
Quantitative Real-Time PCR Assay for the Detection of Pectobacterium parmentieri, a Causal Agent of Potato Soft Rot

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
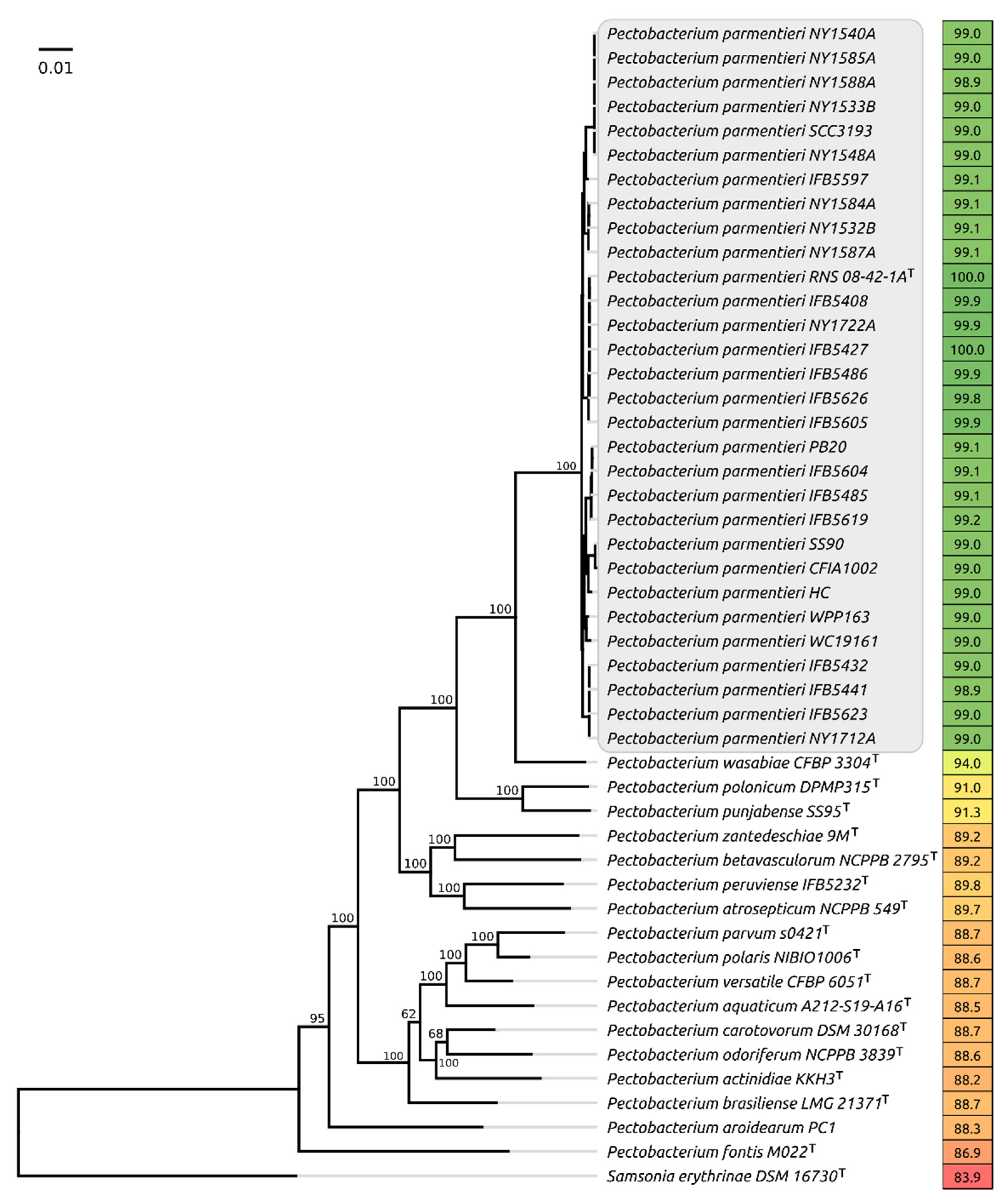
2.1. Phylogenetic Analysis
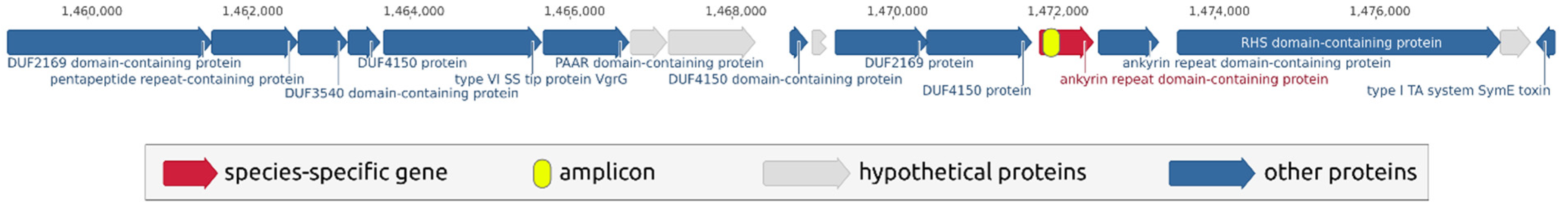
2.2. Search for Species-Specific Primers
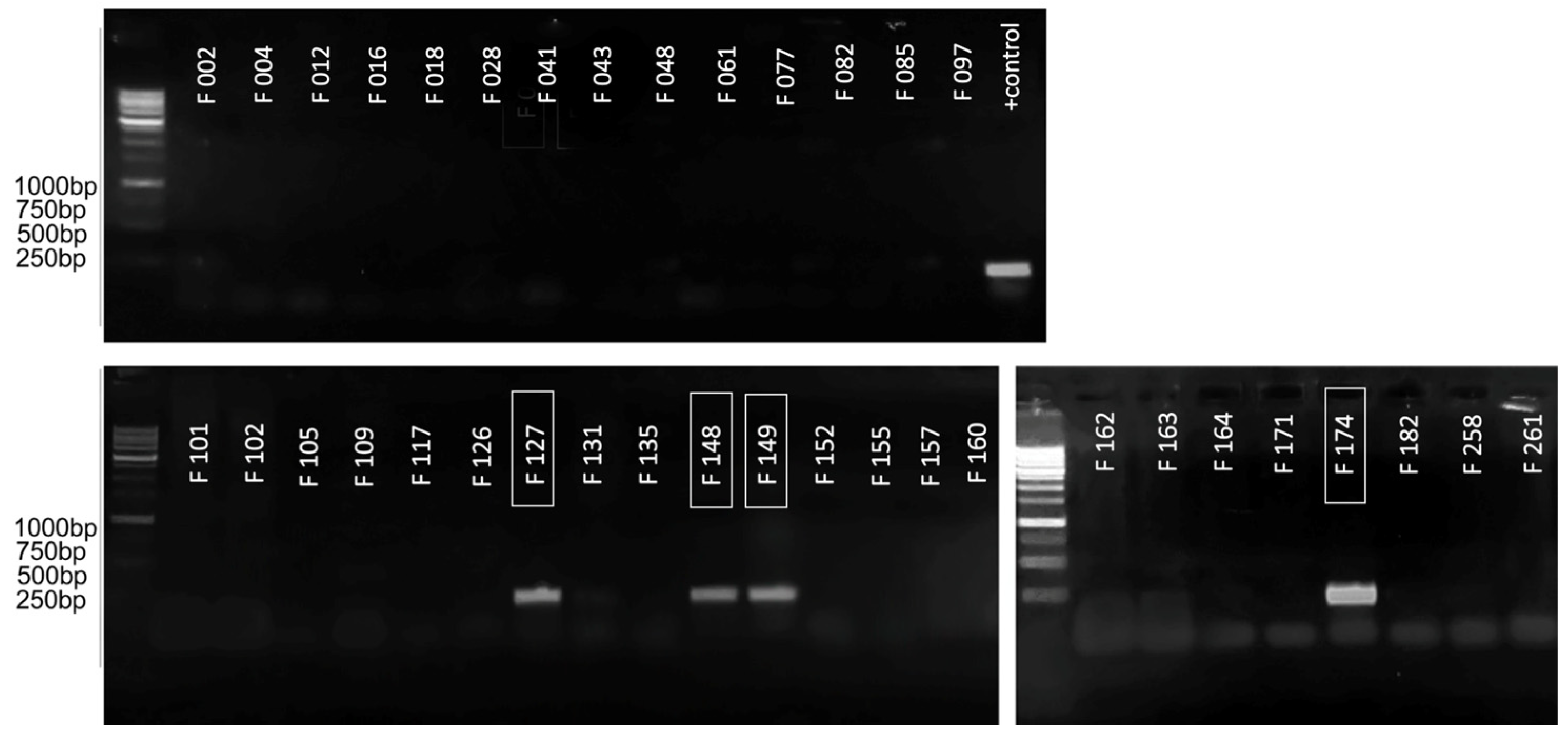
2.3. Primary Analysis by Conventional PCR
2.4. qPCR Analysis on an Extended Set of Strains
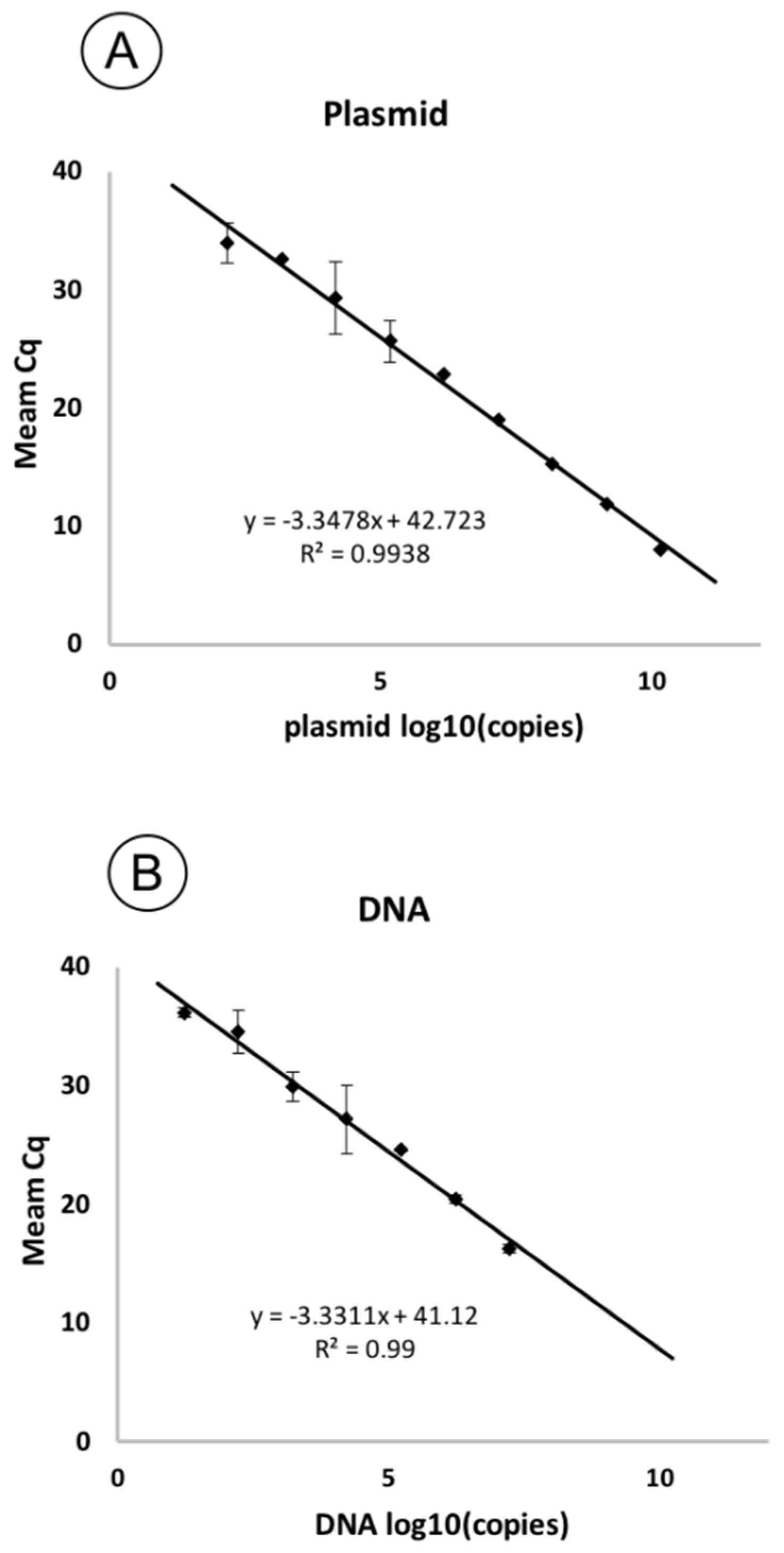
2.5. Sensitivity
2.6. Assays of Plant Samples
3. Discussion
4. Materials and Methods
4.1. Phylogenetic Analysis
4.2. Search for Species-Specific Sequences and Primer Design
4.3. Bacterial Strains, Media and Culture Conditions
4.4. Genomic DNA Isolation
4.5. PCR Conditions
4.6. Plasmid Construction for Sensitivity Assay
4.7. qPCR
4.8. Testing the Detection System on Artificially Infected Tubers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bhat, K.A.; Masood, S.; Bhat, N.; Bhat, M.A.; Razvi, S.; Mir, M.; Akhtar, S.; Wani, N.; Habib, M. Current Status of Post Harvest Soft Rot in Vegetables: A Review. Asian J. Plant Sci. 2010, 9, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khayi, S.; Cigna, J.; Chong, T.M.; Quêtu-Laurent, A.; Chan, K.-G.; Hélias, V.; Faure, D. Transfer of the potato plant isolates of Pectobacterium wasabiae to Pectobacterium parmentieri sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5379–5383. [Google Scholar] [CrossRef] [PubMed]
- Suarez, B.; Feria, F.J.; Martín-Robles, M.J.; Del Rey, F.J.; Palomo, J.L. Pectobacterium parmentieri Causing Soft Rot on Potato Tubers in Southern Europe. Plant Dis. 2017, 101, 1029. [Google Scholar] [CrossRef]
- Ha, V.T.N.; Voronina, M.V.; Kabanova, A.P.; Shneider, M.M.; Korzhenkov, A.A.; Toschakov, S.V.; Miroshnikov, K.; Ignatov, A.N. First Report of Pectobacterium parmentieri Causing Stem Rot Disease of Potato in Russia. Plant Dis. 2019, 103, 144. [Google Scholar] [CrossRef]
- Kamau, J.W.; Ngaira, J.; Kinyua, J.; Gachamba, S.; Ngundo, G.; Janse, J.; Macharia, I. Occurence of pectinolytic bacteria causing blackleg and soft rot of potato in Kenya. J. Plant Pathol. 2019, 101, 689–694. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Sun, Q.; Feng, Z.; Handique, U.; Wu, J.; Li, M.W.; Zhang, R. First Report of Pectobacterium parmentieri Causing Blackleg on Potato in Inner Mongolia, China. Plant Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Sarfraz, S.; Riaz, K.; Oulghazi, S.; Cigna, J.; Sahi, S.T.; Khan, S.; Hameed, A.; Alam, M.W.; Faure, D. First Report of Pectobacterium parmentieri and Pectobacterium polaris Causing Potato Blackleg Disease in Punjab, Pakistan. Plant Dis. 2019, 103, 1405. [Google Scholar] [CrossRef]
- Ge, T.L.; Jiang, H.H.; Hao, J.J.; Johnson, S.B. First Report of Pectobacterium parmentieri Causing Bacterial Soft Rot and Blackleg on Potato in Maine. Plant Dis. 2018, 102, 437. [Google Scholar] [CrossRef]
- McNally, R.R.; Curland, R.; Webster, B.T.; Robinson, A.P.; Ishimaru, C.A. First Report of Blackleg and Tuber Soft Rot of Potato Caused by Pectobacterium parmentieri in Minnesota and North Dakota. Plant Dis. 2017, 101, 2144. [Google Scholar] [CrossRef]
- Zoledowska, S.; Motyka, A.; Zukowska, D.; Sledz, W.; Lojkowska, E. Population Structure and Biodiversity of Pectobacterium parmentieri Isolated from Potato Fields in Temperate Climate. Plant Dis. 2018, 102, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Zoledowska, S.; Motyka-Pomagruk, A.; Sledz, W.; Mengoni, A.; Lojkowska, E. High genomic variability in the plant pathogenic bacterium Pectobacterium parmentieri deciphered from de novo assembled complete genomes. BMC Genom. 2018, 19, 751. [Google Scholar] [CrossRef] [Green Version]
- Waleron, M.; Misztak, A.; Waleron, M.; Jonca, J.; Furmaniak, M.; Waleron, K. Pectobacterium polonicum sp. nov. isolated from vegetable fields. Int. J. Syst. Evol. Microbiol. 2019, 69, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Sarfraz, S.; Riaz, K.; Oulghazi, S.; Cigna, J.; Sahi, S.T.; Khan, S.; Faure, D. Pectobacterium punjabense sp. nov., isolated from blackleg symptoms of potato plants in Pakistan. Int. J. Syst. Evol. Microbiol. 2018, 68, 3551–3556. [Google Scholar] [CrossRef] [PubMed]
- Cigna, J.; Laurent, A.; Waleron, M.; Waleron, K.; Dewaegeneire, P.; van der Wolf, J.; Andrivon, D.; Faure, D.; Hélias, V. European Population of Pectobacterium punjabense: Genomic Diversity, Tuber Maceration Capacity and a Detection Tool for This Rarely Occurring Potato Pathogen. Microorganisms 2021, 9, 781. [Google Scholar] [CrossRef] [PubMed]
- Humphris, S.N.; Cahill, G.; Elphinstone, J.G.; Kelly, R.; Parkinson, N.M.; Pritchard, L.; Toth, I.K.; Saddler, G.S. Detection of the Bacterial Potato Pathogens Pectobacterium and Dickeya spp. Using Conventional and Real-Time PCR. In Plant Pathology; Humana Press: New York, NY, USA, 2015; Volume 1302, pp. 1–16. [Google Scholar]
- van der Wolf, J.M.; Cahill, G.; Van Gijsegem, F.; Helias, V.; Humphris, S.; Li, X.; Lojkowska, E.; Pritchard, L. Isolation, Detection and Characterization of Pectobacterium and Dickeya Species. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Springer International Publishing: Cham, Switzerland, 2021; pp. 149–173. [Google Scholar]
- De Boer, S.H.; Li, X.; Ward, L.J. Pectobacterium spp. Associated with Bacterial Stem Rot Syndrome of Potato in Canada. Phytopathology 2012, 102, 937–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.H.; Cho, M.S.; Kim, B.K.; Choi, H.J.; Hahn, J.H.; Kim, C.; Kang, M.J.; Kim, S.H.; Park, D.S. Quantitative Real-Time Polymerase Chain Reaction Assay for Detection of Pectobacterium wasabiae Using YD Repeat Protein Gene-Based Primers. Plant Dis. 2012, 96, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Lukianova, A.; Evseev, P.; Stakheev, A.; Kotova, I.; Zavriev, S.; Ignatov, A.; Miroshnikov, K. Development of qPCR Detection Assay for Potato Pathogen Pectobacterium atrosepticum Based on a Unique Target Sequence. Plants 2021, 10, 355. [Google Scholar] [CrossRef]
- De Campos, S.B.; Lardi, M.; Gandolfi, A.; Eberl, L.; Pessi, G. Mutations in Two Paraburkholderia phymatum Type VI Secretion Systems Cause Reduced Fitness in Interbacterial Competition. Front. Microbiol. 2017, 8, 2473. [Google Scholar] [CrossRef] [Green Version]
- Van Der Wolf, J.M.; Acuña, I.; De Boer, S.H.; Brurberg, M.B.; Cahill, G.; Charkowski, A.O.; Coutinho, T.; Davey, T.; Dees, M.W.; Degefu, Y.; et al. Diseases Caused by Pectobacterium and Dickeya Species around the World. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Springer Nature: Cham, Switzerland, 2021; pp. 215–261. [Google Scholar]
- Van Gijsegem, F.; Hugouvieux-Cotte-Pattat, N.; Kraepiel, Y.; Lojkowska, E.; Moleleki, L.N.; Gorshkov, V.; Yedidia, I. Molecular Interactions of Pectobacterium and Dickeya with Plants. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Springer Nature: Cham, Switzerland, 2021; pp. 85–147. [Google Scholar]
- Moleleki, L.N.; Onkendi, E.M.; Mongae, A.; Kubheka, G.C. Characterisation of Pectobacterium wasabiae causing blackleg and soft rot diseases in South Africa. Eur. J. Plant Pathol. 2013, 135, 279–288. [Google Scholar] [CrossRef]
- van den Bosch, T.J.M.; Niemi, O.; Welte, C.U. Single gene enables plant pathogenic Pectobacterium to overcome host-specific chemical defence. Mol. Plant Pathol. 2020, 21, 349–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kõiv, V.; Roosaare, M.; Vedler, E.; Kivistik, P.A.; Toppi, K.; Schryer, D.W.; Remm, M.; Tenson, T.; Mäe, A. Microbial population dynamics in response to Pectobacterium atrosepticum infection in potato tubers. Sci. Rep. 2015, 5, 11606. [Google Scholar] [CrossRef] [PubMed]
- Ge, T.; Ekbataniamiri, F.; Johnson, S.; Larkin, R.; Hao, J. Interaction between Dickeya dianthicola and Pectobacterium parmentieri in Potato Infection under Field Conditions. Microorganisms 2021, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.; Hommais, F.; Nasser, H.W.; Reverchon, S. Plant-phytopathogen interactions: Bacterial responses to environmental and plant stimuli. Environ. Microbiol. 2017, 19, 1689–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charkowski, A.O. The Changing Face of Bacterial Soft-Rot Diseases. Annu. Rev. Phytopathol. 2018, 56, 269–288. [Google Scholar] [CrossRef]
- van der Wolf, J.M.; De Boer, S.H.; Czajkowski, R.; Cahill, G.; Van Gijsegem, F.; Davey, T.; Dupuis, B.; Ellicott, J.; Jafra, S.; Kooman, M.; et al. Management of Diseases Caused by Pectobacterium and Dickeya Species. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Springer: Cham, Switzerland, 2021; pp. 175–214. [Google Scholar]
- Holtappels, D.; Fortuna, K.; Lavigne, R.; Wagemans, J. The future of phage biocontrol in integrated plant protection for sustainable crop production. Curr. Opin. Biotechnol. 2021, 68, 60–71. [Google Scholar] [CrossRef]
- Svircev, A.; Roach, D.; Castle, A. Framing the Future with Bacteriophages in Agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef] [Green Version]
- Smolarska, A.; Rabalski, L.; Narajczyk, M.; Czajkowski, R. Isolation and phenotypic and morphological characterization of the first Podoviridae lytic bacteriophages ϕA38 and ϕA41 infecting Pectobacterium parmentieri (former Pectobacterium wasabiae). Eur. J. Plant Pathol. 2018, 150, 413–425. [Google Scholar] [CrossRef]
- Kabanova, A.; Shneider, M.; Bugaeva, E.; Ha, V.T.N.; Miroshnikov, K.; Korzhenkov, A.; Kulikov, E.; Toschakov, S.; Ignatov, A.; Miroshnikov, K. Genomic characteristics of vB_PpaP_PP74, a T7-like Autographivirinae bacteriophage infecting a potato pathogen of the newly proposed species Pectobacterium parmentieri. Arch. Virol. 2018, 163, 1691–1694. [Google Scholar] [CrossRef]
- Charkowski, A.; Blanco, C.; Condemine, G.; Expert, D.; Franza, T.; Hayes, C.; Cotte-Pattat, N.; Solanilla, E.A.L.; Low, D.; Moleleki, L.; et al. The Role of Secretion Systems and Small Molecules in Soft-Rot Enterobacteriaceae Pathogenicity. Annu. Rev. Phytopathol. 2012, 50, 425–449. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Hibbing, M.E.; Kim, H.-S.; Reedy, R.M.; Yedidia, I.; Breuer, J.; Breuer, J.; Glasner, J.D.; Perna, N.T.; Kelman, A.; et al. Host Range and Molecular Phylogenies of the Soft Rot Enterobacterial Genera Pectobacterium and Dickeya. Phytopathology 2007, 97, 1150–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-S.; Ma, B.; Perna, N.T.; Charkowski, A.O. Phylogeny and Virulence of Naturally Occurring Type III Secretion System-Deficient Pectobacterium Strains. Appl. Environ. Microbiol. 2009, 75, 4539–4549. [Google Scholar] [CrossRef] [Green Version]
- Nykyri, J.; Mattinen, L.; Niemi, O.; Adhikari, S.; Kõiv, V.; Somervuo, P.; Fang, X.; Auvinen, P.; Mäe, A.; Palva, E.T.; et al. Role and Regulation of the Flp/Tad Pilus in the Virulence of Pectobacterium atrosepticum SCRI1043 and Pectobacterium wasabiae SCC3193. PLoS ONE 2013, 8, e73718. [Google Scholar] [CrossRef] [Green Version]
- Toth, I.K.; Barny, M.-A.; Brurberg, M.B.; Condemine, G.; Czajkowski, R.; Elphinstone, J.G.; Helias, V.; Johnson, S.B.; Moleleki, L.N.; Pirhonen, M.; et al. Pectobacterium and Dickeya: Environment to Disease Development. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Springer: Cham, Switzerland, 2021; pp. 39–84. [Google Scholar]
- Bugaeva, E.; Voronina, M.; Vasiliev, D.; Lukianova, A.; Landyshev, N.; Ignatov, A.; Miroshnikov, K. Use of a Specific Phage Cocktail for Soft Rot Control on Ware Potatoes: A Case Study. Viruses 2021, 13, 1095. [Google Scholar] [CrossRef] [PubMed]
- Na, S.-I.; Kim, Y.O.; Yoon, S.-H.; Ha, S.-M.; Baek, I.; Chun, J. UBCG: Up-to-date bacterial core gene set and pipeline for phylogenomic tree reconstruction. J. Microbiol. 2018, 56, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Kim, Y.O.; Park, S.-C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Stakheev, A.A.; Khairulina, D.R.; Zavriev, S.K. Four-locus phylogeny of Fusarium avenaceum and related species and their species-specific identification based on partial phosphate permease gene sequences. Int. J. Food Microbiol. 2016, 225, 27–37. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
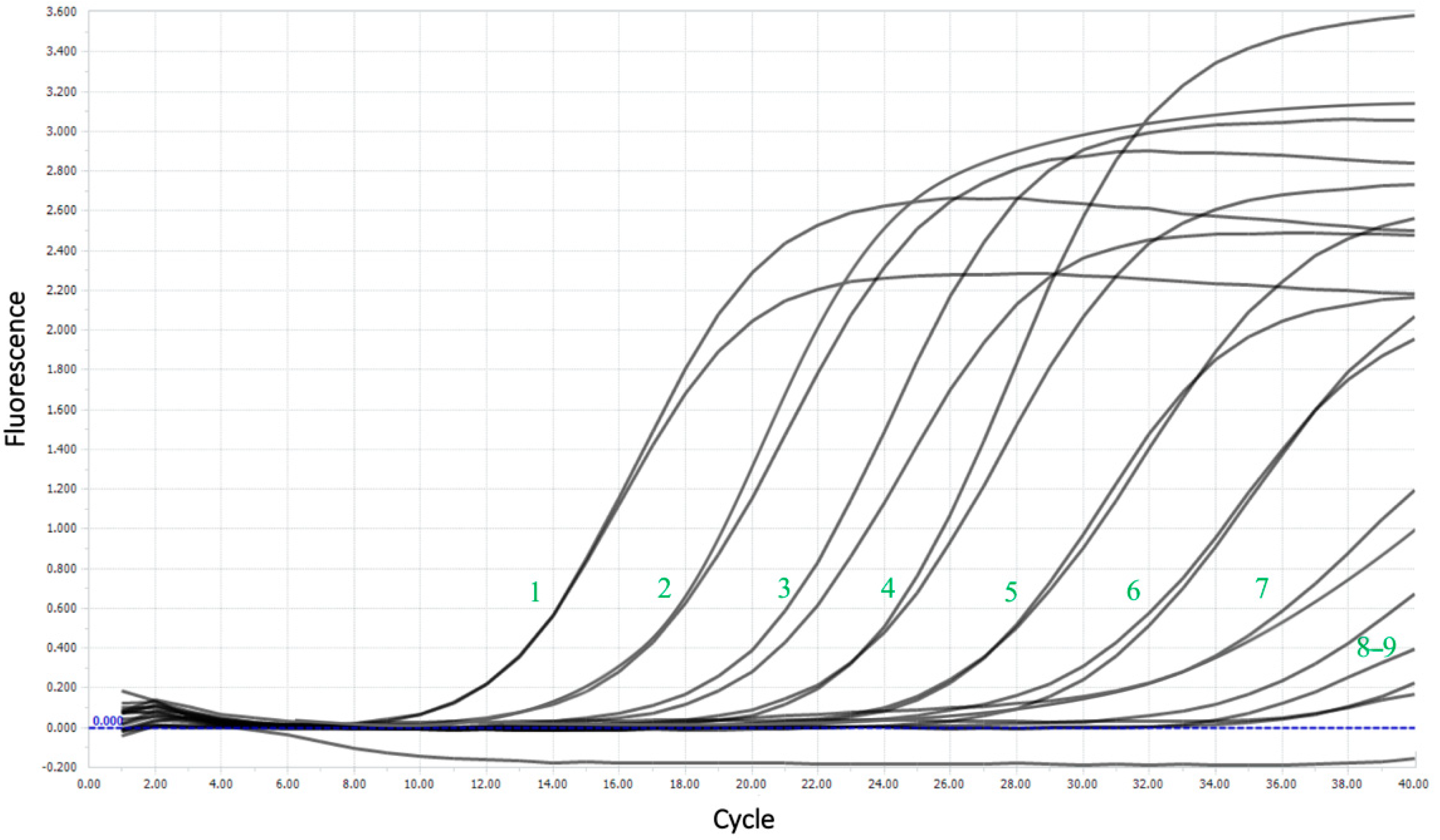{kind=link}
| Type | Sequence |
|---|---|
| F primer | TAT CGC TGG CTC AGG CAA TT |
| R primer | TAC GCT GCG CAT ACT TGG AA |
| Probe | (6-FAM)-CGCCCGGG-(dT-BHQ-1)-GCCCAAGATATGACTT-(Pi) |
| Amplicon | TATCGCTGGCTCAGGCAATTGAAAAAAACGATGAATCAGAAGTCAAAAAACTTTCAGCTCATACGGACCTAAATCGCCCGGGTGCCCAAGATATGACTTTACTATTCTTCGCGATGCAAAATAGTTATGACAAACAAGCCAAACATTTGTCGATAGTCTCATATTTGGTTAGTGCCGGAGCAAGTCCATTACAGAAAGTTCCAAGTATGCGCAGCGTA |
| № | Plasmid | DNA | ||||
|---|---|---|---|---|---|---|
| Copies Per Reaction | Cq | SD | Copies Per Reaction | Cq | SD | |
| 1 | 1.4 × 1010 | 8.01 | 0.12 | 1.68 × 107 | 16.29 | 0.37 |
| 2 | 1.48 × 109 | 11.83 | 0.21 | 1.68 × 106 | 20.48 | 0.36 |
| 3 | 1.48 × 108 | 15.27 | 0.15 | 1.68 × 105 | 24.62 | 0.01 |
| 4 | 1.48 × 107 | 19.04 | 0.71 | 1.68 × 104 | 27.20 | 2.90 |
| 5 | 1.48 × 106 | 22.90 | 0.57 | 1.68 × 103 | 29.92 | 1.25 |
| 6 | 1.48 × 105 | 25.67 | 1.72 | 168 | 34.60 | 1.80 |
| 7 | 1.48 × 104 | 29.37 | 3.07 | 16.8 | 36.20 | 0.37 |
| 8 | 1.48 × 103 | 32.59 | 0.70 | 1.68 | - | - |
| 9 | 148 | 33.95 | 1.72 | 0.16 | - | - |
| Incubation, h | Cq | SD | Copies/mL |
|---|---|---|---|
| Control (120 h) | 38.32 | 0.00 | 6.92 |
| 72 | 31.03 | 0.67 | 1.06 × 103 |
| 96 | 27.38 | 0.67 | 1.3 × 104 |
| 120 | 20.76 | 1.05 | 1.3 × 106 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukianova, A.A.; Evseev, P.V.; Stakheev, A.A.; Kotova, I.B.; Zavriev, S.K.; Ignatov, A.N.; Miroshnikov, K.A. Quantitative Real-Time PCR Assay for the Detection of Pectobacterium parmentieri, a Causal Agent of Potato Soft Rot. Plants 2021, 10, 1880. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091880
Lukianova AA, Evseev PV, Stakheev AA, Kotova IB, Zavriev SK, Ignatov AN, Miroshnikov KA. Quantitative Real-Time PCR Assay for the Detection of Pectobacterium parmentieri, a Causal Agent of Potato Soft Rot. Plants. 2021; 10(9):1880. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091880
Chicago/Turabian StyleLukianova, Anna A., Peter V. Evseev, Alexander A. Stakheev, Irina B. Kotova, Sergey K. Zavriev, Alexander N. Ignatov, and Konstantin A. Miroshnikov. 2021. "Quantitative Real-Time PCR Assay for the Detection of Pectobacterium parmentieri, a Causal Agent of Potato Soft Rot" Plants 10, no. 9: 1880. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091880