
Role of Phaseolus vulgaris L. in the Prevention of Cardiovascular Diseases—Cardioprotective Potential of Bioactive Compounds

 ,
, 
Abstract
:1. Introduction
2. Phaseolus vulgaris L.
3. Relationship between Nutritional Composition and Health Benefits
4. Role of Beans in CVD
4.1. Effect on Hemostasis and Platelet Aggregation
4.2. Effect on the Endothelium
4.3. Effect on Inflammation
4.4. Effect on Metabolic Syndrome
4.5. Effect of Beans on Atherosclerosis
4.6. Other Studies Related to Cardioprotective Role
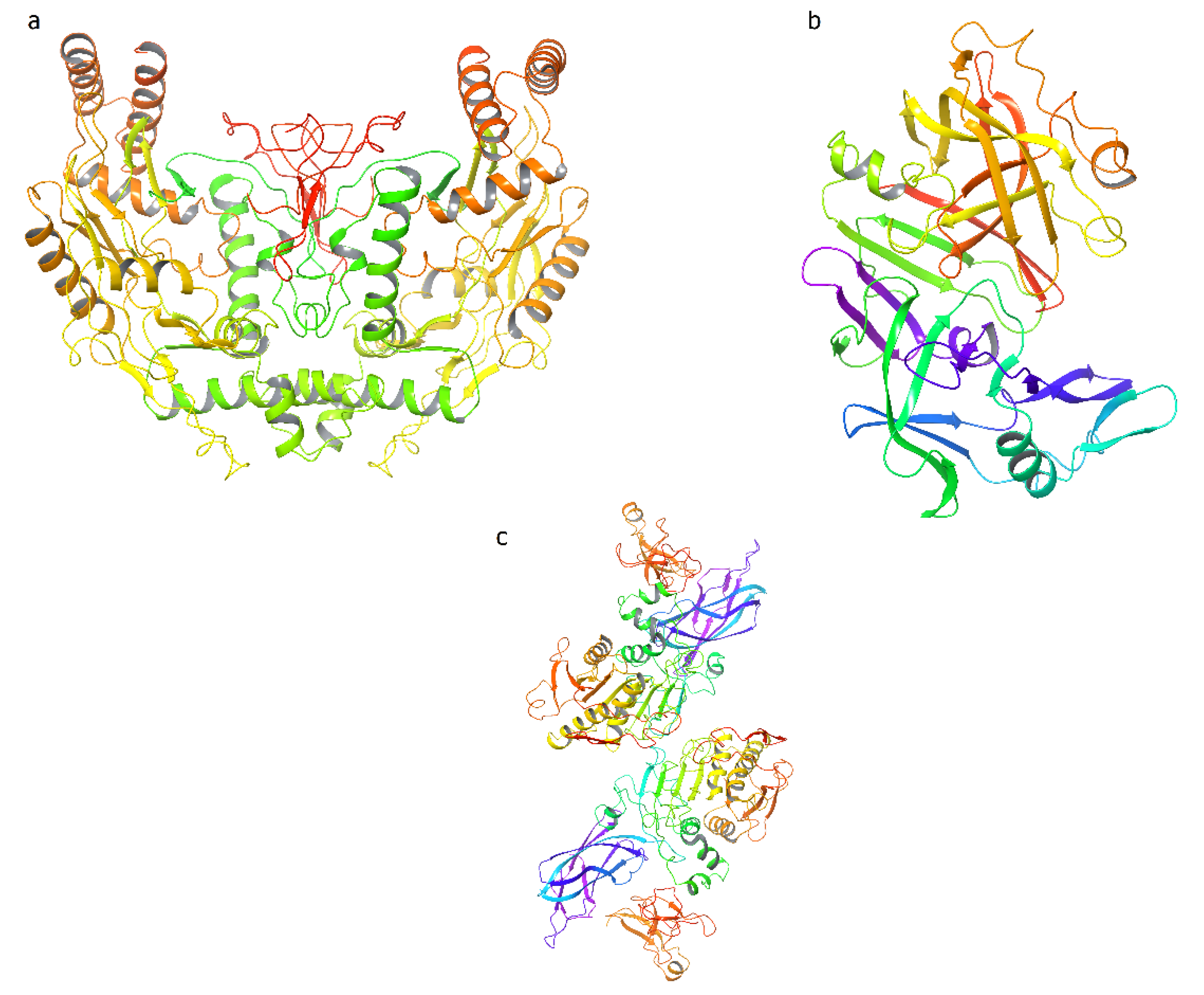
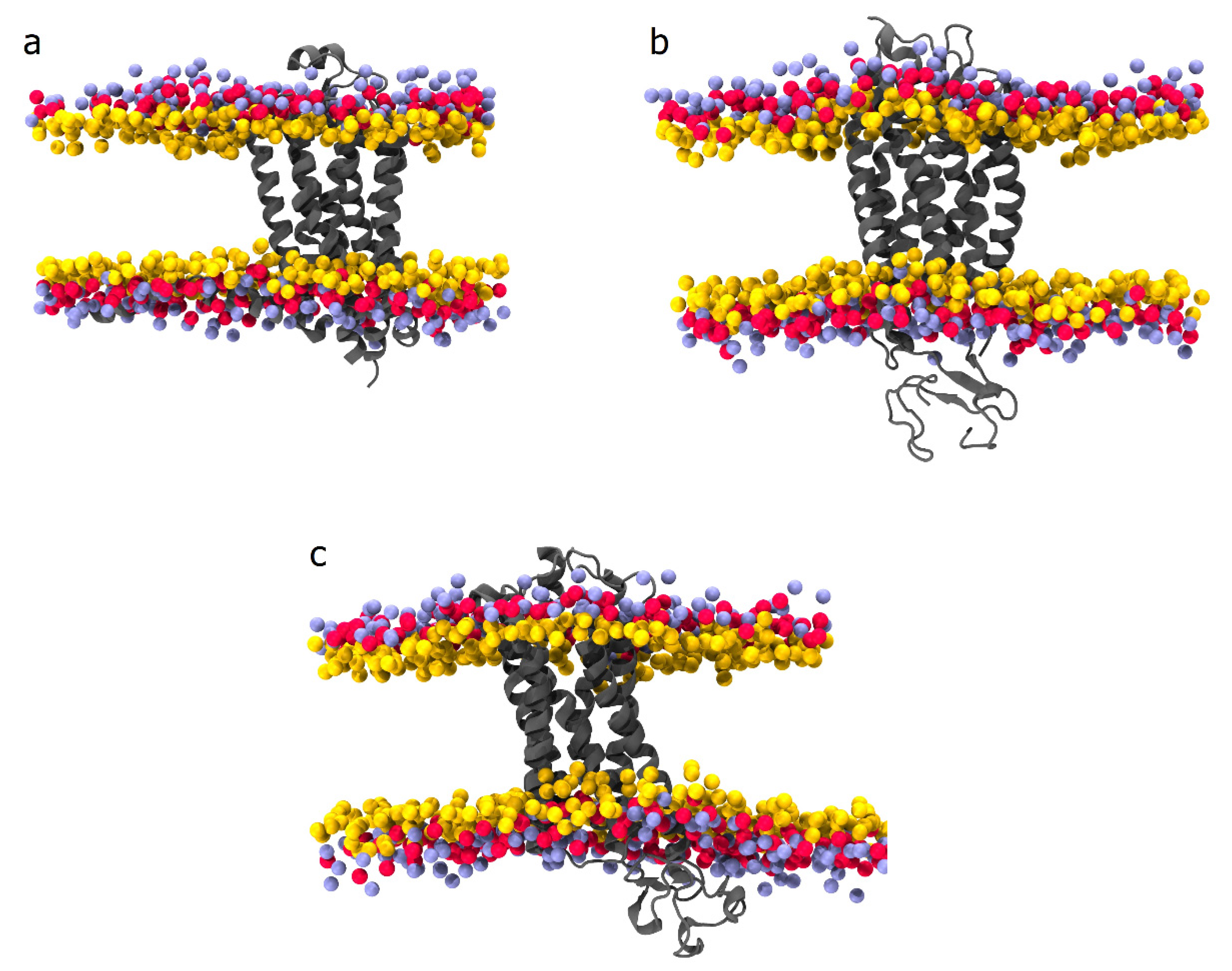
5. In Silico Assays of Bioactive Compounds from Phaseolus vulgaris L.
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Irfan, M.; Kwon, T.-H.; Lee, D.-H.; Hong, S.-B.; Oh, J.-W.; Kim, S.-D.; Rhee, M.H. Antiplatelet and antithrombotic effects of Epimedium koreanum Nakai. Evid.-Based Complement. Altern. Med. 2021, 2021, 7071987. [Google Scholar] [CrossRef]
- Olas, B. Dietary supplements with antiplatelet activity: A solution for everyone? Adv. Nutr. 2018, 9, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.L.; Yang, E.C.; Zhang, Z. Income disparities and cardiovascular risk factors among adolescents. Pediatrics 2018, 142, e20181089. [Google Scholar] [CrossRef] [Green Version]
- Mensah, G.A.; Roth, G.A.; Fuster, V. The global burden of cardiovascular diseases and risk factors: 2020 and beyond. J. Am. Coll. Cardiol. 2019, 74, 2529–2532. [Google Scholar] [CrossRef]
- Rosengren, A.; Hawken, S.; Ôunpuu, S.; Sliwa, K.; Zubaid, M.; Almahmeed, W.A.; Ngu Blackett, K.; Sitthi-Amorn, C.; Sato, H.; Yusuf, S.; et al. Association of psychosocial risk factors with risk of acute myocardial infarction in 11,119 cases and 13,648 controls from 52 countries (the INTERHEART study): Case-control study. Lancet 2004, 364, 953–962. [Google Scholar] [CrossRef]
- Cuadrado-Godia, E.; Jamthikar, A.D.; Gupta, D.; Khanna, N.N.; Araki, T.; Maniruzzaman, M.; Saba, L.; Nicolaides, A.; Sharma, A.; Omerzu, T.; et al. Ranking of stroke and cardiovascular risk factors for an optimal risk calculator design: Logistic regression approach. Comput. Biol. Med. 2019, 108, 182–195. [Google Scholar] [CrossRef]
- Kalantar-Zadeh, K.; Block, G.; Humphreys, M.H.; Kopple, J.D. Reverse epidemiology of cardiovascular risk factors in maintenance dialysis patients. Kidney Int. 2003, 63, 793–808. [Google Scholar] [CrossRef] [Green Version]
- Niccoli, T.; Partridge, L. Ageing as a risk factor for disease. Curr. Biol. 2012, 22, R741–R752. [Google Scholar] [CrossRef] [Green Version]
- Strait, J.B.; Lakatta, E.G. Aging-associated cardiovascular changes and their relationship to heart failure. Heart Fail. Clin. 2012, 8, 143–164. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liu, Y.; Wang, X.; Yang, L.; Li, H.; Wang, Y.; Liu, M.; Zhao, X.; Xie, Y.; Yang, Y.; et al. SARS-CoV-2 binds platelet ACE2 to enhance thrombosis in COVID-19. J. Hematol. Oncol. 2020, 13, 120. [Google Scholar] [CrossRef]
- Huang, J.; Li, X.; Shi, X.; Zhu, M.; Wang, J.; Huang, S.; Huang, X.; Wang, H.; Li, L.; Deng, H.; et al. Platelet integrin αIIbβ3: Signal transduction, regulation, and its therapeutic targeting. J. Hematol. Oncol. 2019, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Meshkini, A.; Tahmasbi, M. Antiplatelet aggregation activity of walnut hull extract via suppression of reactive oxygen species generation and caspase activation. J. Acupunct. Meridian Stud. 2017, 10, 193–203. [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Thrombosis and COVID-19: The potential role of nutrition. Front. Nutr. 2020, 7, 177. [Google Scholar] [CrossRef]
- Tsoupras, A.; Lordan, R.; Zabetakis, I. Inflammation, not cholesterol, is a cause of chronic disease. Nutrients 2018, 10, 604. [Google Scholar] [CrossRef] [Green Version]
- Yu, E.; Malik, V.S.; Hu, F.B. Cardiovascular disease prevention by diet modification: JACC health promotion series. J. Am. Coll. Cardiol. 2018, 72, 914–926. [Google Scholar] [CrossRef]
- Calder, P.C.; Carr, A.C.; Gombart, A.F.; Eggersdorfer, M. optimal nutritional status for a well-functioning immune system is an important factor to protect against viral infections. Nutrients 2020, 12, 1181. [Google Scholar] [CrossRef] [Green Version]
- Bartimoccia, S.; Nocella, C.; Pastori, D.; Pignatelli, P.; Carnevale, R. Platelet oxidative stress and antioxidant nutrients. J. Vasc. Med. Surg. 2014, 2, 1000164. [Google Scholar]
- Violi, F.; Pignatelli, P. Platelet oxidative stress and thrombosis. Thromb. Res. 2012, 129, 378–381. [Google Scholar] [CrossRef]
- Câmara, C.R.S.; Urrea, C.A.; Schlegel, V. Pinto beans (Phaseolus vulgaris L.) as a functional food: Implications on human health. Agriculture 2013, 3, 90–111. [Google Scholar] [CrossRef]
- Bellucci, E.; Bitocchi, E.; Rau, D.; Rodriguez, M.; Biagetti, E.; Giardini, A.; Attene, G.; Nanni, L.; Papa, R. Genomics of origin, domestication and evolution of Phaseolus vulgaris. Genom. Plant Genet. Resour. 2014, 483–507. [Google Scholar] [CrossRef]
- Yanez, E.; Zacarias, I.; Aguayo, M.; Vasquez, M.; Guzman, E. Nutritive value evaluated on rats of new cultivars of common beans (Phaseolus vulgaris) released in Chile. Plant Foods Hum. Nutr. 1995, 47, 301–307. [Google Scholar] [CrossRef]
- de Almeida Costa, G.E.; da Silva Queiroz-Monici, K.; Reis, S.M.P.M.; de Oliveira, A.C. Chemical composition, dietary fibre and resistant starch contents of raw and cooked pea, common bean, chickpea and lentil legumes. Food Chem. 2006, 94, 327–330. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Harnly, J.M.; Pastor-Corrales, M.S.; Luthria, D.L. The polyphenolic profiles of common bean (Phaseolus vulgaris L.). Food Chem. 2008, 107, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Tharanathan, R.N.; Mahadevamma, S. Grain legumes—A boon to human nutrition. Trends Food Sci. Technol. 2003, 14, 507–518. [Google Scholar] [CrossRef]
- Shimelis, E.A.; Rakshit, S.K. Proximate composition and physico-chemical properties of improved dry bean (Phaseolus vulgaris L.) varieties grown in Ethiopia. LWT-Food Sci. Technol. 2005, 38, 331–338. [Google Scholar] [CrossRef]
- Hayat, I.; Ahmad, A.; Masud, T.; Ahmed, A.; Bashir, S. Nutritional and health perspectives of beans (Phaseolus vulgaris L.): An overview. Crit. Rev. Food Sci. Nutr. 2014, 54, 580–592. [Google Scholar] [CrossRef]
- Siddiq, M.; Ravi, R.; Harte, J.B.; Dolan, K.D. Physical and functional characteristics of selected dry bean (Phaseolus vulgaris L.) flours. LWT-Food Sci. Technol. 2010, 43, 232–237. [Google Scholar] [CrossRef]
- Landon, A.J. The “how” of the three sisters: The origins of agriculture in Mesoamerica and the human niche. Neb. Anthropol. 2008, 23, 110–124. [Google Scholar]
- Celmeli, T.; Sari, H.; Canci, H.; Sari, D.; Adak, A.; Eker, T.; Toker, C. The nutritional content of common bean (Phaseolus vulgaris L.) landraces in comparison to modern varieties. Agronomy 2018, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- Gepts, P. Phaseolus vulgaris (beans). Encycl. Genet. 2001, 1444–1445. [Google Scholar]
- Singh, S.P. Broadening the genetic base of common bean cultivars: A review. Crop Sci. 2001, 41, 1659–1675. [Google Scholar] [CrossRef]
- Bitocchi, E.; Rau, D.; Bellucci, E.; Rodriguez, M.; Murgia, M.L.; Gioia, T.; Santo, D.; Nanni, L.; Attene, G.; Papa, R. Beans (Phaseolus ssp.) as a model for understanding crop evolution. Front. Plant Sci. 2017, 8, 722. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Gallegos, J.A.; Kelly, J.D.; Gepts, P. Prebreeding in common bean and use of genetic diversity from wild germplasm. Crop Sci. 2007, 47, S-44–S-59. [Google Scholar] [CrossRef] [Green Version]
- Marquezi, M.; Gervin, V.M.; Watanabe, L.B.; Moresco, R.; Amante, E.R. Chemical and functional properties of different common Brazilian bean (Phaseolus vulgaris L.) cultivars. Braz. J. Food Technol. 2017, 20, e2016006. [Google Scholar] [CrossRef] [Green Version]
- Leterme, P.; Muũoz, L.C. Factors influencing pulse consumption in Latin America. Br. J. Nutr. 2002, 88, 251–254. [Google Scholar] [CrossRef]
- Luthria, D.L.; Pastor-Corrales, M.A. Phenolic acids content of fifteen dry edible bean (Phaseolus vulgaris L.) varieties. J. Food Compos. Anal. 2006, 19, 205–211. [Google Scholar] [CrossRef]
- Akibode, C.S.; Maredia, M.K. Global and Regional Trends in Production, Trade and Consumption of Food Legume Crops. 2012. Available online: https://ageconsearch.umn.edu/record/136293 (accessed on 15 October 2012).
- Paredes, M.; Becerra, V.; Tay, J. Inorganic nutritional composition of common bean (Phaseolus vulgaris L.) genotypes race Chile. Chil. J. Agric. Res. 2009, 69, 486–495. [Google Scholar] [CrossRef]
- Shellie-Dessert, K.C.; Bliss, F.A. Genetic improvement of food quality factors. In Common Beans: Research for Crop Improvement; Schoonhoven, A., vanVoysest, O., Eds.; CAB International: Wallingford, UK, 1991; pp. 649–677. [Google Scholar]
- Choung, M.-G.; Choi, B.-R.; An, Y.-N.; Chu, Y.-H.; Cho, Y.-S. Anthocyanin profile of Korean cultivated kidney bean (Phaseolus vulgaris L.). J. Agric. Food Chem. 2003, 51, 7040–7043. [Google Scholar] [CrossRef]
- Aparicio-Fernández, X.; García-Gasca, T.; Yousef, G.G.; Lila, M.A.; González de Mejia, E.; Loarca-Pina, G. Chemopreventive activity of polyphenolics from black Jamapa bean (Phaseolus vulgaris L.) on HeLa and HaCaT cells. J. Agric. Food Chem. 2006, 54, 2116–2122. [Google Scholar] [CrossRef]
- Hosfield, G.L.; Varner, G.V.; Uebersax, M.A.; Kelly, J.D. Registration of ‘Merlot’ small red bean. Crop Sci. 2004, 44, 351–353. [Google Scholar] [CrossRef]
- Amir, Y.; Haenni, A.L.; Youyou, A. Differences in the biochemical composition of dry legumes cultivated in north Algeria. Electron J. Environ. Agric Food Chem. 2006, 5, 1411–1418. [Google Scholar]
- Baginsky, C.; Ramos, L. Situación de las legumbres en Chile: Una mirada agronómica. Rev. Chil. Nutr. 2018, 45, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Broughton, W.J.; Hernandez, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef] [Green Version]
- Baginsky, C.; Brito, B.; Scherson, R.; Pertuzé, R.; Seguel, O.; Cañete, A.; Araneda, C.; Johnson, W. Genetic diversity of Rhizobium from nodulating beans grown in a variety of Mediterranean climate soils of Chile. Arch. Microbiol. 2015, 197, 419–429. [Google Scholar] [CrossRef]
- Olanipekun, O.T.; Omenna, E.C.; Olapade, O.A.; Suleiman, P.; Omodara, O.G. Effect of boiling and roasting on the nutrient composition of kidney beans seed flour. Sky J. Food Sci. 2015, 4, 24–29. [Google Scholar]
- Singh, S.P.; Gepts, P.; Debouck, D.G. Races of common bean (Phaseolus vulgaris, Fabaceae). Econ. Bot. 1991, 45, 379–396. [Google Scholar] [CrossRef]
- Bascur, G.B.; Tay, J.U. Collection, characterization and use of genetic variation in Chilean bean germplasm (Phaseolus vulgaris L.) 1. Agric. Técnica 2005, 65, 135–146. [Google Scholar]
- Trucchi, E.; Benazzo, A.; Lari, M.; Iob, A.; Vai, S.; Nanni, L.; Bellucci, E.; Bitocchi, E.; Raffini, F.; Xu, C.; et al. Ancient genomes reveal early Andean farmers selected common beans while preserving diversity. Nat. Plants 2021, 7, 123–128. [Google Scholar] [CrossRef]
- Yang, Q.Q.; Gan, R.Y.; Ge, Y.Y.; Zhang, D.; Corke, H. Polyphenols in common beans (Phaseolus vulgaris L.): Chemistry, analysis, and factors affecting composition. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1518–1539. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.X.; Tang, Y.; Marcone, M.F.; Pauls, P.K.; Zhang, B.; Liu, R.; Tsao, R. Characterization of free, conjugated and bound phenolics and lipophilic antioxidants in regular-and non-darkening cranberry beans (Phaseolus vulgaris L.). Food Chem. 2015, 185, 298–308. [Google Scholar] [CrossRef]
- Teixeira-Guedes, C.I.; Oppolzer, D.; Barros, A.I.; Pereira-Wilson, C. Impact of cooking method on phenolic composition and antioxidant potential of four varieties of Phaseolus vulgaris L. and Glycine max L. LWT 2019, 103, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Los, F.G.B.; Zielinski, A.A.F.; Wojeicchowski, J.P.; Nogueira, A.; Demiate, I.M. Beans (Phaseolus vulgaris L.): Whole seeds with complex chemical composition. Curr. Opin. Food Sci. 2018, 19, 63–71. [Google Scholar] [CrossRef]
- Madrera, R.R.; Valles, B.S. Development and validation of ultrasound assisted extraction (UAE) and HPLC-DAD method for determination of polyphenols in dry beans (Phaseolus vulgaris). J. Food Compos. Anal. 2020, 85, 103334. [Google Scholar] [CrossRef]
- Yang, Q.-Q.; Gan, R.-Y.; Ge, Y.-Y.; Zhang, D.; Corke, H. Ultrasonic treatment increases extraction rate of common bean (Phaseolus vulgaris L.) antioxidants. Antioxidants 2019, 8, 83. [Google Scholar] [CrossRef] [Green Version]
- Pitura, K.; Arntfield, S.D. Characteristics of flavonol glycosides in bean (Phaseolus vulgaris L.) seed coats. Food Chem. 2019, 272, 26–32. [Google Scholar] [CrossRef]
- Flores-Sosa, Á.R.; Aquino-Bolaños, E.N.; Cardador-Martínez, A.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Carrillo-Rodríguez, J.C.; Jiménez, J.E.A. Variation in protein and amino acids content among landraces of common bean (Phaseolus vulgaris L.). Emir. J. Food Agric. 2020, 32, 750–760. [Google Scholar] [CrossRef]
- Yoshida, H.; Yoshida, N.; Tomiyama, Y.; Mizushina, Y. Fatty acid characteristics of triacylglycerols and phospholipids in adzuki beans (Vigna angularis). Food Sci. Technol. Res. 2010, 16, 209–214. [Google Scholar] [CrossRef]
- Kotha, R.R.; Finley, J.W.; Luthria, D.L. Determination of soluble mono, di, and oligosaccharide content in 23 dry beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2020, 68, 6412–6419. [Google Scholar] [CrossRef]
- Díaz-Batalla, L.; Widholm, J.M.; Fahey, G.C.; Castaño-Tostado, E.; Paredes-López, O. Chemical components with health implications in wild and cultivated Mexican common bean seeds (Phaseolus vulgaris L.). J. Agric. Food Chem. 2006, 54, 2045–2052. [Google Scholar] [CrossRef]
- Pereira, H.S.; Alvares, R.C.; Melo, L.C.; Da Costa, A.F.; De Carvalho, H.W.L. Culinary and nutritional quality of common bean lines with Carioca grain type and interaction with environments. Rev. Ceres 2017, 64, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Nouri, F.; Sarrafzadegan, N.; Mohammadifard, N.; Sadeghi, M.; Mansourian, M. Intake of legumes and the risk of cardiovascular disease: Frailty modeling of a prospective cohort study in the Iranian middle-aged and older population. Eur. J. Clin. Nutr. 2016, 70, 217–221. [Google Scholar] [CrossRef]
- Batista, K.A.; Prudêncio, S.H.; Fernandes, K.F. Wheat bread enrichment with hard-to-cook bean extruded flours: Nutritional and acceptance evaluation. J. Food Sci. 2011, 76, S108–S113. [Google Scholar] [CrossRef]
- Seena, S.; Sridhar, K.R.; Bhagya, B. Biochemical and biological evaluation of an unconventional legume, Canavalia maritima of coastal sand dunes of India. Trop. Subtrop. Agroecosyst. 2005, 5, 1–14. [Google Scholar]
- Słupski, J. Effect of cooking and sterilisation on the composition of amino acids in immature seeds of flageolet bean (Phaseolus vulgaris L.) cultivars. Food Chem. 2010, 121, 1171–1176. [Google Scholar] [CrossRef]
- Derbyshire, E.; Wright, D.J.; Boulter, D. Legumin and vicilin, storage proteins of legume seeds. Phytochemistry 1976, 15, 3–24. [Google Scholar] [CrossRef]
- Guzmán-Maldonado, S.H.; Acosta-Gallegos, J.; Paredes-López, O. Protein and mineral content of a novel collection of wild and weedy common bean (Phaseolus vulgaris L.). J. Sci. Food Agric. 2000, 80, 1874–1881. [Google Scholar] [CrossRef]
- Baptista, A.; Pinho, O.; Pinto, E.; Casal, S.; Mota, C.; Ferreira, I.M. Characterization of protein and fat composition of seeds from common beans (Phaseolus vulgaris L.), cowpea (Vigna unguiculata L. Walp) and bambara groundnuts (Vigna subterranea L. Verdc) from Mozambique. J. Food Meas. Charact. 2017, 11, 442–450. [Google Scholar] [CrossRef]
- Carbas, B.; Machado, N.; Oppolzer, D.; Ferreira, L.; Queiroz, M.; Brites, C.; Rosa, E.A.; Barros, A.I. Nutrients, antinutrients, phenolic composition, and antioxidant activity of common bean cultivars and their potential for food applications. Antioxidants 2020, 9, 186. [Google Scholar] [CrossRef] [Green Version]
- Boye, J.; Wijesinha-Bettoni, R.; Burlingame, B. Protein quality evaluation twenty years after the introduction of the protein digestibility corrected amino acid score method. Br. J. Nutr. 2012, 108, S183–S211. [Google Scholar] [CrossRef] [Green Version]
- Grimble, R.F. The effects of sulfur amino acid intake on immune function in humans. J. Nutr. 2006, 136, 1660S–1665S. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Torres, A.; Osorio-Díaz, P.; Islas-Hernández, J.J.; Tovar, J.; Paredes-López, O.; Bello-Pérez, L.A. Starch digestibility of five cooked black bean (Phaseolus vulgaris L.) varieties. J. Food Compos. Anal. 2004, 17, 605–612. [Google Scholar] [CrossRef]
- Ovando-Martínez, M.; Bello-Pérez, L.A.; Whitney, K.; Osorio-Díaz, P.; Simsek, S. Starch characteristics of bean (Phaseolus vulgaris L.) grown in different localities. Carbohydr. Polym. 2011, 85, 54–64. [Google Scholar] [CrossRef]
- Tharanathan, R.N. Food-derived carbohydrates—Structural complexity and functional diversity. Crit. Rev. Biotechnol. 2002, 22, 65–84. [Google Scholar] [CrossRef]
- Zhou, Y.; Hoover, R.; Liu, Q. Relationship between α-amylase degradation and the structure and physicochemical properties of legume starches. Carbohydr. Polym. 2004, 57, 299–317. [Google Scholar] [CrossRef]
- Reynoso-Camacho, R.; Ramos-Gomez, M.; Loarca-Pina, G. Bioactive components in common beans (Phaseolus vulgaris L.). Adv. Agric. Food Biotechnol. 2006, 10, 217–236. [Google Scholar]
- Yoshida, H.; Tomiyama, Y.; Mizushina, Y. Characterization in the fatty acid distributions of triacylglycerols and phospholipids in kidney beans (Phaseolus vulgaris L.). J. Food Lipids 2005, 12, 169–180. [Google Scholar] [CrossRef]
- Ryan, E.; Galvin, K.; O’Connor, T.P.; Maguire, A.; O’Brien, N.M. Phytosterol, squalene, tocopherol content and fatty acid profile of selected seeds, grains, and legumes. Mater. Veg. 2007, 62, 85–91. [Google Scholar] [CrossRef]
- Grela, E.R.; Günter, K.D. Fatty acid composition and tocopherol content of some legume seeds. Anim. Feed Sci. Technol. 1995, 52, 325–331. [Google Scholar] [CrossRef]
- Beninger, C.W.; Hosfield, G.L. Antioxidant activity of extracts, condensed tannin fractions, and pure flavonoids from Phaseolus vulgaris L. seed coat color genotypes. J. Agric. Food Chem. 2003, 51, 7879–7883. [Google Scholar] [CrossRef]
- Vucenik, I.; Shamsuddin, A.M. Protection against cancer by dietary IP6 and inositol. Nutr. Cancer 2006, 55, 109–125. [Google Scholar] [CrossRef]
- Bennink, M.R. Consumption of black beans and navy beans (Phaseolus vulgaris) reduced azoxymethane-induced colon cancer in rats. Nutr. Cancer 2002, 44, 60–65. [Google Scholar] [CrossRef]
- Xu, B.; Chang, S.K.C. Comparative study on antiproliferation properties and cellular antioxidant activities of commonly consumed food legumes against nine human cancer cell lines. Food Chem. 2012, 134, 1287–1296. [Google Scholar] [CrossRef]
- Sangronis, E.; Machado, C.J. Influence of germination on the nutritional quality of PhaseolusLWT vulgaris and Cajanus cajan. LWT-Food Sci. Technol. 2007, 40, 116–120. [Google Scholar] [CrossRef]
- Mitchell, D.C.; Lawrence, F.R.; Hartman, T.J.; Curran, J.M. Consumption of dry beans, peas, and lentils could improve diet quality in the US population. J. Am. Diet. Assoc. 2009, 109, 909–913. [Google Scholar] [CrossRef]
- Rosa, C.O.; Costa, N.M.; Leal, P.F.; Oliveira, T.T. The cholesterol-lowering effect of black beans (Phaseolus vulgaris, L.) without hulls in hypercholesterolemic rats. Arch. Latinoam. Nutr. 1998, 48, 299–305. [Google Scholar]
- Anderson, J.W.; Smith, B.M.; Washnock, C.S. Cardiovascular and renal benefits of dry bean and soybean intake. Am. J. Clin. Nutr. 1999, 70, 464S–474S. [Google Scholar] [CrossRef] [Green Version]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Romani, A. Rapid tests to assess the antioxidant activity of Phaseolus vulgaris L. dry beans. J. Agric. Food Chem. 2005, 53, 3053–3056. [Google Scholar] [CrossRef]
- Olas, B. The multifunctionality of berries toward blood platelets and the role of berry phenolics in cardiovascular disorders. Platelets 2017, 28, 540–549. [Google Scholar] [CrossRef]
- Fuentes, E.; Palomo, I. Antiplatelet effects of natural bioactive compounds by multiple targets: Food and drug interactions. J. Funct. Foods 2014, 6, 73–81. [Google Scholar] [CrossRef]
- Rodríguez-Azúa, R.; Quinteros, E.F.; Olate-Briones, A.; Moore-Carrasco, R. Phaseolus vulgaris exerts an inhibitory effect on platelet aggregation through AKT dependent way. Prev. Nutr. Food Sci. 2018, 23, 102. [Google Scholar] [CrossRef]
- Vaiyapuri, S.; Roweth, H.; Ali, M.S.; Unsworth, A.J.; Stainer, A.R.; Flora, G.D.; Crescente, M.; Jones, C.I.; Moraes, L.A.; Gibbins, J.M. Pharmacological actions of nobiletin in the modulation of platelet function. Br. J. Pharmacol. 2015, 172, 4133–4145. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, P.; Gould, N.L.; Sidhu, P. Interaction of lectins with human platelets: Effects of platelet stimulation by thrombin and ristocetin. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1979, 586, 574–583. [Google Scholar] [CrossRef]
- Signorello, M.G.; Ravera, S.; Leoncini, G. Lectin-induced oxidative stress in human platelets. Redox Biol. 2020, 32, 101456. [Google Scholar] [CrossRef]
- Kinlough-Rathbone, R.L.; Mustard, J.F.; Packham, M.A.; Perry, D.W.; Reimers, H.-J.; Cazenave, J.-P. Properties of washed human platelets. Thromb. Haemost. 1977, 37, 291–308. [Google Scholar] [CrossRef]
- Cazenave, J.P.; Guccione, M.A.; Packham, M.A.; Mustard, J.F. Effects of cephalothin and penicillin G on platelet function in vitro. Br. J. Haematol. 1977, 35, 135–152. [Google Scholar] [CrossRef]
- Signorello, M.G.; Leoncini, G. The Ca2+/calmodulin kinase/AMP-activated protein kinase pathway regulates the lectin Phaseolus vulgaris agglutinin induced NO production in human platelets. Integr. Mol. Med. 2019, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-R.; Zhu, Y.; Halushka, P.V.; Lincoln, T.M.; Mendelsohn, M.E. Mechanism of platelet inhibition by nitric oxide: In vivo phosphorylation of thromboxane receptor by cyclic GMP-dependent protein kinase. Proc. Natl. Acad. Sci. USA 1998, 95, 4888–4893. [Google Scholar] [CrossRef] [Green Version]
- Schemmer, P.; Zhong, Z.; Galli, U.; Wheeler, M.D.; Xiangli, L.; Bradford, B.U.; Conzelmann, L.O.; Forman, D.; Boyer, J.; Thurman, R.G. Glycine reduces platelet aggregation. Amino Acids 2013, 44, 925–931. [Google Scholar] [CrossRef] [Green Version]
- Wolf, A.; Zalpour, C.; Theilmeier, G.; Wang, B.-Y.; Ma, A.; Anderson, B.; Tsao, P.S.; Cooke, J.P. Dietary L-arginine supplementation normalizes platelet aggregation in hypercholesterolemic humans. J. Am. Coll. Cardiol. 1997, 29, 479–485. [Google Scholar] [CrossRef]
- Thompson, K.; Hosking, H.; Pederick, W.; Singh, I.; Santhakumar, A.B. The effect of anthocyanin supplementation in modulating platelet function in sedentary population: A randomised, double-blind, placebo-controlled, cross-over trial. Br. J. Nutr. 2017, 118, 368–374. [Google Scholar] [CrossRef]
- Adili, R.; Hawley, M.; Holinstat, M. Regulation of platelet function and thrombosis by omega-3 and omega-6 polyunsaturated fatty acids. Prostaglandins Other Lipid Mediat. 2018, 139, 10–18. [Google Scholar] [CrossRef]
- Dratewka-Kos, E.; Tinker, D.O.; Kindl, B. Unsaturated fatty acids inhibit ADP-arachidonate-induced platelet aggregation without affecting thromboxane synthesis. Biochem. Cell Biol. 1986, 64, 906–913. [Google Scholar] [CrossRef]
- McEwen, B.J.; Morel-Kopp, M.-C.; Chen, W.; Tofler, G.H.; Ward, C.M. Effects of omega-3 polyunsaturated fatty acids on platelet function in healthy subjects and subjects with cardiovascular disease. Semin. Thromb. Hemost. 2013, 39, 25–32. [Google Scholar] [CrossRef]
- Hodgson, J.M.; Wahlqvist, M.L.; Boxall, J.A.; Balazs, N.D. Can linoleic acid contribute to coronary artery disease? Am. J. Clin. Nutr. 1993, 58, 228–234. [Google Scholar] [CrossRef]
- Dinicolantonio, J.; Okeefe, J. Importance of maintaining a low omega-6/omega-3 ratio for reducing platelet aggregation, coagulation and thrombosis. Open Heart 2019, 6, e001011. [Google Scholar] [CrossRef]
- Miyake, K.; Tanaka, T.; McNeil, P.L. Lectin-based food poisoning: A new mechanism of protein toxicity. PLoS ONE 2007, 2, e687. [Google Scholar] [CrossRef] [Green Version]
- Luna-Vital, D.A.; De Mejía, E.G.; Mendoza, S.; Loarca-Piña, G. Peptides present in the non-digestible fraction of common beans (Phaseolus vulgaris L.) inhibit the angiotensin-I converting enzyme by interacting with its catalytic cavity independent of their antioxidant capacity. Food Funct. 2015, 6, 1470–1479. [Google Scholar] [CrossRef]
- Gomes, M.J.; Lima, S.L.; Alves, N.E.G.; Assis, A.; Moreira, M.E.; Toledo, R.C.; Rosa, C.O.; Teixeira, O.R.; Bassinello, P.Z.; De Mejía, E.G.; et al. Common bean protein hydrolysate modulates lipid metabolism and prevents endothelial dysfunction in BALB/c mice fed an atherogenic diet. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Davis, H.R.; Glagov, S. Lectin binding to distinguish cell types in fixed atherosclerotic arteries. Atherosclerosis 1986, 61, 193–203. [Google Scholar] [CrossRef]
- Zehr, K.R.; Walker, M.K. Omega-3 polyunsaturated fatty acids improve endothelial function in humans at risk for atherosclerosis: A review. Prostaglandins Other Lipid Mediat. 2018, 134, 131–140. [Google Scholar] [CrossRef]
- Kakoki, M.; Kim, H.-S.; Edgell, C.-J.S.; Maeda, N.; Smithies, O.; Mattson, D.L. Amino acids as modulators of endothelium-derived nitric oxide. Am. J. Physiol.-Ren. Physiol. 2006, 291, F297–F304. [Google Scholar] [CrossRef] [Green Version]
- Gamboa-Gomez, C.I.; Rocha-Guzman, N.E.; Gallegos-Infante, J.A.; Moreno-Jimenez, M.R.; Vazquez-Cabral, B.D.; Gonzalez-Laredo, R.F. Plants with potential use on obesity and its complications. EXCLI J. 2015, 14, 809–831. [Google Scholar]
- Lefkowitz, D.L.; Lefkowitz, S.S. Macrophage–neutrophil interaction: A paradigm for chronic inflammation revisited. Immunol. Cell Biol. 2001, 79, 502–506. [Google Scholar] [CrossRef]
- Williams, C.S.; Mann, M.; DuBois, R.N. The role of cyclooxygenases in inflammation, cancer, and development. Oncogene 1999, 18, 7908–7916. [Google Scholar] [CrossRef] [Green Version]
- Alves, N.E.G.; de Mejía, E.G.; Vasconcelos, C.M.; Bassinello, P.Z.; Martino, H.S.D. Postharvest storage of Carioca bean (Phaseolus vulgaris L.) did not impair inhibition of inflammation in lipopolysaccharide-induced human THP-1 macrophage-like cells. J. Funct. Foods 2016, 23, 154–166. [Google Scholar] [CrossRef]
- Monsalve, F.A.; Pyarasani, R.D.; Delgado-Lopez, F.; Moore-Carrasco, R. Peroxisome proliferator-activated receptor targets for the treatment of metabolic diseases. Mediat. Inflamm. 2013, 2013, 549627. [Google Scholar] [CrossRef] [Green Version]
- de Lima, S.L.S.; Gomes, M.J.C.; da Silva, B.P.; Alves, N.E.G.; Toledo, R.C.L.; Theodoro, J.M.V.; Moreira, M.E.C.; Bento, J.A.C.; Bassinello, P.Z.; da Matta, S.L.P.; et al. Whole flour and protein hydrolysate from common beans reduce the inflammation in BALB/c mice fed with high fat high cholesterol diet. Food Res. Int. 2019, 122, 330–339. [Google Scholar] [CrossRef]
- Oseguera-Toledo, M.E.; De Mejia, E.G.; Dia, V.P.; Amaya-Llano, S.L. Common bean (Phaseolus vulgaris L.) hydrolysates inhibit inflammation in LPS-induced macrophages through suppression of NF-κB pathways. Food Chem. 2011, 127, 1175–1185. [Google Scholar] [CrossRef]
- Grancieri, M.; Martino, H.S.D.; de Mejia, E.G. Protein Digests and pure peptides from chia seed prevented adipogenesis and inflammation by inhibiting PPARγ and NF-κB pathways in 3T3L-1 adipocytes. Nutrients 2021, 13, 176. [Google Scholar] [CrossRef]
- Hwang, S.-J.; Song, Y.-S.; Lee, H.-J. Phaseolin attenuates lipopolysaccharide-induced inflammation in RAW 264.7 cells and zebrafish. Biomedicines 2021, 9, 420. [Google Scholar] [CrossRef]
- Kim, S.; Hong, J.; Jeon, R.; Kim, H.-S. Adzuki bean ameliorates hepatic lipogenesis and proinflammatory mediator expression in mice fed a high-cholesterol and high-fat diet to induce nonalcoholic fatty liver disease. Nutr. Res. 2016, 36, 90–100. [Google Scholar] [CrossRef]
- Rodríguez, E.; Ribot, J.; Rodríguez, A.M.; Palou, A. PPAR-γ2 expression in response to cafeteria diet: Gender- and depot-specific effects. Obes. Res. 2004, 12, 1455–1463. [Google Scholar] [CrossRef]
- Oomah, B.D.; Corbé, A.; Balasubramanian, P. Antioxidant and anti-inflammatory activities of bean (Phaseolus vulgaris L.) hulls. J. Agric. Food Chem. 2010, 58, 8225–8230. [Google Scholar] [CrossRef]
- Borresen, E.C.; Brown, D.G.; Harbison, G.; Taylor, L.; Fairbanks, A.; O’Malia, J.; Bazan, M.; Rao, S.; Bailey, S.M.; Wdowik, M.; et al. A randomized controlled trial to increase navy bean or rice bran consumption in colorectal cancer survivors. Nutr. Cancer 2016, 68, 1269–1280. [Google Scholar] [CrossRef] [Green Version]
- Kunnumakkara, A.B.; Sailo, B.L.; Banik, K.; Harsha, C.; Prasad, S.; Gupta, S.C.; Bharti, A.C.; Aggarwal, B.B. Chronic diseases, inflammation, and spices: How are they linked? J. Transl. Med. 2018, 16, 14. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Zou, Y.; Shen, Z.; Xiong, Y.; Zhang, W.; Liu, C.; Chen, S. Trace Elements, PPARs, and Metabolic Syndrome. Int. J. Mol. Sci. 2020, 21, 2612. [Google Scholar] [CrossRef] [Green Version]
- Katsimardou, A.; Imprialos, K.; Stavropoulos, K.; Sachinidis, A.; Doumas, M.; Athyros, V. Hypertension in metabolic syndrome: Novel Insights. Curr. Hypertens. Rev. 2020, 16, 12–18. [Google Scholar] [CrossRef]
- Castro-Barquero, S.; Ruiz-Leon, A.M.; Sierra-Perez, M.; Estruch, R.; Casas, R. Dietary strategies for metabolic syndrome: A comprehensive review. Nutrients 2020, 12, 2983. [Google Scholar] [CrossRef]
- Perez-Martinez, P.; Mikhailidis, D.P.; Athyros, V.G.; Bullo, M.; Couture, P.; Covas, M.I.; de Koning, L.; Delgado-Lista, J.; Díaz-López, A.; Drevon, C.A.; et al. Lifestyle recommendations for the prevention and management of metabolic syndrome: An international panel recommendation. Nutr. Rev. 2017, 75, 307–326. [Google Scholar] [CrossRef] [Green Version]
- Jakubczyk, A.; Karas, M.; Zlotek, U.; Szymanowska, U. Identification of potential inhibitory peptides of enzymes involved in the metabolic syndrome obtained by simulated gastrointestinal digestion of fermented bean (Phaseolus vulgaris L.) seeds. Food Res. Int. 2017, 100, 489–496. [Google Scholar] [CrossRef]
- Barrett, M.L.; Udani, J.K. A proprietary alpha-amylase inhibitor from white bean (Phaseolus vulgaris): A review of clinical studies on weight loss and glycemic control. Nutr. J. 2011, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Sales, P.M.; Souza, P.M.; Simeoni, L.A.; Magalhães, P.O.; Silveira, D. α-Amylase inhibitors: A review of raw material and isolated compounds from plant source. J. Pharm. Pharm. Sci. 2012, 15, 141–183. [Google Scholar] [CrossRef] [Green Version]
- Payab, M.; Hasani-Ranjbar, S.; Shahbal, N.; Qorbani, M.; Aletaha, A.; Haghi-Aminjan, H.; Soltani, A.; Khatami, F.; Nikfar, S.; Hassani, S.; et al. Effect of the herbal medicines in obesity and metabolic syndrome: A systematic review and meta-analysis of clinical trials. Phytother. Res. 2020, 34, 526–545. [Google Scholar] [CrossRef]
- Celleno, L.; Tolaini, M.V.; D’Amore, A.; Perricone, N.V.; Preuss, H.G. A dietary supplement containing standardized Phaseolus vulgaris extract influences body composition of overweight men and women. Int. J. Med. Sci. 2007, 4, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Shutler, S.M.; Bircher, G.M.; Tredger, J.A.; Morgan, L.M.; Walker, A.F.; Low, A.G. The effect of daily baked bean (Phaseolus vulgaris) consumption on the plasma lipid levels of young, normo-cholesterolaemic men. Br. J. Nutr. 1989, 61, 257–265. [Google Scholar] [CrossRef]
- Anderson, J.W.; Gustafson, N.J.; Spencer, D.B.; Tietyen, J.; Bryant, C.A. Serum lipid response of hypercholesterolemic men to single and divided doses of canned beans. Am. J. Clin. Nutr. 1990, 51, 1013–1019. [Google Scholar] [CrossRef]
- Reverri, E.J.; Randolph, J.M.; Steinberg, F.M.; Kappagoda, C.T.; Edirisinghe, I.; Burton-Freeman, B.M. black beans, fiber, and antioxidant capacity pilot study: Examination of whole foods vs. functional components on postprandial metabolic, oxidative stress, and inflammation in adults with metabolic syndrome. Nutrients 2015, 7, 6139–6154. [Google Scholar] [CrossRef]
- Spadafranca, A.; Rinelli, S.; Riva, A.; Morazzoni, P.; Magni, P.; Bertoli, S.; Battezzati, A. Phaseolus vulgaris extract affects glycometabolic and appetite control in healthy human subjects. Br. J. Nutr. 2013, 109, 1789–1795. [Google Scholar] [CrossRef] [Green Version]
- Reverri, E.J.; Randolph, J.M.; Kappagoda, C.T.; Park, E.; Edirisinghe, I.; Burton-Freeman, B.M. Assessing beans as a source of intrinsic fiber on satiety in men and women with metabolic syndrome. Appetite 2017, 118, 75–81. [Google Scholar] [CrossRef]
- Micheli, L.; Lucarini, E.; Trallori, E.; Avagliano, C.; De Caro, C.; Russo, R.; Calignano, A.; Ghelardini, C.; Pacini, A.; Di Cesare Mannelli, L. Phaseolus vulgaris L. extract: Alpha-amylase inhibition against metabolic syndrome in mice. Nutrients 2019, 11, 1778. [Google Scholar] [CrossRef] [Green Version]
- Zaru, A.; Maccioni, P.; Riva, A.; Morazzoni, P.; Bombardelli, E.; Gessa, G.L.; Morazzoni, P.; Carai, M.A.M. Reducing effect of a combination of Phaseolus vulgaris and Cynara scolymus extracts on operant self-administration of a chocolate-flavoured beverage in rats. Phytother. Res. 2013, 27, 944–947. [Google Scholar] [CrossRef]
- Li, G.-H.; Shi, Y.-H.; Liu, H.; Le, G.-W. Antihypertensive effect of alcalase generated mung bean protein hydrolysates in spontaneously hypertensive rats. Eur. Food Res. Technol. 2006, 222, 733–736. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, Y.; Zhang, Y.; Ruan, X.; Zhang, R. Purification, characterization, synthesis, in vitro ACE inhibition and in vivo antihypertensive activity of bioactive peptides derived from oil palm kernel glutelin-2 hydrolysates. J. Funct. Foods 2017, 28, 48–58. [Google Scholar] [CrossRef]
- Mojica, L.; de Mejia, E.G.; Granados-Silvestre, M.Á.; Menjivar, M. Evaluation of the hypoglycemic potential of a black bean hydrolyzed protein isolate and its pure peptides using in silico, in vitro and in vivo approaches. J. Funct. Foods 2017, 31, 274–286. [Google Scholar] [CrossRef]
- Nichenametla, S.N.; Weidauer, L.A.; Wey, H.E.; Beare, T.M.; Specker, B.L.; Dey, M. Resistant starch type 4-enriched diet lowered blood cholesterols and improved body composition in a double blind controlled cross-over intervention. Mol. Nutr. Food Res. 2014, 58, 1365–1369. [Google Scholar] [CrossRef]
- Park, O.J.; Ekang, N.; Chang, M.J.; Kim, W.K. Resistant starch supplementation influences blood lipid concentrations and glucose control in overweight subjects. J. Nutr. Sci. Vitaminol. 2004, 50, 93–99. [Google Scholar] [CrossRef] [Green Version]
- den Hartigh, L.J. Conjugated linoleic acid effects on cancer, obesity, and atherosclerosis: A review of pre-clinical and human trials with current perspectives. Nutrients 2019, 11, 370. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Albright, K.J.; Liu, W.; Storkson, J.M.; Cook, M.E.; Pariza, M.W. Effect of conjugated linoleic acid on body composition in mice. Lipids 1997, 32, 853–858. [Google Scholar] [CrossRef]
- Miller, J.R.; Siripurkpong, P.; Hawes, J.; Majdalawieh, A.; Ro, H.-S.; McLeod, R.S. The trans-10, cis-12 isomer of conjugated linoleic acid decreases adiponectin assembly by PPARγ-dependent and PPARγ-independent mechanisms. J. Lipid Res. 2008, 49, 550–562. [Google Scholar] [CrossRef] [Green Version]
- Carai, M.A.; Fantini, N.; Loi, B.; Colombo, G.; Riva, A.; Morazzoni, P. Potential efficacy of preparations derived from Phaseolus vulgaris in the control of appetite, energy intake, and carbohydrate metabolism. Diabetes Metab. Syndr. Obes. Targets Ther. 2009, 2, 145–153. [Google Scholar] [CrossRef]
- Guo, W.; Kim, S.H.; Wu, D.; Li, L.; Ortega, E.F.; Thomas, M.; Meydani, S.N.; Meydani, M. Dietary fruit and vegetable supplementation suppresses diet-induced atherosclerosis in LDL receptor knockout mice. J. Nutr. 2021, 151, 902–910. [Google Scholar] [CrossRef]
- Khan, R.; Spagnoli, V.; Tardif, J.-C.; L’Allier, P.L. Novel anti-inflammatory therapies for the treatment of atherosclerosis. Atherosclerosis 2015, 240, 497–509. [Google Scholar] [CrossRef]
- Alves, N.E.G.; Vasconcelos, C.M.; Bassinello, P.Z.; de Mejia, E.G.; Martino, H.S.D. Digested protein isolate from fresh and stored Carioca beans reduced markers of atherosclerosis in oxidized LDL-induced THP-1 macrophages. J. Funct. Foods 2016, 24, 97–111. [Google Scholar] [CrossRef]
- Neele, A.E.; Van den Bossche, J.; Hoeksema, M.A.; De Winther, M.P.J. Epigenetic pathways in macrophages emerge as novel targets in atherosclerosis. Eur. J. Pharmacol. 2015, 763, 79–89. [Google Scholar] [CrossRef]
- Soltani, S.; Boozari, M.; Cicero, A.F.G.; Jamialahmadi, T.; Sahebkar, A. Effects of phytochemicals on macrophage cholesterol efflux capacity: Impact on atherosclerosis. Phytother. Res. 2021, 35, 2854–2878. [Google Scholar] [CrossRef]
- Finley, J.W.; Burrell, J.B.; Reeves, P.G. Pinto bean consumption changes SCFA profiles in fecal fermentations, bacterial populations of the lower bowel, and lipid profiles in blood of humans. J. Nutr. 2007, 137, 2391–2398. [Google Scholar] [CrossRef]
- Taga, M.S.; Miller, E.E.; Pratt, D.E. Chia seeds as a source of natural lipid antioxidants. J. Am. Oil Chem. Soc. 1984, 61, 928–931. [Google Scholar] [CrossRef]
- Ayerza, R. The seed’s oil content and fatty acid composition of chia (Salvia hispanica L.) Var. Iztac 1, grown under six tropical ecosystems conditions. Interciencia 2011, 36, 620–624. [Google Scholar]
- Ciftci, O.N.; Przybylski, R.; Rudzińska, M. Lipid components of flax, perilla, and chia seeds. Eur. J. Lipid Sci. Technol. 2012, 114, 794–800. [Google Scholar] [CrossRef]
- Carrero, J.J.; Martín-Bautista, E.; Baró, L.; Fonollá, J.; Jiménez, J.; Boza, J.J.; López-Huertas, E. Efectos cardiovasculares de los ácidos grasos omega-3 y alternativas para incrementar su ingesta. Nutr. Hosp. 2005, 20, 63–69. [Google Scholar]
- Kritchevsky, D. Conjugated linoleic acid in experimental atherosclerosis. In Advances in Conjugated Linoleic Acid Research; AOCS Publishing: New York, NY, USA, 2020; Volume 2, pp. 292–301. [Google Scholar]
- Kritchevsky, D.; Tepper, S.A.; Wright, S.; Czarnecki, S.K.; Wilson, T.A.; Nicolosi, R.J. Conjugated linoleic acid isomer effects in atherosclerosis: Growth and regression of lesions. Lipids 2004, 39, 611–616. [Google Scholar] [CrossRef]
- Wilson, T.A.; Nicolosi, R.J.; Chrysam, M.; Kritchevsky, D. Conjugated linoleic acid reduces early aortic atherosclerosis greater than linoleic acid in hypercholesterolemic hamsters. Nutr. Res. 2000, 20, 1795–1805. [Google Scholar] [CrossRef]
- Glore, S.R.; Van Treeck, D.; Knehans, A.W.; Guild, M. Soluble fiber and serum lipids: A literature review. J. Am. Diet. Assoc. 1994, 94, 425–436. [Google Scholar] [CrossRef]
- Kritchevsky, D. Protein and atherosclerosis. J. Nutr. Sci. Vitaminol. 1990, 36, S81–S86. [Google Scholar] [CrossRef]
- Navab, M.; Anantharamaiah, G.; Hama, S.; Garber, D.W.; Chaddha, M.; Hough, G.; Lallone, R.; Fogelman, A.M. Oral administration of an Apo AI mimetic peptide synthesized from D-amino acids dramatically reduces atherosclerosis in mice independent of plasma cholesterol. Circulation 2002, 105, 290–292. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wu, D.; Matthan, N.R.; Lamon-Fava, S.; Lecker, J.L.; Lichtenstein, A.H. Reduction in dietary omega-6 polyunsaturated fatty acids: Eicosapentaenoic acid plus docosahexaenoic acid ratio minimizes atherosclerotic lesion formation and inflammatory response in the LDL receptor null mouse. Atherosclerosis 2009, 204, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Chavez-Mendoza, C.; Sanchez, E. Bioactive Compounds from Mexican varieties of the common bean (Phaseolus vulgaris): Implications for Health. Molecules 2017, 22, 1360. [Google Scholar] [CrossRef] [Green Version]
- Frassinetti, S.; Gabriele, M.; Caltavuturo, L.; Longo, V.; Pucci, L. Antimutagenic and antioxidant activity of a selected lectin-free common bean (Phaseolus vulgaris L.) in two cell-based models. Plant Foods Hum. Nutr. 2015, 70, 35–41. [Google Scholar] [CrossRef]
- Giugliano, D.; Ceriello, A.; Esposito, K. The effects of diet on inflammation: Emphasis on the metabolic syndrome. J. Am. Coll. Cardiol. 2006, 48, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Madrera, R.R.; Negrillo, A.C.; Valles, B.S.; Fernandez, J.J.F. Characterization of extractable phenolic profile of common bean seeds (Phaseolus vulgaris L.) in a Spanish diversity panel. Food Res. Int. 2020, 138, 109713. [Google Scholar] [CrossRef]
- Perez-Hernandez, L.M.; Nugraheni, K.; Benohoud, M.; Sun, W.; Hernández-Álvarez, A.J.; Morgan, M.R.A.; Boesch, C.; Orfila, C. Starch Digestion Enhances Bioaccessibility of Anti-Inflammatory Polyphenols from Borlotti Beans (Phaseolus vulgaris). Nutrients 2020, 12, 295. [Google Scholar] [CrossRef] [Green Version]
- Hutchins, A.M.; Winham, D.M.; Thompson, S.V. Phaseolus beans: Impact on glycaemic response and chronic disease risk in human subjects. Br. J. Nutr. 2012, 108, S52–S65. [Google Scholar] [CrossRef] [Green Version]
- Chavez-Santoscoy, R.A.; Gutierrez-Uribe, J.A.; Serna-Saldivar, S.O. Effect of flavonoids and saponins extracted from black bean (Phaseolus vulgaris L.) seed coats as cholesterol micelle disruptors. Plant Foods Hum. Nutr. 2013, 68, 416–423. [Google Scholar] [CrossRef]
- Clark, J.L.; Taylor, C.G.; Zahradka, P. Black beans and red kidney beans induce positive postprandial vascular responses in healthy adults: A pilot randomized cross-over study. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 216–226. [Google Scholar] [CrossRef]
- Graziani, D.; Ribeiro, J.; Cruz, V.; Gomes, R.; Araújo, E.; Júnior, A.S.; Tomaz, H.; Castro, C.; Fontes, W.; Batista, K.; et al. Oxidonitrergic and antioxidant effects of a low molecular weight peptide fraction from hardened bean (Phaseolus vulgaris) on endothelium. Braz. J. Med Biol. Res. 2021, 54, e10423. [Google Scholar] [CrossRef]
- Zhu, Z.; Jiang, W.; Thompson, H.J. Edible dry bean consumption (Phaseolus vulgaris L.) modulates cardiovascular risk factors and diet-induced obesity in rats and mice. Br. J. Nutr. 2012, 108, S66–S73. [Google Scholar] [CrossRef] [Green Version]
- Rubio, L.A.; Aranda-Olmedo, I.; Martin-Pedrosa, M. Inclusion of limited amounts of extruded legumes plus cereal mixes in normocaloric or obesogenic diets for rats: Effects on lipid profile. Foods 2020, 9, 704. [Google Scholar] [CrossRef]
- Howard, B.V.; Kritchevsky, D. Phytochemicals and cardiovascular disease: A statement for healthcare professionals from the American Heart Association. Circulation 1997, 95, 2591–2593. [Google Scholar] [CrossRef]
- Anderson, J.W.; Smith, B.M.; Washnock, C.S. Cardiovascular and renal benefits of dry bean and soya bean intake. Oilseeds Focus 2020, 6, 43–45. [Google Scholar]
- Van Horn, L. Fiber, lipids, and coronary heart disease: A statement for healthcare professionals from the Nutrition Committee, American Heart Association. Circulation 1997, 95, 2701–2704. [Google Scholar] [CrossRef]
- Lavie, C.J.; Milani, R.V.; Mehra, M.R.; Ventura, H.O. Omega-3 polyunsaturated fatty acids and cardiovascular diseases. J. Am. Coll. Cardiol. 2009, 54, 585–594. [Google Scholar] [CrossRef] [Green Version]
- Anand, R.G.; Alkadri, M.; Lavie, C.J.; Milani, R.V. The role of fish oil in arrhythmia prevention. J. Cardiopulm. Rehabil. Prev. 2008, 28, 92–98. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Bryson, C.L.; Lemaitre, R.N.; Burke, G.L.; Siscovick, D.S. Fish intake and risk of incident heart failure. J. Am. Coll. Cardiol. 2005, 45, 2015–2021. [Google Scholar] [CrossRef] [Green Version]
- Yamagishi, K.; Iso, H.; Date, C.; Fukui, M.; Wakai, K.; Kikuchi, S.; Inaba, Y.; Tanabe, N.; Tamakoshi, A. Fish, ω-3 polyunsaturated fatty acids, and mortality from cardiovascular diseases in a nationwide community-based cohort of Japanese men and women: The JACC (Japan Collaborative Cohort Study for Evaluation of Cancer Risk) study. J. Am. Coll. Cardiol. 2008, 52, 988–996. [Google Scholar] [CrossRef] [Green Version]
- Galan, P.; Kesse-Guyot, E.; Czernichow, S.; Briancon, S.; Blacher, J.; Hercberg, S. Effects of B vitamins and omega 3 fatty acids on cardiovascular diseases: A randomised placebo controlled trial. BMJ 2010, 341, c6273. [Google Scholar] [CrossRef] [Green Version]
- Luceri, C.; Giannini, L.; Lodovici, M.; Antonucci, E.; Abbate, R.; Masini, E.; Dolara, P. p-Coumaric acid, a common dietary phenol, inhibits platelet activity in vitro and in vivo. Br. J. Nutr. 2007, 97, 458–463. [Google Scholar] [CrossRef] [Green Version]
- Prince, P.S.M.; Roy, A.J. p-Coumaric acid attenuates apoptosis in isoproterenol-induced myocardial infarcted rats by inhibiting oxidative stress. Int. J. Cardiol. 2013, 168, 3259–3266. [Google Scholar] [CrossRef]
- Ostertag, L.M.; O’Kennedy, N.; Horgan, G.W.; Kroon, P.A.; Duthie, G.G.; de Roos, B. In vitro anti-platelet effects of simple plant-derived phenolic compounds are only found at high, non-physiological concentrations. Mol. Nutr. Food Res. 2011, 55, 1624–1636. [Google Scholar] [CrossRef]
- Zang, L.Y.; Cosma, G.; Gardner, H.; Shi, X.; Castranova, V.; Vallyathan, V. Effect of antioxidant protection by p-coumaric acid on low-density lipoprotein cholesterol oxidation. Am. J. Physiol.-Cell Physiol. 2000, 279, C954–C960. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Wahab, M.H.; El-Mahdy, M.A.; Abd-Ellah, M.F.; Helal, G.K.; Khalifa, F.; Hamada, F.M. Influence of p-coumaric acid on doxorubicin-induced oxidative stress in rat’s heart. Pharmacol. Res. 2003, 48, 61–65. [Google Scholar] [CrossRef]
- Fuentes, E.; Caballero, J.; Alarcon, M.; Rojas, A.; Palomo, I. Chlorogenic acid inhibits human platelet activation and thrombus formation. PLoS ONE 2014, 9, e90699. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-J.; Kang, H.-J.; Kim, Y.-J.; Lee, D.-H.; Kwon, H.-W.; Kim, Y.-Y.; Park, H.-J. Inhibition of platelet aggregation by chlorogenic acid via cAMP and cGMP-dependent manner. Blood Coagul. Fibrinolysis 2012, 23, 629–635. [Google Scholar] [CrossRef]
- Fuentes, E.; Forero-Doria, O.; Carrasco, G.; Maricán, A.; Santos, L.S.; Alarcón, M.; Palomo, I. Effect of tomato industrial processing on phenolic profile and antiplatelet activity. Molecules 2013, 18, 11526–11536. [Google Scholar] [CrossRef] [Green Version]
- Hong, Q.; Ma, Z.-C.; Huang, H.; Wang, Y.-G.; Tan, H.-L.; Xiao, C.-R.; Liang, Q.-D.; Zhang, H.-T.; Gao, Y. Antithrombotic activities of ferulic acid via intracellular cyclic nucleotide signaling. Eur. J. Pharmacol. 2016, 777, 1–8. [Google Scholar] [CrossRef]
- Kim, M.-S.; Shin, W.-C.; Kang, D.-K.; Sohn, H.-Y. Anti-thrombosis activity of sinapic acid isolated from the lees of bokbunja wine. J. Microbiol. Biotechnol. 2016, 26, 61–65. [Google Scholar] [CrossRef]
- Kim, K.; Bae, O.-N.; Lim, K.-M.; Noh, J.-Y.; Kang, S.; Chung, K.Y.; Chung, J.-H. Novel antiplatelet activity of protocatechuic acid through the inhibition of high shear stress-induced platelet aggregation. J. Pharmacol. Exp. Ther. 2012, 343, 704–711. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.-L.; Liu, J.-X.; Dong, W.; Li, P.; Li, L.; Lin, C.-R.; Zheng, Y.-Q.; Cong, W.-H.; Hou, J.-C. The cardioprotective effect of protocatechuic acid on myocardial ischemia/reperfusion injury. J. Pharmacol. Sci. 2014, 125, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Ya, F.; Li, K.; Chen, H.; Tian, Z.; Fan, D.; Shi, Y.; Song, F.; Xu, X.; Ling, W.; Adili, R.; et al. Protocatechuic acid protects platelets from apoptosis via inhibiting oxidative stress-mediated PI3K/Akt/GSK3β signaling. Thromb. Haemost. 2021, 121, 931–943. [Google Scholar] [CrossRef]
- Choi, J.-H.; Kim, S. Mechanisms of attenuation of clot formation and acute thromboembolism by syringic acid in mice. J. Funct. Foods 2018, 43, 112–122. [Google Scholar] [CrossRef]
- Gaspar, R.S.; da Silva, S.A.; Stapleton, J.; Fontelles, J.L.L.; Sousa, H.R.; Chagas, V.T.; Alsufyani, S.; Trostchansky, A.; Gibbins, J.M.; de Andrade Paes, A.M. Myricetin, the main flavonoid in Syzygium cumini leaf, is a novel inhibitor of platelet thiol isomerases PDI and ERp5. Front. Pharmacol. 2020, 10, 1678. [Google Scholar] [CrossRef]
- Zang, B.X.; Jin, M.; Wu, W.; Chen, W.M.; Piao, Y.Z.; Li, J.R. Antagonistic effect of myricetin on platelet activing factor. Yao Xue Xue Bao 2003, 38, 831–833. [Google Scholar]
- Robak, J.; Korbut, R.; Shridi, F.; Swies, J.; Rzadkowska-Bodalska, H. On the mechanism of antiaggregatory effect of myricetin. Pol. J. Pharmacol. Pharm. 1988, 40, 337–340. [Google Scholar]
- Landolfi, R.; Mower, R.L.; Steiner, M. Modification of platelet function and arachidonic acid metabolism by bioflavonoids: Structure-activity relations. Biochem. Pharmacol. 1984, 33, 1525–1530. [Google Scholar] [CrossRef]
- Nakashima, S.; Koike, T.; Nozawa, Y. Genistein, a protein tyrosine kinase inhibitor, inhibits thromboxane A2-mediated human platelet responses. Mol. Pharmacol. 1991, 39, 475–480. [Google Scholar]
- Gottstein, N.; Ewins, B.A.; Eccleston, C.; Hubbard, G.P.; Kavanagh, I.C.; Minihane, A.-M.; Weinberg, P.D.; Rimbach, G. Effect of genistein and daidzein on platelet aggregation and monocyte and endothelial function. Br. J. Nutr. 2003, 89, 607–615. [Google Scholar] [CrossRef]
- Ozaki, Y.; Yatomi, Y.; Jinnai, Y.; Kume, S. Effects of genistein, a tyrosine kinase inhibitor, on platelet functions: Genistein attenuates thrombin-induced Ca2+ mobilization in human platelets by affecting polyphosphoinositide turnover. Biochem. Pharmacol. 1993, 46, 395–403. [Google Scholar] [CrossRef]
- Dhar, A.; Paul, A.K.; Shukla, S.D. Platelet-activating factor stimulation of tyrosine kinase and its relationship to phospholipase C in rabbit platelets: Studies with genistein and monoclonal antibody to phosphotyrosine. Mol. Pharmacol. 1990, 37, 519–525. [Google Scholar]
- Pan, W.; Ikeda, K.; Takebe, M.; Yamori, Y. Genistein, daidzein and glycitein inhibit growth and DNA synthesis of aortic smooth muscle cells from stroke-prone spontaneously hypertensive rats. J. Nutr. 2001, 131, 1154–1158. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.J.; Zhuang, P.W.; Zhang, Y.J. Antithrombotic activity of Formononetin sodium and its mechanism. Chin. J. New Drugs 2016, 25, 1355–1358. [Google Scholar]
- Chen, Z.; Liu, S.; Cai, Y.; Xie, K.; Zhang, W.; Dong, L.; Liu, Y.; Zheng, F.; Dun, Y.; Li, N. Suppressive effect of formononetin on platelet-derived growth factor-BB-stimulated proliferation and migration of vascular smooth muscle cells. Exp. Ther. Med. 2016, 12, 1901–1907. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Zou, L.; Tian, J.; Lin, F.; He, J.; Hou, J. Protective effects of sulphonated formononetin in a rat model of cerebral ischemia and reperfusion injury. Planta Medica 2014, 80, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Chtourou, Y.; Kamoun, Z.; Zarrouk, W.; Kebieche, M.; Kallel, C.; Gdoura, R.; Fetoui, H. Naringenin ameliorates renal and platelet purinergic signalling alterations in high-cholesterol fed rats through the suppression of ROS and NF-κB signaling pathways. Food Funct. 2016, 7, 183–193. [Google Scholar] [CrossRef]
- Jin, Y.-R.; Han, X.-H.; Zhang, Y.-H.; Lee, J.-J.; Lim, Y.; Chung, J.-H.; Yun, Y.-P. Antiplatelet activity of hesperetin, a bioflavonoid, is mainly mediated by inhibition of PLC-γ2 phosphorylation and cyclooxygenase-1 activity. Atherosclerosis 2007, 194, 144–152. [Google Scholar] [CrossRef]
- McGregor, L.; Bellangeon, M.; Chignier, E.; Lerond, L.; Rousselle, C.; McGregor, J.L. Effect of a micronized purified flavonoid fraction on in vivo platelet functions in the rat. Thromb. Res. 1999, 94, 235–240. [Google Scholar] [CrossRef]
- Pignatelli, P.; Pulcinelli, F.M.; Celestini, A.; Lenti, L.; Ghiselli, A.; Gazzaniga, P.P.; Violi, F. The flavonoids quercetin and catechin synergistically inhibit platelet function by antagonizing the intracellular production of hydrogen peroxide. Am. J. Clin. Nutr. 2000, 72, 1150–1155. [Google Scholar] [CrossRef] [Green Version]
- Neiva, T.J.C.; Morais, L.; Polack, M.; Simoes, C.M.O.; D’Amico, E.A. Effects of catechins on human blood platelet aggregation and lipid peroxidation. Phytother. Res. 1999, 13, 597–600. [Google Scholar] [CrossRef]
- Blache, D.; Durand, P.; Prost, M.; Loreau, N. (+)-Catechin inhibits platelet hyperactivity induced by an acute iron load in vivo. Free. Radic. Biol. Med. 2002, 33, 1670–1680. [Google Scholar] [CrossRef]
- Tzeng, S.-H.; Ko, W.-C.; Ko, F.-N.; Teng, C.-M. Inhibition of platelet aggregation by some flavonoids. Thromb. Res. 1991, 64, 91–100. [Google Scholar] [CrossRef]
- Chung, M.-I.; Gan, K.-H.; Lin, C.-N.; Ko, F.-N.; Teng, C.-M. Antiplatelet effects and vasorelaxing action of some constituents of Formosan plants. J. Nat. Prod. 1993, 56, 929–934. [Google Scholar] [CrossRef]
- Kim, T.-J.; Kim, J.-H.; Jin, Y.-R.; Yun, Y.-P. The inhibitory effect and mechanism of luteolin 7-glucoside on rat aortic vascular smooth muscle cell proliferation. Arch. Pharmacal Res. 2006, 29, 67–72. [Google Scholar] [CrossRef]
- Teng, C.-M.; Li, H.-L.; Wu, T.-S.; Huang, S.-C.; Huang, T.-F. Antiplatelet actions of some coumarin compounds isolated from plant sources. Thromb. Res. 1992, 66, 549–557. [Google Scholar] [CrossRef]
- Zaragozá, C.; Monserrat, J.; Mantecón, C.; Villaescusa, L.; Zaragozá, F.; Álvarez-Mon, M. Antiplatelet activity of flavonoid and coumarin drugs. Vasc. Pharmacol. 2016, 87, 139–149. [Google Scholar] [CrossRef]
- Macáková, K.; Řeháková, Z.; Mladěnka, P.; Karlíčková, J.; Filipský, T.; Říha, M.; Prasad, A.K.; Parmar, V.S.; Jahodář, L.; Pávek, P.; et al. In vitro platelet antiaggregatory properties of 4-methylcoumarins. Biochimie 2012, 94, 2681–2686. [Google Scholar] [CrossRef]
- Deusch, E.; Gamsjäger, T.; Kress, H.-G.; Kozek-Langenecker, S.A. Binding of hydroxyethyl starch molecules to the platelet surface. Anesthesia Analg. 2003, 97, 680–683. [Google Scholar] [CrossRef]
- Freedman, J.E.; Farhat, J.H.; Loscalzo, J.; Keaney, J.F. α-tocopherol inhibits aggregation of human platelets by a protein kinase C-dependent mechanism. Circulation 1996, 94, 2434–2440. [Google Scholar] [CrossRef]
- Liu, M.; Wallmon, A.; Olsson-Mortlock, C.; Wallin, R.; Saldeen, T. Mixed tocopherols inhibit platelet aggregation in humans: Potential mechanisms. Am. J. Clin. Nutr. 2003, 77, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Gajos, G.; Rostoff, P.; Undas, A.; Piwowarska, W. Effects of polyunsaturated omega-3 fatty acids on responsiveness to dual antiplatelet therapy in patients undergoing percutaneous coronary intervention: The OMEGA-PCI (OMEGA-3 fatty acids after pci to modify responsiveness to dual antiplatelet therapy) study. J. Am. Coll. Cardiol. 2010, 55, 1671–1678. [Google Scholar]
- Gajos, G.; Zalewski, J.; Rostoff, P.; Nessler, J.; Piwowarska, W.; Undas, A. Reduced thrombin formation and altered fibrin clot properties induced by polyunsaturated omega-3 fatty acids on top of dual antiplatelet therapy in patients undergoing percutaneous coronary intervention (OMEGA-PCI clot). Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1696–1702. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.G.; Rossi, J.S.; Garbarino, J.; Bowling, R.; Motsinger-Reif, A.A.; Schuler, C.; Dupont, A.G.; Gabriel, D. Insights into the inhibition of platelet activation by omega-3 polyunsaturated fatty acids: Beyond aspirin and clopidogrel. Thromb. Res. 2011, 128, 335–340. [Google Scholar] [CrossRef]
- Song, F.; Zhu, Y.; Shi, Z.; Tian, J.; Deng, X.; Ren, J.; Andrews, M.C.; Ni, H.; Ling, W.; Yang, Y. Plant food anthocyanins inhibit platelet granule secretion in hypercholesterolaemia: Involving the signalling pathway of PI3K–Akt. Thromb. Haemost. 2014, 111, 981–991. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, Z.; Reheman, A.; Jin, J.W.; Li, C.; Wang, Y.; Andrews, M.C.; Chen, P.; Zhu, G.; Ling, W.; et al. Plant food delphinidin-3-glucoside significantly inhibits platelet activation and thrombosis: Novel protective roles against cardiovascular diseases. PLoS ONE 2012, 7, e37323. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Chen, Y.; Adili, R.; McKeown, T.; Chen, P.; Zhu, G.; Li, D.; Ling, W.; Ni, H.; Yang, Y. Plant-based food cyanidin-3-glucoside modulates human platelet glycoprotein VI signaling and inhibits platelet activation and thrombus formation. J. Nutr. 2017, 147, 1917–1925. [Google Scholar] [CrossRef] [Green Version]
- Ku, S.-K.; Yoon, E.-K.; Lee, W.; Kwon, S.; Lee, T.; Bae, J.-S. Antithrombotic and antiplatelet activities of pelargonidin in vivo and in vitro. Arch. Pharmacal Res. 2016, 39, 398–408. [Google Scholar] [CrossRef]
- Ji, H.; Delker, S.L.; Li, H.; Martásek, P.; Roman, L.J.; Poulos, T.L.; Silverman, R.B. Exploration of the active site of neuronal nitric oxide synthase by the design and synthesis of pyrrolidinomethyl 2-aminopyridine derivatives. J. Med. Chem. 2010, 53, 7804–7824. [Google Scholar] [CrossRef] [Green Version]
- Hopper, P.C.W. Virtual Screening and Molecular Docking Studies of Compounds from Phaseolus vulgaris against Nitric Oxide Synthase. In Proceedings of the International Conference on Bioscience, Biochemistry and Bioinformatics (CBBB 2011), Singapore, 26–28 February 2011. [Google Scholar]
- Ngoh, Y.-Y.; Lim, T.S.; Gan, C.-Y. Screening and identification of five peptides from pinto bean with inhibitory activities against α-amylase using phage display technique. Enzym. Microb. Technol. 2016, 89, 76–84. [Google Scholar] [CrossRef]
- Hermoso, J.; Pignol, D.; Kerfelec, B.; Crenon, I.; Chapus, C.; Fontecilla-Camps, J.C. Lipase activation by nonionic detergents: The crystal structure of the porcine lipase-colipase-tetraethylene glycol monooctyl ether complex. J. Biol. Chem. 1996, 271, 18007–18016. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.B.; Khan, G.; Taylor, G.; Tickle, I.J.; Blundell, T.L. X-ray analyses of aspartic proteinases: II. Three-dimensional structure of the hexagonal crystal form of porcine pepsin at 2.3 A resolution. J. Mol. Biol. 1990, 214, 199–222. [Google Scholar] [CrossRef]
- Lebon, G.; Warne, T.; Edwards, P.C.; Bennett, K.; Langmead, C.J.; Leslie, A.G.W.; Tate, C.G. Agonist-bound adenosine A 2A receptor structures reveal common features of GPCR activation. Nature 2011, 474, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, Z.-G.; Zhang, K.; Kiselev, E.; Crane, S.; Wang, J.; Paoletta, S.; Yi, C.; Ma, L.; Zhang, W.; et al. Two disparate ligand-binding sites in the human P2Y1 receptor. Nature 2015, 520, 317–321. [Google Scholar] [CrossRef]
- Fan, H.; Chen, S.; Yuan, X.; Han, S.; Zhang, H.; Xia, W.; Xu, Y.; Zhao, Q.; Wu, B. Structural basis for ligand recognition of the human thromboxane A(2) receptor. Nat. Chem. Biol. 2019, 15, 27–33. [Google Scholar] [CrossRef]
- Grodsky, N.; Li, Y.; Bouzida, D.; Love, R.; Jensen, J.; Nodes, B.; Nonomiya, J.; Grant, S. Structure of the catalytic domain of human protein kinase C beta II complexed with a bisindolylmaleimide inhibitor. Biochemistry 2006, 45, 13970–13981. [Google Scholar] [CrossRef]
- Sutton, R.B.; Sprang, S.R. Structure of the protein kinase Cbeta phospholipid-binding C2 domain complexed with Ca2+. Structure 1998, 6, 1395–1405. [Google Scholar] [CrossRef] [Green Version]
- Newport, T.D.; Sansom, M.S.P.; Stansfeld, P.J. The MemProtMD database: A resource for membrane-embedded protein structures and their lipid interactions. Nucleic Acids Res. 2018, 47, D390–D397. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, E.; Badimon, L.; Caballero, J.; Padró, T.; Vilahur, G.; Alarcón, M.; Pérez, P.; Palomo, I. Protective mechanisms of adenosine 5′-monophosphate in platelet activation and thrombus formation. Thromb. Haemost. 2014, 111, 491–507. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, K.; Gao, Z.-G.; Paoletta, S.; Zhang, D.; Han, G.W.; Li, T.; Ma, L.; Zhang, W.; Müller, C.E.; et al. Agonist-bound structure of the human P2Y 12 receptor. Nature 2014, 509, 119–122. [Google Scholar] [CrossRef]
- Bugert, P.; Fischer, L.; Althaus, K.; Knöfler, R.; Bakchoul, T. Platelet dysfunction caused by a novel thromboxane A2 receptor mutation and congenital thrombocytopenia in a case of mild bleeding. Platelets 2020, 31, 276–279. [Google Scholar] [CrossRef]
- Bauer, J.; Ripperger, A.; Frantz, S.; Ergün, S.; Schwedhelm, E.; Benndorf, R.A. Pathophysiology of isoprostanes in the cardiovascular system: Implications of isoprostane-mediated thromboxane A 2 receptor activation. Br. J. Pharmacol. 2014, 171, 3115–3131. [Google Scholar] [CrossRef] [Green Version]
- Harper, M.T.; Poole, A.W. Diverse functions of protein kinase C isoforms in platelet activation and thrombus formation. J. Thromb. Haemost. 2010, 8, 454–462. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Classification | Mechanisms |
|---|---|---|
| Coumaric acid | Phenolic acid | Reduces ADP-induced platelet aggregation in vitro. Modifies platelet function, measured with PFA-100 (500 and 1 µM). Mechanism: inhibits the AA cascade, reducing TxA2 production and the generation of prostaglandin E2 induced by lipopolysaccharides (IC50: 371 and 126 μM) [189]. Inhibits ADP-induced platelet aggregation in vivo without affecting blood coagulation. Mechanism: associated with a marked increase in plasma antioxidant activity, measured as plasma iron-reducing capacity, and with a reduction in TxA2 production [189]. |
| Reduces apoptosis in vivo in rats with isoproterenol-induced myocardial infarction by inhibiting oxidative stress (2.4 and 8 mg/kg). Mechanism: protective effect due to anti-lipid peroxidative, antioxidant, and anti-apoptotic properties [190]. | ||
| Inhibits in vitro TRAP-induced surface P-selectin expression (100 μmol/L) [191]. | ||
| Inhibits in vivo LDL oxidation and decreases MDA production, causing a decrease in atherosclerosis (100 mg/kg) [192]. | ||
| Protects the heart in vivo against DOX-induced oxidative stress (100 mg/kg) [193]. | ||
| Chlorogenic acid | Phenolic acid | Inhibits in vitro platelet secretion and aggregation induced by ADP, collagen, AA, and TRAP-6. Decreases platelet adhesion, aggregation, and platelet–leukocyte interactions under flow conditions. Inhibits platelet inflammatory mediators (P-selectin, sCD40L, CCL5, and IL-1β) and increases intra-platelet cAMP levels by activation of PKA (0.1 to 1 mmol/L). Mechanism: adenylate cyclase/cAMP/PKA receptor signaling pathway [194]. |
| Inhibits collagen-induced platelet aggregation and suppresses TxA2 production, associated with COX-1 inhibition in platelet microsomes that have activity on cytochrome c reductase. Increases the formation of cAMP and cGMP and intracellular Ca2+ (10, 30, and 50 mmol/L). Mechanism: reduction of TxA2 and increased levels of cAMP and cGMP [195]. | ||
| Ferulic acid | Phenolic acid | Inhibits in vitro the expression of P-selectin on the surface induced by TRAP (100 μmol/L). Inhibits platelet aggregation induced with collagen 3.5 μg/mL (100 μmol/L) [191]. |
| Inhibits in vitro platelet aggregation stimulated with ADP 8 μmol/L and collagen 1.5 μg/mL (0.5 mmol/L) [196]. | ||
| Protects in vivo death produced by pulmonary thrombosis and prolongs tail bleeding time in mice and rats (20, 40, and 80 mg/kg for mice, or 10, 20, and 40 mg/kg for rats). Inhibits in vitro platelet aggregation induced by ADP, thrombin, collagen, and AA. It decreases the mobilization of intracellular Ca2+ and the production of TxA2. Increases levels of cAMP and cGMP and phosphoprotein stimulated by VASP while decreasing phospho-MAPK and PDE (200 mM). Mechanism: activation of cAMP and cGMP signaling [197]. | ||
| Synapic acid | Phenolic acid | Inhibits in vitro platelet aggregation and coagulation with antithrombotic effect (0.25 mg/mL, IC50: 1.03 mM) [198]. |
| Protocatechic acid | Phenolic acid | Decreases in vitro SIPA and attenuates platelet activation, intracellular Ca2+ mobilization, granule secretion, and adhesion receptor expression (10–25 µM). Mechanism: blockade of the binding of VWF to activated GPIb, primary and initial event for the performance of SIPA [199]. |
| Reduces myocardial infarction size, serum TNF-α levels, and platelet aggregation in vitro. Inhibits apoptosis and caspase-3 expression and positively regulates phosphorylated Akt expression in cardiomyocytes subjected to injury by H/R (250–500 mg/kg). Mechanism: provides protection against MI/R injury, inflammatory response, platelet aggregation, and apoptosis of cardiomyocytes [200]. | ||
| Inhibits platelet apoptosis induced by decreased dissipation of mitochondrial membrane potential, activation of caspase-9 and caspase-3, and decreased exposure to PS. Modulates the distributions of Bax, Bcl-xL, and cytochrome c mediated by H2O2 in mitochondria and cytochrome. It decreases ROS generation and intracellular Ca2+ concentration, caspase-3 signaling and activation, and PS exposure (0.5 or 1 µM). Mechanism: protects platelets from oxidative stress-induced apoptosis by regulating ROS-mediated PI3K/Akt/SK3β signaling [201]. | ||
| Syringic acid | Phenolic acid | Inhibits clotting factors, decreases the secretion of dense granules, and retraction of the clot. Regulates the downstream signaling pathway of DEP-1/PTP-1B/αIIbβ3/kinases. It decreases the expression of density-enhanced phosphatase-1 (DEP-1)/protein tyrosine phosphatase-1B (PTP1B)/αIIbβ3, as well as the phosphorylation of platelet kinases stimulated with collagen/epinephrine both in vitro and in vivo (5, 10 and 20 µg). It inhibits the secretion of granular components, clot retraction, and FeCl3-induced vascular occlusion of the carotid artery. Mechanism: attenuates the development of thrombosis and thromboembolism by inhibiting fibrin, clot formation, clotting factors, and platelet stimulation through DEP-1/PTP1B/αIIbβ3/kinases [202]. |
| Myricerin | Flavonoid | Reduces the ability of platelets to spread on collagen and form thrombi in vitro without affecting hemostasis in vivo. Inhibits the activities of PDI and ERp5 reductase (10, 30, or 100 µM). Mechanism: ERp5 and PDI inhibitor [203]. |
| Inhibits in vitro PAF-induced platelet adhesion (IC50: 13.1 mmol/L) and internal free Ca2+ concentration. It inhibits platelet aggregation PAF 1, 2, and 4 nmol/L with IC50: 34.8, 85.7, and 118.6 mmol/L. Mechanism: antagonizes the specific binding of the PAF receptor [204]. | ||
| Inhibits in vivo 3.6 µg/kg cat blood platelet aggregation. Reduces platelet thrombi in vitro at a concentration of 60 nM. Mechanism: inhibits PGI2 synthase [205]. | ||
| Inhibits platelet aggregation. Increases the cAMP stimulated by PGI2. Inhibits lipoxygenase activity (50 µM). Mechanism: modification of cAMP metabolism through inhibition of phosphodiesterase activity [206]. | ||
| Genistein | Isoflavonoid | Inhibits in vitro human platelet aggregation, serotonin secretion, and phosphorylation of protein tyrosine induced by collagen and TxA2. It slightly attenuates thrombin-induced protein tyrosine phosphorylation (100 μg/mL). Mechanism: preventive action on TxA2 binding, through inhibition of protein tyrosine phosphorylation [207]. |
| Inhibits in vitro dose-dependent collagen-induced platelet aggregation, NO production, and TNF-α secretion. Decreased secretion of MCP-1 induced by TNF-α in endothelial cells of the human umbilical vein (50 µM). Mechanism: through TNF-α [208]. | ||
| Inhibits platelet aggregation or the release of serotonin-induced by thrombin and Ca2+. Inhibits the COX pathway and PI3 and PI(4,5)P2 (50 µM) production. Mechanism: acts on the passage of inositol phospholipids [209]. | ||
| Decreases PAF stimulation of PLC activity at baseline. Inhibits PAF-stimulated platelet aggregation. Inhibits PI3 production and reduces induced phosphorylation (0.5 mM). Mechanism: inhibits tyrosine kinase early in signal transduction by inhibiting PLC. Decreases the activation of PKC and causes a reduction in protein phosphorylation [210]. | ||
| Glycythein | Isoflavonoid | Inhibits proliferation and synthesis of SMC DNA induced by PDGF-BB (3–10 μmol/L). Mechanism: inhibitory effect on SMC proliferation attenuates said proliferation (basic mechanism involved in atherosclerotic vascular change) [211]. |
| Formononetin | Isoflavonoid | Inhibits ferric chloride-induced arterial thrombus formation in rats and ADP- and thrombin-stimulated platelet aggregation in rats. Prolongs bleeding time and aPTT in mice Mechanism: inhibits platelet aggregation induced by ADP and thrombin and reduces the function of the endogenous clotting pathway [212]. |
| Inhibits PDGF-BB-induced proliferation and migration of human VSMCs. Inhibits upregulation of cell cycle-related proteins, matrix metalloproteinase, and PDGF-BB-induced phosphorylation of AKT in VSMC (1 µM). Mechanism: suppressive effect on PDGF-BB-stimulated proliferation and migration of VSMC, through inhibition of the AKT signaling pathway [213]. | ||
| Increases cell migration, tube formation, and levels of PECAM-1 and VEGF and platelet endothelial cells of the human umbilical vein. Protects against cerebral ischemia and reperfusion injury in rats. Improves cerebrovascular angiogenesis in human umbilical vein endothelial cells (10 and 20 µg/mL). Mechanism: suppresses cell apoptosis and improves cerebrovascular angiogenesis by promoting the expression of VEGF and PECAM-1 [214]. | ||
| Naringenin | Flavanone | Decreases kidney failure in rats and the lipid profile. Inhibits levels of inflammation markers and pro-oxidants in the kidney of rats. Moderate platelet parameters (50 mg/kg/90 days). Mechanism: improves kidney failure and platelet abnormalities through its antioxidant effects [215]. |
| Hesperetin | Flavanone | Inhibits in vitro platelet aggregation induced with collagen 5 μg/mL and AA 0.5 μmol/L (IC50: 20.5 and IC50: 69.2). Collagen-induced cytosolic Ca2+ mobilization decreases from 10 μg/mL to 20–50 μM. Inhibits collagen-stimulated serotonin secretion at IC50: 10.5 and IC50: 25.2. Mechanism: inhibition of PLCγ2 phosphorylation and collagen-induced COX-1 activity [216]. |
| Inhibits in vivo platelet aggregation induced with ADP and collagen (100 mg/kg) [217]. | ||
| Daidzein | Isoflavonoid | Inhibits in vitro dose-dependent collagen-induced platelet aggregation, NO production, and TNF-α secretion. Decreases secretion of MCP-1 induced by TNF-α in endothelial cells of the human umbilical vein (50 µM). Mechanism: through TNF-α [208]. |
| Inhibits proliferation and synthesis of SMC DNA induced by PDGF-BB (3–10 μmol/L). Mechanism: inhibits SMC proliferation (basic mechanism involved in atherosclerotic vascular change) [211]. | ||
| Catechin | Flavonol | Inhibits collagen-induced platelet aggregation and platelet adhesion to collagen (50–100 µmol/L). Mechanism: inhibits platelet function by reducing the production of hydrogen peroxide and PLC [218]. |
| Inhibits platelet aggregation induced by AA, ADP, and Mepinephrine (200 µg/mL). Decreases MDA production from AA-stimulated platelets (20–200 µg/mL). Mechanism: protects from peroxidative stress [219]. | ||
| Ex vivo platelet function improvement in iron-loaded rats, associated with impaired antioxidant defense, including free radical-induced hemolysis (10 mg/kg). Mechanism: through normalization of antioxidant status [220]. | ||
| Kaempferol-3-glucoside | Flavonol | Inhibits in vitro platelet aggregation induced with 100 μM AA and 10 μg/mL collagen to 22 μM. Inhibits ATP release stimulated with 7 µM ADP and 7 µM epinephrine to 25 µM. Mechanism: inhibits collagen 10 μg/mL induced TxA2 and PG formation L to 5 μM [221]. |
| Inhibits in vitro platelet aggregation induced by AA 100 μM and collagen 10 μg/mL to 100 μg/mL [222]. | ||
| Inhibits in vitro of platelet aggregation induced with AA 150 μM at IC50: 24 μM [206]. | ||
| Luteolin-7 glycoside | Flavone | Inhibits the proliferation of CMLV induced by PDGF-BB 1/2) and DNA synthesis in CMLV (50 uM). Mechanism: inhibits PDGF-BB 1/2 (ERK1/2)-induced extracellular signal-regulated kinase, Akt, and PLC activation [223]. |
| Coumarin | Benzopyrone | Inhibits platelet aggregation and release of ATP from rabbit platelets induced by AA, collagen, ADP, PAF) and U46619 (analogous to TxA2). Inhibits degradation of phosphoinositide caused by collagen and PAF (200 uM). Mechanism: inhibits TxA2 formation and phosphoinositide degradation [224]. |
| Inhibits platelet aggregation (25.75 ± 4.12%). Shows the percentage of binding with GPIIb/IIIa receptor (0.5 and 2 mM) [225]. | ||
| Inhibits AA-induced platelet aggregation. It does not interfere with the function of TxA2 synthase, but they were competitive antagonists of TxA2 receptors and inhibited COX-1 (50 µM). Mechanism: TxA2 stimulates its receptors on platelets, promoting platelet aggregation [226]. | ||
| Starch | Sugar | Reduces availability of the fibrinogen receptor. Mechanism: prevents the platelet from binding to fibrinogen, by blocking the access of ligands to the platelet fibrinogen receptor [227]. |
| Tocopherol | Tocopherol | Inhibits in vitro the aggregation induced by ADP, AA, PMA. Inhibits in vivo PMA-induced stimulation of PKC (400 to 1200 µL/d). Mechanism: inhibits human platelet aggregation through a PKC-dependent mechanism [228]. |
| Inhibits platelet aggregation induced by ADP and PMA. Increases NO release, ecNOS activation, and platelet protein SOD content. Decreases the activation of PKC (45 mg of α-tocopherol equivalents). Mechanism: increased NO release, ecNOS activation, and SOD protein content in platelets [229]. | ||
| Omega-3 and 6 | Fatty acid | Decreases in vivo reactivity of P2Y12 by 22.2%. Inhibits ADP-induced platelet aggregation. Mechanism: potentiates platelet response to clopidogrel then percutaneous coronary intervention [230]. |
| Reduces in vivo thrombin formation and oxidative stress and favorably alters the properties of the fibrin clot (1 g/day of PUFA n-3) [231]. | ||
| Increases the total surface load of platelets and attenuates platelet activation (1 to 8 g/day) [232]. | ||
| Delphinidin | Anthocyanin | Inhibits in vitro the secretion of alpha granules: PF4, β-TG, P-selectin, TGF-β1, RANTES, ATP, and serotonin and CD63 induced with thrombin 0.5 U/mL. Inhibits the secretion of dense granules: ATP and serotonin-induced with thrombin 0.5 U/mL (320 mg/day). Mechanism: inhibits the phosphorylation of the MAPK family stimulated by thrombin 0.5 U/mL. Inhibits the activation of PI3K/Akt, phosphorylation of eNOS, and production of cGMP induced by thrombin 0.5 U/mL. Inhibits in vivo the secretion of alpha granules PF4, β-TG, P-selectin, TGF-β1, RANTES stimulated with thrombin 0.5 U/mL (320 mg/day) [233]. |
| Inhibits platelet aggregation induced with 5 µM ADP, 2 µg/mL collagen, and 100 µM TRAP. Inhibits in vitro the activation and secretion of P-selectin, CD63, CD40L, αllbβ3, and fibrinogen induced with 200 μM ADP, 10 μg/mL collagen, 1 U/mL thrombin, and 250 μM TRAP. Mechanism: inhibits the phosphorylation of MAPK induced by collagen 25 μg/mL. Inhibits in vivo the formation of the thrombus induced by collagen 100 μg/mL, under controlled flow. Inhibits FeCl3-induced thrombus formation at 50 μg/mL [234]. | ||
| Cyanidin-3-O-glucoside | Anthocyanin | Inhibits in vitro the secretion of alpha granules: PF4, β-TG, P-selectin, TGF-β1, RANTES, ATP, and serotonin and CD63 induced with thrombin 0.5 U/mL. Mechanism: inhibits the phosphorylation of the MAPK family induced by thrombin 0.5 U/mL. Inhibits PI3K/Akt activation, eNOS phosphorylation, and 0.5 U/mL thrombin-induced cGMP production [233]. Inhibits in vivo the secretion of alpha granules PF4, β-TG, P-selectin, TGF-β1, RANTES induced with thrombin 0.5 U/mL at 320 mg/day [233]. |
| Inhibits platelet aggregation induced with 2.5 µg/mL collagen, 0.1 U/mL thrombin, and 100 µM TRAP-6. Mechanism: GPVI collagen receptor pathway. Inhibits collagen 2.5 µg/mL induced phosphorylation of protein tyrosine at 5–50 µM [235]. Inhibits 0.5–50 µM collagen-induced thrombus formation in vivo, under controlled flow. Inhibits FeCl3-induced thrombus formation (5–50 µM) [234]. | ||
| Pelargonidine-3-O-glucoside | Anthocyanin | Prolongs aPTT and PT. Inhibits thrombin and FXa activity and production in human umbilical vein endothelial cells. Inhibits thrombin-catalyzed polymerization of fibrin and platelet aggregation and anticoagulant effect elicited in mice (10 µM) [236]. |
| Inhibits in vitro the secretion of alpha granules: PF4, β-TG, P-selectin, TGF-β1, RANTES, ATP, and serotonin and CD63 induced with thrombin 0.5 U/mL. Inhibits the secretion of dense granules, ATP, and serotonin with thrombin 0.5 U/mL. Mechanism: inhibition of the phosphorylation of the MAPK family induced by thrombin 0.5 U/mL. Inhibits PI3K/Akt activation, eNOS phosphorylation, and 0.5 U/mL thrombin-induced cGMP production [233]. Inhibits in vivo the secretion of alpha granules PF4, β-TG, P-selectin, TGF-β1, RANTES induced with thrombin 0.5 U/mL (320 mg/day) [233]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez, L.; Mendez, D.; Montecino, H.; Carrasco, B.; Arevalo, B.; Palomo, I.; Fuentes, E. Role of Phaseolus vulgaris L. in the Prevention of Cardiovascular Diseases—Cardioprotective Potential of Bioactive Compounds. Plants 2022, 11, 186. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11020186
Rodríguez L, Mendez D, Montecino H, Carrasco B, Arevalo B, Palomo I, Fuentes E. Role of Phaseolus vulgaris L. in the Prevention of Cardiovascular Diseases—Cardioprotective Potential of Bioactive Compounds. Plants. 2022; 11(2):186. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11020186
Chicago/Turabian StyleRodríguez, Lyanne, Diego Mendez, Hector Montecino, Basilio Carrasco, Barbara Arevalo, Iván Palomo, and Eduardo Fuentes. 2022. "Role of Phaseolus vulgaris L. in the Prevention of Cardiovascular Diseases—Cardioprotective Potential of Bioactive Compounds" Plants 11, no. 2: 186. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11020186