
Screening for Resistance in Selected Tomato Varieties against the Root-Knot Nematode Meloidogyne incognita in the Philippines Using a Molecular Marker and Biochemical Analysis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Nematode Cultures
2.2. Nematode Extraction and Inoculum Standardization
2.3. Preparation of Planting Materials for Resistance Screening
2.4. Resistance Screening Using a Bioassay Method
2.5. Statistical Analysis
2.6. Marker Analysis Using Mi23
2.7. Biochemical Analysis
3. Results
3.1. Growth and Disease Reaction of Tomato Varieties Infected with Meloidogyne incognita
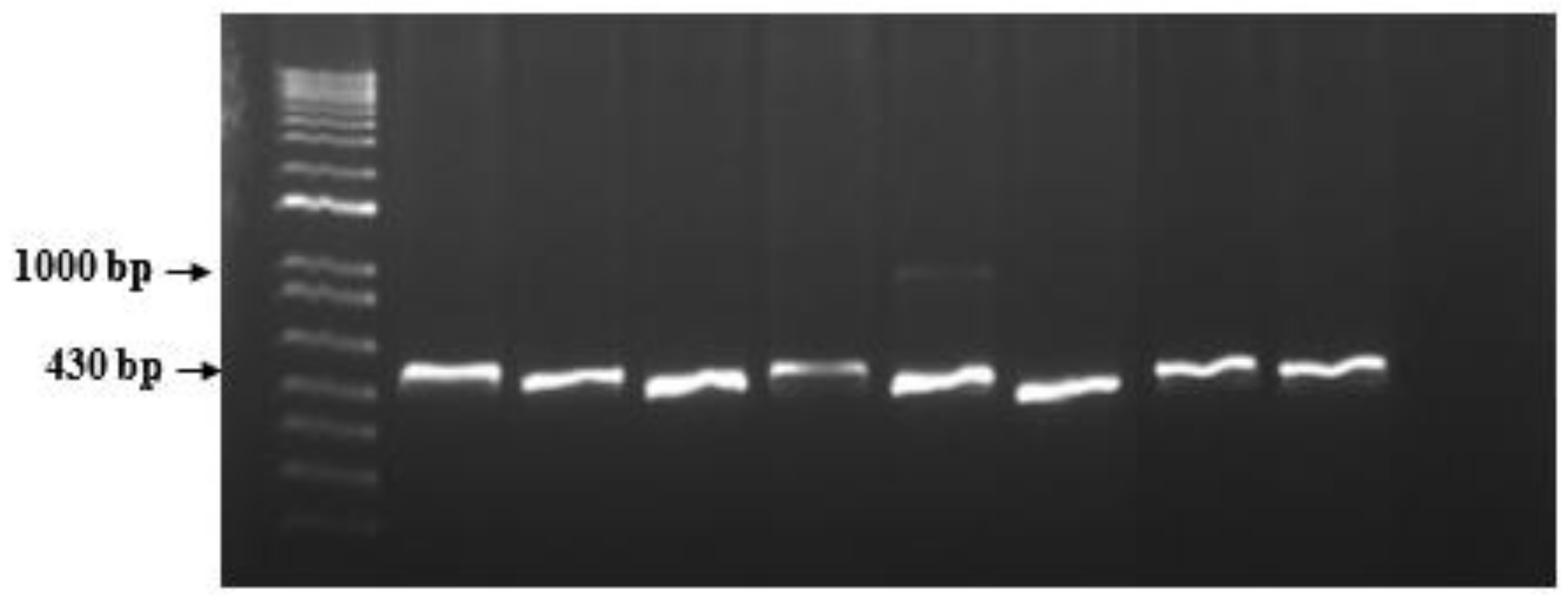
3.2. Mi23 Marker and Phylogenetic Analyses
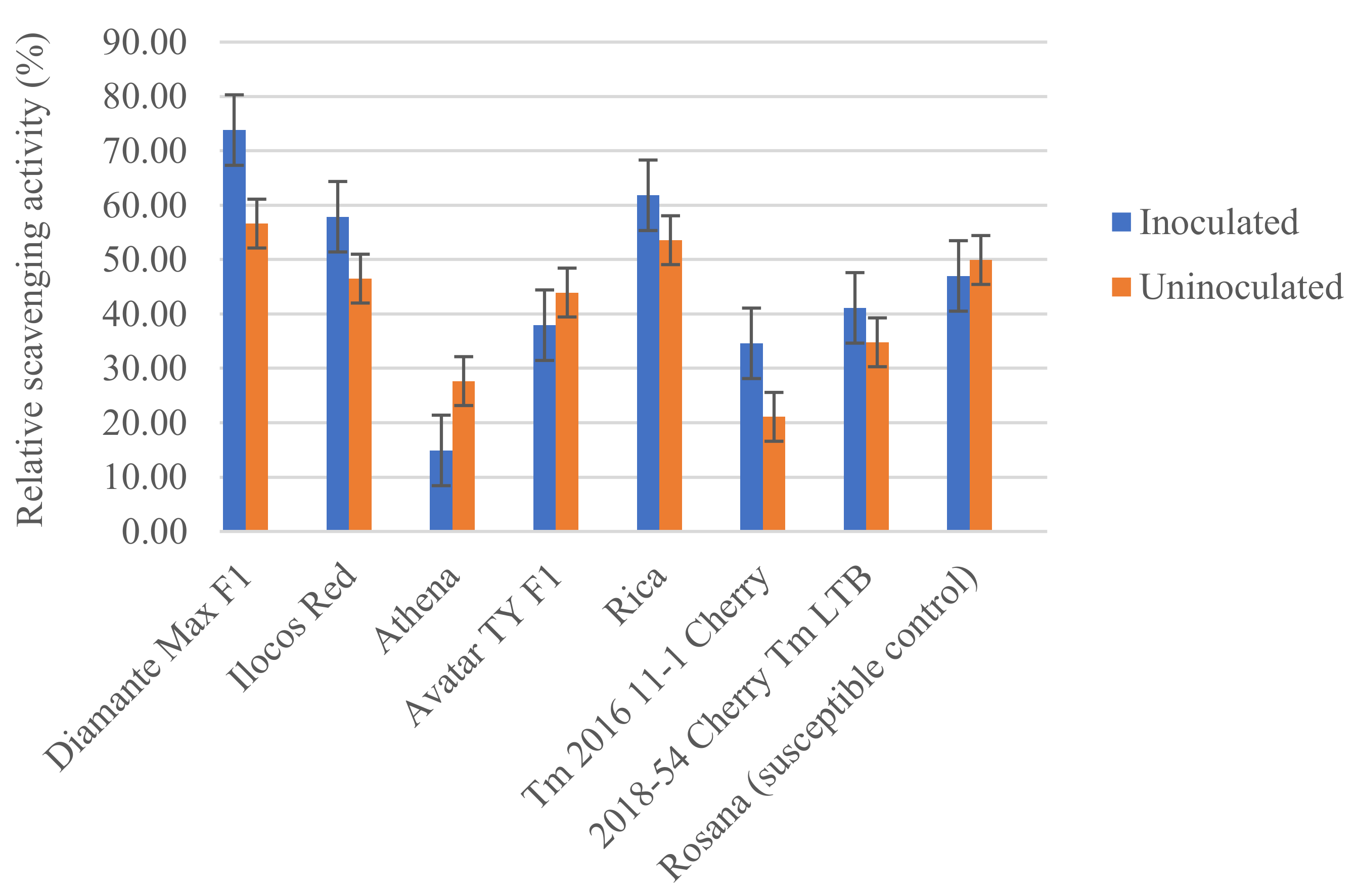
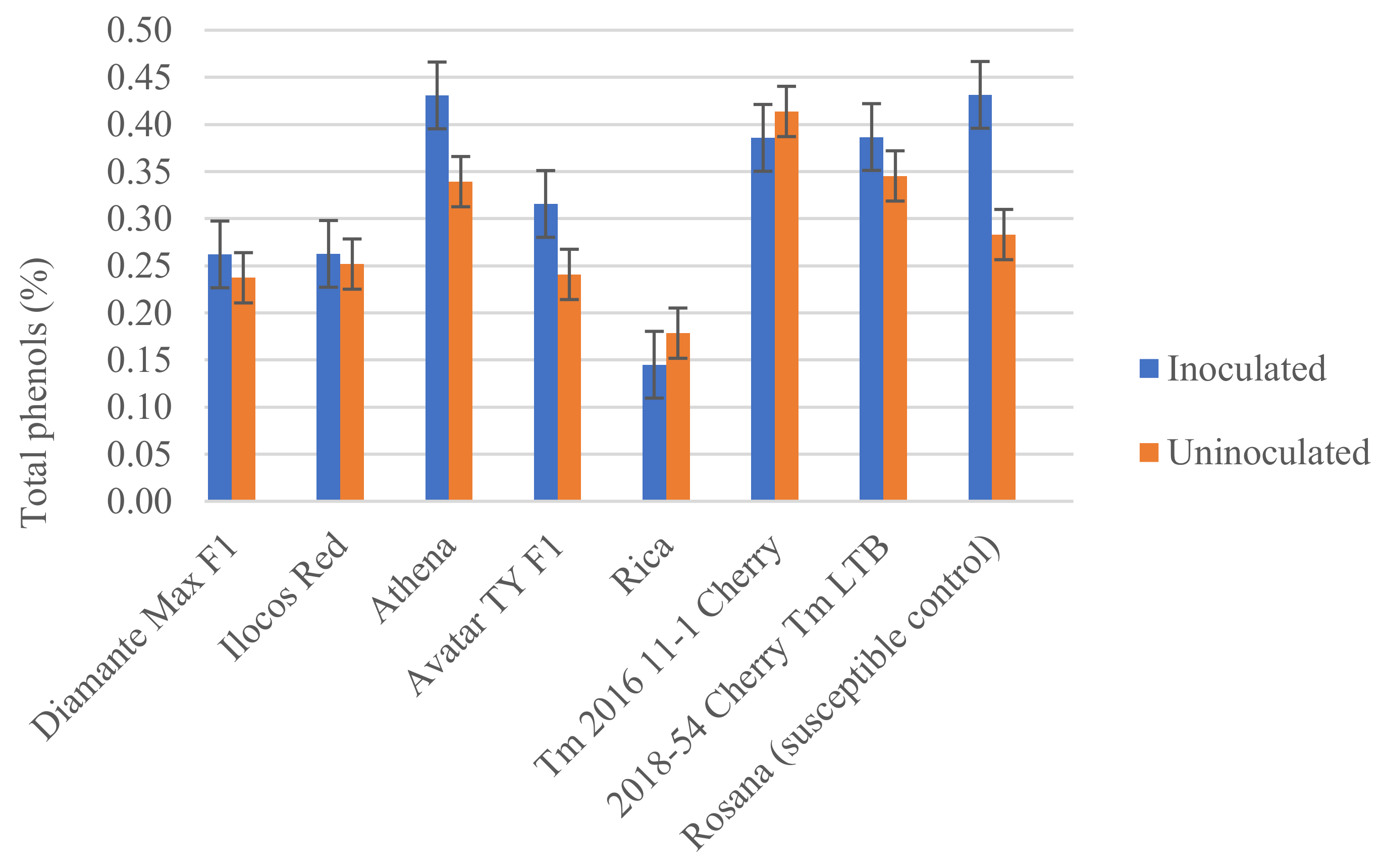
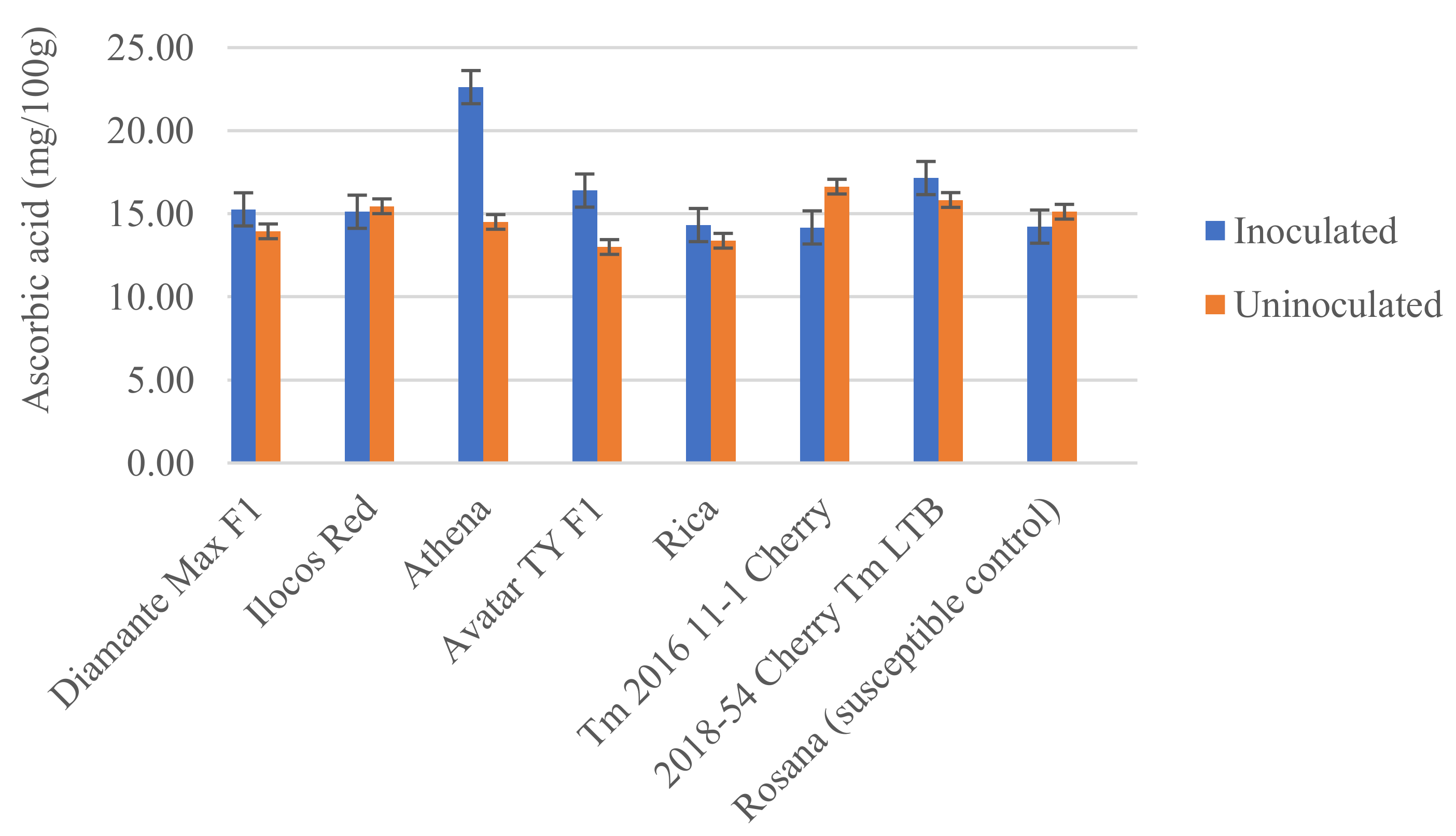
3.3. Biochemical Response of Tomato Varieties to Infection by Meloidogyne incognita
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization Corporate Statistical Database. FAOSTAT: Crops. Food Agriculture Organization, United Nations. 2017. Available online: https://www.fao.org/faostat/en/#home (accessed on 5 March 2019).
- Philippines Statistics Authority. Costs and Returns of Tomato Production; Philippine Statistics Authority: Quezon City, Philippines, 2017. [Google Scholar]
- Sikora, R.A.; Fernandez, E. Nematode Parasites of Vegetables. In Plant Parasitic Nematodes in Tropical and Subtropical Agriculture; Luc, M., Sikora, R.A., Bridge, J., Eds.; Centre for Agriculture and Bioscience International: Oxfordshire, UK, 2005; pp. 319–392. [Google Scholar]
- Mitkowski, N.A.; Abawi, G.S. Root-knot nematodes. Plant Health Instr. 2003. [Google Scholar] [CrossRef]
- Valdez, R.B. Nematodes Attacking Tomato and Their Control. In Proceedings of the 1st International Symposium on Tropical Tomato, Shanhua, Taiwan, 23–27 October 1978; Asian Vegetable Research and Development Center: Shanhua, Taiwan, 1978; pp. 136–150. [Google Scholar]
- Madamba, C.P. Distribution and identification of Meloidogyne spp. in the Philippines and five other Asian countries. Philipp. Agric. 1981, 64, 21–39. [Google Scholar]
- Davide, R.G. Nematode problems affecting agriculture in the Philippines. J. Nematol. 1988, 20, 214–218. [Google Scholar] [PubMed]
- Roberts, P.A.; May, D.; Matthews, W.C. Root-knot nematode resistance in processing tomatoes. Calif. Agric. 1986, 40, 24–26. [Google Scholar]
- Milligan, S.B.; Bodeau, J.; Yaghoobi, J.; Kaloshian, I.; Zabel, P.; Williamson, V.M. The Root knot nematode resistance gene Mi from tomato is a member of the leucine zipper, nucleotide binding, leucine-rich repeat family of plant genes. Plant Cell 1998, 10, 1307–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, P.G. Embryo culture of a tomato species hybrid. Proc. Am. Soc. Hortic. Sci. 1944, 44, 413–416. [Google Scholar]
- Medina Filho, H.P.; Stevens, M.A. Tomato breeding for nematode resistance: Survey of resistant varieties for horticultural characteristics and genotype of acid phosphates. Acta Hortic. 1980, 100, 383–393. [Google Scholar] [CrossRef]
- Arrigoni, O.; Zacheo, G.; Arrigoni-Liso, R.; Bleve-Zacheo, T.; Lamberti, F. Relationship between ascorbic acid and resistance in tomato plants to Meloidogyne incognita. Phytopathology 1979, 69, 579–581. [Google Scholar] [CrossRef]
- Branch, C.; Hwang, C.-F.; Navarre, D.A.; Williamson, V.M. Salicylic acid is part of the Mi-1-mediated defense response to root-knot nematode in tomato. Mol. Plant Microbe Interact. 2007, 17, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Chawla, N.; Choudhary, K.; Kaur, S.; Jindal, S. Changes in antioxidative enzymes in resistant and susceptible genotypes of tomato infected with root-knot nematode (Meloidogyne incognita). Indian J. Nematol. 2013, 43, 1–12. [Google Scholar]
- Cooper, W.R.; Jia, L.; Goggin, L. Effect of jasmonate-induced defenses on root-knot nematode infection of resistant and susceptible tomato cultivars. J. Chem. Ecol. 2005, 31, 1953–1967. [Google Scholar] [CrossRef] [PubMed]
- Giebel, J. Phenolic content in roots of some Solanaceae and its influence on IAA-oxidase activity as an indication of resistance to Heterodera rostochiensis. Nematologica 1966, 16, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Devran, Z.; Başköylü, B.; Taner, A.; Doǧan, F. Comparison of PCR-based molecular markers for identification of Mi gene. Acta Agric. Scand. Sect. B Soil Plant Sci. 2013, 63, 395–402. [Google Scholar] [CrossRef]
- Bhavana, P.; Singh, A.K.; Kumar, R.; Prajapati, G.K.; Thamilarasi, K.; Manickam, R.; Maurya, S. Identification of resistance in tomato against root knot nematode (Meloidogyne incognita) and comparison of molecular markers for Mi gene. Australas. Plant Pathol. 2018, 48, 93–100. [Google Scholar] [CrossRef]
- Seah, S.; Williamson, V.M.; Garcia, B.E.; Mejia, L.; Salus, M.S.; Martin, C.T.; Maxwell, D.P. Evaluation of a co-dominant SCAR marker for detection of the Mi-1 locus for resistance to root-knot nematode in tomato germplasm. Tomato Genet. Coop. Rep. 2007, 57, 37–40. [Google Scholar]
- Davide, R.G. Summary Report on the Current Status, Progress and Needs for Meloidogyne Research in Region VI. In An Advance Treatise on Meloidogyne; Sasser, J.N., Carter, C.C., Eds.; North Carolina State University Press Graphics: Raleigh, NC, USA, 1985; Volume 1, pp. 369–372. [Google Scholar]
- Eisenback, J.D. Diagnostic Characters Useful in the Identification of the Four Most Common Species of Root-Knot Nematodes (Meloidogyne spp.). In An Advance Treatise on Meloidogyne; Sasser, J.N., Carter, C.C., Eds.; North Carolina State University Press Graphics: Raleigh, NC, USA, 1985; Volume 1: Biology and Control, pp. 95–112. [Google Scholar]
- Hartman, K.M.; Sasser, J.N. Identification of Meloidogyne species on the Basisi of Differential Host Test and Perineal-Pattern Morphology. In An Advance Treatise on Meloidogyne; Barker, K.R., Carter, C.C., Sasser, J.N., Eds.; North Carolina State University Press Graphics: Raleigh, NC, USA, 1985; Volume 2: Methodology, pp. 69–77. [Google Scholar]
- Coyne, D.L.; Ross, J.L. Nematode Resistance Screening Root-Knot Nematodes Meloidogyne spp.; The International Institute of Tropical Agriculture: Ibadan, Nigeria, 2014. [Google Scholar]
- Taylor, A.L.; Sasser, J.N. Biology, Identification and Control of Root-Knot Nematodes (Meloidogyne spp.); North Carolina State University Press Graphics: Raleigh, NC, USA, 1978. [Google Scholar]
- Fox, J.; Bouchet-Valat, M. Rcmdr: R Commander. R Packag, Version 2.6-2: 2020; Available online: https://cran.r-project.org/web/packages/Rcmdr/Rcmdr.pdf (accessed on 15 November 2019).
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acid Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant Activity and Total Phenolics in Selected Fruits, Vegetables, and Grain Products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Jagota, S.K.; Dani, H.M. A new calorimetric technique for the estimation of vitamin C using folin phenol reagent. Anal. Biochem. 1982, 127, 178–182. [Google Scholar] [CrossRef]
- Garcia, B.E.; Mejia, L.; Salus, M.S.; Martin, C.T.; Seah, S.; Williamson, V.M.; Maxwell, D.P. A Co-Dominant SCAR Marker, Mi23, for Detection of the Mi-1.2 Gene for Resistance to Root-Knot Nematode in Tomato Germplasm. 2007. Available online: http://invirlab.plantpath.wisc.edu/GeminivirusResistantTomatoes/Markers/MAS-Protocols/Mi23-SCAR.pdf (accessed on 5 March 2019).
- Bajaj, K.L.; Mahajan, R. Phenolic compounds in tomato susceptible and resistant to Meloidogyne incognita (Kofoid et White) Chitwood. Nematol. Mediterr. 1977, 5, 329–333. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Plant Height | Fresh Weight | Shoot Weight | Root Weight | Number of Galls | Gall Index |
|---|---|---|---|---|---|---|
| Diamante Max F1 | −31.2 a | −22.9 ab | −30.5 ab | 76.7 a | 189.8 abc | 4.8 ab |
| Ilocos Red | −22.8 a | −3.9 ab | −13.4 ab | 145.6 a | 265.3 a | 5.0 a |
| Avatar TY F1 | −32.5 a | −53.2 a | −58.8 a | 92.4 a | 133.3 bcd | 4.8 ab |
| Athena | −23.6 a | −32.2 ab | −40.2 ab | 122.2 a | 48.5 d | 3.8 b |
| Rica | −44.9 a | −60.6 a | −67.9 a | 66.7 a | 54.8 d | 4.0 b |
| Tm 2016 11−1 Cherry | −3.2 a | 30.0 b | 22.0 b | 216.7 a | 94.3 cd | 4.5 ab |
| 2018−54 Cherry Tm LTB | −18.7 a | −18.0 ab | −28.6 ab | 135.6 a | 255.8 ab | 5.0 a |
| Rosanna (susceptible control) | −9.3 a | −3.4 ab | −10.2 ab | 117.7 a | 122.7 cd | 4.5 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lizardo, R.C.M.; Pinili, M.S.; Diaz, M.G.Q.; Cumagun, C.J.R. Screening for Resistance in Selected Tomato Varieties against the Root-Knot Nematode Meloidogyne incognita in the Philippines Using a Molecular Marker and Biochemical Analysis. Plants 2022, 11, 1354. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101354
Lizardo RCM, Pinili MS, Diaz MGQ, Cumagun CJR. Screening for Resistance in Selected Tomato Varieties against the Root-Knot Nematode Meloidogyne incognita in the Philippines Using a Molecular Marker and Biochemical Analysis. Plants. 2022; 11(10):1354. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101354
Chicago/Turabian StyleLizardo, Roden Carlo M., Marita S. Pinili, Maria Genaleen Q. Diaz, and Christian Joseph R. Cumagun. 2022. "Screening for Resistance in Selected Tomato Varieties against the Root-Knot Nematode Meloidogyne incognita in the Philippines Using a Molecular Marker and Biochemical Analysis" Plants 11, no. 10: 1354. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11101354