
Variation in Phytochemical, Antioxidant and Volatile Composition of Pomelo Fruit (Citrus grandis (L.) Osbeck) during Seasonal Growth and Development

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Collection
2.3. Juice Extraction and Extract Preparation of Seeds, Membrane, Albedo, Flavedo, and Pulp
2.4. Spectrophotometric Determination of Phytochemical Properties
2.5. Determination of Naringin Content
2.6. Ferric Reducing Antioxidant Power (FRAP), DPPH Free Radical Scavenging, and Antioxidant Capacity
2.7. Extraction of Volatile Compounds and GC-MS/FID Analysis
2.8. Statistical Analysis
3. Results and Discussion
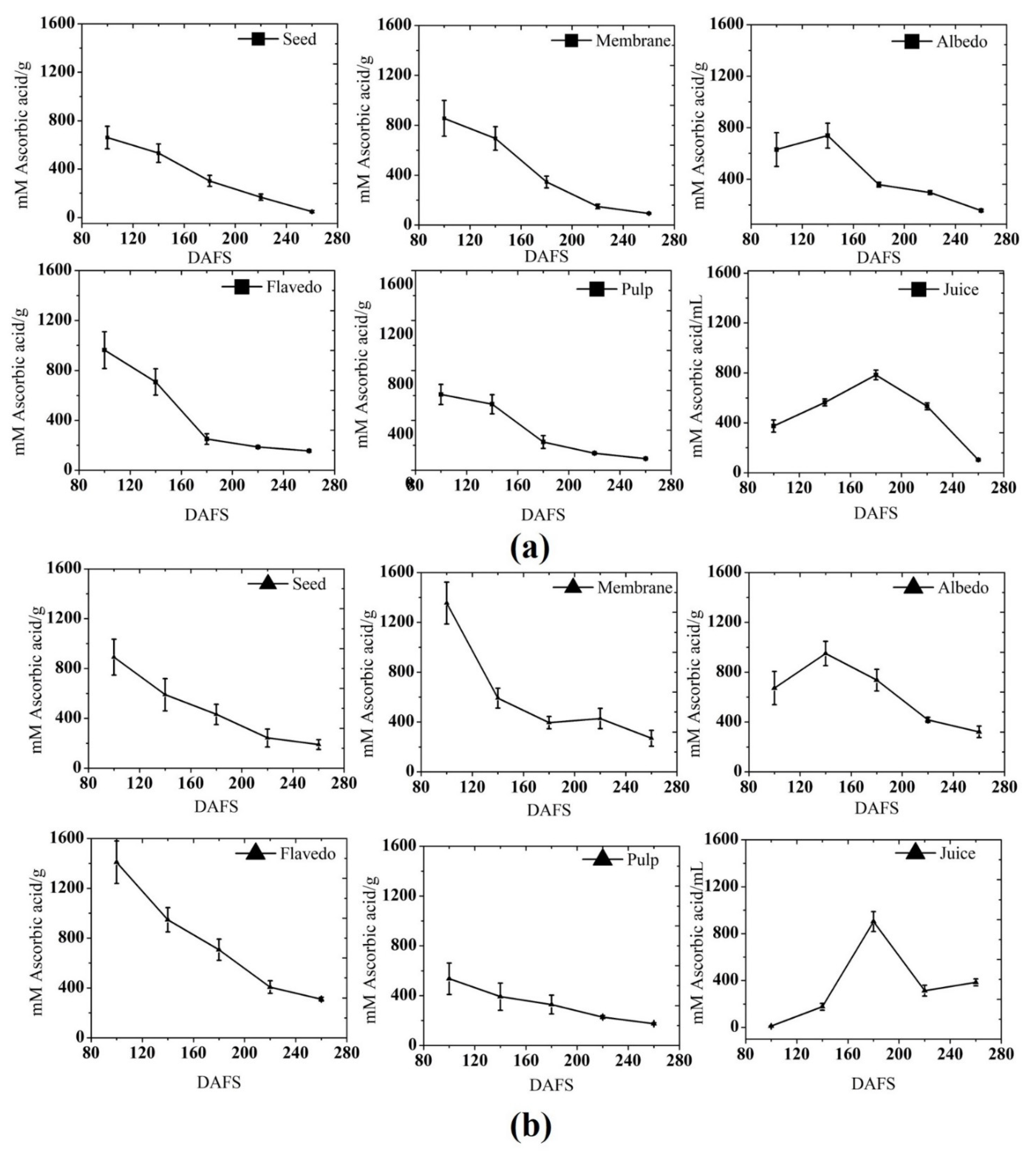
3.1. Phytochemical Content
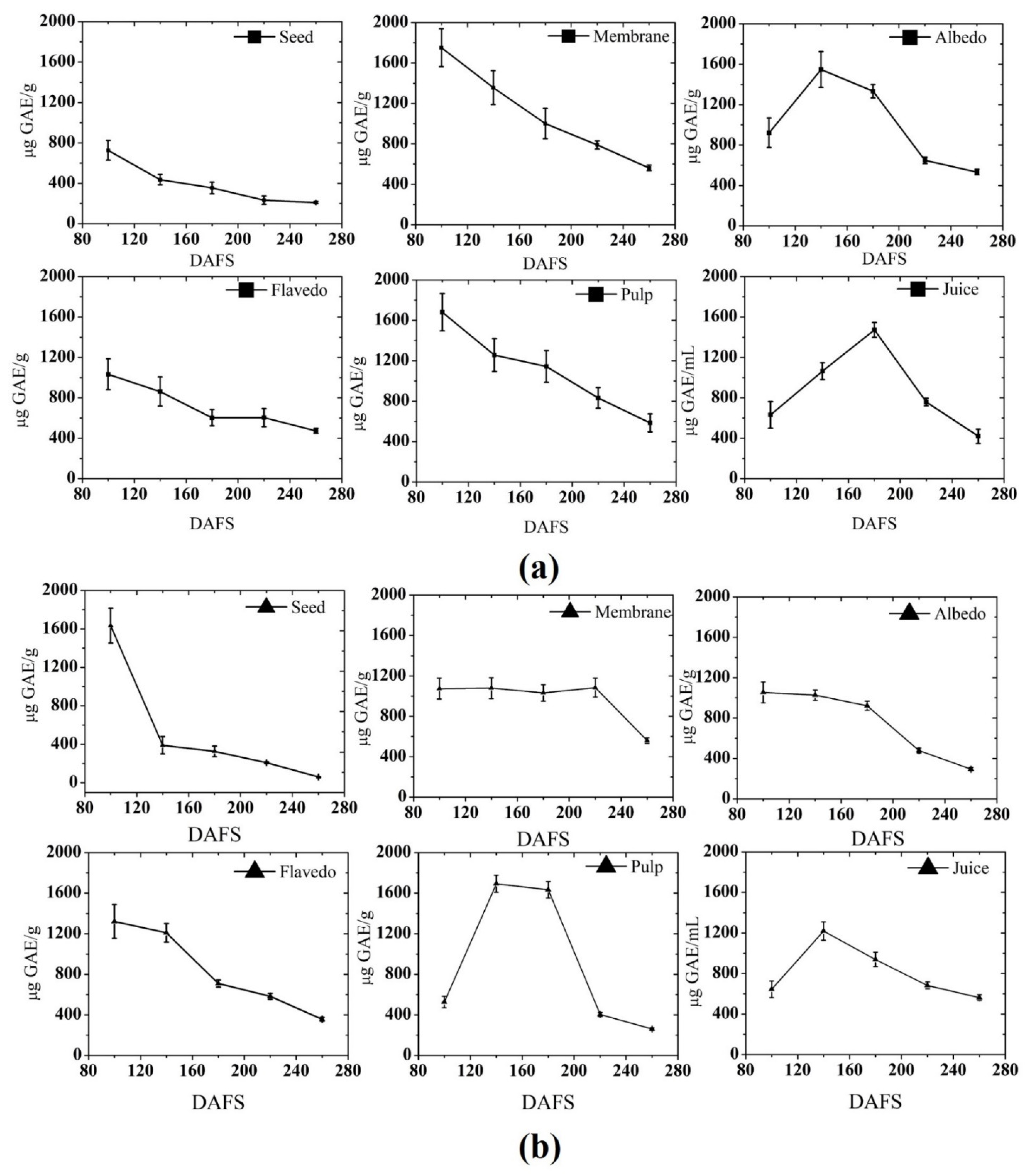
3.1.1. Total Phenolic Content
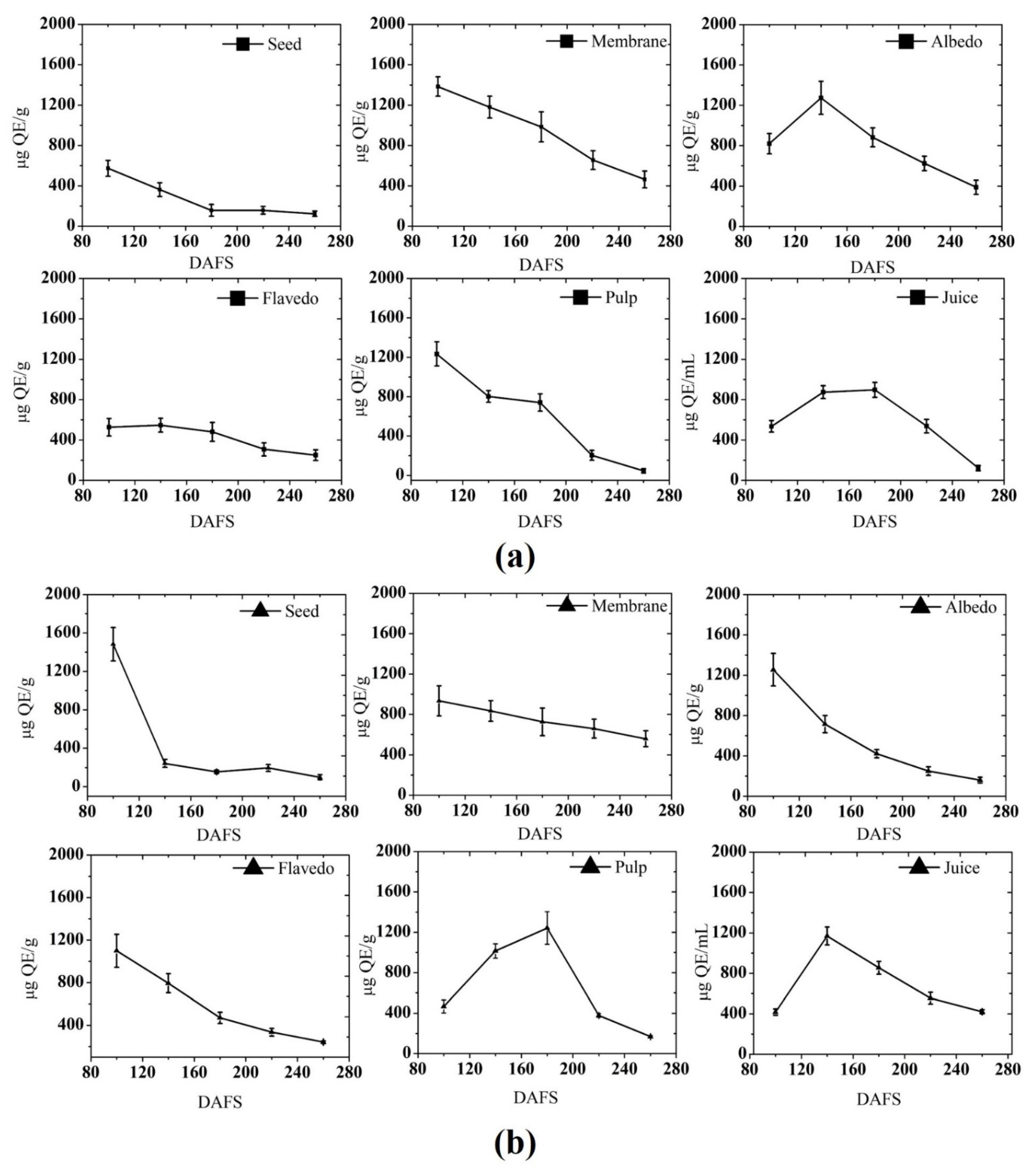
3.1.2. Total Flavonoid Content
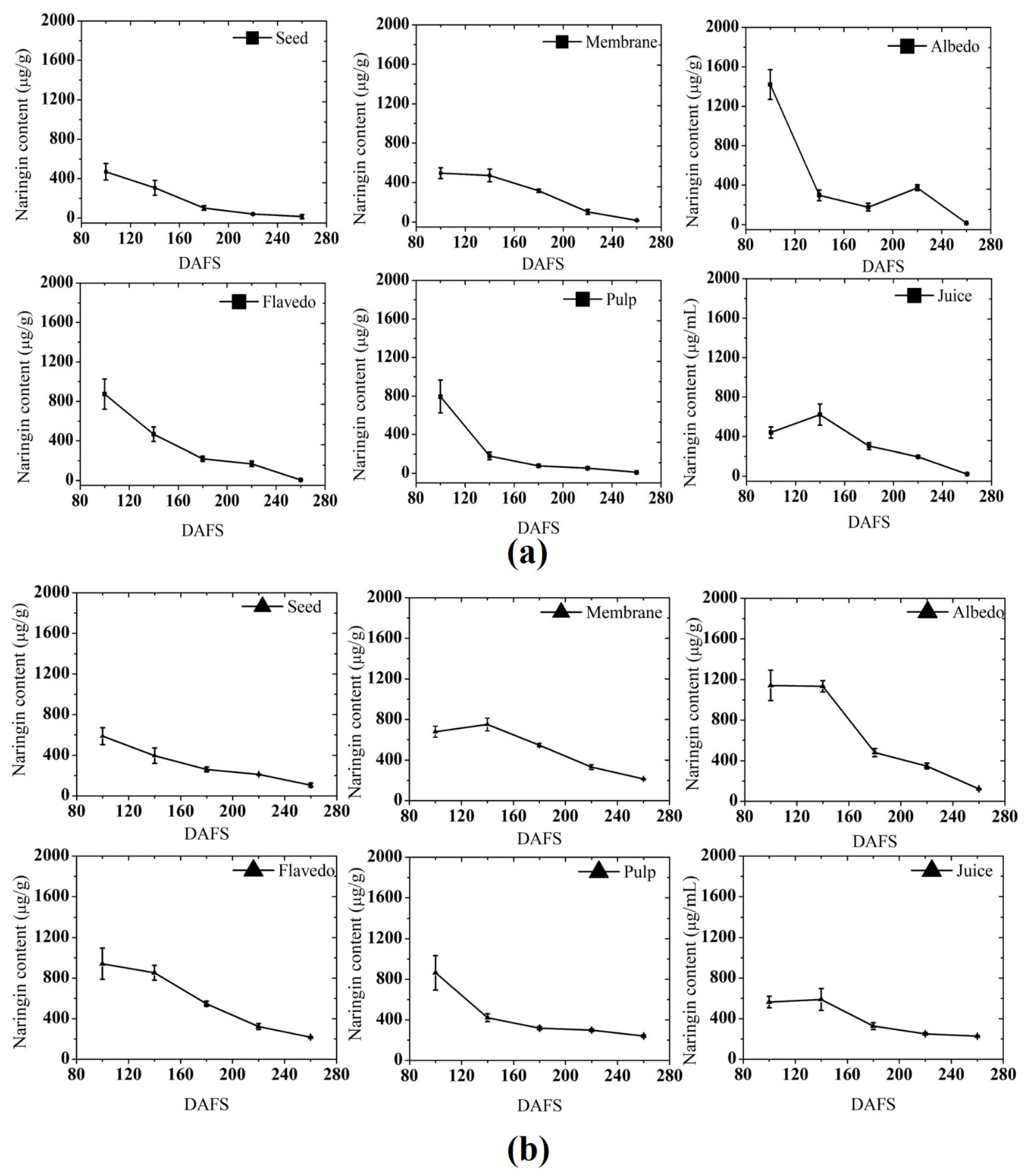
3.1.3. Naringin Content
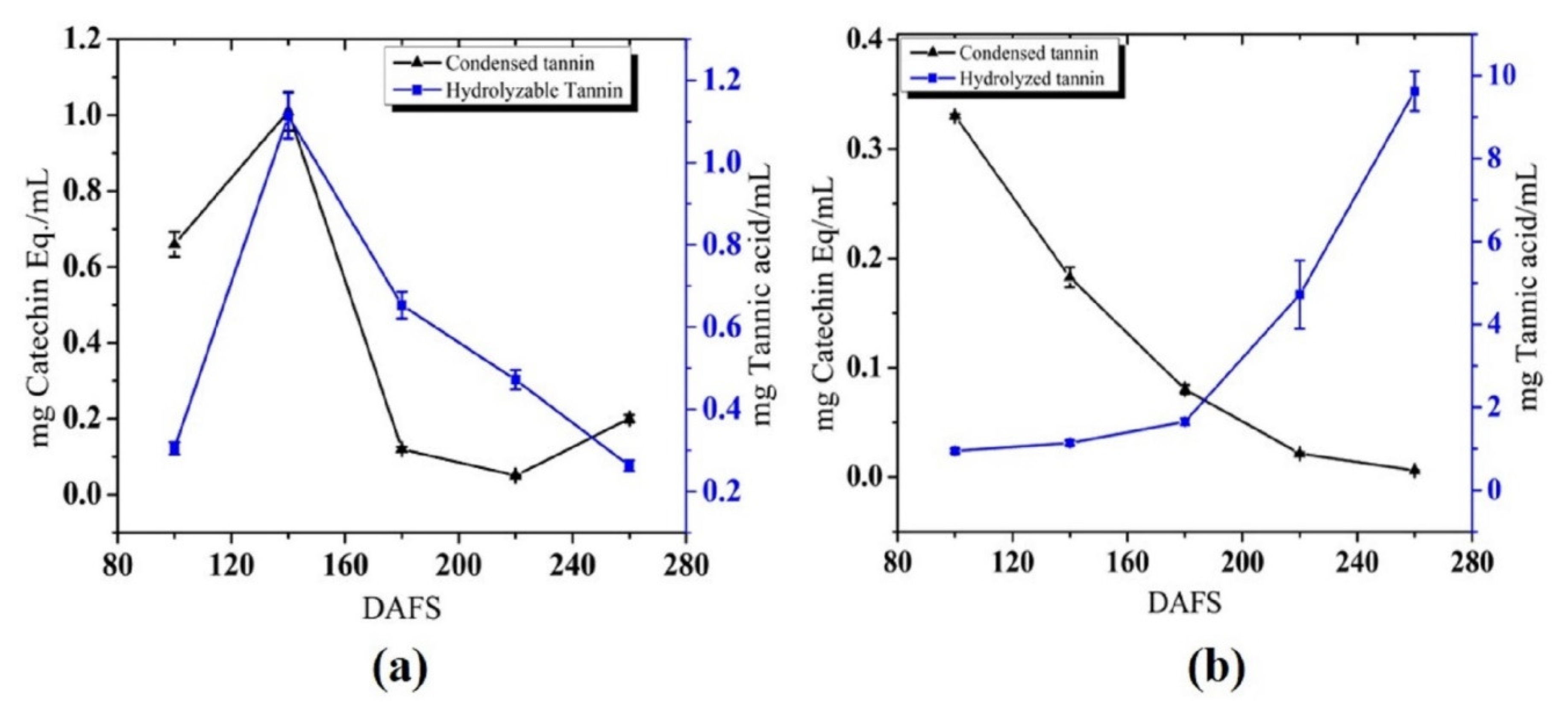
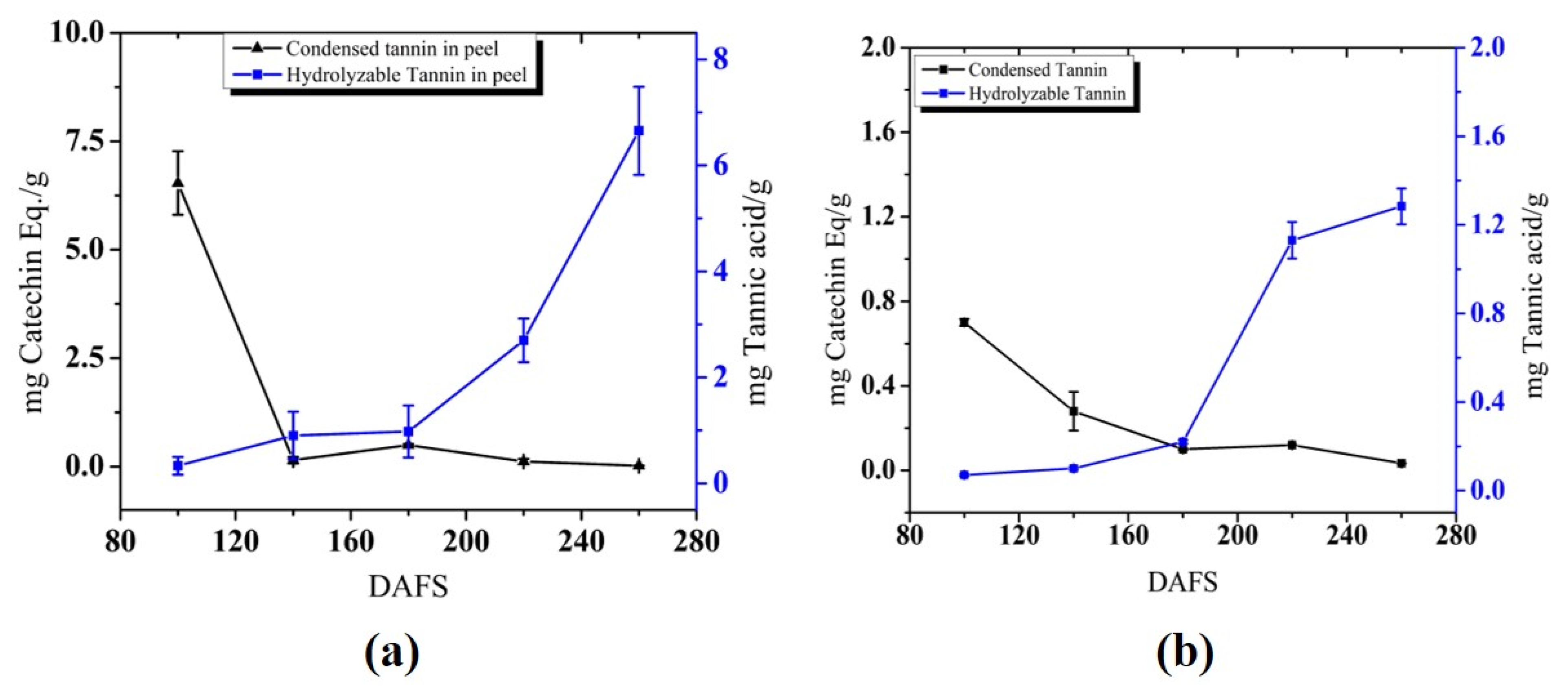
3.1.4. Tannin Content
3.2. Ferric Reducing Antioxidant Power, DPPH Free Radical Scavenging Activity, and Antioxidant Capacity
3.2.1. Antioxidant Capacity by the Phosphomolybdenum Method
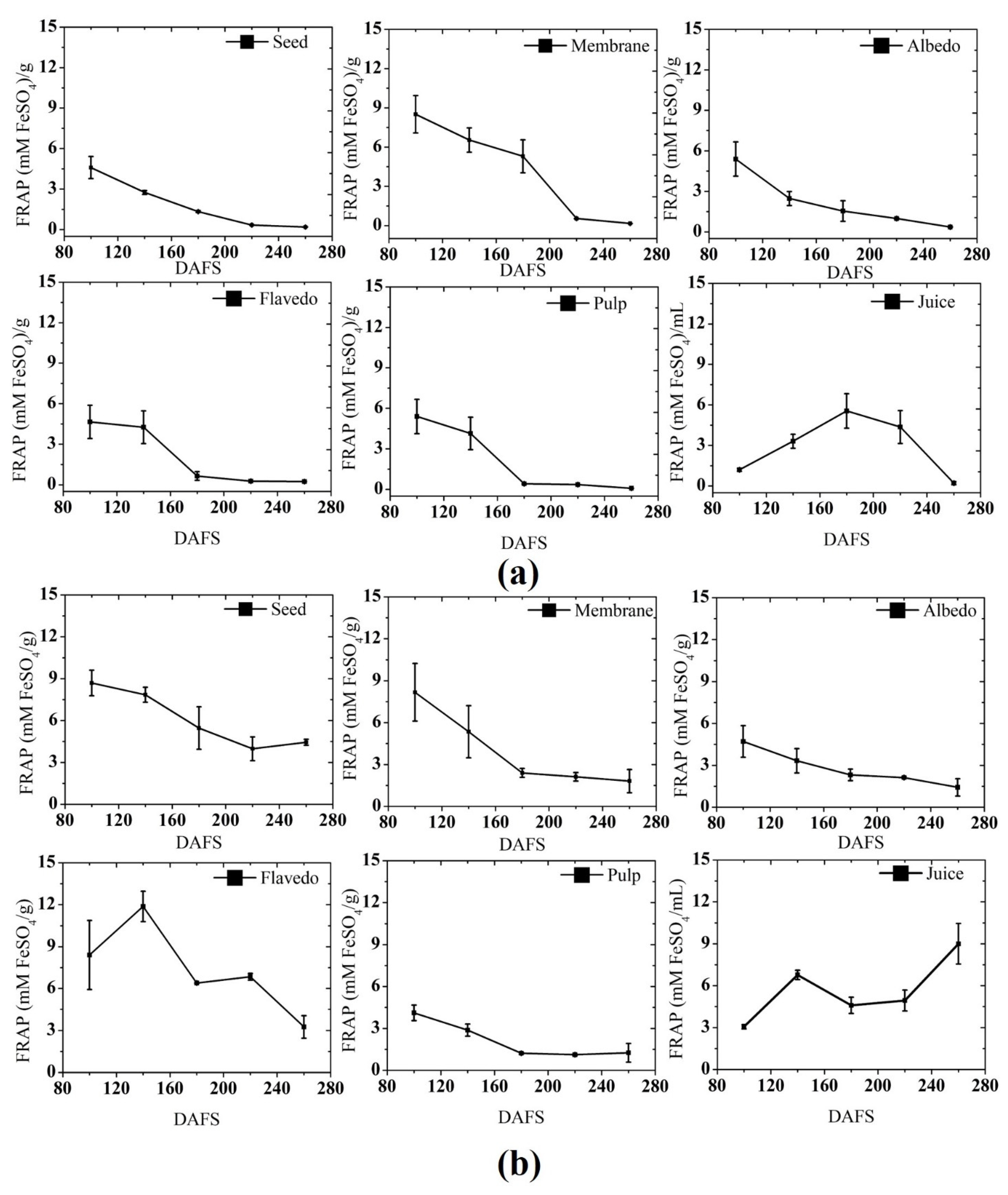
3.2.2. Ferric Reducing Antioxidant Power (FRAP)
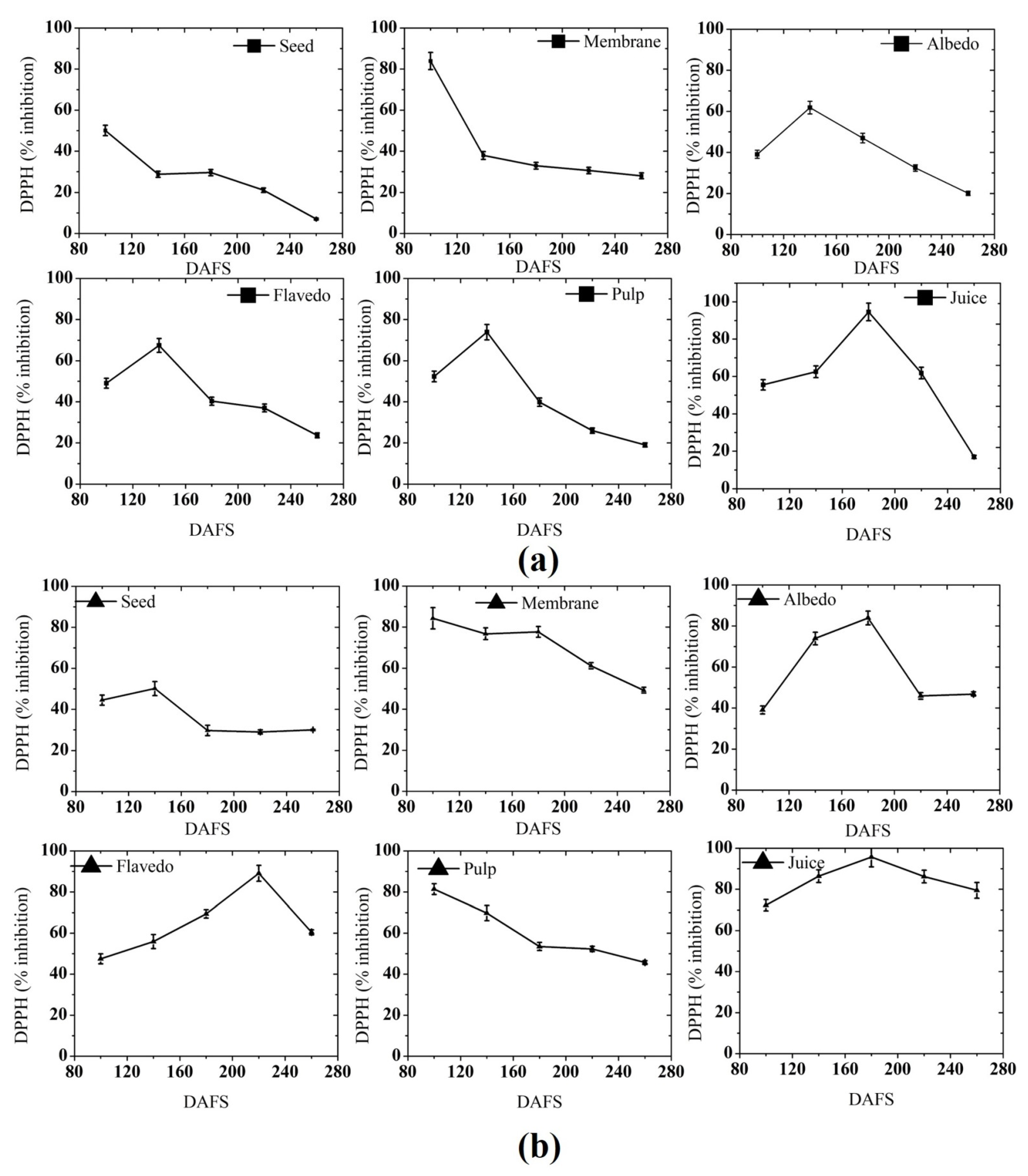
3.2.3. DPPH Free Radical Scavenging Activity
3.3. Variation in Volatile Composition of Pomelo Juice
3.4. Correlation of DAFS with Bioactive Compounds and Antioxidant Capacity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abate, G.; Zhang, L.; Pucci, M.; Morbini, G.; Mac Sweeney, E.; Maccarinelli, G.; Ribaudo, G.; Gianoncelli, A.; Uberti, D.; Memo, M.; et al. Phytochemical Analysis and Anti-Inflammatory Activity of Different Ethanolic Phyto-Extracts of Artemisia annua L. Biomolecules 2021, 11, 975. [Google Scholar] [CrossRef] [PubMed]
- Mastinu, A.; Bonini, S.A.; Premoli, M.; Maccarinelli, G.; Mac Sweeney, E.; Zhang, L.; Lucini, L.; Memo, M. Protective effects of Gynostemma pentaphyllum (var. Ginpent) against lipopolysaccharide-induced inflammation and motor alteration in mice. Molecules 2021, 26, 570. [Google Scholar] [CrossRef]
- Gupta, A.K.; Rather, M.A.; Kumar Jha, A.; Shashank, A.; Singhal, S.; Sharma, M.; Pathak, U.; Sharma, D.; Mastinu, A. Artocarpus lakoocha Roxb. and Artocarpus heterophyllus Lam. Flowers: New Sources of Bioactive Compounds. Plants 2020, 9, 1329. [Google Scholar] [CrossRef]
- Abate, G.; Vezzoli, M.; Sandri, M.; Rungratanawanich, W.; Memo, M.; Uberti, D. Mitochondria and cellular redox state on the route from ageing to Alzheimer’s disease. Mech. Ageing Dev. 2020, 192, 111385. [Google Scholar] [CrossRef] [PubMed]
- Rautiainen, S.; Levitan, E.B.; Orsini, N.; Åkesson, A.; Morgenstern, R.; Mittleman, M.A.; Wolk, A. Total antioxidant capacity from diet and risk of myocardial infarction: A prospective cohort of women. Am. J. Med. 2012, 125, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Zainudin, M.A.M.; Hamid, A.A.; Anwar, F.; Osman, A.; Saari, N. Variation of bioactive compounds and antioxidant activity of carambola (Averrhoa carambola L.) fruit at different ripening stages. Sci. Hortic. 2014, 172, 325–331. [Google Scholar] [CrossRef]
- Morton, J.F. Fruits of Warm Climates; Florida Flair Books; J.F. Morton Publisher: Miami, FL, USA, 1987; ISBN 0961018410. [Google Scholar]
- Sawamura, M.; Kuwahara, S.; Shichiri, K.; Aoki, T. Volatile Constituents of Several Varieties of Pummelos and a Comparison of the Nootkatone Levels in Pummelos and Other Citrus Fruits. Agric. Biol. Chem. 1990, 54, 803–805. [Google Scholar] [CrossRef]
- Tocmo, R.; Pena-Fronteras, J.; Calumba, K.F.; Mendoza, M.; Johnson, J.J. Valorization of pomelo (Citrus grandis Osbeck) peel: A review of current utilization, phytochemistry, bioactivities, and mechanisms of action. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1969–2012. [Google Scholar] [CrossRef]
- Shaw, P.E.; Goodner, K.L.; Moshonas, M.G.; Hearn, C.J. Comparison of grapefruit hybrid fruit with parent fruit based on composition of volatile components. Sci. Hortic. 2001, 91, 71–80. [Google Scholar] [CrossRef]
- González-Mas, M.C.; Rambla, J.L.; Alamar, M.C.; Gutiérrez, A.; Granell, A. Comparative Analysis of the Volatile Fraction of Fruit Juice from Different Citrus Species. PLoS ONE 2011, 6, e22016. [Google Scholar] [CrossRef] [Green Version]
- Cheong, M.-W.; Loke, X.-Q.; Liu, S.-Q.; Pramudya, K.; Curran, P.; Yu, B. Characterization of Volatile Compounds and Aroma Profiles of Malaysian Pomelo (Citrus grandis (L.) Osbeck) Blossom and Peel. J. Essent. Oil Res. 2011, 23, 34–44. [Google Scholar] [CrossRef]
- Cheong, M.W.; Liu, S.Q.; Zhou, W.; Curran, P.; Yu, B. Chemical composition and sensory profile of pomelo (Citrus grandis (L.) Osbeck) juice. Food Chem. 2012, 135, 2505–2513. [Google Scholar] [CrossRef] [PubMed]
- Kefford, J.F.; Chandler, B.V. The Chemical Constituents of Citrus Fruits; Advances in Food & Nutrition Research; Academic Press: Cambridge, MA, USA, 1970; ISBN 9780120164622. [Google Scholar]
- Holden, J.M.; Bhagwat, S.A.; Haytowitz, D.B.; Gebhardt, S.E.; Dwyer, J.T.; Peterson, J.; Beecher, G.R.; Eldridge, A.L.; Balentine, D. Development of a database of critically evaluated flavonoids data: Application of USDA’s data quality evaluation system. J. Food Compos. Anal. 2005, 18, 829–844. [Google Scholar] [CrossRef]
- Kim, K.; Park, K.-I. A Review of Antiplatelet Activity of Traditional Medicinal Herbs on Integrative Medicine Studies. Evid. Based. Complement. Alternat. Med. 2019, 2019, 7125162. [Google Scholar] [CrossRef]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Ortuño, A.; Díaz, L.; Alvarez, N.; Porras, I.; García-Lidón, A.; Del Río, J.A. Comparative study of flavonoid and scoparone accumulation in different Citrus species and their susceptibility to Penicillium digitatum. Food Chem. 2011, 125, 232–239. [Google Scholar] [CrossRef]
- Reza Yousefi, A.; Rashidi, S.; Moradi, P.; Mastinu, A. Germination and Seedling Growth Responses of Zygophyllum fabago, Salsola kali L. and Atriplex canescens to PEG-Induced Drought Stress. Environments 2020, 7, 107. [Google Scholar] [CrossRef]
- Naservafaei, S.; Sohrabi, Y.; Moradi, P.; Mac Sweeney, E.; Mastinu, A. Biological Response of Lallemantia iberica to Brassinolide Treatment under Different Watering Conditions. Plants 2021, 10, 496. [Google Scholar] [CrossRef]
- Aghajanlou, F.; Mirdavoudi, H.; Shojaee, M.; Mac Sweeney, E.; Mastinu, A.; Moradi, P. Rangeland Management and Ecological Adaptation Analysis Model for Astragalus curvirostris Boiss. Horticulturae 2021, 7, 67. [Google Scholar] [CrossRef]
- Zangani, E.; Afsahi, K.; Shekari, F.; Mac Sweeney, E.; Mastinu, A. Nitrogen and Phosphorus Addition to Soil Improves Seed Yield, Foliar Stomatal Conductance, and the Photosynthetic Response of Rapeseed (Brassica napus L.). Agriculture 2021, 11, 483. [Google Scholar] [CrossRef]
- Moradi, P.; Aghajanloo, F.; Moosavi, A.; Monfared, H.H.; Khalafi, J.; Taghiloo, M.; Khoshzaman, T.; Shojaee, M.; Mastinu, A. Anthropic Effects on the Biodiversity of the Habitats of Ferula gummosa. Sustainability 2021, 13, 7874. [Google Scholar] [CrossRef]
- Khaleghnezhad, V.; Yousefi, A.R.; Tavakoli, A.; Farajmand, B.; Mastinu, A. Concentrations-dependent effect of exogenous abscisic acid on photosynthesis, growth and phenolic content of Dracocephalum moldavica L. under drought stress. Planta 2021, 253, 127. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, A.; Moradi, P.; Mastinu, A. Variation in Terpene Profiles of Thymus vulgaris in Water Deficit Stress Response. Molecules 2020, 25, 1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, S.; Yu, O. Metabolic engineering of flavonoids in plants and microorganisms. Appl. Microbiol. Biotechnol. 2011, 91, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Schweiggert, R.M.; Steingass, C.B.; Mora, E.; Esquivel, P.; Carle, R. Carotenogenesis and physico-chemical characteristics during maturation of red fleshed papaya fruit (Carica papaya L.). Food Res. Int. 2011, 44, 1373–1380. [Google Scholar] [CrossRef]
- Castrejón, A.D.R.; Eichholz, I.; Rohn, S.; Kroh, L.W.; Huyskens-Keil, S. Phenolic profile and antioxidant activity of highbush blueberry (Vaccinium corymbosum L.) during fruit maturation and ripening. Food Chem. 2008, 109, 564–572. [Google Scholar] [CrossRef]
- Huang, R.; Xia, R.; Hu, L.; Lu, Y.; Wang, M. Antioxidant activity and oxygen-scavenging system in orange pulp during fruit ripening and maturation. Sci. Hortic. 2007, 113, 166–172. [Google Scholar] [CrossRef]
- Ilahy, R.; Hdider, C.; Lenucci, M.S.; Tlili, I.; Dalessandro, G. Antioxidant activity and bioactive compound changes during fruit ripening of high-lycopene tomato cultivars. J. Food Compos. Anal. 2011, 24, 588–595. [Google Scholar] [CrossRef]
- Wu, C.-S.; Gao, Q.-H.; Guo, X.; Yu, J.-G.; Wang, M. Effect of ripening stage on physicochemical properties and antioxidant profiles of a promising table fruit_pear-jujube_(Zizyphus jujuba Mill.). Sci. Hortic. 2012, 148, 177–184. [Google Scholar] [CrossRef]
- Karydas, C.; Iatrou, M.; Kouretas, D.; Patouna, A.; Iatrou, G.; Lazos, N.; Gewehr, S.; Tseni, X.; Tekos, F.; Zartaloudis, Z.; et al. Prediction of Antioxidant Activity of Cherry Fruits from UAS Multispectral Imagery Using Machine Learning. Antioxidants 2020, 9, 156. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.; Kalita, D. Assessment of pre-treatment of edible coatings prepared from gelatine and polyphenols (extracted under optimized conditions from amla fruit powder) on stability of banana chips. Acta Aliment. 2017, 46, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Broadhurst, R.; Jones, W. Analysis of condensed tannins using acidified vanillin. J. Sci. Food Agric. 1978, 29, 788–794. [Google Scholar] [CrossRef]
- Bossu, C.M.; Ferreira, E.C.; Chaves, F.S.; Menezes, E.A.; Nogueira, A.R.A. Flow injection system for hydrolysable tannin determination. Microchem. J. 2006, 84, 88–92. [Google Scholar] [CrossRef]
- Bao, Y.; Yuan, F.; Zhao, X.; Liu, Q.; Gao, Y. Equilibrium and kinetic studies on the adsorption debittering process of ponkan (Citrus reticulata Blanco) juice using macroporous resins. Food Bioprod. Process. 2015, 94, 199–207. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Jena, B.S.; Negi, P.S.; Sakariah, K.K. Evaluation of antioxidant activities and antimutagenicity of turmeric oil: A byproduct from curcumin production. Z. Naturforsch. C. 2002, 57, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Marziano, M.; Tonello, S.; Cantù, E.; Abate, G.; Vezzoli, M.; Rungratanawanich, W.; Serpelloni, M.; Lopomo, N.F.; Memo, M.; Sardini, E.; et al. Monitoring Caco-2 to enterocyte-like cells differentiation by means of electric impedance analysis on printed sensors. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Phenolics: From Chemistry to Biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- Shui, G.; Leong, L.P. Analysis of polyphenolic antioxidants in star fruit using liquid chromatography and mass spectrometry. J. Chromatogr. A 2004, 1022, 67–75. [Google Scholar] [CrossRef]
- Ortega-García, F.; Peragón, J. Phenylalanine ammonia-lyase, polyphenol oxidase, and phenol concentration in fruits of Olea europaea L. cv. Picual, Verdial, Arbequina, and Frantoio during ripening. J. Agric. Food Chem. 2009, 57, 10331–10340. [Google Scholar] [CrossRef]
- Ayaz, F.A.; Demir, O.; Torun, H.; Kolcuoglu, Y.; Colak, A. Characterization of polyphenoloxidase (PPO) and total phenolic contents in medlar (Mespilus germanica L.) fruit during ripening and over ripening. Food Chem. 2008, 106, 291–298. [Google Scholar] [CrossRef]
- Albertini, M.-V.; Carcouet, E.; Pailly, O.; Gambotti, C.; Luro, F.; Berti, L. Changes in organic acids and sugars during early stages of development of acidic and acidless citrus fruit. J. Agric. Food Chem. 2006, 54, 8335–8339. [Google Scholar] [CrossRef] [PubMed]
- Mansour, R. Determination of nutritional composition in citrus fruits (C. aurantium) during maturity. Nutr. Food Sci. 2019, 49, 299–317. [Google Scholar] [CrossRef]
- Samaniego, I.; Brito, B.; Viera, W.; Cabrera, A.; Llerena, W.; Kannangara, T.; Vilcacundo, R.; Angós, I.; Carrillo, W. Influence of the Maturity Stage on the Phytochemical Composition and the Antioxidant Activity of Four Andean Blackberry Cultivars (Rubus glaucus Benth) from Ecuador. Plants 2020, 9, 1027. [Google Scholar] [CrossRef] [PubMed]
- Ersus, S.; Cam, M. Determination of organic acids, total phenolic content, and antioxidant capacity of sour Citrus aurantium fruits. Chem. Nat. Compd. 2007, 43, 607–609. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, C.; Bucheli, P.; Wei, D. Citrus Flavonoids in Fruit and Traditional Chinese Medicinal Food Ingredients in China. Plant Foods Hum. Nutr. 2006, 61, 55–63. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, C.A.; Mansell, R.L. Three-Dimensional Distribution of Limonin, Limonoate A-Ring Monolactone, and Naringin in the Fruit Tissues of Three Varieties of Citrus paradisi. J. Agric. Food Chem. 1997, 45, 2876–2883. [Google Scholar] [CrossRef]
- Hasegawa, S.; Ou, P.; Fong, C.H.; Herman, Z.; Coggins, C.W., Jr.; Atkin, D.R. Changes in the limonoate A-ring lactone and limonin 17-. beta.-D-glucopyranoside content of navel oranges during fruit growth and maturation. J. Agric. Food Chem. 1991, 39, 262–265. [Google Scholar] [CrossRef]
- Sun, C.; Chen, K.; Chen, Y.; Chen, Q. Contents and antioxidant capacity of limonin and nomilin in different tissues of citrus fruit of four cultivars during fruit growth and maturation. Food Chem. 2005, 93, 599–605. [Google Scholar] [CrossRef]
- Li, S.; Wang, Z.; Ding, F.; Sun, D.; Ma, Z.; Cheng, Y.; Xu, J. Content changes of bitter compounds in “Guoqing No.1” Satsuma mandarin (Citrus unshiu Marc.) during fruit development of consecutive 3 seasons. Food Chem. 2014, 145, 963–969. [Google Scholar] [CrossRef]
- Hagen, R.E.; Dunlap, W.J.; Wender, S.H. Seasonal Variation of Naringin and Certain Other Flavanone Glycosides in Juice Sacs of Texas Ruby Red Grapefruit. J. Food Sci. 1966, 31, 542–547. [Google Scholar] [CrossRef]
- Castillo, J.; Benavente, O.; Del Río, J.A. Naringin and Neohesperidin Levels during Development of Leaves, Flower Buds, and Fruits of Citrus aurantium. Plant Physiol. 1992, 99, 67–73. [Google Scholar] [CrossRef]
- Ezeabara, C.A.; Okeke, C.U.; Ilodibia, C.V.; Aziagba, O.B. Determination of Tannin Content in Various Parts of Six Citrus Species. J. Sci. Res. Rep. 2014, 3, 1384–1392. [Google Scholar] [CrossRef]
- Diba, D.; Bultosa, G.; Tolesa, G.N. Effect of maturity stages on nutritive quality and sensory properties of Fig fruits. Botsw. J. Agric. Appl. Sci. 2019, 13, 3–10. [Google Scholar]
- Kyraleou, M.; Kallithraka, S.; Theodorou, N.; Teissedre, P.L.; Kotseridis, Y.; Koundouras, S. Changes in tannin composition of syrah grape skins and seeds during fruit ripening under contrasting water conditions. Molecules 2017, 22, 1453. [Google Scholar] [CrossRef] [Green Version]
- de Brandão, T.S.O.; de Sena, A.R.; Teshima, E.; David, J.M.; Assis, S.A. Changes in enzymes, phenolic compounds, tannins, and vitamin C in various stages of Jambolan (Syzygium cumini Lamark) development. Cienc. Tecnol. Aliment. 2011, 31, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Abate, G.; Vezzoli, M.; Polito, L.; Guaita, A.; Albani, D.; Marizzoni, M.; Garrafa, E.; Marengoni, A.; Forloni, G.; Frisoni, G.B. A Conformation Variant of p53 Combined with Machine Learning Identifies Alzheimer Disease in Preclinical and Prodromal Stages. J. Pers. Med. 2021, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Habibi, F.; Ramezanian, A.; Guillén, F.; Castillo, S.; Serrano, M.; Valero, D. Changes in Bioactive Compounds, Antioxidant Activity, and Nutritional Quality of Blood Orange Cultivars at Different Storage Temperatures. Antioxidants 2020, 9, 1016. [Google Scholar] [CrossRef] [PubMed]
- Menichini, F.; Tundis, R.; Bonesi, M.; Loizzo, M.R.; Conforti, F.; Statti, G.; De Cindio, B.; Houghton, P.J.; Menichini, F. The influence of fruit ripening on the phytochemical content and biological activity of Capsicum chinense Jacq. cv Habanero. Food Chem. 2009, 114, 553–560. [Google Scholar] [CrossRef]
- Kalt, W.; Lawand, C.; Ryan, D.A.J.; McDonald, J.E.; Donner, H.; Forney, C.F. Oxygen Radical Absorbing Capacity, Anthocyanin and Phenolic Content of Highbush Blueberries (Vaccinium corymbosum L.) during Ripening and Storage. J. Am. Soc. Hortic. Sci. JASHS 2003, 128, 917–923. [Google Scholar] [CrossRef]
- Namiesnik, J.; Vearasilp, K.; Kupska, M.; Ham, K.-S.; Kang, S.-G.; Park, Y.-K.; Barasch, D.; Nemirovski, A.; Gorinstein, S. Antioxidant activities and bioactive components in some berries. Eur. Food Res. Technol. 2013, 237, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi, K.; Ghasemi, Y.; Ebrahimzadeh, M.A. Antioxidant activity, phenol and flavonoid contents of 13 citrus species peels and tissues. Pak. J. Pharm. Sci. 2009, 22, 277–281. [Google Scholar] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Muda, R.J.; Nurhanan, A.R.; Rosli, W.; Mohsin, S. Total polyphenol content and free radical scavenging activity of cornsilk (Zea mays hairs). Sains Malays 2012, 41, 1217–1221. [Google Scholar]
- Perez-Cacho, P.R.; Rouseff, R. Processing and storage effects on orange juice aroma: A review. J. Agric. Food Chem. 2008, 56, 9785–9796. [Google Scholar] [CrossRef]
- Furusawa, M.; Hashimoto, T.; Noma, Y.; Asakawa, Y. Highly efficient production of nootkatone, the grapefruit aroma from valencene, by biotransformation. Chem. Pharm. Bull. 2005, 53, 1513–1514. [Google Scholar] [CrossRef] [Green Version]
- Ledesma-Escobar, C.A.; Priego-Capote, F.; Robles Olvera, V.J.; Luque de Castro, M.D. Targeted analysis of the concentration changes of phenolic compounds in Persian lime (Citrus latifolia) during fruit growth. J. Agric. Food Chem. 2018, 66, 1813–1820. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Overall Compounds | Identified in 2018 | Identified in 2019 |
|---|---|---|---|
| Hydrocarbons | |||
| 1. | α-pinene | + | + |
| 2. | β-pinene | + | + |
| 3. | Sabinene | + | + |
| 4. | β-Myrcene | + | + |
| 5. | α-terpinene | + | + |
| 6. | Camphene | + | + |
| 7. | Cyclohexane | + | + |
| 8. | m-cymene | + | + |
| 9. | Benzene | + | + |
| 10. | Dodecane | + | + |
| 11. | 9-tricosene | + | − |
| 12. | Caryophyllene | − | + |
| 13. | Alloaromadendrene | − | + |
| 14. | α-Copaene | + | + |
| 15. | β-Copaene | + | + |
| 16. | α,β-Cubebene | + | + |
| 17. | Limonene | + | + |
| 18. | 17-Acetyloxy | + | + |
| 19. | β-Phellandrene | + | + |
| 20. | trans-β-Ocimene | − | + |
| 21. | cis-β-Ocimene | − | + |
| 22. | q-Cymene | + | + |
| 23. | Terpinolene | + | + |
| 24. | 4,8-Dimethyl-1,3,7-nonatriene | + | − |
| 25. | d-Elemene | + | + |
| 26. | γ-Elemene | + | + |
| 27. | β-Caryophyllene | − | + |
| 28. | c-Elemene | + | + |
| 29. | α-Humulene | + | + |
| 30. | β-Humulene | + | + |
| 31. | Germacrene D | + | − |
| 32. | α-Selinene | + | + |
| 33. | 1,1-Biphenyl | + | + |
| 34. | z,z,z-4,6,9-Nonadecatriene | + | − |
| 35. | Coumarin | + | + |
| 36. | Longifolene | − | + |
| 37. | Cyclohexene | + | + |
| 38. | 4-Carene | − | + |
| Esters | |||
| 39. | Neryl acetate | + | + |
| 40. | Ethyl palmitate | + | + |
| 41. | Methyl esters | + | + |
| Acids | |||
| 42. | Acetic acid | + | + |
| 43. | Malonic acid | + | + |
| 44. | 2-Pentenoic acid | + | + |
| 45. | Propionic acid | + | + |
| 46. | Hexadecanoic acid | + | + |
| 47. | Nonanoic acid | + | + |
| 48. | Butyric acid | + | + |
| 49. | Benzoic acid | + | + |
| 50. | Oleic acid | + | + |
| 51. | Linoleic acid | + | + |
| 52. | α-Linoleic acid | + | + |
| 53. | Oxalic acid | + | + |
| 54. | 1,2,3,4-Tetrahydroisoquinolin-6-ol-1-carboxylic acid | + | − |
| 55. | Mercaptobenzoic acid | + | − |
| 56. | 4-Oxatricyclo[6.3.0.0]-undecan-5-one-1-carboxylic acid | + | + |
| 57. | Cis-vaccenic acid | − | + |
| Alcohols | |||
| 58. | Phenol | + | + |
| 59. | Ethanol | + | + |
| 60. | Ergoline 8-Methanol | − | + |
| 61. | 1-dodecanol | + | + |
| 62. | Linalool | + | + |
| 63. | Nerol | + | + |
| 64. | Geraniol | + | + |
| 65. | Carveol | + | + |
| 66. | 1-Heptatriacotanol | + | + |
| 67. | Eugenol | + | + |
| 68. | 1-Nonadecanol | + | − |
| 69. | 1-Undecanol | + | − |
| 70. | Corynan-17-ol | + | + |
| Aldehydes | |||
| 71. | Octanal | + | + |
| 72. | 7-Heptadecene | + | + |
| 73. | Decanal | + | + |
| 74. | Benzaldehyde | + | + |
| 75. | Citronellal | + | + |
| 76. | Neral | − | + |
| 77. | Geranial | + | + |
| 78. | trans,cis-2,4-Decadienal | + | + |
| 79. | Dodecanal | + | + |
| 80. | Perilla aldehyde | + | + |
| 81. | trans,trans-2,4-Decadienal | + | − |
| 82. | trans-2-Dodecenal | + | − |
| 83. | cis,trans-2,6-nonadienal | + | + |
| 84. | Octadecanal | + | + |
| 85. | 9-Octadecenoic acid | + | + |
| 86. | 2-Nonadecanone | + | + |
| 87. | D-carvone | + | + |
| 88. | 4H-1-Benzopyran-4-one | + | + |
| 89. | 2,4-Cycloheptadien | − | + |
| 90. | 2,6-dihydroxyacetophenone | + | + |
| 91. | 9,10-Anthracene-dione | + | + |
| 92. | 3H-cycloocta[c]pyran-3-one | + | + |
| 93. | Flavone | + | + |
| Others | |||
| 94. | Limonene oxide | + | + |
| 95. | 3,5-Androstadien-17-one-oxime | + | + |
| 96. | δ-Cadinene | + | + |
| 97. | Bis(trimethylsiyl) ether | + | + |
| 98. | Trienbolone | + | + |
| 99. | (-)-Isolongifolol | + | + |
| 100. | Silane | + | + |
| 101. | Di-Silane | + | + |
| 102. | Copaene | + | + |
| 103. | Bicyclo | + | + |
| 104. | 1H-indole | + | + |
| 105. | Chinchonan | + | + |
| 106. | Giseofulvin | + | + |
| 107. | Quinine | + | + |
| 108. | Indole | + | + |
| 109. | Nootkatone | + | + |
| 110. | Osthole | + | + |
| 111. | 3-Piperdinamine | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, A.K.; Dhua, S.; Sahu, P.P.; Abate, G.; Mishra, P.; Mastinu, A. Variation in Phytochemical, Antioxidant and Volatile Composition of Pomelo Fruit (Citrus grandis (L.) Osbeck) during Seasonal Growth and Development. Plants 2021, 10, 1941. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091941
Gupta AK, Dhua S, Sahu PP, Abate G, Mishra P, Mastinu A. Variation in Phytochemical, Antioxidant and Volatile Composition of Pomelo Fruit (Citrus grandis (L.) Osbeck) during Seasonal Growth and Development. Plants. 2021; 10(9):1941. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091941
Chicago/Turabian StyleGupta, Arun Kumar, Subhamoy Dhua, Partha Pratim Sahu, Giulia Abate, Poonam Mishra, and Andrea Mastinu. 2021. "Variation in Phytochemical, Antioxidant and Volatile Composition of Pomelo Fruit (Citrus grandis (L.) Osbeck) during Seasonal Growth and Development" Plants 10, no. 9: 1941. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091941