 41–52. [Google Scholar] [CrossRef]
- Camejo, D.; Frutos, A.; Mestre, T.C.; del Carmen Piñero, M.; Rivero, R.M.; Martínez, V. Artificial light impacts the physical and nutritional quality of lettuce plants. Hortic. Environ. Biotechnol. 2020, 61, 69–82. [Google Scholar] [CrossRef]
- Parvin, K.; Nahar, K.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Mohsin, S.M.; Fujita, M. Exogenous vanillic acid enhances salt tolerance of tomato: Insight into plant antioxidant defense and glyoxalase systems. Plant Physiol. Biochem. 2020, 150, 109–120. [Google Scholar] [CrossRef]
- Rouphael, Y.; Petropoulos, S.A.; Cardarelli, M.; Colla, G. Salinity as eustressor for enhancing quality of vegetables. Sci. Hortic. 2018, 234, 361–369. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Contreras, M.M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Reversed-phase ultra-high-performance liquid chromatography coupled to electrospray ionization-quadrupole-time-of-flight mass spectrometry as a powerful tool for metabolic profiling of vegetables: Lactuca sativa as an example of its application. J. Chromatogr. 2013, 1313, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Viacava, G.E.; Roura, S.I.; Berrueta, L.A.; Iriondo, C.; Gallo, B.; Alonso-Salces, R.M. Characterization of phenolic compounds in green and red oak-leaf lettuce cultivars by UHPLC-DAD-ESI-QToF/MS using MSE scan mode. J. Mass Spectrom. 2017, 52, 873–902. [Google Scholar] [CrossRef] [PubMed]
- El-Shafey, N.M.; AbdElgawad, H.L. a bioactive flavone compound extracted from Cichorium endivia L. subsp. divaricatum alleviates the harmful effect of salinity on maize. Acta Physiol. Plant. 2012, 34, 2165–2177. [Google Scholar] [CrossRef]
- Sanchez, D.H.; Siahpoosh, M.R.; Roessner, U.; Udvardi, M.; Kopka, J. Plant metabolomics reveals conserved and divergent metabolic responses to salinity. Physiol. Plant. 2008, 132, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, H.; Huang, J.; Gruber, M.Y.; Kaddour, R.; Lachaâl, M.; Ouerghi, Z.; Hannoufa, A. The Impact of Genotype and Salinity on Physiological Function, Secondary Metabolite Accumulation, and Antioxidative Responses in Lettuce. J. Agric. Food Chem. 2010, 58, 5122–5130. [Google Scholar] [CrossRef]
- Ahmed, S.; Ahmed, S.; Roy, S.K.; Woo, S.H.; Sonawane, K.D.; Shohael, A.M. Effect of salinity on the morphological, physiological and biochemical properties of lettuce (Lactuca sativa L.) in Bangladesh. Open Agric. 2019, 4, 361–373. [Google Scholar] [CrossRef]
- Kim, H.-J.; Fonseca, J.M.; Choi, J.-H.; Kubota, C.; Kwon, D.Y. Salt in Irrigation Water Affects the Nutritional and Visual Properties of Romaine Lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef]
- Blasco, B.; Leyva, R.; Romero, L.; Ruiz, J.M. Iodine Effects on Phenolic Metabolism in Lettuce Plants under Salt Stress. J. Agric. Food Chem. 2013, 61, 2591–2596. [Google Scholar] [CrossRef]
- Rauf, A.; Jehan, N. Natural products as a potential enzyme inhibitors from medicinal plants. Enzym. Inhib. Act. InTech Rij. Croat. 2017, 7, 165–177. [Google Scholar]
- Mishra, P.; Kumar, A.; Panda, G. Anti-cholinesterase hybrids as multi-target-directed ligands against Alzheimer’s disease (1998–2018). Biorg. Med. Chem. 2019, 27, 895–930. [Google Scholar] [CrossRef]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2021, 338, 128119. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Biswas, R.; Sharma, A.; Banerjee, S.; Biswas, S.; Katiyar, C.K. Validation of medicinal herbs for anti-tyrosinase potential. J. Herb. Med. 2018, 14, 1–16. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Charoenlertkul, P.; Yibchok-anun, S. α-Glucosidase inhibitory activity of cyanidin-3-galactoside and synergistic effect with acarbose. J. Enzym. Inhib. Med. Chem. 2009, 24, 65–69. [Google Scholar] [CrossRef]
- Hosseini-Zare, M.S.; Sarhadi, M.; Zarei, M.; Thilagavathi, R.; Selvam, C. Synergistic effects of curcumin and its analogs with other bioactive compounds: A comprehensive review. Eur. J. Med. Chem. 2021, 210, 113072. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zheng, Z.; Liu, C.; Wang, Y.; Li, J.; Liu, Y. Identification and quantification of synergetic antioxidants and their application in sunflower oil. LWT 2020, 118, 108726. [Google Scholar] [CrossRef]
- Freeman, B.L.; Eggett, D.L.; Parker, T.L. Synergistic and antagonistic interactions of phenolic compounds found in navel oranges. J. Food Sci. 2010, 75, C570–C576. [Google Scholar] [CrossRef] [PubMed]
- Kopjar, M.; Lončarić, A.; Mikulinjak, M.; Šrajbek, Ž.; Šrajbek, M.; Pichler, A. Evaluation of antioxidant interactions of combined model systems of phenolics in the presence of sugars. Nat. Prod. Commun. 2016, 11, 1934578X1601101008. [Google Scholar] [CrossRef] [Green Version]
- Senizza, B.; Zhang, L.L.; Miras-Moreno, B.; Righetti, L.; Zengin, G.; Ak, G.; Bruni, R.; Lucini, L.; Sifola, M.I.; El-Nakhel, C.; et al. The Strength of the Nutrient Solution Modulates the Functional Profile of Hydroponically Grown Lettuce in a Genotype-Dependent Manner. Foods 2020, 9, 1156. [Google Scholar] [CrossRef]
- Xi, Y.; Jiao, W.; Cao, J.; Jiang, W. Effects of chlorogenic acid on capacity of free radicals scavenging and proteomic changes in postharvest fruit of nectarine. PLoS ONE 2017, 12, e0182494. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Cho, M.C.; Yang, E.Y.; Lee, J.G. Response to Salt Stress in Lettuce: Changes in Chlorophyll Fluorescence Parameters, Phytochemical Contents, and Antioxidant Activities. Agronomy 2020, 10, 1627. [Google Scholar] [CrossRef]
- Vetrano, F.; Moncada, A.; Miceli, A. Use of Gibberellic Acid to Increase the Salt Tolerance of Leaf Lettuce and Rocket Grown in a Floating System. Agron. 2020, 10, 505. [Google Scholar] [CrossRef] [Green Version]
- Arriola, N.D.A.; Chater, P.I.; Wilcox, M.; Lucini, L.; Rocchetti, G.; Dalmina, M.; Pearson, J.P.; de Mello Castanho Amboni, R.D. Encapsulation of stevia rebaudiana Bertoni aqueous crude extracts by ionic gelation–Effects of alginate blends and gelling solutions on the polyphenolic profile. Food Chem. 2019, 275, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. GigaScience 2013, 2, 2047-217X-2-13. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Aktumsek, A. Investigation of antioxidant potentials of solvent extracts from different anatomical parts of Asphodeline anatolica E. Tuzlaci: An endemic plant to Turkey. Afr. J. Tradit. Complementary Altern. Med. 2014, 11, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Zengin, G. A study on in vitro enzyme inhibitory properties of Asphodeline anatolica: New sources of natural inhibitors for public health problems. Ind. Crop. Prod. 2016, 83, 39–43. [Google Scholar] [CrossRef]
- Senizza, B.; Rocchetti, G.; Okur, M.A.; Zengin, G.; Yıldıztugay, E.; Ak, G.; Montesano, D.; Lucini, L. Phytochemical Profile and Biological Properties of Colchicum triphyllum (Meadow Saffron). Foods 2020, 9, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mimmo, T.; Tiziani, R.; Valentinuzzi, F.; Lucini, L.; Nicoletto, C.; Sambo, P.; Scampicchio, M.; Pii, Y.; Cesco, S. Selenium biofortification in Fragaria× ananassa: Implications on strawberry fruits quality, content of bioactive health beneficial compounds and metabolomic profile. Front. Plant Sci. 2017, 8, 1887. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Lucini, L.; Rodriguez, J.M.L.; Barba, F.J.; Giuberti, G. Gluten-free flours from cereals, pseudocereals and legumes: Phenolic fingerprints and in vitro antioxidant properties. Food Chem. 2019, 271, 157–164. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Source of Variance | Anthocyanins | Other Flavonoids | Flavonols | Lignans | Other Polyphenols | Phenolic Acids | Stilbenes |
|---|---|---|---|---|---|---|---|
| mg Eq. g−1 DM | mg Eq. g−1 DM | mg Eq. g−1 DM | mg Eq. g−1 DM | mg Eq. g−1 DM | mg Eq. g−1 DM | mg Eq. g−1 DM | |
| Salinity (S) | |||||||
| Standard | 4.05 ± 0.13 a | 1.08 ± 0.05 | 0.59 ± 0.07 a | 8.88 ± 0.46 a | 14.44 ± 1.07 | 1.97 ± 0.06 a | 0.99 ± 0.18 b |
| Saline | 3.85 ± 0.16 b | 1.15 ± 0.09 | 0.51 ± 0.06 b | 6.81 ± 0.58 b | 12.85 ± 0.78 | 1.71 ± 0.07 b | 1.45 ± 0.27 a |
| Phenolic application (P) | |||||||
| No application | 4.12 ± 0.12 a | 0.96 ± 0.07 c | 0.78 ± 0.02 a | 7.32 ± 1.07 | 14.86 ± 0.98 | 1.92 ± 0.08 | 0.58 ± 0.02 b |
| HES | 3.99 ± 0.15 a | 1.36 ± 0.06 a | 0.24 ± 0.01 d | 7.80 ± 0.84 | 13.22 ± 0.98 | 1.92 ± 0.10 | 1.27 ± 0.31 a |
| CGA | 4.18 ± 0.23 a | 1.15 ± 0.12 b | 0.63 ± 0.05 b | 7.50 ± 0.37 | 15.17 ± 1.94 | 1.87 ± 0.13 | 1.22 ± 0.24 a |
| HES + CGA | 3.53 ± 0.20 b | 0.99 ± 0.07 c | 0.54 ± 0.05 c | 8.76 ± 1.01 | 11.32 ± 0.79 | 1.66 ± 0.07 | 1.81 ± 0.45 a |
| S × P | |||||||
| Standard × No application | 3.89 ± 0.15 cd | 1.06 ± 0.08 bcd | 0.76 ± 0.02 a | 9.54 ± 0.15 ab | 14.45 ± 1.79 abc | 2.02 ± 0.05 | 0.61 ± 0.03 c |
| Standard × HES | 3.71 ± 0.12 d | 1.25 ± 0.06 ab | 0.22 ± 0.01 e | 7.04 ± 0.03 bcd | 12.09 ± 1.87 bc | 2.02 ± 0.12 | 1.64 ± 0.59 b |
| Standard × CGA | 4.70 ± 0.07 a | 0.90 ± 0.09 cd | 0.74 ± 0.02 a | 8.10 ± 0.22 bc | 18.70 ± 1.82 a | 2.05 ± 0.20 | 0.80 ± 0.11 bc |
| Standard × HES + CGA | 3.92 ± 0.14 cd | 1.12 ± 0.07 bc | 0.65 ± 0.02 b | 10.85 ± 0.67 a | 12.51 ± 1.27 bc | 1.80 ± 0.09 | 0.89 ± 0.24 bc |
| Saline × No application | 4.35 ± 0.05 ab | 0.86 ± 0.08 d | 0.80 ± 0.03 a | 5.10 ± 0.85 d | 15.28 ± 1.21 ab | 1.82 ± 0.14 | 0.55 ± 0.03 c |
| Saline × HES | 4.27 ± 0.16 bc | 1.47 ± 0.06 a | 0.27 ± 0.01 e | 8.56 ± 1.73 abc | 14.35 ± 0.18 abc | 1.83 ± 0.17 | 0.90 ± 0.03 bc |
| Saline × CGA | 3.67 ± 0.07 d | 1.40 ± 0.07 a | 0.52 ± 0.02 c | 6.91 ± 0.55 cd | 11.64 ± 1.77 bc | 1.69 ± 0.12 | 1.64 ± 0.31 b |
| Saline × HES + CGA | 3.14 ± 0.18 e | 0.86 ± 0.06 d | 0.44 ± 0.01 d | 6.67 ± 0.55 cd | 10.13 ± 0.30 c | 1.52 ± 0.03 | 2.72 ± 0.35 a |
| Significance | |||||||
| S | * | ns | *** | ** | ns | ** | * |
| P | *** | *** | *** | ns | ns | ns | ** |
| S × P | *** | *** | *** | ** | * | ns | ** |
| Source of Variance | DPPH | ABTS | FRAP | Metal Chelating | Phosphomolybdenum |
|---|---|---|---|---|---|
| mgTE g−1 | mgTE g−1 | mgTE g−1 | mg EDTAE g−1 | mmol TE g−1 | |
| Salinity (S) | |||||
| Standard | 1.17 ± 0.13 b | 8.86 ± 0.14 a | 15.37 ± 0.14 a | 30.50 ± 0.78 a | 0.87 ± 0.02 |
| Saline | 2.49 ± 0.34 a | 8.10 ± 0.47 b | 12.71 ± 0.53 b | 26.46 ± 1.38 b | 0.90 ± 0.04 |
| Phenolic application (P) | |||||
| No application | 0.94 ± 0.00 b | 8.12 ± 0.38 b | 14.59 ± 0.31 b | 25.94 ± 1.58 c | 0.83 ± 0.03 b |
| HES | 2.54 ± 0.56 a | 9.59 ± 0.44 a | 12.93 ± 1.26 c | 28.55 ± 1.86 b | 0.97 ± 0.06 a |
| CGA | 2.24 ± 0.46 a | 8.22 ± 0.15 b | 13.50 ± 0.54 c | 30.51 ± 1.68 a | 0.84 ± 0.02 b |
| HES + CGA | 1.48 ± 0.31 b | 7.99 ± 0.68 b | 15.16 ± 0.41 a | 28.90 ± 1.85 b | 0.90 ± 0.02 ab |
| S × P | |||||
| Standard × No application | nd | 8.89 ± 0.27 c | 15.28 ± 0.07 ab | 29.47 ± 0.10 c | 0.86 ± 0.03 bc |
| Standard × HES | 1.39 ± 0.27 | 8.62 ± 0.09 c | 15.72 ± 0.07 a | 32.71 ± 0.18 b | 0.85 ± 0.05 bc |
| Standard × CGA | 1.25 ± 0.20 | 8.42 ± 0.09 cd | 14.68 ± 0.09 bc | 26.77 ± 0.27 d | 0.84 ± 0.01 bc |
| Standard × HES + CGA | 0.88 ± 0.08 | 9.50 ± 0.09 b | 15.81 ± 0.19 a | 33.02 ± 0.36 b | 0.94 ± 0.01 b |
| Saline × No application | 0.94 ± 0.00 | 7.35 ± 0.23 e | 13.89 ± 0.01 c | 22.40 ± 0.07 f | 0.81 ± 0.07 c |
| Saline × HES | 3.68 ± 0.40 | 10.56 ± 0.08 a | 10.13 ± 0.30 e | 24.40 ± 0.09 e | 1.09 ± 0.03 a |
| Saline × CGA | 3.24 ± 0.10 | 8.03 ± 0.24 d | 12.31 ± 0.18 d | 34.24 ± 0.13 a | 0.85 ± 0.05 bc |
| Saline × HES + CGA | 2.08 ± 0.34 | 6.48 ± 0.08 f | 14.52 ± 0.63 bc | 24.79 ± 0.01 e | 0.85 ± 0.01 bc |
| Significance | |||||
| S | *** | *** | *** | *** | ns |
| P | *** | *** | *** | *** | ** |
| S × P | ns | *** | *** | *** | ** |
| Source of Variance | AChE | BChE | Tyrosinase | α-Amylase | α-Glucosidase |
|---|---|---|---|---|---|
| mg GALAE g−1 | mg GALAE g−1 | mg KAE g−1 | mmol ACAE g−1 | mmol ACAE | |
| Salinity (S) | |||||
| Standard | 2.06 ± 0.08 a | 2.48 ± 0.09 a | 46.80 ± 0.78 | 0.36 ± 0.01 b | 1.05 ± 0.01 a |
| Saline | 1.93 ± 0.06 b | 1.81 ± 0.10 b | 47.59 ± 0.57 | 0.39 ± 0.01 a | 0.83 ± 0.09 b |
| Phenolic application (P) | |||||
| No application | 1.79 ± 0.08 c | 2.20 ± 0.12 | 46.16 ± 0.69 c | 0.38 ± 0.01 a | 1.05 ± 0.01 a |
| HES | 1.89 ± 0.12 c | 2.19 ± 0.18 | 45.17 ± 0.65 c | 0.36 ± 0.01 b | 0.71 ± 0.16 b |
| CGA | 2.05 ± 0.04 b | 1.95 ± 0.21 | 49.65 ± 0.49 a | 0.37 ± 0.01 b | 0.97 ± 0.04 a |
| HES + CGA | 2.26 ± 0.06 a | 2.25 ± 0.26 | 47.79 ± 0.88 b | 0.39 ± 0.01 a | 1.02 ± 0.02 a |
| S × P | |||||
| Standard × No application | 1.67 ± 0.12 d | 2.26 ± 0.12 bc | 46.81 ± 0.88 b | 0.38 ± 0.01 bcd | 1.04 ± 0.01 a |
| Standard × HES | 2.13 ± 0.05 abc | 2.54 ± 0.21 ab | 43.93 ± 0.72 c | 0.33 ± 0.01 f | 1.06 ± 0.01 a |
| Standard × CGA | 2.11 ± 0.05 abc | 2.30 ± 0.06 bc | 50.56 ± 0.51 a | 0.35 ± 0.01 e | 1.03 ± 0.01 a |
| Standard × HES + CGA | 2.34 ± 0.10 a | 2.83 ± 0.10 a | 45.88 ± 0.39 bc | 0.37 ± 0.01 d | 1.06 ± 0.01 a |
| Saline × No application | 1.91 ± 0.02 c | 2.14 ± 0.23 bcd | 45.51 ± 1.08 bc | 0.38 ± 0.01 cd | 1.06 ± 0.01 a |
| Saline × HES | 1.66 ± 0.10 d | 1.85 ± 0.05 cde | 46.42 ± 0.20 b | 0.39 ± 0.01 b | 0.37 ± 0.10 b |
| Saline × CGA | 1.99 ± 0.03 bc | 1.59 ± 0.29 de | 48.74 ± 0.34 a | 0.39 ± 0.01 bc | 0.91 ± 0.07 a |
| Saline × HES + CGA | 2.17 ± 0.02 ab | 1.67 ± 0.03 e | 49.69 ± 0.35 a | 0.41 ± 0.01 a | 0.99 ± 0.03 a |
| Significance | |||||
| S | * | *** | ns | *** | *** |
| P | *** | ns | *** | *** | *** |
| S × P | ** | * | *** | *** | *** |
| Groups | Treatments |
|---|---|
| Control | |
| Hesperidin | 100 µM hesperidin |
| Chlorogenic acid | 50 µM chlorogenic acid |
| Hesperidin + Chlorogenic acid | 100 µM hesperidin + 50 µM chlorogenic acid |
| Salinity | 40 mM NaCl |
| Salinity—Hesperidin | 40 mM NaCl + 100 µM HES |
| Salinity—Chlorogenic acid | 40 mM NaCl + 50 µM CGA |
| Salinity—Hesperidin + Chlorogenic acid | 40 mM NaCl + 100 µM HES + 50 µM CGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Martinelli, E.; Senizza, B.; Miras-Moreno, B.; Yildiztugay, E.; Arikan, B.; Elbasan, F.; Ak, G.; Balci, M.; Zengin, G.; et al. The Combination of Mild Salinity Conditions and Exogenously Applied Phenolics Modulates Functional Traits in Lettuce. Plants 2021, 10, 1457. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071457
Zhang L, Martinelli E, Senizza B, Miras-Moreno B, Yildiztugay E, Arikan B, Elbasan F, Ak G, Balci M, Zengin G, et al. The Combination of Mild Salinity Conditions and Exogenously Applied Phenolics Modulates Functional Traits in Lettuce. Plants. 2021; 10(7):1457. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071457
Chicago/Turabian StyleZhang, Leilei, Erika Martinelli, Biancamaria Senizza, Begoña Miras-Moreno, Evren Yildiztugay, Busra Arikan, Fevzi Elbasan, Gunes Ak, Melike Balci, Gokhan Zengin, and et al. 2021. "The Combination of Mild Salinity Conditions and Exogenously Applied Phenolics Modulates Functional Traits in Lettuce" Plants 10, no. 7: 1457. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10071457