
Antioxidation, Anti-Inflammation, and Regulation of SRD5A Gene Expression of Oryza sativa cv. Bue Bang 3 CMU Husk and Bran Extracts as Androgenetic Alopecia Molecular Treatment Substances

 ,
,  , , ,
, , ,  , ,
, ,  ,
,  , ,
, ,  ,
,  , and
, and 
Abstract
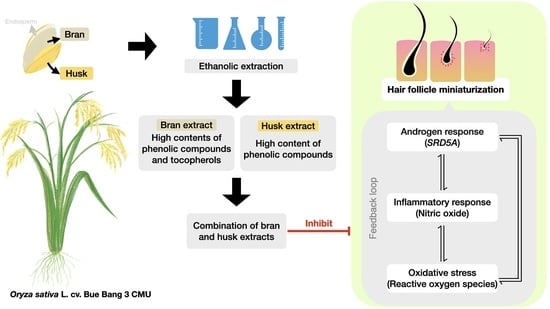
:
1. Introduction
2. Results and Discussion
2.1. Extraction Yield and Bioactive Compound Estimation
2.2. Profile of Phenolic Compounds, Phytic Acid and Tocopherols of BB3CMU-H and BB3CMU-RB
2.3. Antioxidant Activities
2.4. Anti-Inflammatory Activity
2.5. Effect of BB3CMU-H and BB3CMU-RB on Gene Expression of Steroid 5α-Reductase Isoenzymes
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material and Extract Preparation
3.3. Bioactive Compound Estimation
3.3.1. Determination of Total Phenolic Content
3.3.2. Determination of Total Flavonoid Content
3.3.3. Determination of Total Polysaccharide Content
3.3.4. Determination of Total Protein Content
3.4. Bioactive Characterization of Extracts
3.4.1. Determination of Phenolic Compounds and Phytic Acid by Liquid Chromatography-Mass Spectrometry (LC-MS)
3.4.2. Determination of Tocopherols by High-Performance Liquid Chromatography (HPLC)
3.5. Antioxidant Activities
3.5.1. DPPH Radical Scavenging Assay
3.5.2. ABTS Radical Cation Scavenging Assay
3.5.3. Metal Chelating Assay
3.6. Cell Culture Procedure
3.7. Cell Viability Assay
3.8. Anti-Inflammatory Activity
3.9. Semi-Quantitative Reverse Transcription and Polymerase Chain Reaction
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- English, R.S. A hypothetical pathogenesis model for androgenic alopecia: Clarifying the dihydrotestosterone paradox and rate-limiting recovery factors. Med. Hypotheses 2018, 111, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Ceruti, J.M.; Leirós, G.J.; Balañá, M.E. Androgens and androgen receptor action in skin and hair follicles. Mol. Cell. Endocrinol. 2018, 465, 122–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trüeb, R.M. Oxidative stress in ageing of hair. Int. J. Trichology 2009, 1, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inui, S.; Itami, S. Molecular basis of androgenetic alopecia: From androgen to paracrine mediators through dermal papilla. J. Dermatol. Sci. 2011, 61, 1–6. [Google Scholar] [CrossRef]
- Wolf, R.; Schönfelder, G.; Paul, M.; Blume-Peytavi, U. Nitric oxide in the human hair follicle: Constitutive and dihydrotestosterone-induced nitric oxide synthase expression and NO production in dermal papilla cells. J. Mol. Med. 2003, 81, 110–117. [Google Scholar] [CrossRef]
- Balık, A.R.; Balık, Z.B.; Aktaş, A.; Neşelioğlu, S.; Karabulut, E.; Karabulut, A.B. Examination of androgenetic alopecia with serum biomarkers. J. Cosmet. Dermatol. 2021, 20, 1855–1859. [Google Scholar] [CrossRef]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef]
- Sowden, H.M.; Naseem, K.M.; Tobin, D.J. Differential expression of nitric oxide synthases in human scalp epidermal and hair follicle pigmentary units: Implications for regulation of melanogenesis. Br. J. Dermatol. 2005, 153, 301–309. [Google Scholar] [CrossRef]
- Herman, A.; Herman, A.P. Mechanism of action of herbs and their active constituents used in hair loss treatment. Fitoterapia 2016, 114, 18–25. [Google Scholar] [CrossRef]
- Wadley, A.J.; Veldhuijzen van Zanten, J.J.C.S.; Aldred, S. The interactions of oxidative stress and inflammation with vascular dysfunction in ageing: The vascular health triad. Age 2013, 35, 705–718. [Google Scholar] [CrossRef]
- Ustuner, E.T. Baldness may be caused by the weight of the scalp: Gravity as a proposed mechanism for hair loss. Med. Hypotheses 2008, 71, 505–514. [Google Scholar] [CrossRef]
- Upton, J.H.; Hannen, R.F.; Bahta, A.W.; Farjo, N.; Farjo, B.; Philpott, M.P. Oxidative stress–associated senescence in dermal papilla cells of men with androgenetic alopecia. J. Investig. Dermatol. 2015, 135, 1244–1252. [Google Scholar] [CrossRef] [Green Version]
- Prie, B.E.; Iosif, L.; Tivig, I.; Stoian, I.; Giurcaneanu, C. Oxidative stress in androgenetic alopecia. J. Med. Life 2016, 9, 79–83. [Google Scholar]
- Zgonc Škulj, A.; Poljšak, N.; Kočevar Glavač, N.; Kreft, S. Herbal preparations for the treatment of hair loss. Arch. Dermatol. 2020, 312, 395–406. [Google Scholar] [CrossRef]
- Semalty, M.; Semalty, A.; Joshi, G.P.; Rawat, M.S.M. Hair growth and rejuvenation: An overview. J. Dermatol. Treat. 2011, 22, 123–132. [Google Scholar] [CrossRef]
- Peanparkdee, M.; Iwamoto, S. Bioactive compounds from by-products of rice cultivation and rice processing: Extraction and application in the food and pharmaceutical industries. Trends Food Sci. Technol. 2019, 86, 109–117. [Google Scholar] [CrossRef]
- Huang, S.H.; Ng, L.T. Quantification of polyphenolic content and bioactive constituents of some commercial rice varieties in Taiwan. J. Food Compos. Anal. 2012, 26, 122–127. [Google Scholar] [CrossRef]
- Jha, P.; Das, A.J.; Deka, S.C. Optimization of ultrasound and microwave assisted extractions of polyphenols from black rice (Oryza sativa cv. Poireton) husk. J. Food Sci. Technol. 2017, 54, 3847–3858. [Google Scholar] [CrossRef]
- Goufo, P.; Trindade, H. Rice antioxidants: Phenolic acids, flavonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, γ-oryzanol, and phytic acid. Food Sci. Nutr. 2014, 2, 75–104. [Google Scholar] [CrossRef]
- Sreethong, T.; Rerkasem, B.; Dell, B.; Jamjod, S. Identifying rice grains with premium nutritional quality among on-farm germplasm in the highlands of northern Thailand. Qual. Assur. Saf. Crop. Foods. 2020, 12, 12–23. [Google Scholar] [CrossRef]
- Jamjod, S.; Yimyam, N.; Lordkaew, S.; Promuthai, C.; Rerkasem, B. Characterization of on-farm rice germplasm in an area of the crop’s center of diversity. Chiang Mai J. Sci. 2017, 16, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Surin, S.; Seesuriyachan, P.; Thakeow, P.; You, S.; Phimolsiripol, Y. Antioxidant and antimicrobial properties of polysaccharides from rice brans. Chiang Mai J. Sci. 2018, 45, 1372–1382. [Google Scholar]
- Yamuangmorn, S.; Promuthai, C. The potential of high-anthocyanin purple rice as a functional ingredient in human health. Antioxidants 2021, 10, 833. [Google Scholar] [CrossRef]
- Butsat, S.; Siriamornpun, S. Antioxidant capacities and phenolic compounds of the husk, bran and endosperm of Thai rice. Food Chem. 2010, 119, 606–613. [Google Scholar] [CrossRef]
- Park, H.Y.; Lee, K.W.; Choi, H.D. Rice bran constituents: Immunomodulatory and therapeutic activities. Food Funct. 2017, 8, 935–943. [Google Scholar] [CrossRef]
- Phuwadolpaisarn, P. Comparison of β-glucan content in milled rice, rice husk and rice bran from rice cultivars grown in different locations of Thailand and the relationship between β-glucan and amylose contents. Molecules 2021, 26, 6368. [Google Scholar] [CrossRef]
- Yang, L.C.; Hsieh, C.C.; Lin, W.C. Characterization and immunomodulatory activity of rice hull polysaccharides. Carbohydr. Polym. 2015, 124, 150–156. [Google Scholar] [CrossRef]
- Nandi, I.; Ghosh, M. Studies on functional and antioxidant property of dietary fibre extracted from defatted sesame husk, rice bran and flaxseed. Bioact. Carbohydr. Diet. Fibre 2015, 5, 129–136. [Google Scholar] [CrossRef]
- Horvat, D.; Šimić, G.; Drezner, G.; Lalić, A.; Ledenčan, T.; Tucak, M.; Plavšić, H.; Andrić, L.; Zdunić, Z. Phenolic acid profiles and antioxidant activity of major cereal crops. Antioxidants 2020, 9, 527. [Google Scholar] [CrossRef]
- Saji, N.; Francis, N.; Schwarz, L.J.; Blanchard, C.L.; Santhakumar, A.B. The antioxidant and anti-inflammatory properties of rice bran phenolic extracts. Foods 2020, 9, 829. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.Y.; Han, S.N. The role of vitamin E in immunity. Nutrients 2018, 10, 1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.Y.; Lai, H.M. Bioactive compounds in rice during grain development. Food Chem. 2011, 127, 86–93. [Google Scholar] [CrossRef]
- Jadkauskaite, L.; Coulombe, P.A.; Schäfer, M.; Dinkova-Kostova, A.T.; Paus, R.; Haslam, I.S. Oxidative stress management in the hair follicle: Could targeting NRF2 counter age-related hair disorders and beyond? Bioessays 2017, 39, 1700029. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Surin, S.; Surayot, U.; Seesuriyachan, P.; You, S.; Phimolsiripol, Y. Antioxidant and immunomodulatory activities of sulphated polysaccharides from purple glutinous rice bran (Oryza sativa L.). Int. J. Food Sci. Technol. 2018, 53, 994–1004. [Google Scholar] [CrossRef] [Green Version]
- Lai, P.; Li, K.Y.; Lu, S.; Chen, H.H. Phytochemicals and antioxidant properties of solvent extracts from Japonica rice bran. Food Chem. 2009, 117, 538–544. [Google Scholar] [CrossRef]
- Surin, S.; You, S.; Seesuriyachan, P.; Muangrat, R.; Wangtueai, S.; Jambrak, A.R.; Phongthai, S.; Jantanasakulwong, K.; Chaiyaso, T.; Phimolsiripol, Y. Optimization of ultrasonic-assisted extraction of polysaccharides from purple glutinous rice bran (Oryza sativa L.) and their antioxidant activities. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Kiliç, I.; Yeşiloğlu, Y. Spectroscopic studies on the antioxidant activity of p-coumaric acid. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 115, 719–724. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative stress and inflammation: What polyphenols can do for us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Woranam, K.; Senawong, G.; Utaiwat, S.; Yunchalard, S.; Sattayasai, J.; Senawong, T. Anti-inflammatory activity of the dietary supplement Houttuynia cordata fermentation product in RAW264. 7 cells and Wistar rats. PLoS ONE 2020, 15, e0230645. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Lu, X.; Wei, T.; Dong, Y.; Cai, Z.; Tang, L.; Liu, M. Asperuloside and asperulosidic acid exert an anti-inflammatory effect via suppression of the NF-κB and MAPK signaling pathways in LPS-Induced RAW 264.7 macrophages. Int. J. Mol. Sci. 2018, 19, 2027. [Google Scholar] [CrossRef] [Green Version]
- Hämäläinen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-kappaB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007, 2007, 45673. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.J.; Choi, I.W.; Kim, G.Y.; Kim, B.W.; Park, C.; Hong, S.H.; Moon, S.K.; Cha, H.J.; Chang, Y.C.; Paek, K.Y.; et al. Anti-inflammatory potential of saponins derived from cultured wild ginseng roots in lipopolysaccharide-stimulated RAW 264.7 macrophages. Int. J. Mol. Med. 2015, 35, 1690–1698. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Zawistowski, J.; Ling, W.; Kitts, D.D. Black rice (Oryza sativa L. indica) pigmented fraction suppresses both reactive oxygen species and nitric oxide in chemical and biological model systems. J. Agric. Food Chem. 2003, 51, 5271–5277. [Google Scholar] [CrossRef]
- Jeong, C.G.; Park, M.H.; Seong, J.W.; Lee, H.S.; Park, S.K.; Kim, S.Y.; Kim, Y.B.; Jung, H.S.; Sohn, N.W.; Sohn, Y.J. Immunohistochemical study on the hair growth promoting effect of Yonnyuniksoogobon-dan. J. Korean Med. 2008, 29, 77–89. [Google Scholar]
- Shin, K.; Kim, T.S.; Kyung, J.; Kim, D.; Park, D.; Choi, E.K.; Lee, S.P.; Yang, W.S.; Kang, M.H.; Kim, Y.B. Effectiveness of the combinational treatment of Laminaria japonica and Cistanche tubulosa extracts in hair growth. Lab. Anim. Res. 2015, 31, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Zeinali, M.; Rezaee, S.A.; Hosseinzadeh, H. An overview on immunoregulatory and anti-inflammatory properties of chrysin and flavonoids substances. Biomed. Pharmacother. 2017, 92, 998–1009. [Google Scholar] [CrossRef]
- Beharka, A.A.; Wu, D.; Serafini, M.; Meydani, S.N. Mechanism of vitamin E inhibition of cyclooxygenase activity in macrophages from old mice: Role of peroxynitrite. Free Radic. Biol. Med. 2002, 32, 503–511. [Google Scholar] [CrossRef]
- Shen, J.; Yang, T.; Xu, Y.; Luo, Y.; Zhong, X.; Shi, L.; Hu, T.; Guo, T.; Nie, Y.; Luo, F.; et al. δ-Tocotrienol, isolated from rice bran, exerts an anti-inflammatory effect via MAPKs and PPARs signaling pathways in lipopolysaccharide-stimulated macrophages. Int. J. Mol. Sci. 2018, 19, 3022. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.B. Male hormone stimulation is prerequisite and an incitant in common baldness. Am. J. Anat. 1942, 71, 451–480. [Google Scholar] [CrossRef]
- Sánchez, P.; Serrano-Falcón, C.; Torres, J.M.; Serrano, S.; Ortega, E. 5α-Reductase isozymes and aromatase mRNA levels in plucked hair from young women with female pattern hair loss. Arch. Dermatol. 2018, 310, 77–83. [Google Scholar] [CrossRef]
- Sawaya, M.E.; Price, V.H. Different levels of 5α-reductase type I and II, aromatase, and androgen receptor in hair follicles of women and men with androgenetic alopecia. J. Investig. Dermatol. 1997, 109, 296–300. [Google Scholar] [CrossRef] [Green Version]
- Asada, Y.; Sonoda, T.; Ojiro, M.; Kurata, S.; Sato, T.; Ezaki, T.; Takayasu, S. 5α-Reductase type 2 is constitutively expressed in the dermal papilla and connective tissue sheath of the hair follicle in vivo but not during culture in vitro. J. Clin. Endocrinol. Metab. 2001, 86, 2875–2880. [Google Scholar] [CrossRef] [Green Version]
- Yamana, K.; Labrie, F.; Luu-The, V. Human type 3 5α-reductase is expressed in peripheral tissues at higher levels than types 1 and 2 and its activity is potently inhibited by finasteride and dutasteride. Horm. Mol. Biol. Clin. Investig. 2010, 2, 293–299. [Google Scholar] [CrossRef]
- Khantham, C.; Yooin, W.; Sringarm, K.; Sommano, S.R.; Jiranusornkul, S.; Carmona, F.D.; Nimlamool, W.; Jantrawut, P.; Rachtanapun, P.; Ruksiriwanich, W. Effects on steroid 5-alpha reductase gene expression of Thai rice bran extracts and molecular dynamics study on SRD5A2. Biology 2021, 10, 319. [Google Scholar] [CrossRef]
- Manosroi, A.; Chankhampan, C.; Kietthanakorn, B.O.; Ruksiriwanich, W.; Chaikul, P.; Boonpisuttinant, K.; Sainakham, M.; Manosroi, W.; Tangjai, T.; Manosroi, J. Pharmaceutical and cosmeceutical biological activities of hemp (Cannabis sativa L. var. sativa) leaf and seed extracts. Chiang Mai J. Sci. 2019, 46, 180–195. [Google Scholar]
- Manosroi, A.; Ruksiriwanich, W.; Manosroi, W.; Abe, M.; Manosroi, J. In vivo hair growth promotion activity of gel containing niosomes loaded with the Oryza sativa bran fraction (OSF3). Adv. Sci. Lett. 2012, 16, 222–228. [Google Scholar] [CrossRef]
- Manosroi, A.; Ruksiriwanich, W.; Abe, M.; Sakai, H.; Aburai, K.; Manosroi, W.; Manosroi, J. Physico-chemical properties of cationic niosomes loaded with fraction of rice (Oryza sativa) bran extract. J. Nanosci. Nanotechnol. 2012, 12, 7339–7345. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, A.; Chaikul, P.; Chankhampan, C.; Ruksiriwanich, W.; Manosroi, W.; Manosroi, J. 5α-reductase inhibition and melanogenesis induction of the selected Thai plant extracts. Chiang Mai J. Sci. 2018, 45, 220–236. [Google Scholar]
- Ruksiriwanich, W.; Manosroi, J.; Abe, M.; Manosroi, W.; Manosroi, A. 5α-Reductase type 1 inhibition of Oryza sativa bran extract prepared by supercritical carbon dioxide fluid. J. Supercrit. Fluids 2011, 59, 61–71. [Google Scholar] [CrossRef]
- Akbaba, H.; Akbaba, G.E.; Kantarcı, A.G. Development and evaluation of antisense shRNA-encoding plasmid loaded solid lipid nanoparticles against 5-α reductase activity. J. Drug Deliv. Sci. Techno. 2018, 44, 270–277. [Google Scholar] [CrossRef]
- Normington, K.; Russell, D.W. Tissue distribution and kinetic characteristics of rat steroid 5 alpha-reductase isozymes. evidence for distinct physiological functions. J. Biol. Chem. 1992, 267, 19548–19554. [Google Scholar] [CrossRef]
- Liang, T.; Liao, S. Inhibition of steroid 5α-reductase by specific aliphatic unsaturated fatty acids. Biochem. J. 1992, 285, 557–562. [Google Scholar] [CrossRef]
- Hiipakka, R.A.; Zhang, H.Z.; Dai, W.; Dai, Q.; Liao, S. Structure–activity relationships for inhibition of human 5α-reductases by polyphenols. Biochem. Pharmacol. 2002, 63, 1165–1176. [Google Scholar] [CrossRef]
- Young, J.W.; Conte, E.T.; Leavitt, M.L.; Nafz, M.A.; Schroeter, A.L. Cutaneous immunopathology of androgenetic alopecia. J. Am. Osteopath. Assoc. 1991, 90, 765–771. [Google Scholar] [CrossRef]
- Chen, C.; Wang, L.; Wang, R.; Luo, X.; Li, Y.; Li, J.; Li, Y.; Chen, Z. Phenolic contents, cellular antioxidant activity and antiproliferative capacity of different varieties of oats. Food Chem. 2018, 239, 260–267. [Google Scholar] [CrossRef]
- Diniyah, N.; Badrul Alam, M.; Lee, S.H. Antioxidant potential of non-oil seed legumes of Indonesian’s ethnobotanical extracts. Arab. J. Chem. 2020, 13, 5208–5217. [Google Scholar] [CrossRef]
- Luo, D.; Qu, C.; Lin, G.; Zhang, Z.; Xie, J.; Chen, H.; Liang, J.; Li, C.; Wang, H.; Su, Z. Character and laxative activity of polysaccharides isolated from Dendrobium officinale. J. Funct. Foods 2017, 34, 106–117. [Google Scholar] [CrossRef]
- Abbas, Z.; Manoharan, A.L.; Jagadeesan, G.; Nataraj, G.; Muniyandi, K.; Sathyanarayanan, S.; Thangaraj, P. Evaluation of an edible polyherbal formulation against urinary tract infection pathogens, its antioxidant and anti-inflammatory potential. Biocatal. Agric. Biotechnol. 2021, 35, 102104. [Google Scholar] [CrossRef]
- Mighri, H.; Akrout, A.; Bennour, N.; Eljeni, H.; Zammouri, T.; Neffati, M. LC/MS method development for the determination of the phenolic compounds of Tunisian Ephedra alata hydro-methanolic extract and its fractions and evaluation of their antioxidant activities. S. Afr. J. Bot. 2019, 124, 102–110. [Google Scholar] [CrossRef]
- Arribas, C.; Pereira, E.; Barros, L.; Alves, M.J.; Calhelha, R.C.; Guillamón, E.; Pedrosa, M.M.; Ferreira, I.C. Healthy novel gluten-free formulations based on beans, carob fruit and rice: Extrusion effect on organic acids, tocopherols, phenolic compounds and bioactivity. Food Chem. 2019, 292, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Kramer, J.K.; Blais, L.; Fouchard, R.C.; Melnyk, R.A.; Kallury, K.M. A rapid method for the determination of vitamin E forms in tissues and diet by high-performance liquid chromatography using a normal-phase diol column. Lipids 1997, 32, 323–330. [Google Scholar] [CrossRef]
- Manosroi, A.; Lohcharoenkal, W.; Ruksiriwanich, W.; Kietthanakorn, B.; Manosroi, W.; Manosroi, J. In vitro immunostimulating activity of the dried sap from fermented Thai rice on human and murine neutrophils. Adv. Sci. Lett. 2012, 17, 306–311. [Google Scholar] [CrossRef]
- Ruksiriwanich, W.; Khantham, C.; Linsaenkart, P.; Jantrawut, P.; Rajchasom, S. Optimization of placenta extraction for wound healing activity. Chiang Mai J. Sci. 2019, 46, 946–959. [Google Scholar]
- Grochowski, D.M.; Uysal, S.; Aktumsek, A.; Granica, S.; Zengin, G.; Ceylan, R.; Locatelli, M.; Tomczyk, M. In vitro enzyme inhibitory properties, antioxidant activities, and phytochemical profile of Potentilla thuringiaca. Phytochem. Lett. 2017, 20, 365–372. [Google Scholar] [CrossRef]
- Ruksiriwanich, W.; Khantham, C.; Sringarm, K.; Sommano, S.; Jantrawut, P. Depigmented Centella asiatica extraction by pretreated with supercritical carbon dioxide fluid for wound healing application. Processes 2020, 8, 277. [Google Scholar] [CrossRef] [Green Version]
- Ruksiriwanich, W.; Khantham, C.; Linsaenkart, P.; Chaitep, T.; Rachtanapun, P.; Jantanasakulwong, K.; Phimolsiripol, Y.; Režek Jambrak, A.; Nazir, Y.; Yooin, W. Anti-inflammation of bioactive compounds from ethanolic extracts of edible bamboo mushroom (Dictyophora indusiata) as functional health promoting food ingredients. Int. J. Food Sci. Technol. 2022, 57, 110–122. [Google Scholar] [CrossRef]
- Smith, C.M.; Ballard, S.A.; Worman, N.; Buettner, R.; Masters, J.R. 5 alpha-reductase expression by prostate cancer cell lines and benign prostatic hyperplasia in vitro. J. Clin. Endocrinol. Metab. 1996, 81, 1361–1366. [Google Scholar] [CrossRef]
- Kim, D.; Cho, H.; Eggers, R.; Kim, S.K.; Ryu, C.S.; Kim, Y.J. Development of a liquid chromatography/mass spectrometry-based inhibition assay for the screening of steroid 5-α reductase in human and fish cell lines. Molecules 2021, 26, 893. [Google Scholar] [CrossRef]
- Wang, F. Semi-quantitative RT-PCR: An effective method to explore the regulation of gene transcription level affected by environmental pollutants. In Environmental Toxicology and Toxicogenomics: Principles, Methods and Applications; Pan, X., Zhang, B., Eds.; Springer: New York, NY, USA, 2021; pp. 95–103. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Phenolic Content (mg GAE/g Extract) | Total Flavonoid Content (mg EGCGE/g Extract) | Total Polysaccharide Content (mg D-glucose/g Extract) | Total Protein Content (mg BSAE/g Extract) |
|---|---|---|---|---|
| BB3CMU-H | 23.68 ± 0.56 a | 82.57 ± 9.89 | 0.22 ± 0.01 a | 0.40 ± 0.01 a |
| BB3CMU-RB | 11.63 ± 0.40 b | 81.64 ± 4.26 | 0.05 ± 0.01 b | 0.16 ± 0.01 b |
| Bioactive Compound | Molecular Formula | BB3CMU-H | BB3CMU-RB |
|---|---|---|---|
| (μg/g Extract) | (μg/g Extract) | ||
| Caffeic acid | C9H8O4 | 105.5 ± 0.6 | 40.18 ± 0.01 |
| Catechin | C15H14O6 | 295.4 ± 0.4 | ND |
| Chlorogenic acid | C16H18O9 | 204.7 ± 3.7 | 113.8 ± 0.01 |
| o-Coumaric acid | C9H8O3 | 910.1 ± 1.0 | 78.92 ± 0.01 |
| p-Coumaric acid | C9H8O3 | 76.65 ± 3.90 | 99.80 ± 0.08 |
| Epigallocatechin gallate | C22H18O11 | 163.6 ± 3.1 | 174.6 ± 0.01 |
| Ferulic acid | C10H10O4 | 157.4 ± 5.4 | 22.87 ± 0.01 |
| Gallic acid | C7H6O5 | ND | 16.84 ± 0.01 |
| p-Hydroxybenzoic acid | C7H6O3 | 198.7 ± 2.8 | ND |
| Kaempferol | C15H10O6 | 863.6 ± 2.1 | 34.23 ± 0.01 |
| Naringin | C27H32O14 | 68.13 ± 1.74 | 113.2 ± 0.01 |
| Phytic acid | C6H18O24P6 | 7358 ± 6.3 | 428.6 ± 0.01 |
| Quercetin | C15H10O7 | 120.2 ± 0.3 | 34.27 ± 0.01 |
| Rosmarinic acid | C18H16O8 | 51.43 ± 0.38 | ND |
| Rutin | C27H30O16 | 150.9 ± 0.04 | ND |
| α-tocopherol | C29H50O2 | ND | 854.3 ± 0.8 |
| β-tocopherol | C28H48O2 | ND | 62.60 ± 0.20 |
| γ-tocopherol | C28H48O2 | ND | 367.2 ± 1.3 |
| δ-tocopherol | C27H46O2 | ND | 11.80 ± 0.40 |
| Sample | DPPH Assay (mg TE/g Extract) | ABTS Assay (mg TE/g Extract) | Metal Chelation (mg EDTAE/g Extract) |
|---|---|---|---|
| BB3CMU-H | 14.64 ± 1.30 a | 19.94 ± 0.65 a | 6.85 ± 4.53 a |
| BB3CMU-RB | 8.61 ± 0.01 b | 9.16 ± 0.19 b | 90.92 ± 4.11 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khantham, C.; Linsaenkart, P.; Chaitep, T.; Jantrawut, P.; Chittasupho, C.; Rachtanapun, P.; Jantanasakulwong, K.; Phimolsiripol, Y.; Sommano, S.R.; Prom-u-thai, C.; et al. Antioxidation, Anti-Inflammation, and Regulation of SRD5A Gene Expression of Oryza sativa cv. Bue Bang 3 CMU Husk and Bran Extracts as Androgenetic Alopecia Molecular Treatment Substances. Plants 2022, 11, 330. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11030330
Khantham C, Linsaenkart P, Chaitep T, Jantrawut P, Chittasupho C, Rachtanapun P, Jantanasakulwong K, Phimolsiripol Y, Sommano SR, Prom-u-thai C, et al. Antioxidation, Anti-Inflammation, and Regulation of SRD5A Gene Expression of Oryza sativa cv. Bue Bang 3 CMU Husk and Bran Extracts as Androgenetic Alopecia Molecular Treatment Substances. Plants. 2022; 11(3):330. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11030330
Chicago/Turabian StyleKhantham, Chiranan, Pichchapa Linsaenkart, Tanakarn Chaitep, Pensak Jantrawut, Chuda Chittasupho, Pornchai Rachtanapun, Kittisak Jantanasakulwong, Yuthana Phimolsiripol, Sarana Rose Sommano, Chanakan Prom-u-thai, and et al. 2022. "Antioxidation, Anti-Inflammation, and Regulation of SRD5A Gene Expression of Oryza sativa cv. Bue Bang 3 CMU Husk and Bran Extracts as Androgenetic Alopecia Molecular Treatment Substances" Plants 11, no. 3: 330. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11030330