
Comparative Study of Metabolomic Profile and Antioxidant Content of Adult and In Vitro Leaves of Aristotelia chilensis

 , , , , ,
, , , , ,  , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
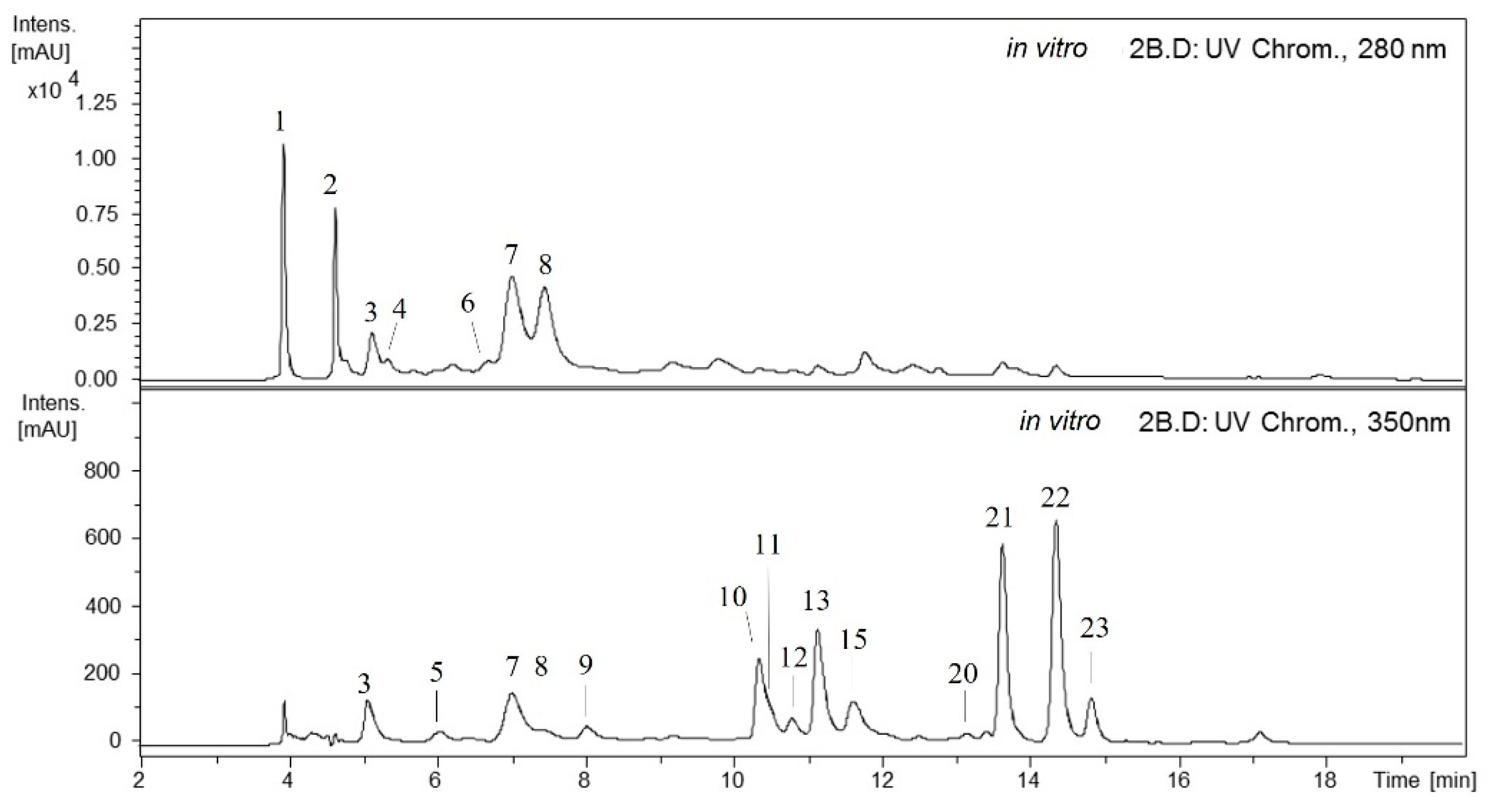
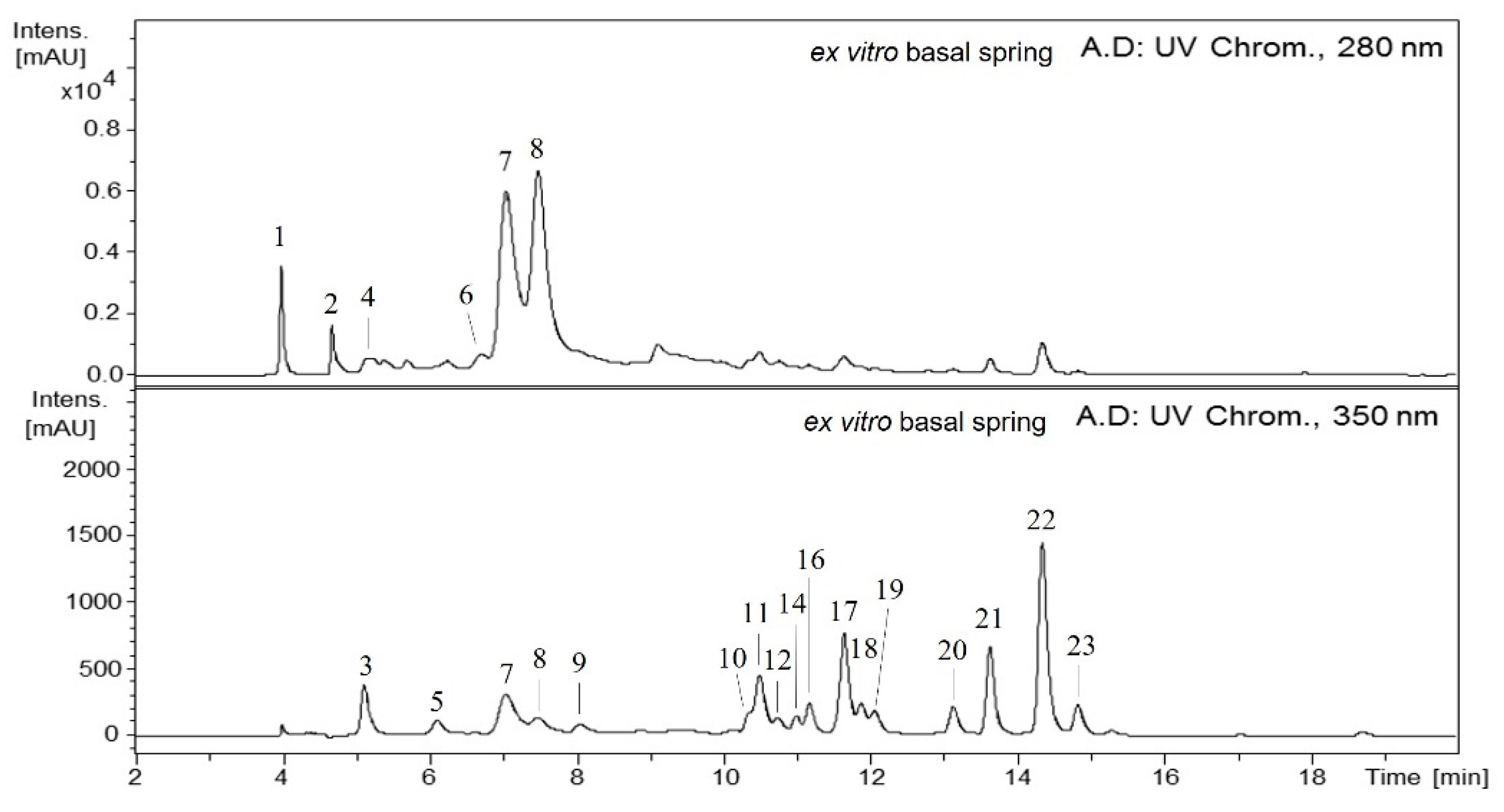
2.1. Qualitative Analysis of the Maqui Leaves
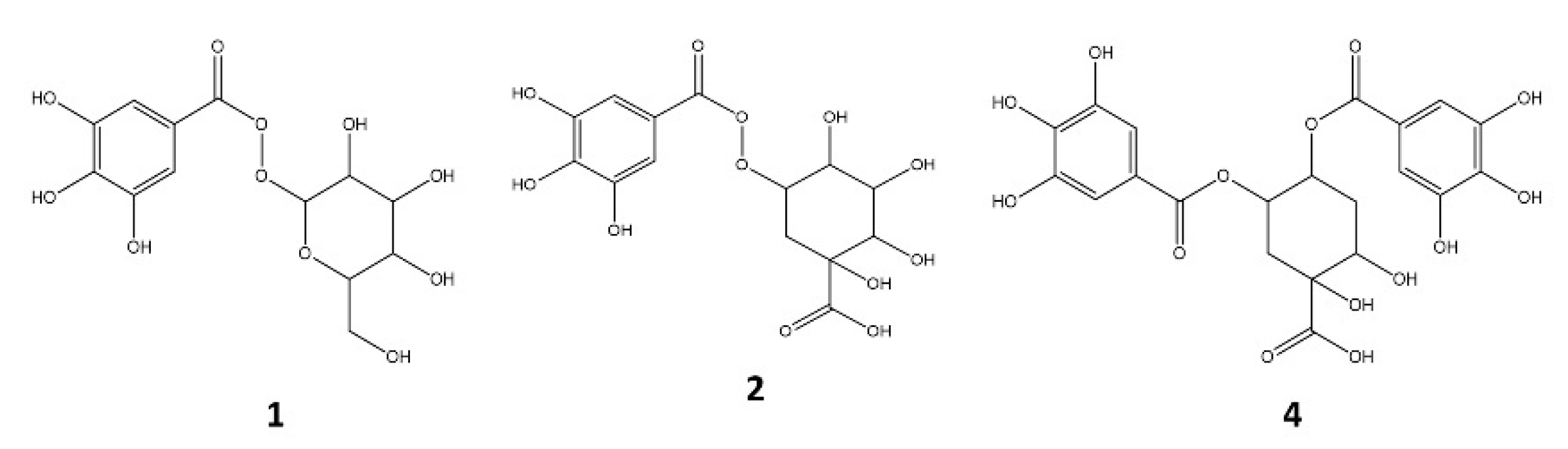
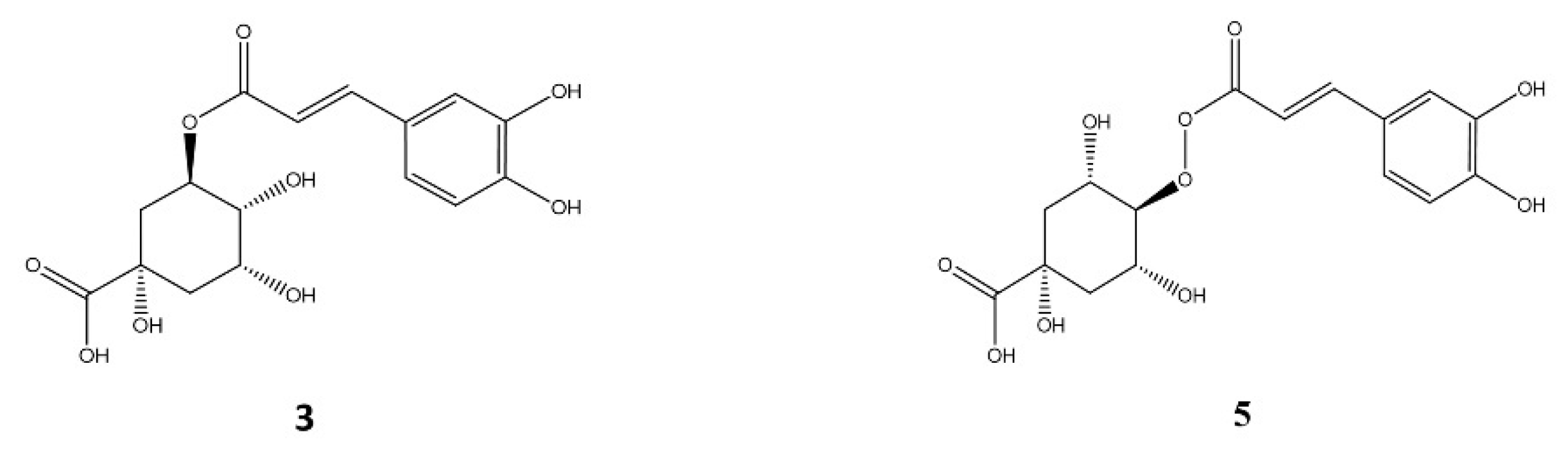
2.1.1. Galloyl Acid Derivatives
2.1.2. Caffeoyl Quinic Acids
2.1.3. Ellagitannins
2.1.4. Ellagic Acid Derivatives
2.1.5. Flavonoid Derivatives
2.2. Qualitative Differences of Phenolics from In Vitro and Ex Vitro Samples
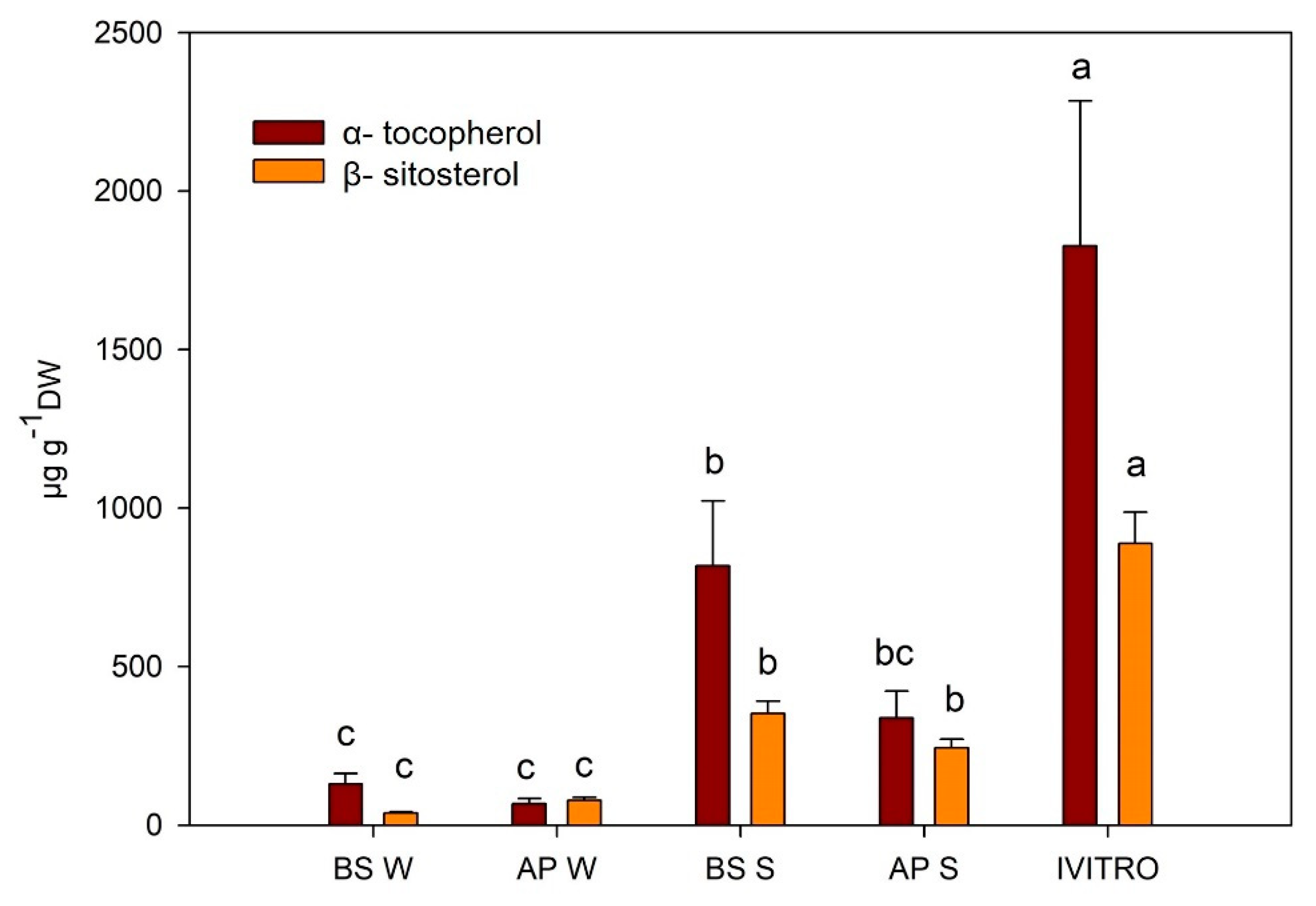
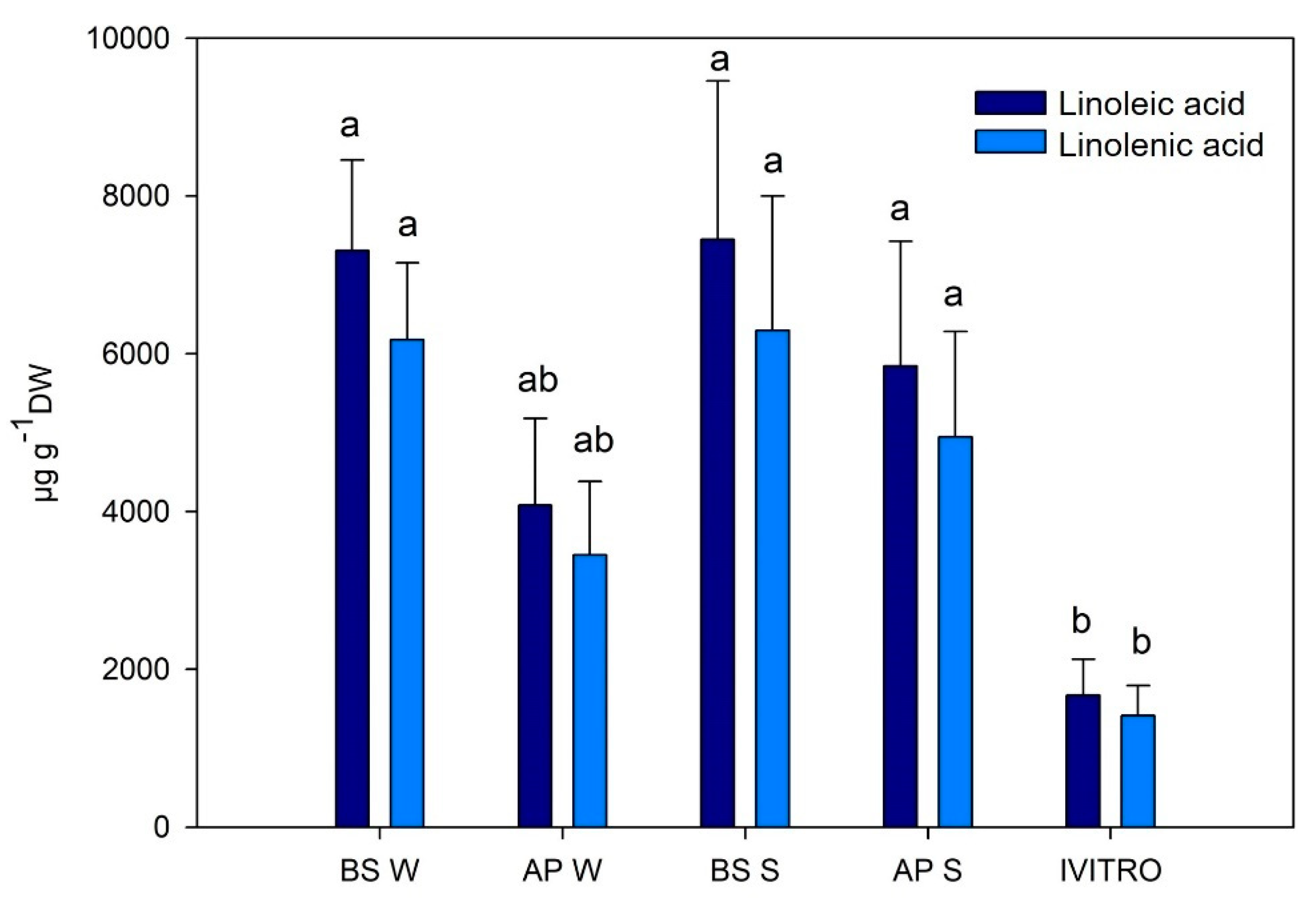
2.3. Quantitative Analysis of the Maqui Leaves
3. Materials and Methods
3.1. Plant Material
3.1.1. In Vitro Plant Cultures
3.1.2. Preparation of the Samples from In Vivo and In Vitro Maqui Leaves
3.2. Determination of Polyphenolic Compounds by HPLC-DAD-ESI (Ion Trap)-MSn
3.3. Determination of Polyphenolic Compounds by HPLC-MS
3.4. Determination of Lipophilic Compounds by GC/MS
3.5. Total Flavonoid Content
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zúñiga, G.E.; Tapia, A.; Arenas, A.; Contreras, R.A.; Zúñiga-Libano, G. Phytochemistry and biological properties of Aristotelia chilensis a Chilean blackberry: A review. Phytochem. Rev. 2017, 16, 1081–1094. [Google Scholar] [CrossRef]
- Girones-Vilaplana, A.; Huertas, J.P.; Moreno, D.A.; Periago, P.M.; Garcia-Viguera, C. Quality and microbial safety evaluation of new isotonic beverages upon thermal treatments. Food Chem. 2016, 194, 455–462. [Google Scholar] [CrossRef]
- Ojeda, J.; Jara, E.; Molina, L.; Parada, F.; Burgos, R.A.; Hidalgo, M.A.; Hancke, J.L. Effects of Aristotelia chilensis berry juice on cyclooxygenase 2 expression. Bol. Latinoam. Caribe Plantas Med. Aromáticas 2011, 10, 543–552. [Google Scholar]
- Mena, J.; Elgueta, E.; Espinola-Gonzales, F.; Cárdenas, H.; Orihuela, P.A. Hydroethanolic Extracts of the Aristotelia Chilensis (Maqui) Berry Reduces Cellular Viability and Invasiveness in the Endometrial Cancer Cell Line Ishikawa. Integr. Cancer Ther. 2021, 20, 15347354211007560. [Google Scholar] [CrossRef]
- Céspedes, C.L.; El-Hafidi, M.; Pavón, N.; Alarcón, J. Antioxidant and cardioprotective activities of phenolic extracts from fruits of Chilean blackberry Aristotelia chilensis (Elaeocarpaceae), Maqui. Food Chem. 2008, 107, 820–829. [Google Scholar] [CrossRef]
- Céspedes, C.L.; Pavón, N.; Domínguez, M.; Alarcón, J.; Balbontín, C.; Kubo, I.; El-Hafidi, M.; Ávila, J.G. The chilean superfruit superfruit black-berry Aristotelia chilensis (Elaeocarpaceae), Maqui as mediator in inflammation-associated disorders. Food Chem. Toxicol. 2017, 108, 438–450. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, T.; Arguelles-Arias, F.; Begines, B.; García-Montes, J.M.; Pereira, A.; Victoriano, M.; Vazquez-Roman, V.; Pérez Bernal, J.L.; Callejón, R.M.; De-Miguel, M.; et al. Native Chilean Berries Preservation and In Vitro Studies of a Polyphenol Highly Antioxidant Extract from Maqui as a Potential Agent against Inflammatory Diseases. Antioxidants 2021, 10, 843. [Google Scholar] [CrossRef]
- Rubilar, M.; Jara, C.; Poo, Y.; Acevedo, F.; Gutiérrez, C.; Sineiro, J.; Shene, C. Extracts of Maqui (Aristotelia chilensis) and Murta (Ugni molinae Turcz.): Sources of antioxidant compounds and alpha-Glucosidase/alpha-Amylase inhibitors. J. Agric. Food Chem. 2011, 59, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Rojo, L.E.; Ribnicky, D.; Logendra, S.; Poulev, A.; Rojas-Silva, P.; Kuhn, P.; Dorn, R.; Grace, M.H.; Lila, M.A.; Raskin, I. In Vitro and in Vivo Anti-Diabetic Effects of Anthocyanins from Maqui Berry (Aristotelia chilensis). Food Chem. 2012, 131, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, L.; Trostchansky, A.; Wood, I.; Mastrogiovanni, M.; Vogel, H.; González, B.; Marostica Junior, M.; Fuentes, E.; Palomo, I. Antiplatelet activity and chemical analysis of leaf and fruit extracts from Aristotelia chilensis. PLoS ONE 2021, 16, e0250852. [Google Scholar] [CrossRef]
- Masoodi, H.; Villano, D.; Zafrilla, P. A comprehensive review on fruit Aristotelia chilensis (Maqui) for modern health: Towards a better understanding. Food Funct. 2019, 10, 3057–3067. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Aspera-Werz, R.H.; Chen, T.; Weng, W.; Braun, B.; Histing, T.; Nussler, A.K. Maqui berry extract prevents cigarette smoke induced oxidative stress in human osteoblasts in vitro. EXCLI J. 2021, 20, 281–296. [Google Scholar] [PubMed]
- Concha-Meyer, A.A.; Sepúlveda, G.; Pérez-Díaz, R.; Torres, C.A. Effect of preservation processing on quality attributes and phenolic profile of maqui (Aristotelia chilensis mol. Stuntz) fruit. LWT 2021, 149, 111920. [Google Scholar] [CrossRef]
- Felgines, C.; Talavera, S.; Texier, O.; Gil-Izquierdo, A.; Lamaison, J.-L.; Rémésy, C. Blackberry Anthocyanins Are Mainly Recovered from Urine asMethylated and Glucuronidated Conjugates in Humans. J. Agric. Food Chem. 2005, 53, 7721–7727. [Google Scholar] [CrossRef] [PubMed]
- Talavera, S.; Felgines, C.; Texier, O.; Besson, C.; Gil-Izquierdo, A.; Lamaison, J.-L.; Rémésy, C. Anthocyanin Metabolism in Rats and Their Distribution to Digestive Area, Kidney, and Brain. J. Agric. Food Chem. 2005, 53, 3902–3908. [Google Scholar] [CrossRef]
- Fredes, C.; Yousef, G.G.; Robert, P.; Grace, M.H.; Lila, M.A.; Gomez, M.; Gebauer, M.; Montenegro, G. Anthocyanin profiling of wild maqui berries (Aristotelia chilensis [Mol.] Stuntz) from different geographical regions in Chile. J. Sci. Food Agric. 2014, 94, 2639–2648. [Google Scholar] [CrossRef]
- Turchetti, G.; Paz, C. Aristotelia chilensis (Mol.) Stuntz: A Natural Source of Bioactive Compounds. Curr. Tradit. Med. 2019, 5, 66–74. [Google Scholar] [CrossRef]
- Muñoz, O.; Christen, P.; Cretton, S.; Backhouse, N.; Torres, V.; Correa, O.; Costa, E.; Miranda, H.; Delporte, C. Chemical study and anti-inflammatory, analgesic and antioxidant activities of the leaves of Aristotelia chilensis (Mol.) Stuntz, Elaeocarpaceae. J. Pharm. Pharmacol. 2011, 63, 849–859. [Google Scholar] [CrossRef]
- Céspedes, C.L.; Balbontín, C.; Ávila, J.G.; Domínguez, M.; Alarcón, J.; Paz, C.; Burgos, V.; Ortiz, L.; Peñaloza-Castro, I.; Seigler, D.S.; et al. Inhibition on cholinesterase and tyrosinase by alkaloids and phenolics from Aristotelia chilensis leaves. Food Chem. Toxicol. 2017, 109, 984–995. [Google Scholar] [CrossRef]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Ramachandra Rao, S.; Ravishankar, G.A. Plant Cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [CrossRef]
- Isah, T.; Umar, S.; Mujib, A.; Sharma, M.P.; Rajasekharan, P.E.; Zafar, N.; Frukh, A. Secondary metabolism of pharmaceuticals in the plant in vitro cultures: Strategies, approaches, and limitations to achieving higher yield. Plant Cell Tissue Organ Cult. 2017, 132, 239–265. [Google Scholar] [CrossRef]
- Ferreres, F.; Gil-Izquierdo, A.; Vinholes, J.; Silva, S.T.; Valentao, P.; Andrade, P.B. Bauhinia asteuriz Link authenticity using flavonoids profile: Relation with their biological properties. Food Chem. 2012, 134, 894–904. [Google Scholar] [CrossRef]
- Motta, E.V.; Lemos, M.; Costa, J.C.; Bandero-Filho, V.C.; Sasse, A.; Sheridan, H.; Bastos, J.K. Galloylquinic acid derivatives from Copaifera langsdorffii leaves display gastroprotective activity. Chem. Biol. Interact. 2017, 261, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Maldini, M.; Montoro, P.; Pizza, C. Phenolic compounds from Byrsonima crassifolia L. bark: Phytochemical investigation and quantitative analysis by LC-ESI MS/MS. J. Pharm. Biomed. Anal. 2011, 56, 1–6. [Google Scholar] [CrossRef]
- Erşan, S.; Güçlü Üstündağ, Ö.; Carle, R.; Schweiggert, R.M. Determination of pistachio (Pistacia vera L.) hull (exo—and mesocarp) phenolics by HPLC-DAD-ESI/MSn and UHPLC-DAD-ELSD after ultrasound-assisted extraction. J. Food Compos. Anal. 2017, 62, 103–114. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Petreska, J.; Stefova, M.; Ferreres, F.; Moreno, D.A.; Tomás-Barberán, F.A.; Stefkov, G.; Kulevanova, S.; Gil-Izquierdo, A. Potential bioactive phenolics of Macedonian Sideritis species used for medicinal “Mountain Tea”. Food Chem. 2011, 125, 13–20. [Google Scholar] [CrossRef]
- Andrade, C.; Ferreres, F.; Gomes, N.G.M.; Duangsrisai, S.; Srisombat, N.; Vajrodaya, S.; Pereira, D.M.; Gil-Izquierdo, A.; Andrade, P.B.; Valentao, P. Phenolic Profiling and Biological Potential of Ficus curtipes Corner Leaves and Stem Bark: 5-Lipoxygenase Inhibition and Interference with NO Levels in LPS-Stimulated RAW 264.7 Macrophages. Biomolecules 2019, 9, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mena, P.; Vegara, S.; Marti, N.; Garcia-Viguera, C.; Saura, D.; Valero, M. Changes on indigenous microbiota, colour, bioactive compounds and antioxidant activity of asteurized pomegranate juice. Food Chem. 2013, 141, 2122–2129. [Google Scholar] [CrossRef]
- Mena, P.; Calani, L.; Dall’Asta, C.; Galaverna, G.; García-Viguera, C.; Bruni, R.; Crozier, A.; Del Rio, D. Rapid and comprehensive evaluation of (poly)phenolic compounds in pomegranate (Punica granatum L.) juice by UHPLC-MSn. Molecules 2012, 17, 14821–14840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreres, F.; Grosso, C.; Gil-Izquierdo, A.; Valentao, P.; Andrade, P.B. Ellagic acid and derivatives from Cochlospermum angolensis Welw. Extracts: HPLC-DAD-ESI/MS(n) profiling, quantification and in vitro anti-depressant, anti-cholinesterase and anti-oxidant activities. Phytochem. Anal. 2013, 24, 534–540. [Google Scholar] [CrossRef]
- Borges, G.; Crozier, A. HPLC-PDA-MS fingerprinting to assess the authenticity of pomegranate beverages. Food Chem. 2012, 135, 1863–1867. [Google Scholar] [CrossRef] [PubMed]
- Medina, S.; Collado-González, J.; Ferreres, F.; Londono-Londono, J.; Jiménez-Cartagena, C.; Guy, A.; Durand, T.; Galano, J.M.; Gil-Izquierdo, A. Quantification of phytoprostanes—bioactive oxylipins—and phenolic compounds of Passiflora edulis Sims shell using UHPLC-QqQ-MS/MS and LC-IT-DAD-MS/MS. Food Chem. 2017, 229, 1–8. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Gil-Izquierdo, A.; Medina, S.; Ferreres, F. Phenolic composition profiling of different edible parts and by-products of date palm (Phoenix dactylifera L.) by using HPLC-DAD-ESI/MS(n). Food Res. Int. 2017, 100, 494–500. [Google Scholar] [CrossRef]
- Gil-Izquierdo, A.; Mellenthin, A. Identification and quantitation of flavonols in rowanberry (Sorbus aucuparia L.) juice. Eur. Food Res. Technol. 2001, 213, 12–17. [Google Scholar]
- Ferreres, F.; Gomes, N.G.M.; Valentao, P.; Pereira, D.M.; Gil-Izquierdo, A.; Araujo, L.; Silva, T.C.; Andrade, P.B. Leaves and stem bark from Allophylus africanus P. Beauv.: An approach to anti-inflammatory properties and characterization of their flavonoid profile. Food Chem. Toxicol. 2018, 118, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Oliveira, A.P.; Gil-Izquierdo, A.; Valentao, P.; Andrade, P.B. Piper betle leaves: Profiling phenolic compounds by HPLC/DAD-ESI/MS(n) and anti-cholinesterase activity. Phytochem. Anal. 2014, 25, 453–460. [Google Scholar] [CrossRef]
- Nikolova, M.; Petrova, M.; Zayova, E. Comparative study of in vitro, ex vitro and in vivo grown plants of Arnica montana —polyphenols and free radical scavenging activity. Acta Bot. Croat. 2013, 72, 13–22. [Google Scholar] [CrossRef]
- Giri, L.; Dhyani, P.; Rawat, S.; Bhatt, I.D.; Nandi, S.K.; Rawal, R.S.; Pande, V. In vitro production of phenolic compounds and antioxidant activity in callus suspension cultures of Habenaria edgeworthii: A rare Himalayan medicinal orchid. Ind. Crop. Prod. 2012, 39, 1–6. [Google Scholar] [CrossRef]
- Arias, H.R.; Ortells, M.O.; Feuerbach, D.; Burgos, V.; Paz, C. Alkaloids Purified from Aristotelia chilensis Inhibit the Human alpha3beta4 Nicotinic Acetylcholine Receptor with Higher Potencies Compared with the Human alpha4beta2 and alpha7 Subtypes. J. Nat. Prod. 2019, 82, 1953–1960. [Google Scholar] [CrossRef]
- Vidal, J.L.; Avello, L.M.; Loyola, C.C.; Campos, P.J.; Aqueveque, M.P.; Dungan, S.R.; Galotto, L.M.; Guarda, M.A. Microencapsulation of maqui (Aristotelia chilensis [Molina] Stuntz) leaf extracts to preserve and control antioxidant properties. Chil. J. Agric. Res. 2013, 73, 17–23. [Google Scholar] [CrossRef] [Green Version]
- González-Villagra, J.; Cohen, J.D.; Reyes-Diaz, M.M. Abscisic acid is involved in phenolic compounds biosynthesis, mainly anthocyanins, in leaves of Aristotelia chilensis plants (Mol.) subjected to drought stress. Physiol. Plant. 2019, 165, 855–866. [Google Scholar] [CrossRef]
- Rivera-Tovar, P.R.; Torres, M.D.; Camilo, C.; Mariotti-Celis, M.S.; Domínguez, H.; Pérez-Correa, J.R. Multi-response optimal hot pressurized liquid recovery of extractable polyphenols from leaves of maqui (Aristotelia chilensis [Mol.] Stuntz). Food Chem. 2021, 357, 129729. [Google Scholar] [CrossRef]
- Sandoval, V.; Femenias, A.; Martínez-Garza, U.; Sanz-Lamora, H.; Castagnini, J.M.; Quifer-Rada, P.; Lamuela-Raventos, R.M.; Marrero, P.F.; Haro, D.; Relat, J. Lyophilized Maqui (Aristotelia chilensis) Berry Induces Browning in the Subcutaneous White Adipose Tissue and Ameliorates the Insulin Resistance in High Fat Diet-Induced Obese Mice. Antioxidants 2019, 8, 360. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhou, G.; Meng, X.S.; Fu, H.Y.; Mo, Q.G.; Wang, Y.W. Photoprotection of maqui berry against ultraviolet B-induced photodamage in vitro and in vivo. Food Funct. 2020, 11, 2749–2762. [Google Scholar] [CrossRef]
- Gironés-Vilaplana, A.; Baenas, N.; Villaño, D.; Speisky, H.; García-Viguera, C.; Moreno, D.A. Evaluation of Latin-American fruits rich in phytochemicals with biological effects. J. Funct. Foods. 2014, 7, 599–608. [Google Scholar] [CrossRef]
- Genskowsky, E.; Puente, L.A.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Determination of polyphenolic profile, antioxidant activity and antibacterial properties of maqui [Aristotelia chilensis (Molina) Stuntz] a Chilean blackberry. J. Sci. Food Agric. 2016, 96, 4235–4242. [Google Scholar] [CrossRef] [PubMed]
- Girones-Vilaplana, A.; Valentao, P.; Moreno, D.A.; Ferreres, F.; García-Viguera, C.; Andrade, P.B. New beverages of lemon juice enriched with the exotic berries maqui, acai, and blackthorn: Bioactive components and in vitro biological properties. J. Agric. Food Chem. 2012, 60, 6571–6580. [Google Scholar] [CrossRef]
- Quispe-Fuentes, I.; Vega-Gálvez, A.; Aranda, M.; Poblete, J.; Pasten, A.; Bilbao-Sainz, C.; Wood, D.; McHugh, T.; Delporte, C. Effects of drying processes on composition, microstructure and health aspects from maqui berries. J. Food Sci. Technol. 2020, 57, 2241–2250. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, O.; Ramos, F. Quantitative analysis of phytosterols in Aristotelia chilensis (Maqui) leaves. Int. Food Res. J. 2016, 23, 822–826. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tohaoco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ferreres, F.; Grosso, C.; Gil-Izquierdo, A.; Fernández, A.; Valentão, P.; Andrade, P.B. Comparing the phenolic profile of Pilocarpus pennatifolius Lem. by HPLC–DAD–ESI/MS n with respect to authentication and enzyme inhibition potential. Ind. Crop. Prod. 2015, 77, 391–401. [Google Scholar] [CrossRef]
- Sánchez-Pujante, P.J.; Gionfriddo, M.; Sabater-Jara, A.B.; Almagro, L.; Pedreño, M.A.; Díaz-Vivancos, P. Enhanced bioactive compound production in broccoli cells due to coronatine and methyl jasmonate is linked to antioxidative metabolism. J. Plant. Physiol. 2020, 248, 153136. [Google Scholar] [CrossRef] [PubMed]
- Sabater-Jara, A.B.; Almagro, L.; Belchi-Navarro, S.; Ferrer, M.A.; Barceló, A.R.; Pedreño, M.A. Induction of sesquiterpenes, phytoesterols and extracellular pathogenesis-related proteins in elicited cell cultures of Capsicum annuum. J. Plant. Physiol. 2010, 167, 1273–1281. [Google Scholar] [CrossRef]
- Ahmed, D.; Khan, M.M.; Saeed, R. Comparative Analysis of Phenolics, Flavonoids, and Antioxidant and Antibacterial Potential of Methanolic, Hexanic and Aqueous Extracts from Adiantum caudatum Leaves. Antioxidants 2015, 4, 394–409. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds 2 | Rt (min) | [M-H]−, m/z | MS2[M-H]−, m/z | |||
|---|---|---|---|---|---|---|
| −132 | −146 | −162 | ||||
| 9 | Ellag ac-Hex | 8.0 | 463 | 301 | ||
| 10 | Ellag ac-Pt | 10.3 | 433 | 301 | ||
| 12 | Ellag ac-Pt | 10.7 | 433 | 301 | ||
| 13 | Ellag ac-Rhmn | 11.0 | 447 | 301 | ||
| 15 | Ellag ac-Rhmn | 11.6 | 447 | 301 | ||
| Compounds 2 | Rt (min) | UV (nm) | [M-H]−, m/z | MS2[M-H]−, m/z | ||
|---|---|---|---|---|---|---|
| −152 | [Aglc-H]− | |||||
| 11 | Qct-3-(Gall)Hex | 10.5 | 268, 288sh, 352 | 615 | 463 (100) | 301 (25) |
| 14 | Qct-3-(Rhmn)Hex | 11.0 | 255, 266sh, 295sh, 354 | 609 | 301 (100) | |
| 16 | Qct-3-(Rhmn)Hex | 11.2 | 256, 266sh, 298sh, 355 | 609 | 301 (100) | |
| 17 | Qct-3-Hex | 11.6 | 256, 266sh, 298sh, 354 | 463 | 301 (100) | |
| 18 | Qct-3-Hex | 11.9 | 256, 266sh, 298sh, 355 | 463 | 301 (100) | |
| 19 | Lut-7-Hex | 12.0 | 256, 266sh, 348 | 447 | 285 (100) | |
| 20 | Qct-3-Pt | 13.1 | 256, 266sh, 298sh, 355 | 433 | 301 (100) | |
| 21 | tetOHFlv-(Rhmn)Hex | 13.6 | 248sh, 268, 290sh, 336 | 593 | 285 (100) | |
| 22 | tetOHFlv-Hex | 14.3 | 248sh, 268, 290sh, 336 | 447 | 285(100) | |
| 23 | triOH-diOMeFlv-der | 14.8 | 252, 266sh, 298sh, 346 | 659 | 329(100) | |
| Sample | Galloyl Acid Derivatives | Caffeoyl Quinic Acids | Ellagitannins | Ellagic Acid Derivatives | Flavonoid Derivatives | Total Polyphenols |
|---|---|---|---|---|---|---|
| AP S | 777.77 ± 15.65 c | 89.67 ± 0.70 b | 3306.80 ± 33.88 b | 27.99 ± 0.33 b | 894.69 ± 2.67 b | 5096.92 ± 37.42 c |
| BS S | 1004.52 ± 10.28 b | 87.83 ± 1.30 b | 3355.70 ± 27.61 b | 36.04 ± 1.45 c | 928.35 ± 4.96 a | 5412.43 ± 29.94 b |
| AP W | 400.96 ± 5.71 d | 21.58 ± 0.14 c | 1055.90 ± 7.28 c | 8.13 ± 0.10 d | 379.24 ± 2.18 d | 1865.81 ± 9.51 d |
| BS W | 362.33 ± 9.20 e | 6.90 ± 0.09 c | 915.33 ± 13.98 c | 5.33 ± 0.05 d | 230.77 ± 2.04 e | 1520.66 ± 16.86 e |
| IVITRO | 4973.71 ± 56.33 a | 852.37 ± 17.79 a | 4513.45 ± 84.90 a | 177.83 ± 2.08 a | 523.51 ± 11.42 c | 10,611.44 ± 78.41 a |
| Leaf Type | Quercetin | Catechin | Kaempferol | 3-Caffeloylquinic Acid |
|---|---|---|---|---|
| In vitro | 35.44 ± 3.54 b | 6.69 ± 0.67 b | 25.93 ± 2.59 b | |
| BS S | 145.73 ± 14.57 a | 132.59 ± 13.26 a | 25.93 ± 2.59 a | 253.77 ± 25.38 a |
| Types of Leaves | TFC (mg RE mL−1) | |
|---|---|---|
| 1 | BS Winter | 0.086 ± 0.004 a |
| 2 | AP Winter | 0.074 ± 0.001 c |
| 3 | BS Spring | 0.081 ± 0.005 bc |
| 4 | AP Spring | 0.083 ± 0.002 b |
| 5 | In Vitro | 0.061 ± 0.002 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crisóstomo-Ayala, K.A.; Sabater-Jara, A.B.; Pérez Manriquez, C.; Ferreres, F.; Gil-Izquierdo, Á.; Pedreño, M.Á.; Hernández de la Torre, M.; Sanchez-Olate, M.; Ríos Leal, D.G. Comparative Study of Metabolomic Profile and Antioxidant Content of Adult and In Vitro Leaves of Aristotelia chilensis. Plants 2022, 11, 37. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11010037
Crisóstomo-Ayala KA, Sabater-Jara AB, Pérez Manriquez C, Ferreres F, Gil-Izquierdo Á, Pedreño MÁ, Hernández de la Torre M, Sanchez-Olate M, Ríos Leal DG. Comparative Study of Metabolomic Profile and Antioxidant Content of Adult and In Vitro Leaves of Aristotelia chilensis. Plants. 2022; 11(1):37. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11010037
Chicago/Turabian StyleCrisóstomo-Ayala, Karina Andrea, Ana Belén Sabater-Jara, Claudia Pérez Manriquez, Federico Ferreres, Ángel Gil-Izquierdo, Maria Ángeles Pedreño, Martha Hernández de la Torre, Manuel Sanchez-Olate, and Darcy Graciela Ríos Leal. 2022. "Comparative Study of Metabolomic Profile and Antioxidant Content of Adult and In Vitro Leaves of Aristotelia chilensis" Plants 11, no. 1: 37. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11010037