
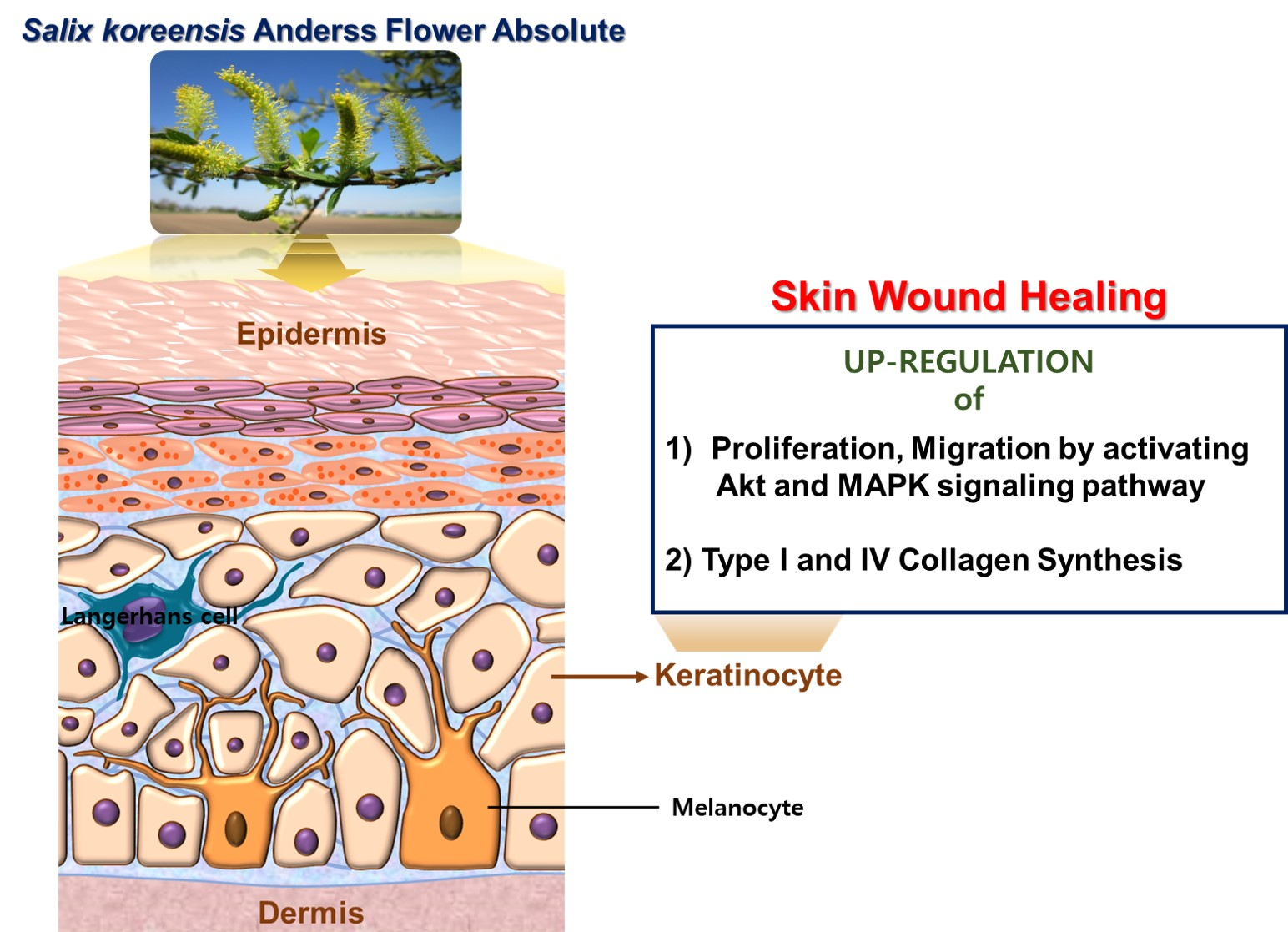
Chemical Composition of Salix koreensis Anderss Flower Absolute and Its Skin Wound Healing Activities In Vitro


Abstract
:
1. Introduction
2. Results
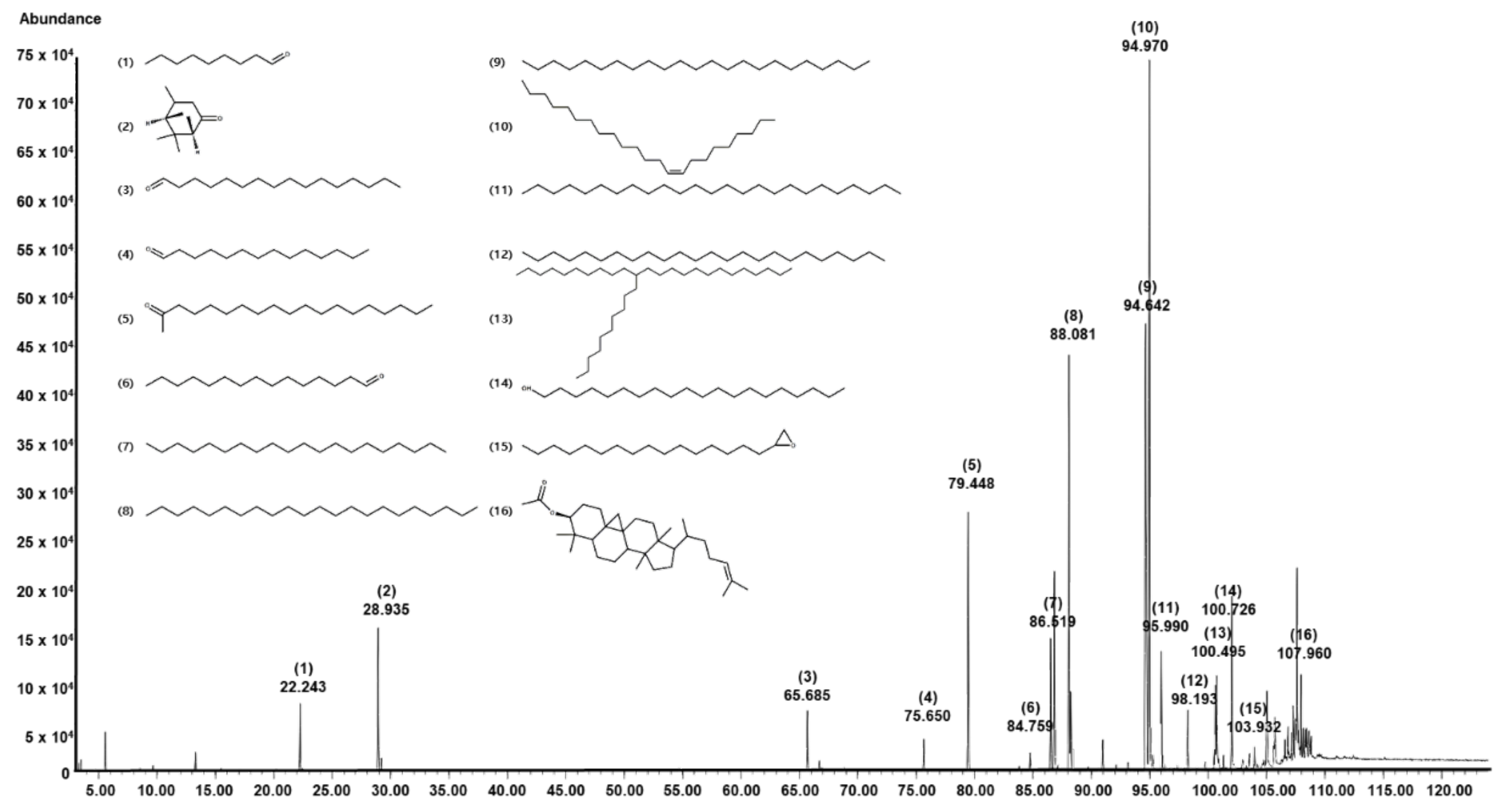
2.1. Chemical Composition of SKAFAb
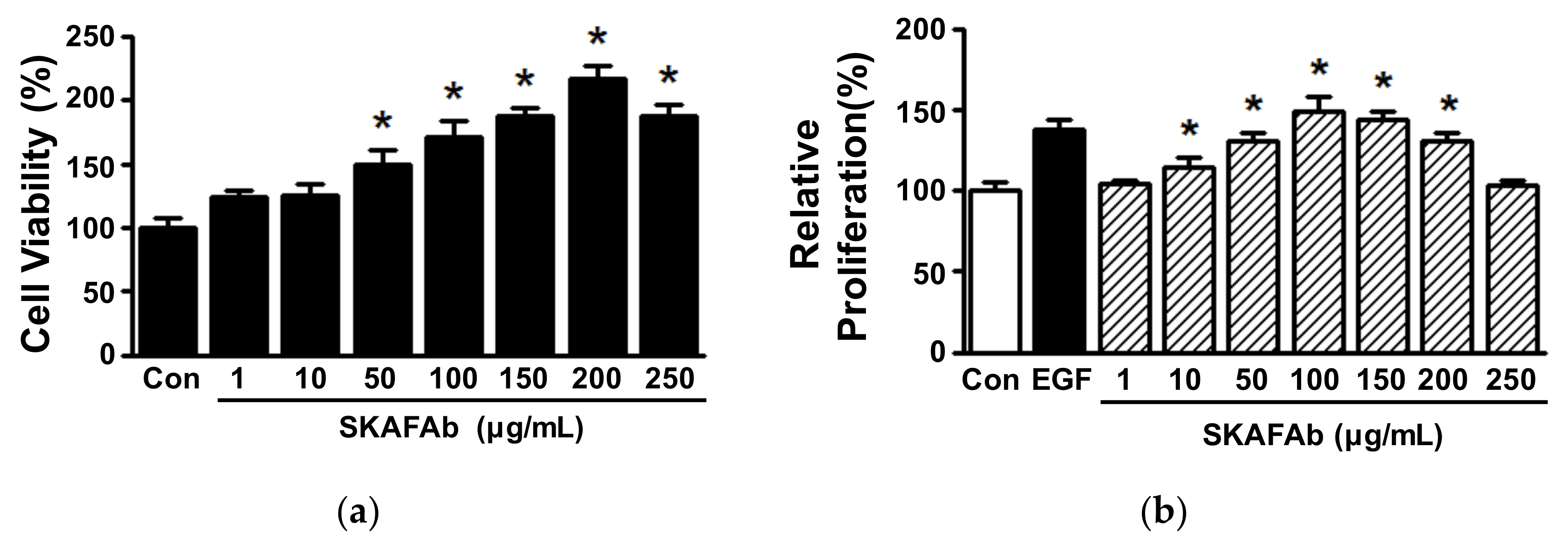
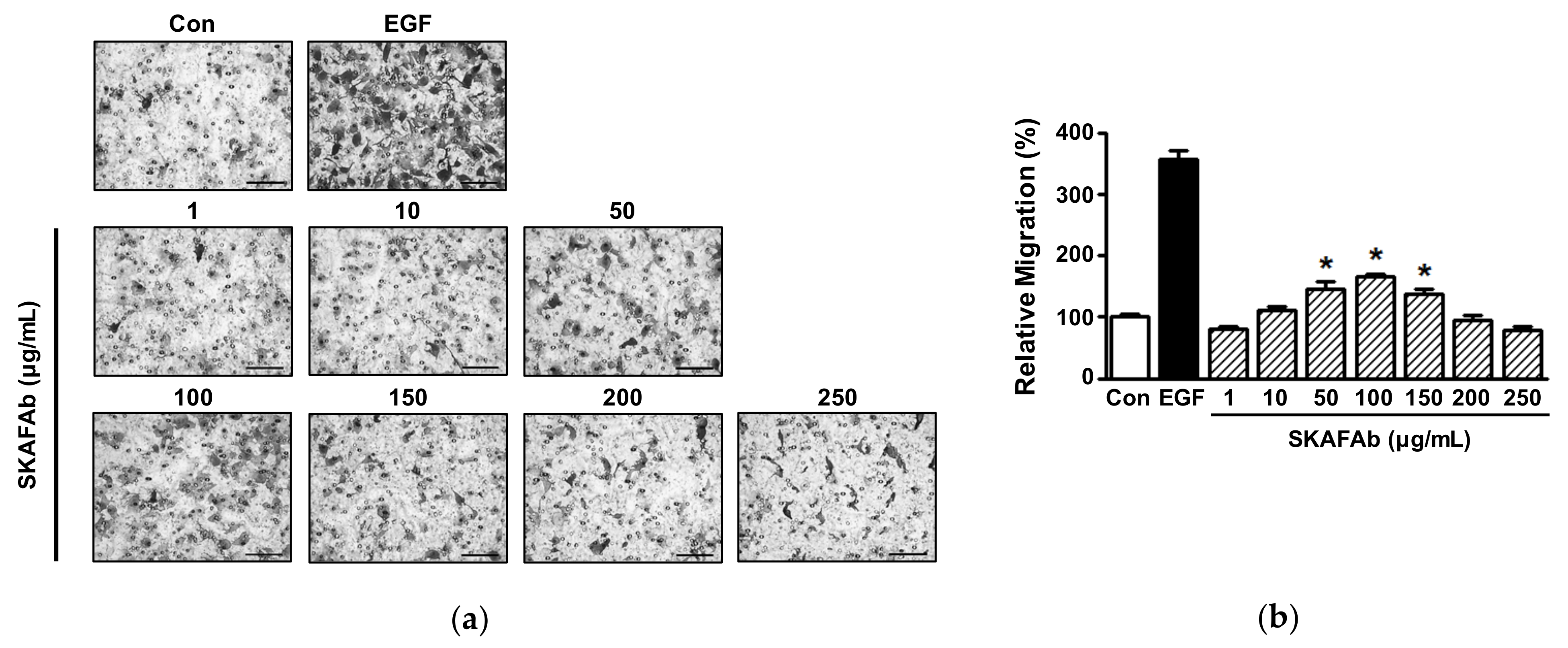
2.2. Proliferative and Migratory Activities of Keratinocytes Exposed to SKAFAb
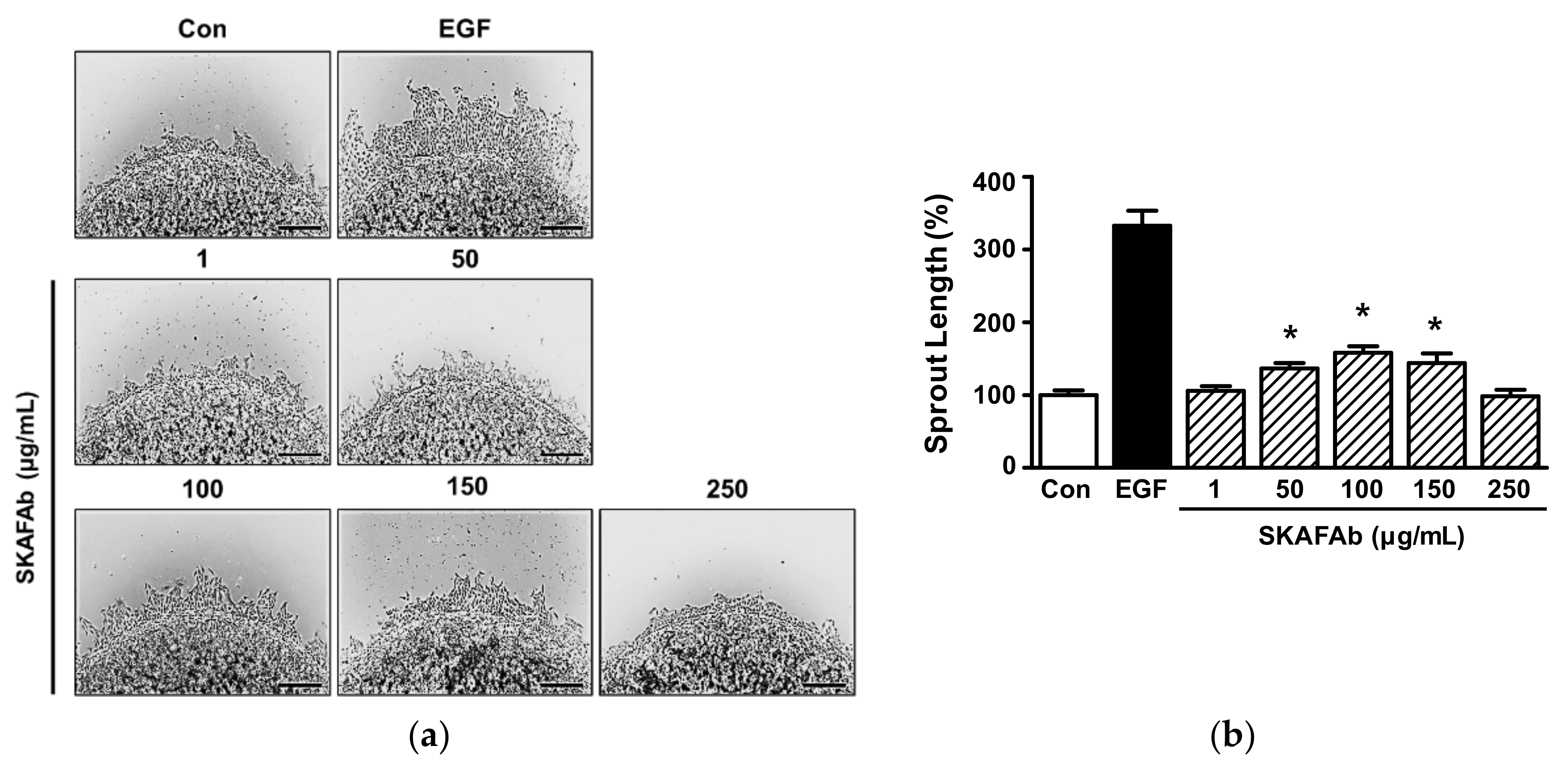
2.3. Effect of SKAFAb on Keratinocyte Sprout Outgrowth
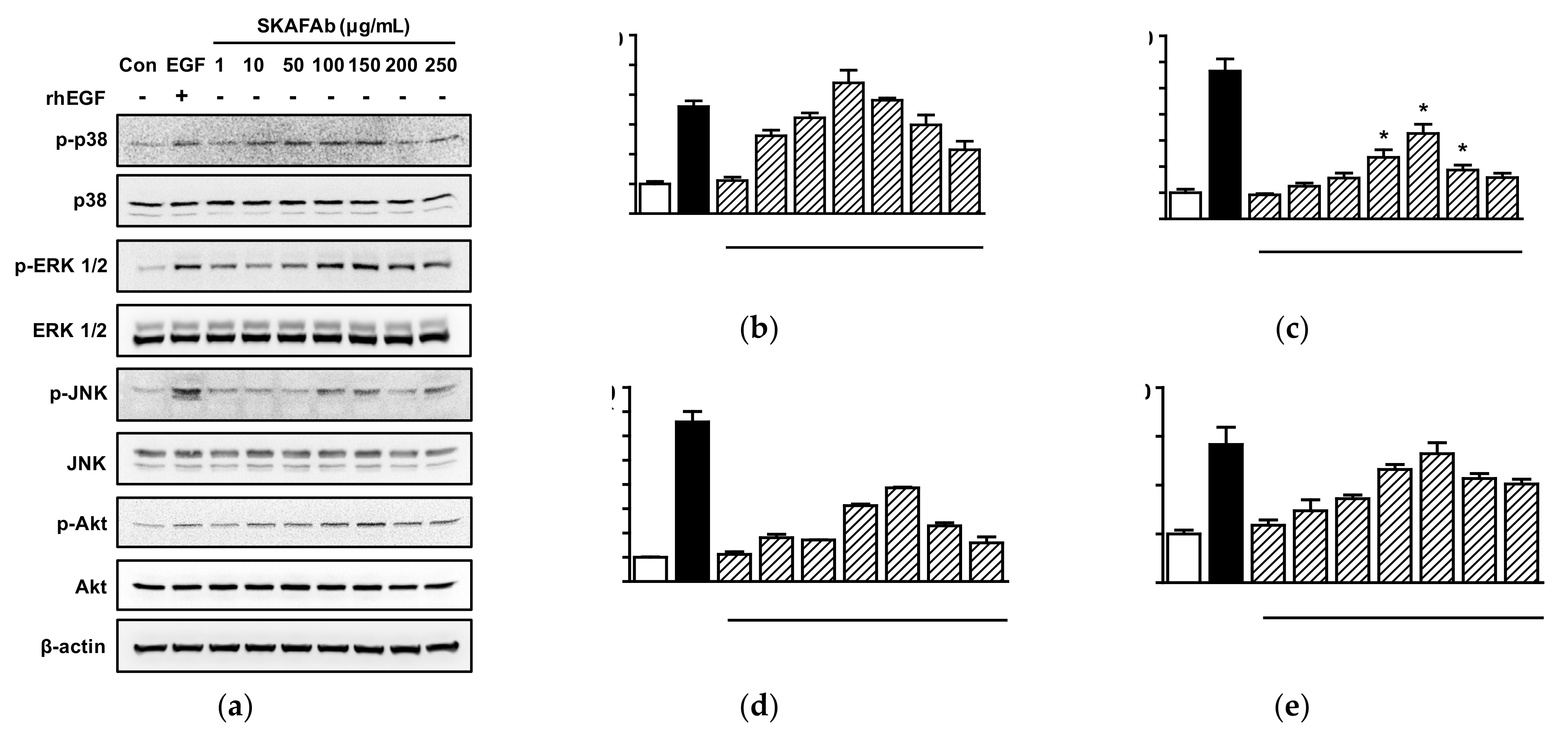
2.4. Effects of SKAFAb on Signaling Kinases in Keratinocytes
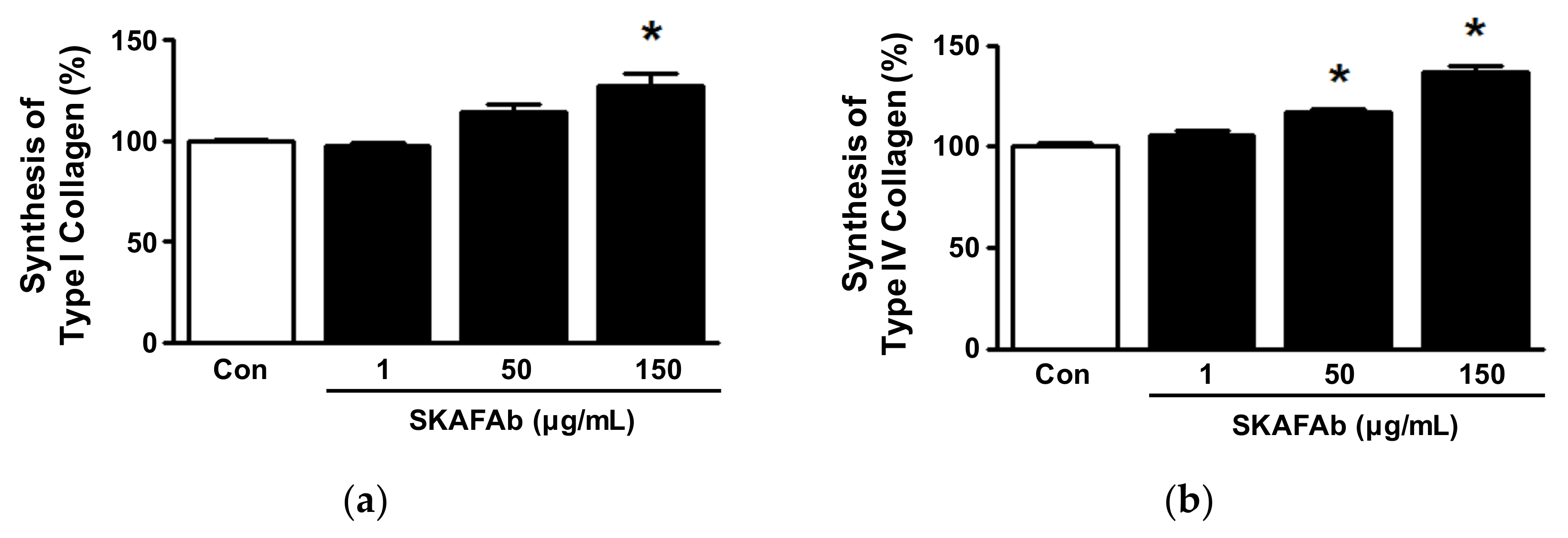
2.5. Collagen Synthesis in Keratinocytes Exposed to SKAFAb
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Extraction of Salix koreensis Anderss Flower Absolute
4.3. Identification of Components in SKAFAb
4.4. Cell Culture
4.5. Cell Viability Assay
4.6. Proliferation Assay
4.7. Migration Assay
4.8. Collagen Sprout Assay
4.9. Collagen Synthesis Assay
4.10. Western Blotting
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Dipietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Cellular signalling and photobiomodulation in chronic wound repair. Int. J. Mol. Sci. 2021, 22, 11223. [Google Scholar] [CrossRef]
- Michopoulou, A.; Rousselle, P. How do epidermal matrix metalloproteinases support re-epithelialization during skin healing? Eur. J. Dermatol. 2015, 25, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Ter Horst, B.; Chouhan, G.; Moiemen, N.S.; Grover, L.M. Advances in keratinocyte delivery in burn wound care. Adv. Drug Deliv. Rev. 2018, 123, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, P.; Braye, F.; Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 344–365. [Google Scholar] [CrossRef]
- Freedberg, I.M.; Tomic-Canic, M.; Komine, M.; Blumenberg, M. Keratins and the keratinocyte activation cycle. J. Investig. Dermatol. 2001, 116, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, M.M.; Gaudino, G. Cellular and molecular facets of keratinocyte reepithelization during wound healing. Exp. Cell Res. 2005, 304, 274–286. [Google Scholar] [CrossRef]
- Puccinelli, T.J.; Bertics, P.J.; Masters, K.S. Regulation of keratinocyte signaling and function via changes in epidermal growth factor presentation. Acta Biomater. 2010, 6, 3415–3425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chen, J.; Kirsner, R. Pathophysiology of acute wound healing. Clin. Dermatol. 2007, 25, 9–18. [Google Scholar] [CrossRef]
- Sharma, S.; Rai, V.K.; Narang, R.K.; Markandeywar, T.S. Collagen-based formulations for wound healing: A literature review. Life Sci. 2022, 290, 120096. [Google Scholar] [CrossRef] [PubMed]
- Brett, D.A. A Review of collagen and collagen-based wound dressings. Wounds 2008, 20, 347–356. [Google Scholar] [PubMed]
- Pereira, R.F.; Bártolo, P.J. Traditional Therapies for Skin Wound Healing. Adv. Wound Care 2016, 5, 208–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasic-Kostov, M.; Arsic, I.; Pavlovic, D.; Stojanovic, S.; Najman, S.; Naumovic, S.; Tadic, V. Towards a modern approach to traditional use: In vitro and in vivo evaluation of Alchemilla vulgaris L. gel wound healing potential. J. Ethnopharmacol. 2019, 238, 111789. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Vijayakumar, M.; Govindarajan, R.; Pushpangadan, P. Ethnopharmacological approaches to wound healing—Exploring medicinal plants of India. J. Ethnopharmacol. 2007, 114, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The traditional medicine and modern medicine from natural products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karapanagioti, E.G.; Assimopoulou, A.N. Naturally occurring wound healing agents: An evidence-based review. Curr. Med. Chem. 2016, 23, 3285–3321. [Google Scholar] [CrossRef]
- Tawfeek, N.; Mahmoud, M.F.; Hamdan, D.I.; Sobeh, M.; Farrag, N.; Wink, M.; El-Shazly, A.M. Phytochemistry, pharmacology and medicinal uses of plants of the genus Salix: An updated review. Front. Pharmacol. 2021, 12, 593856. [Google Scholar] [CrossRef] [PubMed]
- Koo, Y.B.; Woo, K.S.; Yeo, J.K.; Kim, Y.S. Selection of superior poplar and willow clones in growth performance and adaptation abilities at Sudokwon landfill site. J. Korean For. Soc. 2006, 95, 743–750. [Google Scholar]
- Kim, E.J.; Kim, M.H. Anti-oxidant and anti-inflammatory effects of Salix koreensis Andersson in DC. leaf methanol extract in vitro models. CELLMED 2016, 6, e28. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Min, J.; Kang, H. Hepatoprotective effects of willow (Salix koreensis ANDERSS.) branch extracts against cytotoxicity induced by tert-butyl hydroperoxide in human hepatoma cells. J. Korean Soc. Food Sci. Nutr. 2020, 49, 1328–1334. [Google Scholar] [CrossRef]
- Sen, C.K.; Gordillo, G.M.; Roy, S.; Kirsner, R.; Lambert, L.; Hunt, T.K.; Gottrup, F.; Gurtner, G.C.; Longaker, M.T. Human skin wounds: A major and snowballing threat to public health and the economy. Wound Repair Regen. 2009, 17, 763–771. [Google Scholar] [CrossRef] [Green Version]
- Takazawa, Y.; Ogawa, E.; Saito, R.; Uchiyama, R.; Ikawa, S.; Uhara, H.; Okuyama, R. Notch down-regulation in regenerated epidermis contributes to enhanced expression of interleukin-36α and suppression of keratinocyte differentiation during wound healing. J. Dermatol. Sci. 2015, 79, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Hwang, D.I.; Won, K.J.; Kim, D.Y.; Kim, H.B.; Li, Y.; Lee, H.M. Chemical composition of Patrinia scabiosifolia flower absolute and its migratory and proliferative activities in human keratinocytes. Chem. Biodivers. 2019, 16, e1900252. [Google Scholar] [CrossRef]
- Li, Y.; Won, K.J.; Kim, D.Y.; Kim, H.B.; Kang, H.M.; Lee, S.Y.; Lee, H.M. Positive promoting effects of Smilax China flower absolute on the wound healing/skin barrier repair-related responses of HaCaT human skin keratinocytes. Chem. Biodivers. 2021, 18, e2001051. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, J.Q.; Zheng, Z.; Zhang, J.; Wang, S.Y.; Han, S.C.; Zhou, Q.; Guan, H.; Li, C.; Su, L.L.; et al. Human amniotic epithelial stem cells promote wound healing by facilitating migration and proliferation of keratinocytes via ERK, JNK and AKT signaling pathways. Cell Tissue Res. 2016, 365, 85–99. [Google Scholar] [CrossRef]
- Haase, I.; Evans, R.; Pofahl, R.; Watt, F.M. Regulation of keratinocyte shape, migration and wound epithelialization by IGF-1- and EGF-dependent signalling pathways. J. Cell Sci. 2003, 116, 3227–3238. [Google Scholar] [CrossRef] [Green Version]
- Pazyar, N.; Yaghoobi, R.; Rafiee, E.; Mehrabian, A.; Feily, A. Skin wound healing and phytomedicine: A review. Skin Pharmacol. Physiol. 2014, 27, 303–310. [Google Scholar] [CrossRef]
- Negahdari, S.; Galehdari, H.; Kesmati, M.; Rezaie, A.; Shariati, G. Wound healing activity of extracts and formulations of Aloe vera, Henna, Adiantum capillus-veneris, and Myrrh on mouse dermal fibroblast cells. Int. J. Prev. Med. 2017, 8, 18. [Google Scholar] [CrossRef]
- Lee, S.Y.; Won, K.J.; Kim, D.Y.; Kim, M.J.; Won, Y.R.; Kim, N.Y.; Lee, H.M. Wound healing-promoting and melanogenesis-inhibiting activities of Angelica polymorpha Maxim. flower absolute in vitro and its chemical composition. Molecules 2021, 26, 6172. [Google Scholar] [CrossRef]
- Park, S.; Kang, W.; Choi, D.; Son, B.; Park, T. Nonanal stimulates growth factors via cyclic adenosine monophosphate (cAMP) signaling in human hair follicle dermal papilla cells. Int. J. Mol. Sci. 2020, 21, 8054. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Hernández, Y.; Sánchez-Hernández, G.R.; Reyes-Cervantes, E.; Romero-Arenas, O.; Pérez-Xochipa, I.; Villa-Ruano, N. Chemical variation and pharmacological properties of Dyssodia decipiens essential oil. Chem. Biodivers. 2020, 17, e2000487. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, C.R.; Matsuo, A.L.; Massaoka, M.H.; Girola, N.; Azevedo, R.A.; Rabaça, A.N.; Farias, C.F.; Pereira, F.V.; Matias, N.S.; Silva, L.P.; et al. Antitumor activity of kielmeyera coriacea leaf constituents in experimental melanoma, tested in vitro and in vivo in syngeneic mice. Adv. Pharm. Bull. 2014, 4, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Seger, R.; Krebs, E.G. The MAPK signaling cascade. FASEB J. 1995, 9, 726–735. [Google Scholar] [CrossRef]
- Su, L.; Fu, L.; Li, X.; Zhang, Y.; Li, Z.; Wu, X.; Li, Y.; Bai, X.; Hu, D. Loss of CAR promotes migration and proliferation of HaCaT cells, and accelerates wound healing in rats via Src-p38 MAPK pathway. Sci. Rep. 2016, 6, 19735. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, L.; Zhang, Q.; Wang, W.; Zhang, D.; Jia, J.; Lv, Y.; Yuan, H.; Song, H.; Xiang, F.; et al. Microtubule-associated protein 4 phosphorylation regulates epidermal keratinocyte migration and proliferation. Int. J. Biol. Sci. 2019, 15, 1962–1976. [Google Scholar] [CrossRef]
- Hoq, M.I.; Niyonsaba, F.; Ushio, H.; Aung, G.; Okumura, K.; Ogawa, H. Human catestatin enhances migration and proliferation of normal human epidermal keratinocytes. J. Dermatol. Sci. 2011, 64, 108–118. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, Y.K.; Eun, H.C.; Cho, K.H.; Chung, J.H. All-trans retinoic acid antagonizes UV-induced VEGF production and angiogenesis via the inhibition of ERK activation in human skin keratinocytes. J. Investig. Dermatol. 2006, 126, 2697–2706. [Google Scholar] [CrossRef] [Green Version]
- Gazel, A.; Nijhawan, R.I.; Walsh, R.; Blumenberg, M. Transcriptional profiling defines the roles of ERK and p38 kinases in epidermal keratinocytes. J. Cell Physiol. 2008, 215, 292–308. [Google Scholar] [CrossRef]
- Kang, J.; Chen, W.; Xia, J.; Li, Y.; Yang, B.; Chen, B.; Sun, W.; Song, X.; Xiang, W.; Wang, X.; et al. Extracellular matrix secreted by senescent fibroblasts induced by UVB promotes cell proliferation in HaCaT cells through PI3K/AKT and ERK signaling pathways. Int. J. Mol. Med. 2008, 21, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.E.; Cho, H.; Ishihara, A.; Kim, B.; Kim, O. Cell proliferation and migration mechanism of caffeoylserotonin and serotonin via serotonin 2B receptor in human keratinocyte HaCaT cells. BMB Rep. 2018, 51, 188–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.M.; Li, W.; Man, X.Y.; Zhou, J.; Chen, J.Q.; Cai, S.Q.; Zheng, M. Pigment epithelium-derived factor plays an inhibitory role in proliferation and migration of HaCaT cells. Mol. Biol. Rep. 2011, 38, 2099–2105. [Google Scholar] [CrossRef]
- Schüppel, M.; Kürschner, U.; Kleuser, U.; Schäfer-Korting, M.; Kleuser, B. Sphingosine 1-phosphate restrains insulin-mediated keratinocyte proliferation via inhibition of Akt through the S1P2 receptor subtype. J. Investig. Dermatol. 2008, 128, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, E.A. Extracellular matrix and keratinocyte migration. Clin. Exp. Dermatol. 2001, 26, 525–530. [Google Scholar] [CrossRef]
- Uitto, J. Connective tissue biochemistry of the aging dermis. Age-related alterations in collagen and elastin. Dermatol. Clin. 1986, 4, 433–446. [Google Scholar] [CrossRef]
- Savvas, M.; Bishop, J.; Laurent, G.; Watson, N.; Studd, J. Type III collagen content in the skin of postmenopausal women receiving oestradiol and testosterone implants. Br. J. Obstet. Gynaecol. 1993, 100, 154–156. [Google Scholar] [CrossRef]
- Seo, W.Y.; Kim, J.H.; Baek, D.S.; Kim, S.J.; Kang, S.; Yang, W.S.; Song, J.A.; Lee, M.S.; Kim, S.; Kim, Y.S. Production of recombinant human procollagen type I C-terminal propeptide and establishment of a sandwich ELISA for quantification. Sci. Rep. 2017, 7, 15946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Sierra, J.O.; Kelly, R.; Kirsner, R.S.; Li, J. Wool-derived keratin stimulates human keratinocyte migration and types IV and VII collagen expression. Exp. Dermatol. 2012, 21, 458–460. [Google Scholar] [CrossRef]
- Woodley, D.T.; Bachmann, P.M.; O’Keefe, E.J. Laminin inhibits human keratinocyte migration. J. Cell Physiol. 1988, 136, 140–146. [Google Scholar] [CrossRef]
- Kang, H.M.; Won, K.J.; Kim, D.Y.; Lee, S.Y.; Kim, M.J.; Won, Y.R.; Kim, B.; Lee, H.M. Chemical composition of Miscanthus sinensis var. purpurascens flower absolute and its beneficial effects on skin wound healing and melanogenesis-related cell activities. Chem. Biodivers. 2021, 18, e2100383. [Google Scholar] [CrossRef]
- Kovats, E. Gas chromatographic characterization of organic substances in the retention index system. Adv. Chromatogr. 1965, 1, 229–247. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Component Name | RT 1 | RI 2 | Area (%) | CAS No. | |
|---|---|---|---|---|---|---|
| Observed | Literature | |||||
| 1 | Nonanal | 22.24 | 1104 | 1104 | 1.84 | 124-19-6 |
| 2 | Verbenone | 28.94 | 1201 | 1204 | 4.38 | 1196-01-6 |
| 3 | Hexadecanal | 65.69 | 1814 | 1814 | 1.78 | 629-80-1 |
| 4 | Tetradecanal | 75.65 | 2018 | 1940 | 0.90 | 124-25-4 |
| 5 | 2-Nonadecanone | 79.45 | 2101 | 2101 | 10.16 | 629-66-3 |
| 6 | Pentadecanal | 84.76 | 2222 | - | 0.56 | 2765-11-9 |
| 7 | Eicosane | 86.52 | 2263 | 2000 | 4.31 | 112-95-8 |
| 8 | Tricosane | 88.08 | 2300 | 2300 | 14.26 | 638-67-5 |
| 9 | Docosane | 94.64 | 2465 | 2200 | 23.36 | 629-97-0 |
| 10 | 9-Tricosene, (Z)- | 94.97 | 2474 | 2272 | 26.69 | 27519-02-4 |
| 11 | Pentacosane | 95.99 | 2500 | 2500 | 3.81 | 629-99-2 |
| 12 | Tetracosane | 98.19 | 2574 | 2400 | 1.70 | 646-31-1 |
| 13 | Tetracosane, 11-decyl- | 100.50 | 2665 | - | 0.54 | 55429-84-0 |
| 14 | 1-Eicosanol | 100.73 | 2675 | 2755 | 3.24 | 629-96-9 |
| 15 | Oxirane, hexadecyl- | 103.93 | 2836 | - | 0.51 | 7390-81-0 |
| 16 | Cycloartenol acetate | 107.96 | 3089 | 3389 | 1.98 | 1259-10-5 |
| Total Identified (%) | 100.00 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, N.Y.; Won, K.J.; Kim, H.B.; Kim, D.Y.; Kim, M.J.; Won, Y.R.; Lee, H.M. Chemical Composition of Salix koreensis Anderss Flower Absolute and Its Skin Wound Healing Activities In Vitro. Plants 2022, 11, 246. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11030246
Kim NY, Won KJ, Kim HB, Kim DY, Kim MJ, Won YR, Lee HM. Chemical Composition of Salix koreensis Anderss Flower Absolute and Its Skin Wound Healing Activities In Vitro. Plants. 2022; 11(3):246. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11030246
Chicago/Turabian StyleKim, Nan Young, Kyung Jong Won, Ha Bin Kim, Do Yoon Kim, Mi Jung Kim, Yu Rim Won, and Hwan Myung Lee. 2022. "Chemical Composition of Salix koreensis Anderss Flower Absolute and Its Skin Wound Healing Activities In Vitro" Plants 11, no. 3: 246. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11030246