
Phytochemical Profiling and Assessment of Anticancer Activity of Leptocarpha rivularis Extracts Obtained from In Vitro Cultures

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. In Vitro Micropropagation of Leptocarpha Rivularis
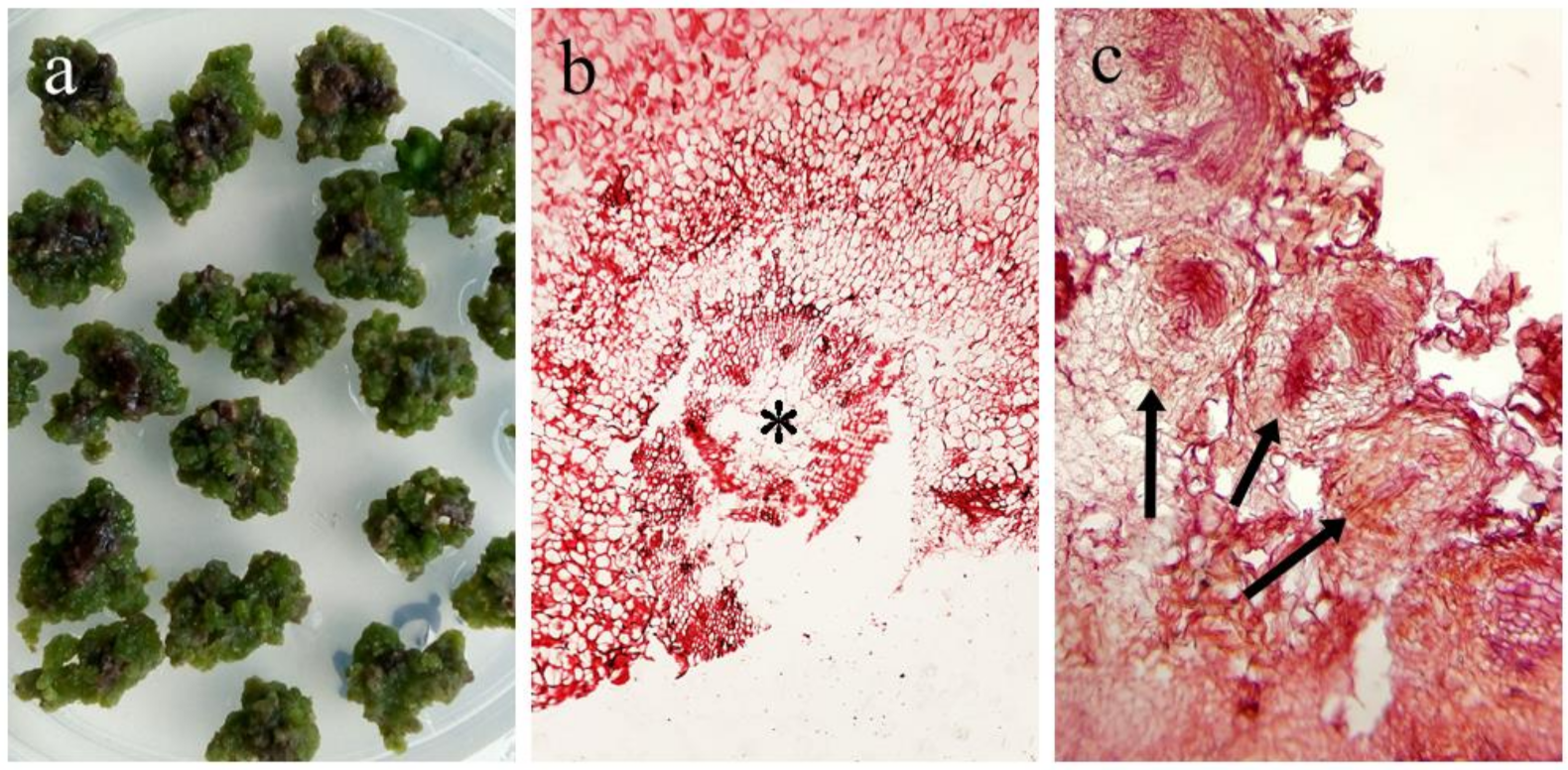
2.2. Callus Induction
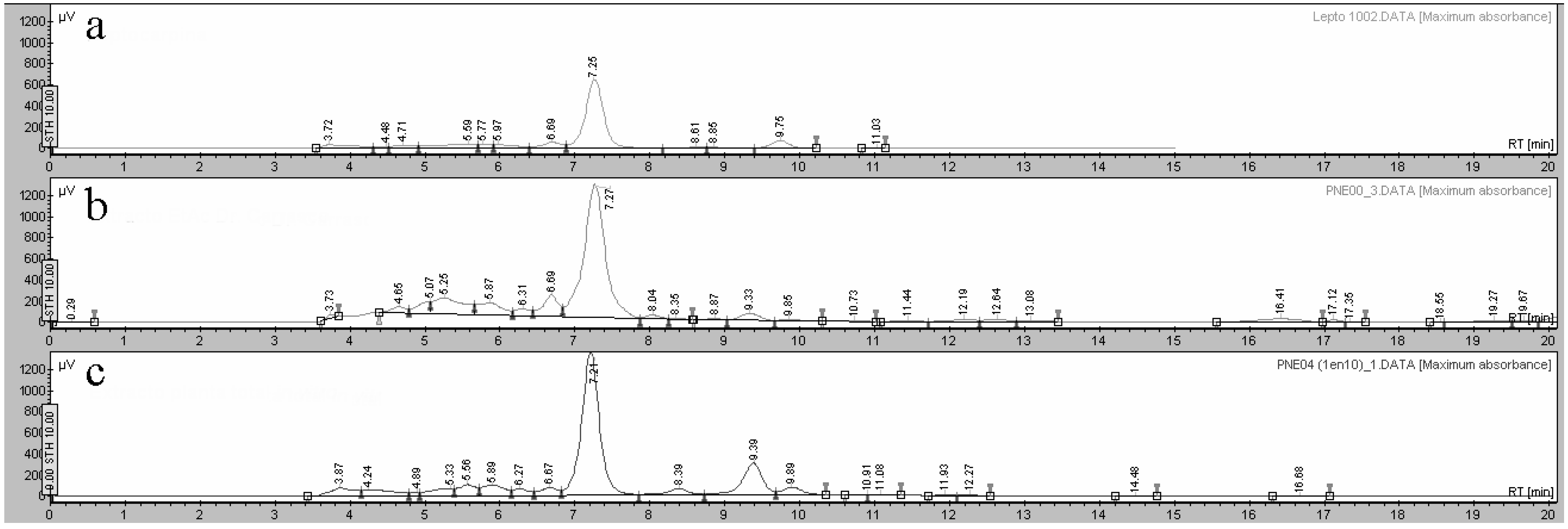
2.3. Phytochemical Analysis of Extracts
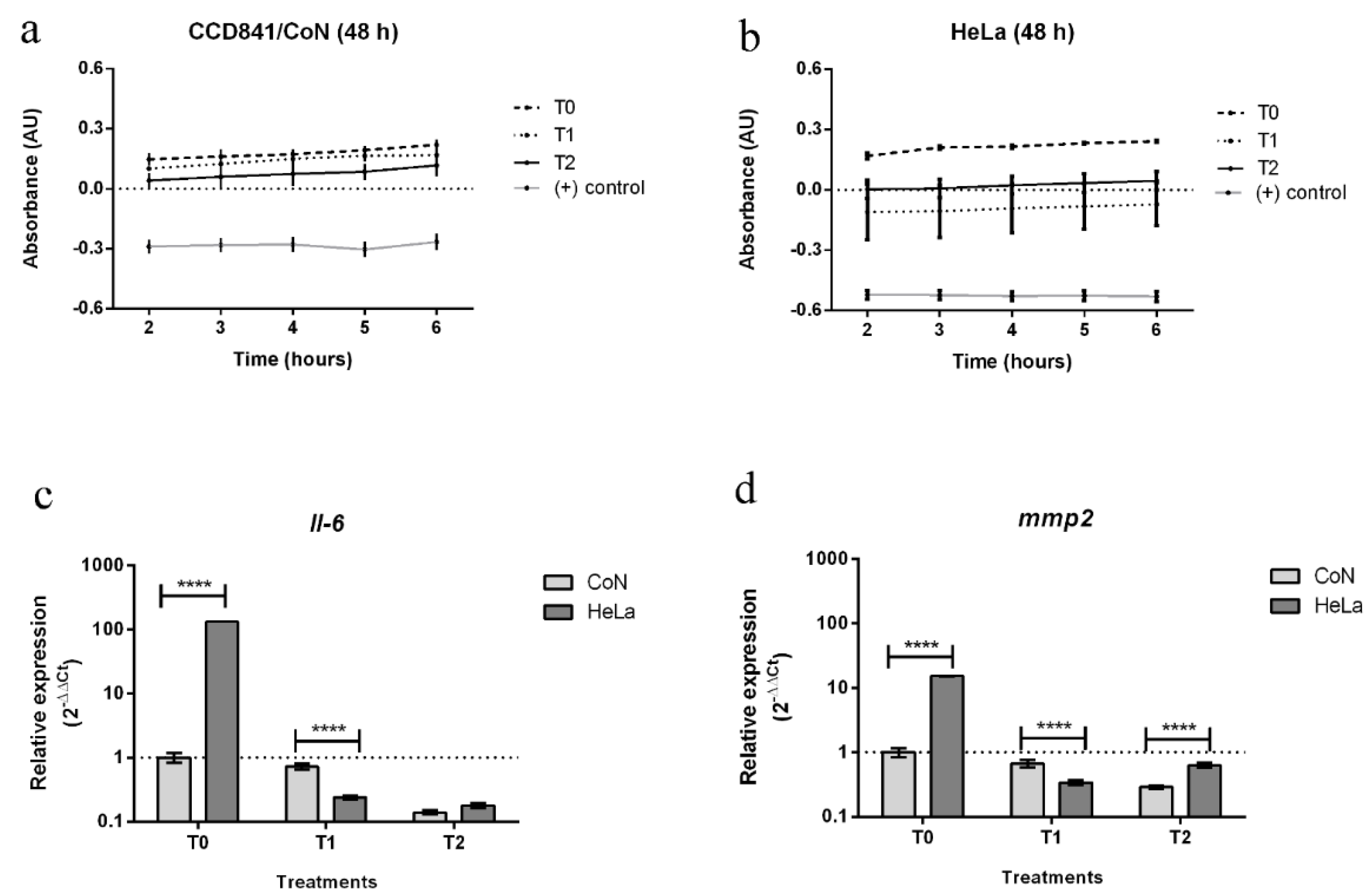
2.4. Asseessment of Biological Activity of In Vitro Plant Extracts
3. Materials and Methods
3.1. In Vitro Micropropagation Platform for Leptocarpha Rivularis
3.2. Callus Induction of Leptocarpha Rivularis
3.3. Extraction and Composition Analysis of Leptocarpha Rivularis Extracts
3.4. Biological Activity of the Total Extracts of Leptocarpha Rivularis In Vitro
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuloaga, F.; Belgrano, M. The Catalogue of Vascular Plants of the Southern Cone and the Flora of Argentina: Their contribution to the World Flora. Rodriguésia 2015, 66, 989–1024. [Google Scholar] [CrossRef] [Green Version]
- Obando, M. Scientific evidence of the medicinal properties of plants for use in intercultural medicine in Chile. Bol. Latinoam. Caribe Plantas Med. Aromat. 2020, 19, 207. [Google Scholar]
- Uquiche, E.; Garcés, F. Recovery and antioxidant activity of extracts from leptocarpha rivularis by supercritical carbon dioxide extraction. J. Supercrit. Fluids 2016, 110, 257–264. [Google Scholar] [CrossRef]
- Jimenez-Gonzalez, A.; Quispe, C.; Borquez, J.; Sepulveda, B.; Riveros, F.; Areche, C.; Nagles, E.; Garcia-Beltran, O.; Simirgiotis, M.J. UHPLC-ESI-ORBITRAP-MS analysis of the native Mapuche medicinal plant palo negro (Leptocarpha rivularis DC.-Asteraceae) and evaluation of its antioxidant and cholinesterase inhibitory properties. J. Enzyme Inhib. Med. Chem. 2018, 33, 936–944. [Google Scholar] [CrossRef]
- Montenegro, I.; Moreira, J.; Ramirez, I.; Dorta, F.; Sanchez, E.; Alfaro, J.F.; Valenzuela, M.; Jara-Gutierrez, C.; Munoz, O.; Alvear, M.; et al. Chemical Composition, Antioxidant and Anticancer Activities of Leptocarpha rivularis DC Flower Extracts. Molecules 2021, 26, 67. [Google Scholar] [CrossRef] [PubMed]
- Olea, A.F.; Villena, J.; Moller, A.; Martinez, R.; Carrasco, H. Enhancement of cytotoxic activity by encapsulation in pluronic polymer micelles: Leptocarpha rivularis extracts against human cancer cell lines. J. Chil. Chem. Soc. 2019, 64, 4437–4440. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Contreras, C.; Martinez, R.; Cubillos, V. Efecto Hipoglicemiante de la Infusión de Leptocarpha rivularis: Estudio en Ratas Diabéticas Tipo II por Inducción con Aloxano; Editorial Académica Española: Chisinau, Moldava, 2018. [Google Scholar]
- Martinez, R.; Kesternich, V.; Gutierrez, E.; Dolz, H.; Mansilla, H. Conformational-Analysis and biological activity of leptocarpin and leptocarpin acetate. Planta Med. 1995, 61, 188–189. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.; Kesternich, V.; Carrasco, H.; Alvarez-Contreras, C.; Montenegro, C.; Ugarte, R.; Gutierrez, E.; Moreno, J.; Garcia, C.; Werner, E.; et al. Synthesis and conformational analysis of leptocarpin derivatives. Influence of modification of the oxirane ring on leptocarpin’s cytotoxic activity. J. Chil. Chem. Soc. 2006, 51, 1010–1014. [Google Scholar] [CrossRef]
- Bosio, C.; Tomasoni, G.; Martinez, R.; Olea, A.F.; Carrasco, H.; Villena, J. Cytotoxic and apoptotic effects of leptocarpin, a plant-derived sesquiterpene lactone, on human cancer cell lines. Chem. Biol. Inter. 2015, 242, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.W.; Wan, W.G.; Miao, H.X.; Tang, R.; Wang, B.; Huang, Q.Z.; Liu, W.L.; Zheng, J.P.; Chen, C.Q.; Zhong, H.B.; et al. Leptocarpin Suppresses Proliferation, Migration, and Invasion of Human Osteosarcoma by Targeting Type-1 Insulin-Like Growth Factor Receptor (IGF-1R). Med. Sci. Monit. 2017, 23, 4132–4140. [Google Scholar] [CrossRef] [Green Version]
- Cordero, C. Propagación Vegetativa de Leptocarpha rivularis DC, Especie Nativa de Uso Medicinal. Bachelor’s Thesis, Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile, 2000. [Google Scholar]
- Dorta, F.; Polanco, V.; Ramirez, I.; Seeger, M.; Madrid, A.; Montenegro, I. In vitro propagation of Leptocarpha rivularis, a native medicinal plant. In Vitro Cell. Dev. Biol. Plant 2020, 56, 827–832. [Google Scholar] [CrossRef]
- Bairu, M.W.; Amoo, S.O.; Van Staden, J. Comparative phytochemical analysis of wild and in vitro-derived greenhouse-grown tubers, in vitro shoots and callus-like basal tissues of Harpagophytum procumbens. S. Afr. J. Bot. 2011, 77, 479–484. [Google Scholar] [CrossRef] [Green Version]
- Mangoale, R.M.; Afolayan, A.J. Comparative Phytochemical Constituents and Antioxidant Activity of Wild and Cultivated Alepidea amatymbica Eckl & Zeyh. Biomed. Res. Int. 2020, 2020, 5808624. [Google Scholar] [PubMed]
- Mulabagal, V.; Tsay, H.-S. Plant Cell Cultures-An Alternative and Efficient Source for the Production of Biologically Important Secondary Metabolites. Int. J. Appl. Sci. Eng. 2004, 2, 29–48. [Google Scholar]
- Ochoa-Villarreal, M.; Howat, S.; Hong, S.; Jang, M.O.; Jin, Y.-W.; Lee, E.-K.; Loake, G.J. Plant cell culture strategies for the production of natural products. BMB Rep. 2016, 49, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Bhojwani, S.S.; Dantu, P.K. Production of Industrial Phytochemicals. In Plant Tissue Culture: An Introductory Text; Springer: New Delhi, India, 2013; pp. 275–286. [Google Scholar]
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; Garcia-Lara, S. In vitro plant tissue culture: Means for production of biological active compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T. Biotechnology Applications of Plant Callus Cultures. Engineering 2019, 5, 50–59. [Google Scholar] [CrossRef]
- Benitez-Garcia, I.; Vanegas-Espinoza, P.E.; Melendez-Martinez, A.J.; Heredia, F.J.; Paredes-Lopez, O.; Del Villar-Martinez, A.A. Callus culture development of two varieties of Tagetes erecta and carotenoid production. Electron. J. Biotechnol. 2014, 17, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, E.; Temporiti, M.E.E.; Cella, R. Improvement of phytochemical production by plant cells and organ culture and by genetic engineering. Plant Cell Rep. 2019, 38, 1199–1215. [Google Scholar] [CrossRef]
- Mukhia, S.; Mandal, P.; Singh, D.K.; Singh, D. Comparison of pharmacological properties and phytochemical constituents of in vitro propagated and naturally occurring liverwort Lunularia cruciata. BMC Complement. Altern. Med. 2019, 19, 181. [Google Scholar] [CrossRef] [Green Version]
- Voronova, O. Callus formation and plant regeneration in sunflower (Helianthus L., asteraceae) in vitro tissue culture. In Proceedings of the 19th International Sunflower Conference, Edirne, Turkey, 29 May–3 June 2016; p. 211. [Google Scholar]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant Callus: Mechanisms of Induction and Repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, K.; Meyerowitz, E.M. Regeneration in Arabidopsis tissue culture. Methods Mol. Biol. 2013, 959, 265–275. [Google Scholar]
- Osman, N.I.; Jaafar Sidik, N.; Awal, A. Effects of variations in culture media and hormonal treatments upon callus induction potential in endosperm explant of Barringtonia racemosa L. Asian Pac. J. Trop. Biomed. 2016, 6, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Feher, A. Callus, Dedifferentiation, Totipotency, Somatic Embryogenesis: What These Terms Mean in the Era of Molecular Plant Biology? Front Plant. Sci. 2019, 10, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S. Chapter 2-Plant tissue culture. In Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Bhatia, S., Sharma, K., Dahiya, R., Bera, T., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 31–107. [Google Scholar]
- Martinez, R.; Ayamante, B.; Nunez-Alarcon, J.A.; de Vivar, A.R. Leptocarpin and 17,18-dihydroleptocarpin, two new heliangolides from Leptocarpha rivularis. Phytochemistry 1979, 18, 1527–1528. [Google Scholar] [CrossRef]
- Martinez, R.; Kesternich, V.; Carrasco, H.; Bustos, C.; Fernandez, S. Structure, conformation and biological activity studies on rivularin, a new heliangolide isolated from Leptocarpha rivularis. Bol. Soc. Chil. Quim. 1998, 43, 7–12. [Google Scholar]
- Präbst, K.; Engelhardt, H.; Ringgeler, S.; Hübner, H. Basic Colorimetric Proliferation Assays: MTT, WST, and Resazurin. Methods Mol. Biol. 2017, 1601, 1–17. [Google Scholar] [PubMed]
- Rampersad, S.N. Multiple applications of Alamar Blue as an indicator of metabolic function and cellular health in cell viability bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef] [PubMed]
- Badache, A.; Hynes, N.E. Interleukin 6 inhibits proliferation and, in cooperation with an epidermal growth factor receptor autocrine loop, increases migration of T47D breast cancer cells. Cancer Res. 2001, 61, 383–391. [Google Scholar]
- Paduch, R.; Walter-Croneck, A.; Zdzisińska, B.; Szuster-Ciesielska, A.; Kandefer-Szerszeń, M. Role of reactive oxygen species (ROS), metalloproteinase-2 (MMP-2) and interleukin-6 (IL-6) in direct interactions between tumour cell spheroids and endothelial cell monolayer. Cell Biol. Int. 2005, 29, 497–505. [Google Scholar] [CrossRef]
- Roomi, M.W.; Monterrey, J.C.; Kalinovsky, T.; Rath, M.; Niedzwiecki, A. In vitro modulation of MMP-2 and MMP-9 in human cervical and ovarian cancer cell lines by cytokines, inducers and inhibitors. Oncol. Rep. 2010, 23, 605–614. [Google Scholar] [PubMed]
- Luo, Y.; Zheng, S.G. Hall of Fame among Pro-inflammatory Cytokines: Interleukin-6 Gene and Its Transcriptional Regulation Mechanisms. Front. Immunol. 2016, 7, 604. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Cheng, F.W.; Wang, F.; Jia, B.; Luo, X.; Zhang, S.Q. The activation of TLR7 regulates the expression of VEGF, TIMP1, MMP2, IL-6, and IL-15 in Hela cells. Mol. Cell Biochem. 2014, 389, 43–49. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, R.; Cheng, L.; Xu, H. Celastrol inhibit the proliferation, invasion and migration of human cervical HeLa cancer cells through down-regulation of MMP-2 and MMP-9. J. Cell Mol. Med. 2021, 25, 5335–5338. [Google Scholar] [CrossRef]
- Suradej, B.; Sookkhee, S.; Panyakaew, J.; Mungkornasawakul, P.; Wikan, N.; Smith, D.R.; Potikanond, S.; Nimlamool, W. Kaempferia parviflora Extract Inhibits STAT3 Activation and Interleukin-6 Production in HeLa Cervical Cancer Cells. Int. J. Mol. Sci. 2019, 20, 4226. [Google Scholar] [CrossRef] [Green Version]
- Shih, Y.W.; Wu, P.F.; Lee, Y.C.; Shi, M.D.; Chiang, T.A. Myricetin suppresses invasion and migration of human lung adenocarcinoma A549 cells: Possible mediation by blocking the ERK signaling pathway. J. Agric. Food Chem. 2009, 57, 3490–3499. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Luo, X.; Chen, T.; Guo, W.; Liang, C.; Tang, S.; Mo, J. Naringenin inhibits migration, invasion, induces apoptosis in human lung cancer cells and arrests tumour progression in vitro. J. Cell Mol. Med. 2021, 25, 2563–2571. [Google Scholar] [CrossRef]
- Yoon, S.; Woo, S.U.; Kang, J.H.; Kim, K.; Shin, H.J.; Gwak, H.S.; Park, S.; Chwae, Y.J. NF-κB and STAT3 cooperatively induce IL6 in starved cancer cells. Oncogene 2012, 31, 3467–3481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarland, B.C.; Hong, S.W.; Rajbhandari, R.; Twitty, G.B., Jr.; Gray, G.K.; Yu, H.; Benveniste, E.N.; Nozell, S.E. NF-κB-induced IL-6 ensures STAT3 activation and tumor aggressiveness in glioblastoma. PLoS ONE 2013, 8, e78728. [Google Scholar] [CrossRef]
- Morgan, E.L.; Macdonald, A. Autocrine STAT3 activation in HPV positive cervical cancer through a virus-driven Rac1-NFκB-IL-6 signalling axis. PLoS Pathog. 2019, 15, e1007835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasempour, H.; Soheilikhah, Z.; Zebarjadi, Z.; Ghasempour, S.; Karimi, N. In vitro micro propagation, callus induction and shoot regeneration in safflower L. cv. Lesaf. Iran. J. Plant Physiol. 2014, 4, 999–1004. [Google Scholar]
- Soukup, A.; Tylová, E. Essential methods of plant sample preparation for light microscopy. In Plant Cell Morphogenesis. Methods in Molecular Biology; Cvrčková, F., Žárský, V., Eds.; Humana Press: Totowa, NY, USA, 2019; Volume 1080, pp. 1–23. [Google Scholar]
- Johansen, D.A. Plant Microtechnique. Nature 1941, 147, 222. [Google Scholar]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO induces drastic changes in human cellular processes and epigenetic landscape in vitro. Sci. Rep. 2019, 9, 4641. [Google Scholar] [CrossRef] [Green Version]
- Partridge, J.J.; Madsen, M.A.; Ardi, V.C.; Papagiannakopoulos, T.; Kupriyanova, T.A.; Quigley, J.P.; Deryugina, E.I. Functional analysis of matrix metalloproteinases and tissue inhibitors of metalloproteinases differentially expressed by variants of human HT-1080 fibrosarcoma exhibiting high and low levels of intravasation and metastasis. J. Biol. Chem. 2007, 282, 35964–35977. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Folco, E.J.; Shimizu, K.; Libby, P. Adiponectin induces pro-inflammatory programs in human macrophages and CD4+ T cells. J. Biol. Chem. 2012, 287, 36896–36904. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Jiménez, T.; Millán-Pérez Peña, L.; Flores-Mendoza, L.; Sedeño-Monge, V.; Santos-López, G.; Rosas-Murrieta, N.; Reyes-Carmona, S.; Terán-Cabanillas, E.; Hernández, J.; Herrera-Camacho, I.; et al. Upregulation of the Suppressors of Cytokine Signaling 1 and 3 Is Associated with Arrest of Phosphorylated-STAT1 Nuclear Importation and Reduced Innate Response in Denguevirus-Infected Macrophages. Viral Immunol. 2016, 29, 95–104. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Introduction (I) | Propagation (P) | Rooting (R) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| I1 | I2 | I3 | I4 | I5 * | P1 * | P2 | P3 | R1 | ||
| Hormones (µM) | BAP | 2.20 | 0.44 | 0.44 | 3.50 | 4.40 | 4.40 | 4.40 | 4.40 | 3.50 |
| IBA | 0.49 | 0.49 | 0.49 | 0.49 | 0.49 | 0.49 | 0.49 | |||
| NAA | 0.27 | |||||||||
| GA3 | 0.14 | 0.14 | 0.14 | 0.14 | 0.14 | |||||
| Hormones | ||||||||
|---|---|---|---|---|---|---|---|---|
| Induction Media | Ref | Auxins | Conc. (µM) | Cytokines | Conc. (µM) | CIF (%) * (Mean ± S.E.) | Plant Organ | Callus Charact † |
| C0 | 0 | leaf, intermodal section | n.c.i. | |||||
| C1 | NAA | 2.7 | TDZ | 4.5 | 0 | leaf | n.c.i. | |
| C2 | NAA | 2.7 | KIN | 4.6 | 0 | leaf | n.c.i. | |
| C3 | NAA | 2.7 | KIN | 2.3 | 33.3 ± 1.2 | leaf | compact, rooting | |
| C4 | NAA | 5.5 | KIN | 2.3 | 0 | leaf | n.c.i. | |
| C5 | NAA | 5.5 | KIN | 2.3 | 0 | leaf | n.c.i. | |
| C6 | [24] | NAA | 5.5 | BAP | 4.4 | 0 | root | n.c.i. |
| C6 | [24] | NAA | 5.5 | BAP | 4.4 | 36.1 ± 2.6 | leaf | compact, greenish |
| C7 | NAA | 2.7 | BAP | 4.4 | 94.4 ± 1.5 | internodal section | friable, green | |
| C8 | NAA | 5.5 | BAP | 4.4 | 91.8 ± 1.3 | internodal section | friable, green | |
| C9 | [21] | 2,4D | 4.9 | BAP | 4.4 | 0 | leaf | n.c.i. |
| C10 | [21] | 2,4D | 9.8 | BAP | 8.8 | 27.8 ± 3.2 | leaf, root | compact, brown |
| C11 | [21] | 2,4D | 13.5 | BAP | 8.8 | 0 | leaf | n.c.i. |
| C12 | BAP-KIN | 0 | leaf, intermodal section | n.c.i. | ||||
| C13 | BAP-Z | 2.7 ± 0.6 | leaf | compact, rooting | ||||
| C14 | 2ip | 0 | leaf | n.c.i. | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio, J.; Arias, G.; Robles-Kelly, C.; Silva-Moreno, E.; Espinoza, L.; Carrasco, H.; Olea, A.F. Phytochemical Profiling and Assessment of Anticancer Activity of Leptocarpha rivularis Extracts Obtained from In Vitro Cultures. Plants 2022, 11, 546. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11040546
Rubio J, Arias G, Robles-Kelly C, Silva-Moreno E, Espinoza L, Carrasco H, Olea AF. Phytochemical Profiling and Assessment of Anticancer Activity of Leptocarpha rivularis Extracts Obtained from In Vitro Cultures. Plants. 2022; 11(4):546. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11040546
Chicago/Turabian StyleRubio, Julia, Guisselle Arias, Christian Robles-Kelly, Evelyn Silva-Moreno, Luis Espinoza, Héctor Carrasco, and Andrés F. Olea. 2022. "Phytochemical Profiling and Assessment of Anticancer Activity of Leptocarpha rivularis Extracts Obtained from In Vitro Cultures" Plants 11, no. 4: 546. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11040546