
Metabolome Profiling: A Breeding Prediction Tool for Legume Performance under Biotic Stress Conditions


1
Department of Agriculture and Animal Health, School of Agriculture and Life Sciences, College of Agriculture and Environmental Sciences, University of South Africa, 28 Pioneer Ave, Florida Park, Roodeport 1709, South Africa
2
Research Support Services, North West Provincial Department of Agriculture and Rural Development, 114 Chris Hani Street, Potchefstroom 2531, South Africa
*
Author to whom correspondence should be addressed.
Plants 2022, 11(13), 1756; https://0-doi-org.brum.beds.ac.uk/10.3390/plants11131756
Submission received: 25 May 2022
/
Revised: 17 June 2022
/
Accepted: 22 June 2022
/
Published: 1 July 2022
(This article belongs to the Special Issue Epidemiology and Control of Plant Diseases)
Abstract
:Legume crops such as common bean, pea, alfalfa, cowpea, peanut, soybean and others contribute significantly to the diet of both humans and animals. They are also important in the improvement of cropping systems that employ rotation and fix atmospheric nitrogen. Biotic stresses hinder the production of leguminous crops, significantly limiting their yield potential. There is a need to understand the molecular and biochemical mechanisms involved in the response of these crops to biotic stressors. Simultaneous expressions of a number of genes responsible for specific traits of interest in legumes under biotic stress conditions have been reported, often with the functions of the identified genes unknown. Metabolomics can, therefore, be a complementary tool to understand the pathways involved in biotic stress response in legumes. Reports on legume metabolomic studies in response to biotic stress have paved the way in understanding stress-signalling pathways. This review provides a progress update on metabolomic studies of legumes in response to different biotic stresses. Metabolome annotation and data analysis platforms are discussed together with future prospects. The integration of metabolomics with other “omics” tools in breeding programmes can aid greatly in ensuring food security through the production of stress tolerant cultivars.
1. Introduction
Leguminous crops such as Arachis hypogaea (groundnut), Glycine max (soybean), Phaseolus vulgaris (common bean), Pisum sativum (common pea), Cicier arietinum (chickpea), Vigna anguiculata (cowpea), Vicia faba (faba bean), Lens culinaris (lentil), Cajanus cajan (pigeon pea), Lupinus spp. (lupin), and Vigna subterranean (bambara bean) contribute to the improvement of ecosystems [1,2,3], nutrition and food security [4,5,6,7]. Although legumes contribute greatly to food security, their production globally is hindered by biotic stresses that include nematodes, viruses, insect pests, and bacterial and fungal pathogens [8,9,10]. The occurrence of biotic stresses in legume production systems has impacted negatively on production and has resulted in significant yield losses globally [11,12,13]. In many breeding programmes, the key objective is to develop crop varieties that are adaptable to an array of stressors in order to meet global food demands [14,15,16], thus addressing sustainable development goals 1 and 2 of the United Nations [17]. Legume programmes have been improving gradually over the years and have advanced from traditional methods of breeding to using genomic tools [18]. Traditional breeding techniques rely mostly on manual selection and the crossing of genotypes with desirable traits, and although these methods have contributed greatly to legume breeding, the genetic gain was often not statistically significant [19].
Contemporary biotechnology tools including next generation sequencing (NGS) platforms have aided many breeding programmes with provision of genetic data that traditional breeding techniques cannot fully reveal [20]. Biotechnological “omics” approaches have contributed greatly to breeding aimed at the improvement of plant stress tolerance by providing insight into genetic diversity, genotype variations, genetic maps and other useful information pertaining to the genetics of plant populations [21,22]. Despite the importance of genomic data generated by the other omics platforms (transcriptomics, transgenomics, epigenomics), plants produce molecular compounds with molecular weights expressed in abundance and are responsible for biochemical functions under different environments [23]. Metabolomics highlights metabolite expressions and changes, together with their interactions and phenotypic characters of plants under stress conditions. When plants are exposed to stress, metabolic homeostasis alterations occur, requiring the plant to adjust its metabolomic pathways, and this phenomenon is referred to as acclimation [24,25,26,27]. When this process occurs, the plant activates signal transduction pathways that set off the assembly of proteins and metabolomic compounds that aid in reaching a new homeostasis [28,29]. Furthermore, metabolome analysis provides information on the metabolomic pathways that are responsible for complex processes that occur when a plant is exposed to stress conditions [20].
A detailed review of metabolomic studies focused on specific biotic stressors of legumes can aid in identifying gaps and create an interactive platform for researchers to conduct, and possibly collaborate on, more studies aimed at improving legume production in the world. This is because the dimensionality of large data sets generated through metabolomics can be interpreted holistically utilising multivariate data analysis [30]. This will further highlight the importance of metabolite detection in breeding programmes and techniques that can be employed for different objectives since metabolites relate to phenotypic and genomic data [9]. This review reports on metabolomics as a breeding prediction tool in legume breeding under biotic stress. We also briefly discuss the impact of metabolomics in legume breeding programmes aimed at improving biotic stress tolerance.
2. Biotic Stressors of Legumes
2.1. Insect Pests
Insect pests attack legume crops by boring, webbing and damaging plant parts such as the leaves, pods, stems and roots [31,32]. In addition to attacking plants, insect pests may also act as vectors for pathogens that negatively impact crop production systems [33]. Insect pests such as aphids [33,34], pod borers [31,35], thrips [36,37] and whiteflies [38,39] have been reported to feed on legume crops, among others. The use of biological enemies of pests, cultural control (crop rotation, mulching, intercropping, etc.), mechanical control (water hosing at high pressure), chemical application and integrated pest management strategies have been recommended for the control of insect pests in legumes [39,40,41,42]. These efforts have been found to be effective in reducing insect severity in legumes [39,43]. However, the insects are constantly adapting to control measures used in production systems [44]. Breeding for tolerance to insect pests is the most sustainable approach and this requires an understanding of the plant’s signal pathways that respond to insect attack [45].
Pathways expressed in rice infested with caterpillars included flavonoids, phenolic acids, amino acids and derivatives. These improved the production of cytosolic calcium ions that signal herbivore attack to the plant [46]. Maize infested with Monolepta hieroglyphica revealed significant up-/down-regulation of metabolites derived from sugar and amino acid pathways that might be responsible for resistance. Similar results were reported in cabbage infested with aphids [47]. Insect–plant metabolomic response of leguminous crops has been conducted for red clover, pea and alfalfa in a composite study with aphid infestation. Triterpene, flavonoid and saponin enriched pathways were found to be responsive to aphid attack [34]. Flavonoids and amino acids have also been found to be significantly enriched in alfalfa infested with thrips [48]. However, limited studies have been conducted on the host-plant metabolomic response of leguminous crops to insects, as well as to other biotic stressors. These studies could have far-reaching impacts on stress biomarker identification with potential benefits in legume improvement programmes.
2.2. Diseases of Legumes
2.2.1. Bacterial Diseases
Bacterial diseases of legumes can be categorised into leaf blights, leaf spots/bacterial wilts and other multiple symptoms of sprout rot and dwarfism [49]. Their symptoms are based on the tissues that they infiltrate (leaves, stems and roots) [50]. Legume bacterial diseases are known to cause yield losses of up to 50%, which negatively impacts economic gains and food security [51]. The two plant bacterial pathogens Xanthomonas axonopodis and Pseudomonas syringae are known worldwide for causing bacterial blight [49,52]. Symptoms of infection usually occur on all aerial parts of the plant, and in severe incidences, defoliation and wilting occur [52,53]. Like bacterial blight, another disease that threatens legume production is bacterial wilt, caused by Curtobacterium flaccumfaciens pv. Flaccumfaciens [54]. The pathogen has created new variants that cause damage to legume crops worldwide by causing leaf chlorosis in plants. In fields where the disease occurs, upon plant maturation and shattering of seeds, the infected seed replants itself and allows the pathogen to thrive from generation to generation [54,55]. The control of bacterial diseases has relied on integrated approaches that limit the survival of pathogens. This includes crop rotation and the use of pathogen free certified seed [52]. These measures are only effective to a limited extent, and detecting pathogens in seed is not an easy task for farmers. A promising and more long-term method for the control of bacterial diseases would be the utilisation/breeding of tolerant varieties [56,57].
The evaluation of metabolite profiles in citrus infected with huanlongbing caused by the bacterium ‘Candidatus Liberibacter asiaticus’ reported distinct sugars as well as amino and organic acids expressed in the roots, thus giving insight on resistance [58]. Metabolomic compounds synthesized from flavonoids, amino and phenolic acids act as protective agents in the xylem of oat plants when infected with halo blights caused by P. syringae pv. by repairing the cell wall [59]. Similar metabolomic pathways including phenols and acetates have been reported in tomato infected with bacterial wilt caused by Ralstonia solanacearum [60]. To date, there is little to no information from metabolomic studies on the response of leguminous crops to bacterial disease infection to aid breeders with biomarker discovery.
2.2.2. Fungal Diseases
The occurrence of fungal diseases in legume production areas is known to cause substantial yield losses of up to 100% [59]. Fungal pathogens can cause infection at any plant growth stage (emergence, seedling, vegetative and reproductive stage) by attacking organs and tissues that are involved in the transportation of water and nutrients [61,62]. Upon infection, these pathogens degrade the plant cell wall, which consequently results in the death of the plant, especially if the variety grown does not have any resistant genes [63]. Root rot caused by Rhizoctonia solani, Fusarium solani, Fusarium oxysporum and Aphanomyces euteiches and fungal wilt caused by Formae speciales are some of the most destructive fungal diseases that limit the productivity of legume crops worldwide [64]. The pathogen R. solani is considered one of the most destructive fungal pathogens that usually infects the roots and hypocotyl of the plant through penetration of the appressoria [63]. At pre-emergence and post-emergence plant growth stages, R. solani causes symptoms of damping-off, root rot and stem canker [65]. Under greenhouse conditions, the seedling survival of some leguminous crops may be less than 5% [66]. The pathogen may further infect the plant’s fruits in highly humid conditions, thus reducing crop quality and yield [67]. Fusarium spp. are also predominant pathogens that interfere with plant growth by causing damping off and root rot [68]. In African small-scale farms, yield losses of up to 100% caused by the F. solani pathogen in common bean have been reported [69]. In addition, A. euteiches is a soil-borne fungal pathogen that poses a threat to legume production by causing wilting, root rot and consequently yield losses of up to 80% [70,71].
The management of fungal diseases is problematic due to the complexity of these pathogens [72]. Over the years, management has been implemented by integrating conventional methods such as crop rotations, increased greenhouse temperatures, biological enemies and chemical use [73]. The use of fungicides has been a promising avenue for the control of fungal pathogens. However, chemicals used to control pathogens have an immense economic and environmental impact [74]. This has led to the exploration of using biological control measures such as bacterium and fungal strains as environmentally friendly alternatives to control pathogens that attack plants [75]. Trichoderma spp. are widely used strains for the biological control of fungal diseases. Beneficial strains of T. velutinum have been found to be an effective biological control measure that promotes the accumulation of metabolites that are responsible for defence in common bean infected with F. solani. Even though numerous strains have been found to be effective in controlling fungal diseases, legislation in many countries regarding the use of biopesticides and their shelf life is still a challenge [76,77]. The development of disease-resistant cultivars using genomic technologies can aid in improving legume productivity worldwide [54]. Legume metabolomics focussed on breeding for disease resistance can be beneficial to breeding programmes by increasing the availability of resistant genotypes that are released to farmers [78].
The metabolomic profiling of leguminous crops has been conducted in common bean and provided major findings in relation to metabolomic pathways including amino acids, flavonoids, isoflavanoids, purines and proline metabolism, which were shown to promote plants’ potential for defence against Fusarium pathogens [79]. In addition, Mayo-Prieto et al. [80] also reported amino acids, peptides, carbohydrates, flavonoids, lipids, phenols, terpenes and glycosides that were up-/down-regulated as a defence mechanism by the common bean plant against the pathogen R. solani. Similar results have been reported in other leguminous crops including chickpea infected with F. oxysporum, soybean infected with Aspergillus oryzae/Rhizopus oligosporus, pea infected with Dydymella pinodes and R. solani (Table 1) [81,82]. Intensifying the fungal–legume metabolomic research worldwide will aid in understanding the biochemical properties of these leguminous crops in response to disease stress.
2.2.3. Viral Diseases
Viral pathogens attack many crops, including legumes, by causing the yellowing of leaves, stunting and poor pod setting, which result in poor yields [65]. Major viral diseases causing production losses in legumes belong to the Nanoviridae, Luteovridae and Poltyvridae families. These diseases cause the necrosis of plants, and their identification requires molecular techniques. Over the years, the accurate identification of viruses has improved because of an increasing number of available genomic platforms. [49,66]. Viruses attach themselves to specific sites of vectors such as insects (aphids, beetles, etc.) and remain there until transmission to their host occurs [67]. The control of viral diseases is difficult and thus requires adherence to quarantine prescripts, removal of inoculum sources, adjustments of planting dates, intercropping, crop rotation, chemical application aimed at controlling pests (elimination of vectors) and the use of tolerant/resistant genotypes [68].
Utilising metabolomic techniques on the Citrus tristeza virus of Mexican lime Citrus aurantifolia revealed up-/down-regulation of amino acids, alkaloids and phenols during infection, thus signalling pathogen defence when different strains of the virus were utilised [83]. In stems of Amarathus hypochondriacus L. infected with Ageratum enation virus, alkaloids, amino acids, dicarboxylic acids, glutamine and sugars may increase or decrease in concentration as a mechanism to improve overall respiratory metabolism [84]. Studies on the response of leguminous crops to viral disease infection are limited, thus requiring more research in order to fully understand the underlying information relating to metabolites expressed under virus pressure.
2.3. Parasitic Weeds
Unlike “normal” weeds that disadvantage the plant greatly, parasitic weeds on the other hand extensively extract moisture, nutrients, photosynthates and other resources from the host plant [69]. When parasitic weeds are not controlled, the extraction of resources continues, consequently extinguishing the crop [70]. Roomrape species, Striga gesnerioides and Alectra vogelii are problematic parasitic weeds that cause yield losses in many legume production areas in Sub-Saharan Africa [71]. Biological control [69], intercropping [72], chemical application and cultural practices (timely planting) are recommended for the control of parasitic weeds [73]. However, these are often not successful, and the fight against parasitic weeds lies within breeding for resistance [71,73]. Although breeding for resistance will aid in controlling parasitic weeds, the complexity and low heritability is a challenge that breeders face when breeding for parasitic weed resistance [71,73,74]. Initiatives to use breeding prediction tools such as metabolomic techniques for parasitic weed resistance have been explored in rice to study and dissect S. hermonthica resistance [85]. This study reported the phenylpropanoid pathway, which contributes to the formation of lignin in rice, to be an important pathway that can be utilised for resistance to S. hermonthica. There is a deficit on metabolomic experiments that evaluate the performance of legumes under parasitic weed conditions.
2.4. Parasitic Nematodes
Legumes are famous for their ability to fix nitrogen by using rhizobium, which is a mutualist bacterium [75]. However, the presence of parasitic nematodes reduces rhizobia activity, which leads to poor nodulation [76]. Parasitic nematodes invade the roots of plants and form an indefinite feeding area, which, in turn, can affect root development, thus leading to poor plant growth [77]. Heterodera and Globodera spp. are root knot and cyst nematodes that affect many crops including legumes, resulting in over 12% yield losses [78]. The presence of parasitic nematodes often leads to infection by other pathogens including fusarium spp.; therefore, the utilisation of sustainable control strategies for other pathogens is essential for legumes [74]. Soybean evaluated under Melodegyne pinodes and Heterodera glycines pressure exhibited phenylpropanoids, cysteine, methionine, alkaloid and tropane pathways that can be attributed to resistance properties of the crop to nematodes [86]. The in-depth exploration of metabolites of other crops including legumes would be beneficial to understanding nematode–crop biological interactions.

{kind=link}
{kind=link}
Table 1.
Summary of metabolomic studies conducted in response to biotic stress in leguminous crops using different platforms such as GC-MS, LC-QqQ-MS, LC-MS, LC-obitrap-MS, UHPLC-MS, 1H NMR and GC-MS/TOF.
Table 1.
Summary of metabolomic studies conducted in response to biotic stress in leguminous crops using different platforms such as GC-MS, LC-QqQ-MS, LC-MS, LC-obitrap-MS, UHPLC-MS, 1H NMR and GC-MS/TOF.
| Legume | Biotic Stress | Classification | Method | Total Metabolites | Reference |
|---|---|---|---|---|---|
| C. arietinum | Fusarium oxysporum | Fungal | GC-MS | 72 | [87] |
| G. max | Aspergillus oryzae/Rhizopus oligosporus | Fungal | LC-QqQ-MS | 489 | [88] |
| Heterodera glycines | Nematode | GC-MS | 20 | [86] | |
| M. sativa | Thysanoptera spp. | Insect | LC-MS | 772 | [48] |
| Acyrthosiphon pisum Harris | Insect | LC-Obitrap-MS/UHPLC-MS | 107 | [34] | |
| P. sativum | Acyrthosiphon pisum Harris | Insect | LC-Obitrap-MS/UHPLC-MS | 57 | [34] |
| Didymella pinodes | Fungal | LC-MS/MS | 31 | [89] | |
| Rhizoctonia solani | Fungal | 1H NMR | 126 | [81] | |
| Didymella pinodes | Fungal | GC-MS/TOF | 39 | [82] | |
| P. vulgaris | Fusarium solani | Fungal | UPLC | 743 | [79] |
| Trichoderma velutinum/Rhizoctotonia solani | Fungal | LC-MS | 216 | [80] | |
| T. pratense | Acyrthosiphon pisum Harris | Insect | LC-Obitrap-MS/UHPLC-MS | 103 | [34] |
| V. faba | Acyrthosiphon pisum Harris | Insect | LC-Obitrap-MS/UHPLC-MS | 13 | [34] |
3. Legume Metabolomics
3.1. Metabolome Profiling Techniques
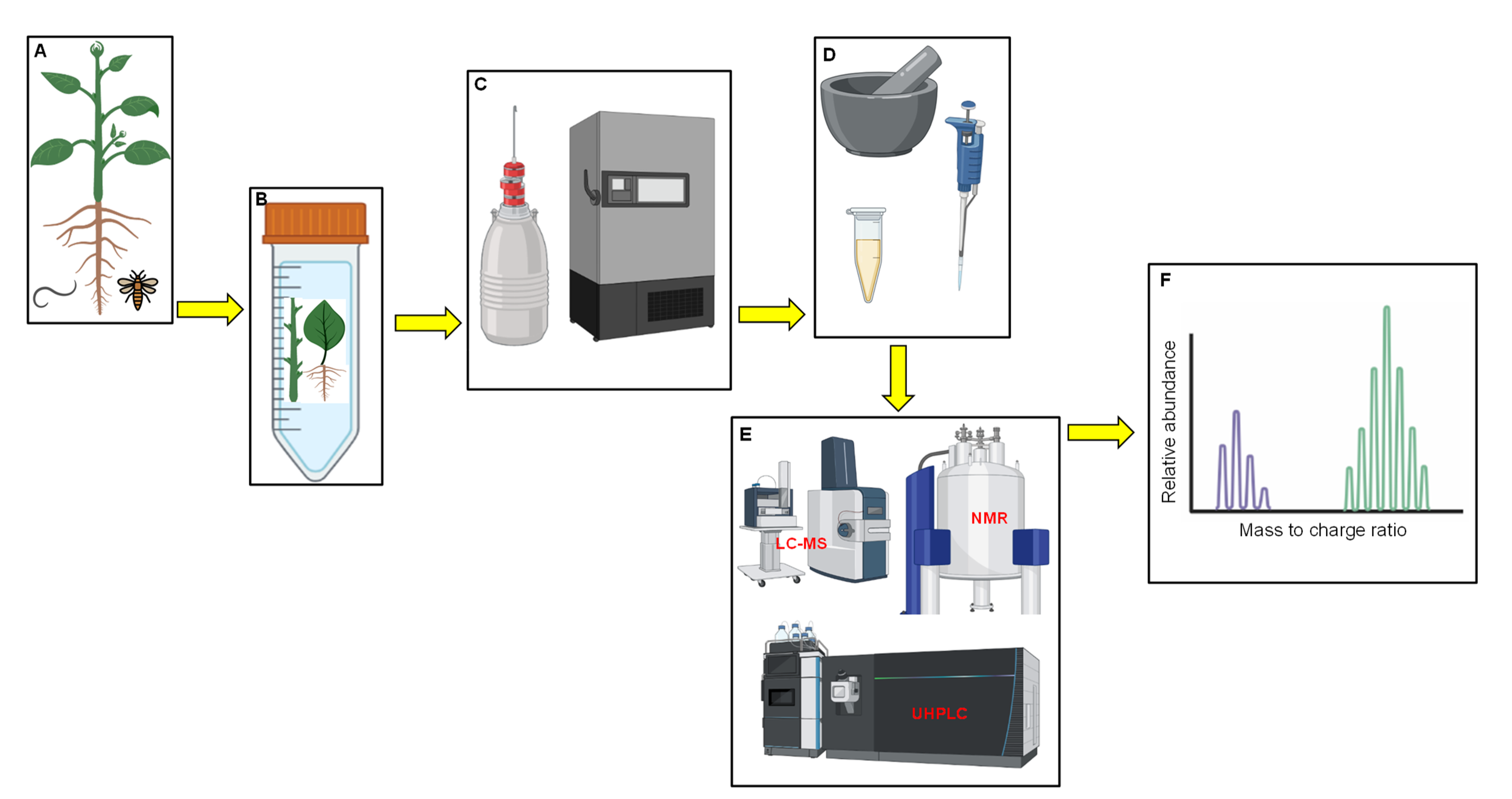
The use of metabolomics has been applauded for its ability to provide detailed prospects by in-depth study of crop biology. Information that is derived from metabolomic tools can be translated to assess phenotypic changes/biomarkers, gene changes and, also, to distinctively support other genomic experiments [79,80]. Furthermore, metabolomic studies can be applied for polygenic traits and prediction of epistatic effects [79,88]. The overall success of detecting metabolites and their changes depends on utilising analytical techniques that can detect compound concentrations, proportions and molecular weights [81,82,89]. The concept of metabolome profiling was introduced with the use of mass spectrometry and at a later stage, gas chromatography was also introduced [87]. Since the inception of the latter, metabolome profiling using both spectrometric and chromatographic techniques have been improving [30,90]. Different strategies are utilised for compound profiling in metabolomics, including metabolite profiling, fingerprinting and target analysis [91,92]. Metabolite fingerprinting compares “fingerprints” of metabolites [93]. The profiling analyses broader groups of metabolites that are related to specific pathways or compound classes, while target analysis is utilised for targeting specific metabolic pathways and observes the occurrences of modifications [94]. Protocols for both metabolite profiling and fingerprinting in stress experiments involve the sample acquisition from a stressed plant (leaves, stems or roots; Figure 1A) that are cut and placed in a labelled tube (Figure 1B). Dewar with liquid nitrogen is ideal for snap freezing samples in the field and a laboratory ultra-freezer with a temperature above −60 °C is recommended for sample preservation to avoid dehydration (Figure 1C). The stored samples are then crushed, and extraction is conducted in preparation for metabolite analysis, using the appropriate technology that generates spectral data (Figure 1D–F).
3.2. Metabolite Profiling
Metabolite profiling is important in studying organisms’ biochemical pathways [88]. Numerous technologies such as gas chromatography-mass spectrometry (GC-MS), liquid chromatography-mass spectrometry (LC-MS), nuclear magnetic resonance (NMR), capillary electrophoresis-MS (CE-MS) and Fourier transform-infrared (FT-IR) spectroscopy are commonly used analytical platforms for metabolite profiling [49,95]. The unique properties of these profiling techniques together with their applications, limitations and successes in plant metabolomics have been discussed by numerous researchers [30,96,97,98,99]. There are limited studies on the metabolome profiling of legume crops evaluated under insect stress. Although not a model for legume crops, metabolomic profiling has been performed on Medicago sativa (a close relative of the model legume crop M. truncatula) under insect stress (Table 1) [34,48]. In plant–insect interactions, a metabolome profiling study on alfalfa cultivars reported the production of numerous up-regulated metabolites in response to infestation by thrips using LC-MS (Table 1). Among the metabolite classes were amino acids together with derivatives that produced toxic amino acids released by the plant in response to insect attack [48]. Similar metabolites analysed using UHPLC-MS were also reported for pea (P. sativum), red clover (Trifolium pratense) and other alfalfa genotypes in response to biotic stress [34]. In addition, Narula et al. [87] reported a large number of metabolites that were up-regulated and down-regulated when chickpea was infected with F. oxysporum using GC-MS as a metabolome profiling tool. Similar results were also reported for common bean infected with F. solani [79], T. velutinum and R. solani [80] (Table 1). Among the primary metabolites reported, amino acids, alcohols and alkaloids were upregulated. Precursor molecules of these metabolites were found to be responsible for defence and energy provision for the plant [91]. More studies have been reported on P. sativum focusing on metabolite profiling under biotic stress (Table 1), particularly fungal pathogens [92,100,101]. For example, using 1H NMR, young pea plants showed a heightened production of amino acids that signal the production of the metabolite proline during fungal infection [81]. However, as the plant grows older, its energy requirements change, and proline production reduces. Overall, the down-regulation of metabolites can be used as a guideline for selecting resistant/tolerant varieties. Varieties resistant to pathogens also produce sulphur as a defence strategy. Resistant cultivars tend to have increased sulphur assimilation with high energy accumulation from sugar metabolites (nitrogen mobilization) for restoration of damaged plant cells [92].
4. Metabolome Data Processing and Annotation Tools Used in Legume Stress Tolerance
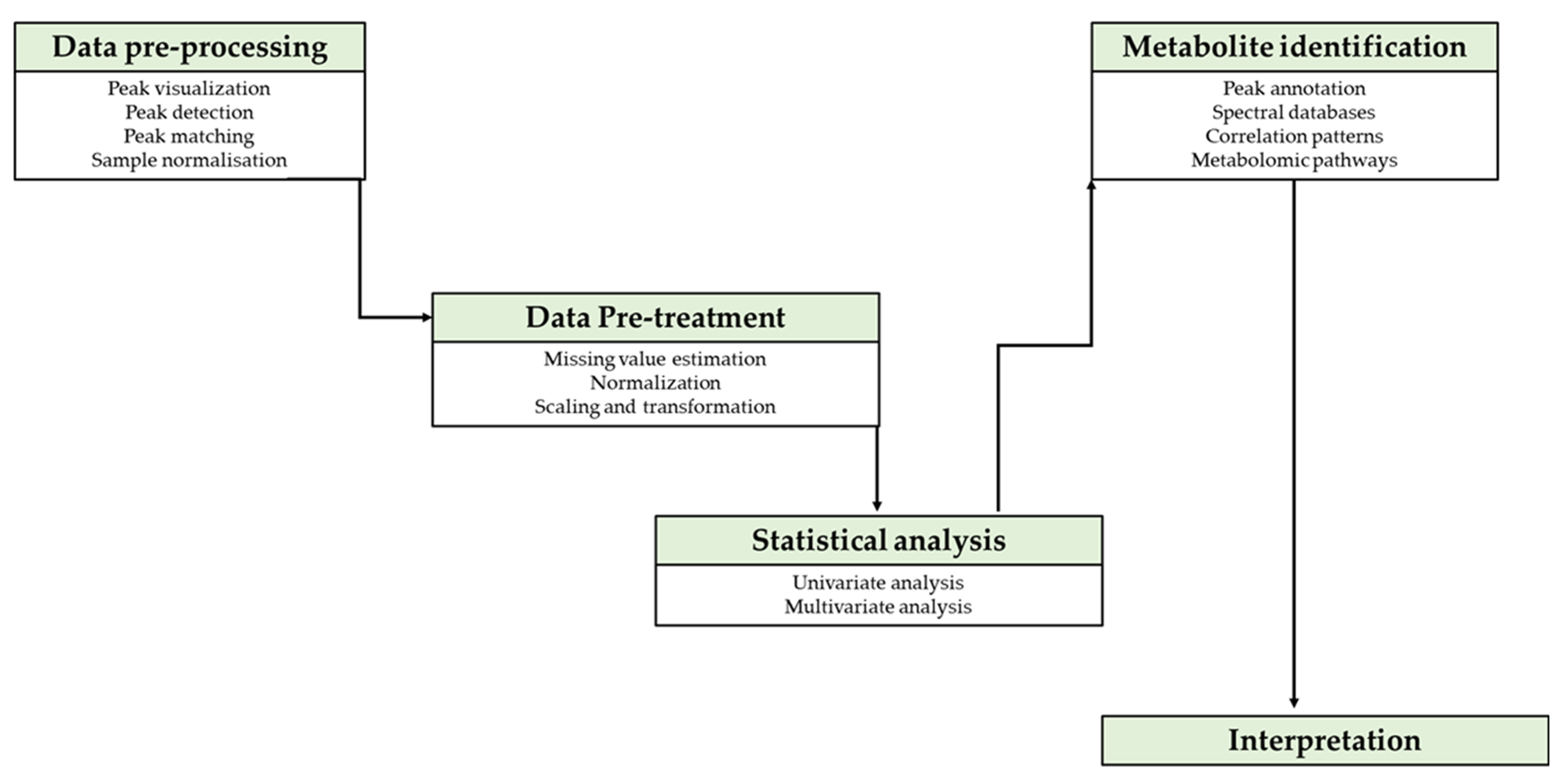
Metabolome usage has grown rapidly because of its provision of the cellular function data of small molecules (<1500 Da) linked to more than 40,000 metabolites that are registered on numerous databases [102]. Data generated by metabolomic technologies such as GC-MS, LC-MS and NMR, amongst others, are enormous and require software tools that are able to visualise, detect peaks, normalize/transform the sample data, annotate, identify, quantify and statistically analyse targeted/untargeted metabolite variations, in accordance with applied algorithms for univariate/multivariate analysis (Figure 2) [103,104]. There is no single tool that can unravel information from a metabolome profile; thus, analysis integrates numerous databases and requires algorithms that are provided by an array of tools [105]. Studies of metabolites in crops use an array of statistical platforms to evaluate variations of metabolites in different stress environment [106]. In legumes, metabolome data processing platforms (Table 2) used in studies of biotic stress for legumes include R and SIMCA [48,81]. Software such as SIMCA, Analyst software, STAT GRAPHICS Centurion, Labsolutions, ChromaTOF and agilent software MassHunter require licensing for metabolome data processing. However, there are numerous web-based accessible platforms that can be used for data processing, metabolome annotation and visualisation such as R, XCMS, MetaboAnalyst, METLIN, KEGG, HMBD, MeV, MetLAB and others (Table 2 and Table 3) [103].
The representation of biological networks is important in metabolomics, as it gives representation of relationships or patterns that occur in metabolomic pathways. There are numerous metabolomic pathway databases that aid in grouping metabolites with similar functions. Metabolomic pathway databases including KEGG, cytoscape, MapMan and iPath, among others, are applicable to plants [103,107].
5. Conclusions
Legume crops are grown in most regions of the world because they provide food security for many households. With the current climate crisis, the production of crops that are adaptable to biotic and abiotic stress is paramount. Legumes are produced in semi-arid environments and in these production areas, multiple stressors are prevalent. Plant stress response is a very complex phenomenon that researchers are constantly striving to understand by making use of high-throughput techniques. The integration and application of omics tools in agriculture has evolved and broadened the understanding of the underlying biochemical and molecular mechanisms of crops grown in diverse environments. Metabolomic studies are already becoming one of the omics tools used for breeding strategies. However, strong bioinformatics skills are needed for the processing and manipulation of the data. Furthermore, metabolomic database availability should be improved in order to accelerate information availability for legume crops. Additionally, studies that integrate metabolomics with other omics tools should aim to elaborate on the metabolomic aspects. For example, in many studies integrating transcriptomics and metabolomics, the information tends to be denser for gene expression than for metabolomics. In such cases, metabolome specific papers should be published separately to avoid complexity of integrating all the data and suppressing metabolomic information. Overall, the integration of metabolomics with other omics tools provides a powerful strategy to unravel plant–pest/pathogen interaction in biotic stress environments.
Author Contributions
Conceptualization, S.F. and M.R.; writing—original draft preparation, P.M., S.F., M.R. and H.M.; writing—review and editing, P.M., S.F., M.R. and H.M.; funding acquisition, S.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data are included in the main text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gao, D.; Wang, X.; Fu, S.; Zhao, J. Legume plants enhance the resistance of soil to ecosystem disturbance. Front. Plant Sci. 2017, 8, 1295. [Google Scholar] [CrossRef] [PubMed]
- Makarov, M.; Onipchenko, V.; Malysheva, T.; Zuev, A.; Tiunov, A. Symbiotic nitrogen fixation by legumes in alpine ecosystems: A vegetation experiment. Russ. J. Ecol. 2021, 52, 9–17. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.; Wang, X.; Fu, S. Legume-soil interactions: Legume addition enhances the complexity of the soil food web. Plant Soil 2014, 385, 273–286. [Google Scholar] [CrossRef]
- Jimenez-Diaz, R.; Castillo, P.; Jimenez-Gasco, M.; Landa, B.; Navas-Cortes, J. Fusarium wilt of chickpeas: Biology, ecology and management. Crop Prot. 2015, 73, 16–27. [Google Scholar] [CrossRef]
- Kulkarni, K.; Tayade, R.; Asekova, S.; Song, J.; Shannon, J.; Lee, J. Harnessing the potential of forage legumes, alfalfa, soybean, and cowpea for sustainable agriculture and global food security. Front. Plant Sci. 2018, 9, 1314. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.; Oyatomi, O.; Babalola, O.; Abberton, M. Breeding potentials of bambara groundnut for food and nutrition security in the face of climate change. Front. Plant Sci. 2022, 12, 798993. [Google Scholar] [CrossRef] [PubMed]
- Sauer, C.; Mason, N.; Maredia, M.; Mofya-Mukuka, R. Does adopting legume-based cropping practices improve the food security of small-scale farm households? panel survey evidence from Zambia. Food Secur. 2018, 10, 1463–1478. [Google Scholar] [CrossRef]
- Kudapa, H.; Ramalingam, A.; Nayakoti, S.; Chen, X.; Zhuang, W.; Liang, X.; Kahl, G.; Edwards, D.; Varshney, R. Functional genomics to study stress responses in crop legumes: Progress and prospects. Funct. Plant Biol. 2013, 40, 1221–1233. [Google Scholar] [CrossRef] [Green Version]
- Ramalingam, A.; Kudapa, H.; Pazhamala, L.; Weckwerth, W.; Varshney, R. Proteomics and metabolomics: Two emerging areas for legume improvement. Front. Plant Sci. 2015, 6, 1116. [Google Scholar] [CrossRef] [Green Version]
- Vrignon-Brenas, S.; Celette, F.; Piquet-Pissaloux, A.; David, C. Biotic and abiotic factors impacting establishment and growth of relay intercropped forage legumes. Eur. J. Agron. 2016, 81, 169–177. [Google Scholar] [CrossRef]
- Dita, M.; Rispail, N.; Prats, E.; Rubiales, D.; Singh, K. Biotechnology approaches to overcome biotic and abiotic stress constraints in legumes. Euphytica 2006, 147, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Sathya, A.; Vijayabharathi, R.; Gopalakrishnan, S. Plant growth-promoting actinobacteria: A new strategy for enhancing sustainable production and protection of grain legumes. 3 Biotech 2017, 7, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldas, D.; Konzen, E.; Recchia, G.; Pereira, A.; Tsai, S. Functional genomics of biotic and abiotic stresses in Phaseolus vulgaris. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; Shanker, A., Shanker, C., Eds.; IntechOpen: London, UK, 2016; p. 151. ISBN 978-953-51-2250-0. [Google Scholar]
- Hussain, B. Modernization in plant breeding approaches for improving biotic stress resistance in crop plants. Turk. J. Agric. For. 2015, 39, 515–530. [Google Scholar] [CrossRef]
- Zivy, M.; Wienkoop, S.; Renaut, J.; Pinheiro, C.; Goulas, E.; Carpentier, S. The quest for tolerant varieties: The importance of integrating “Omics” techniques to phenotyping. Front. Plant Sci. 2015, 6, 448. [Google Scholar] [CrossRef] [Green Version]
- Saed-Moucheshi, A.; Pessarakli, M.; Mozafari, A.; Sohrabi, F.; Moradi, M.; Marvasti, F. Screening barley varieties tolerant to drought stress based on tolerant indices. J. Plant Nutr. 2022, 45, 739–750. [Google Scholar] [CrossRef]
- Khanal, U.; Wilson, C.; Rahman, S.; Lee, B.; Hoang, V. Smallholder farmers’ adaptation to climate change and its potential contribution to UN’s sustainable development goals of zero hunger and no poverty. J. Clean. Prod. 2021, 281, 124999. [Google Scholar] [CrossRef]
- Jacob, C.; Carrasco, B.; Schwember, A. Advances in breeding and biotechnology of legume crops. Plant Cell Tissue Organ Cult. 2016, 127, 561–584. [Google Scholar] [CrossRef]
- Pratap, A.; Das, A.; Kumar, S.; Gupta, S. Current perspectives on introgression breeding in food legumes. Front. Plant Sci. 2021, 11, 589189. [Google Scholar] [CrossRef]
- Singh, D.; Chaudhary, P.; Taunk, J.; Singh, C.; Singh, D.; Tomar, R.; Aski, M.; Konjengbam, N.; Raje, R.; Singh, S. Fab advances in fabaceae for abiotic stress resilience: From “Omics” to artificial intelligence. Int. J. Mol. Sci. 2021, 22, 535. [Google Scholar] [CrossRef]
- Afzal, M.; Alghamdi, S.; Migdadi, H.; Khan, M.; Nurmansyah; Mirza, S.; El-Harty, E. Legume genomics and transcriptomics: From classic breeding to modern technologies. Saudi J. Biol. Sci. 2020, 27, 543–555. [Google Scholar] [CrossRef]
- Varshney, R.; Dubey, A. Novel genomic tools and modern genetic and breeding approaches for crop improvement. J. Plant Biochem. Biotechnol. 2009, 18, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Yang, L.; Zhang, D.; Shi, J. Plant metabolomics: An indispensable system biology tool for plant science. Int. J. Mol. Sci. 2016, 17, 767. [Google Scholar] [CrossRef] [PubMed]
- Lugan, R.; Niogret, M.; Leport, L.; Guegan, J.; Larher, F.; Savoure, A.; Kopka, J.; Bouchereau, A. Metabolome and water homeostasis analysis of Thellungiella salsuginea suggests that dehydration tolerance is a key response to osmotic stress in this halophyte. Plant J. 2010, 64, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Heyneke, E.; Watanabe, M.; Erban, A.; Duan, G.; Buchner, P.; Walther, D.; Kopka, J.; Hawkesford, M.; Hoefgen, R. Characterization of the wheat leaf metabolome during grain filling and under varied N-supply. Front. Plant Sci. 2017, 8, 2048. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Liu, Y.; Zhang, H.; Wang, J.; Chen, Z.; Luo, L.; Liu, G.; Liu, P. Metabolic alterations provide insights into stylosanthes roots responding to phosphorus deficiency. BMC Plant Biol. 2020, 20, 85. [Google Scholar] [CrossRef] [Green Version]
- Joshi, J.; Hasnain, G.; Logue, T.; Lynch, M.; Wu, S.; Guan, J.; Alseekh, S.; Fernie, A.; Hanson, A.; McCarty, D. A core metabolome response of maize leaves subjected to long-duration abiotic stresses. Metabolites 2021, 11, 797. [Google Scholar] [CrossRef]
- Xing, T.; Jordan, M. Genetic engineering of plant signal transduction mechanisms. Plant Mol. Biol. Report. 2000, 18, 309–318. [Google Scholar] [CrossRef]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant. 2008, 132, 199–208. [Google Scholar] [CrossRef]
- Wolfender, J.-L.; Marti, G.; Thomas, A.; Bertrand, S. Current approaches and challenges for the metabolite profiling of complex natural extracts. J. Chromatogr. A 2015, 1382, 136–164. [Google Scholar] [CrossRef]
- Kumar, A.; Jaiwal, R.; Sreevathsa, R.; Chaudhary, D.; Jaiwal, P. Transgenic cowpea plants expressing Bacillus thuringiensis Cry2Aa insecticidal protein imparts resistance to Maruca vitrata legume pod borer. Plant Cell Rep. 2021, 40, 583–594. [Google Scholar] [CrossRef]
- Singh, S.; Singh, P. Biochemical factors associated with resistance to spotted pod borer, Maruca vitrata (Fabricius) in green gram. Legume Res. 2021, 44, 1398–1401. [Google Scholar] [CrossRef]
- Kamphuis, L.; Zulak, K.; Gao, L.; Anderson, J.; Singh, K. Plant-aphid interactions with a focus on legumes. Funct. Plant Biol. 2013, 40, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Arcos, C.; Kai, M.; Svatos, A.; Gershenzon, J.; Kunert, G. Untargeted metabolomics approach reveals differences in host plant chemistry before and after infestation with different pea aphid host races. Front. Plant Sci. 2019, 10, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, H. Bionomics, host plant resistance, and management of the legume pod borer, Maruca Vitrata—A review. Crop Prot. 1998, 17, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Pobozniak, M. The occurrence of thrips (Thysanoptera) on food legumes (Fabaceae). J. Plant Dis. Prot. 2011, 118, 185–193. [Google Scholar] [CrossRef]
- Singh, C.; Singh, N. Occurrence of insect-pests infesting cowpea (Vigna unguiculata walpers) and their natural enemy complex in associations with weather variables. Legume Res. 2014, 37, 658–664. [Google Scholar] [CrossRef]
- Kalyan, R.; Ameta, O. Impact of abiotic factors on seasonal incidence of insect pests of soybean. Legume Res. 2017, 40, 762–767. [Google Scholar] [CrossRef]
- Rajawat, I.; Kumar, A.; Alam, M.; Tiwari, R.; Pandey, A. Insect pests of black gram (Vigna mungo L.) and their management in Vindhya region. Legume Res. 2021, 44, 225–232. [Google Scholar] [CrossRef]
- Ahmed, K.; Awan, M. Integrated management of insect pests of chickpea Cicer arietinum (L. Walp) in South Asian countries: Present status and future strategies—A review. Pak. J. Zool. 2013, 45, 1125–1145. [Google Scholar]
- Abudulai, M.; Kusi, F.; Seini, S.; Seidu, A.; Nboyine, J.; Larbi, A. Effects of planting date, cultivar and insecticide spray application for the management of insect pests of cowpea in Northern Ghana. Crop Prot. 2017, 100, 168–176. [Google Scholar] [CrossRef]
- Mofokeng, M.; Gerrano, A. Efforts in breeding cowpea for aphid resistance: A review. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2021, 71, 489–497. [Google Scholar] [CrossRef]
- Kooner, B.; Malhi, B.; Cheema, H. Insect Pest Management of Mungbean; Shanmugasundaram, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 214–235. [Google Scholar]
- Edwards, O.; Singh, K. Resistance to insect pests: What do legumes have to offer? Euphytica 2006, 147, 273–285. [Google Scholar] [CrossRef]
- Van Dam, N.; van der Meijden, E. A role for metabolomics in plant ecology. In Biology of Plant Metabolomics; Hall, R., Ed.; Wiley: Hoboken, NJ, USA, 2011; Volume 43, pp. 87–107. ISBN 1460-1494. [Google Scholar]
- Wang, Y.; Liu, Q.; Du, L.; Hallerman, E.; Li, Y. Transcriptomic and metabolomic responses of rice plants to Cnaphalocrocis medinalis caterpillar infestation. Insects 2020, 11, 705. [Google Scholar] [CrossRef] [PubMed]
- Kusnierczyk, A.; Winge, P.; Jorstad, T.; Troczynska, J.; Rossiter, J.; Bones, A. Towards global understanding of plant defence against aphids—Timing and dynamics of early arabidopsis defence responses to cabbage aphid (brevicoryne brassicae) attack. Plant Cell Environ. 2008, 31, 1097–1115. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Q.; Tan, Y.; Shuang, S.; Dai, R.; Jiang, X.; Temuer, B. Combined transcriptome and metabolome analysis of alfalfa response to thrips infection. Genes 2021, 12, 1967. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.V.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and challenges in legume breeding for pest and disease resistance. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef] [Green Version]
- Mgbechi-Ezeri, J.; Porter, L.; Johnson, K.; Oraguzie, N. Assessment of sweet cherry (Prunus avium L.) genotypes for response to bacterial canker disease. Euphytica 2017, 213, 145. [Google Scholar] [CrossRef] [Green Version]
- Obasa, K.; Haynes, L. Two new bacterial pathogens of peanut, causing early seedling decline disease, identified in the Texas Panhandle. Plant Dis. 2022, 106, 648–653. [Google Scholar] [CrossRef]
- Chen, N.; Ruh, M.; Darrasse, A.; Foucher, J.; Briand, M.; Costa, J.; Studholme, D.; Jacques, M. Common bacterial blight of bean: A model of seed transmission and pathological convergence. Mol. Plant Pathol. 2021, 22, 1464–1480. [Google Scholar] [CrossRef]
- Tugume, J.; Tusiime, G.; Sekamate, A.; Buruchara, R.; Mukankusi, C. Diversity and interaction of common bacterial blight disease-causing bacteria (Xanthomonas spp.) with Phaseolus vulgaris L. Crop J. 2019, 7, 1–7. [Google Scholar] [CrossRef]
- Osdaghi, E.; Young, A.; Harveson, R. Bacterial wilt of dry beans caused by Curtobacterium flaccumfaciens pv. flaccumfaciens: A new threat from an old enemy. Mol. Plant Pathol. 2020, 21, 605–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaghefi, N.; Adorada, D.; Huth, L.; Kelly, L.; Poudel, B.; Young, A.; Sparks, A. Whole-genome data from Curtobacterium flaccumfaciens pv. flaccumfaciens strains associated with tan spot of mungbean and soybean reveal diverse plasmid profiles. Mol. Plant. Microbe Interact. 2021, 34, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.; Pandey, A.; War, A.; Hanumantharao, B.; Shwe, T.; Alam, A.; Pratap, A.; Malik, S.; Karimi, R.; Mbeyagala, E. Biotic and abiotic constraints in mungbean production-progress in genetic improvement. Front. Plant Sci. 2019, 10, 1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.; Yuste-Lisbona, F.; Fernandez-Lozano, A.; Lozano, R.; Santalla, M. Genetic mapping and QTL analysis in common bean. In Common Bean Genome; DeLaVega, M., Santalla, M., Marsolais, F., Eds.; Springer: Cham, Switzerland, 2017; pp. 69–107. ISBN 2199-4781. [Google Scholar]
- Padhi, E.; Maharaj, N.; Lin, S.; Mishchuk, D.; Chin, E.; Godfrey, K.; Foster, E.; Polek, M.; Leveau, J.; Slupsky, C. Metabolome and microbiome signatures in the roots of citrus affected by huanglongbing. Phytopathology 2019, 109, 2022–2032. [Google Scholar] [CrossRef]
- Andam, A.; Azizi, A.; Majdi, M.; Abdolahzadeh, J. Comparative expression profile of some putative resistance genes of chickpea genotypes in response to ascomycete fungus, Ascochyta rabiei (Pass.) Labr. Braz. J. Bot. 2020, 43, 123–130. [Google Scholar] [CrossRef]
- Gerlin, L.; Escourrou, A.; Cassan, C.; Macia, F.; Peeters, N.; Genin, S.; Baroukh, C. Unravelling physiological signatures of tomato bacterial wilt and xylem metabolites exploited by Ralstonia solanacearum. Environ. Microbiol. 2021, 23, 5962–5978. [Google Scholar] [CrossRef]
- Pandey, A.; Burlakoti, R.; Kenyon, L.; Nair, R. Perspectives and challenges for sustainable management of fungal diseases of mungbean [Vigna radiata (L.) R. wilczek var. radiata]: A review. Front. Environ. Sci. 2018, 6, 53. [Google Scholar] [CrossRef]
- Kowalska, B. Management of the soil-borne fungal pathogen—Verticillium dahliae Kleb. causing vascular wilt diseases. J. Plant Pathol. 2021, 103, 1185–1194. [Google Scholar] [CrossRef]
- Anderson, J.; Lichtenzveig, J.; Oliver, R.; Singh, K. Medicago truncatula as a model host for studying legume infecting Rhizoctonia solani and identification of a locus affecting resistance to root canker. Plant Pathol. 2013, 62, 908–921. [Google Scholar] [CrossRef]
- Infantino, A.; Kharrat, M.; Riccioni, L.; Coyne, C.; McPhee, K.; Grunwald, N. Screening techniques and sources of resistance to root diseases in cool season food legumes. Euphytica 2006, 147, 201–221. [Google Scholar] [CrossRef]
- Marzouk, T.; Chaouachi, M.; Sharma, A.; Jallouli, S.; Mhamdi, R.; Kaushik, N.; Djebali, N. Biocontrol of Rhizoctonia solani using volatile organic compounds of solanaceae seed-borne endophytic bacteria. Postharvest Biol. Technol. 2021, 181, 111655. [Google Scholar] [CrossRef]
- Wang, H.; Chang, K.; Hwang, S.; Gossen, B.; Turnbull, G.; Howard, R.; Strelkov, S. Response of lentil cultivars to Rhizoctonia seedling diseases in Canada. J. Plant Dis. Prot. 2006, 113, 219–223. [Google Scholar] [CrossRef]
- Wolfgang, A.; Taffner, J.; Guimarães, R.A.; Coyne, D.; Berg, G. Novel strategies for soil-borne diseases: Exploiting the microbiome and volatile-based mechanisms toward controlling Meloidogyne-based disease complexes. Front. Microbiol. 2019, 10, 1296. [Google Scholar] [CrossRef] [PubMed]
- Willsey, T.; Chatterton, S.; Heynen, M.; Erickson, A. Detection of interactions between the pea root rot pathogens Aphanomyces euteiches and Fusarium spp. using a multiplex QPCR assay. Plant Pathol. 2018, 67, 1912–1923. [Google Scholar] [CrossRef]
- Diaz, L.; Arredondo, V.; Ariza-Suarez, D.; Aparicio, J.; Buendia, H.; Cajiao, C.; Mosquera, G.; Beebe, S.; Mukankusi, C.; Raatz, B. Genetic analyses and genomic predictions of root rot resistance in common bean across trials and populations. Front. Plant Sci. 2021, 12, 629221. [Google Scholar] [CrossRef] [PubMed]
- Gaulin, E.; Jacquet, C.; Bottin, A.; Dumas, B. Root rot disease of legumes caused by Aphanomyces euteiches. Mol. Plant Pathol. 2007, 8, 539–548. [Google Scholar] [CrossRef]
- Wu, L.; Chang, K.; Conner, R.; Strelkov, S.; Fredua-Agyeman, R.; Hwang, S.; Feindel, D. Aphanomyces euteiches: A threat to Canadian field pea production. Engineering 2018, 4, 542–551. [Google Scholar] [CrossRef]
- Smolinska, U.; Kowalska, B. Biological control of the soil-borne fungal pathogen Sclerotinia sclerotiorum—A review. J. Plant Pathol. 2018, 100, 1–12. [Google Scholar] [CrossRef] [Green Version]
- You, M.; Lamichhane, J.; Aubertot, J.; Barbetti, M. Understanding why effective fungicides against individual soilborne pathogens are ineffective with soilborne pathogen complexes. Plant Dis. 2020, 104, 904–920. [Google Scholar] [CrossRef]
- Bilkiss, M.; Shiddiky, M.J.A.; Ford, R. Advanced diagnostic approaches for necrotrophic fungal pathogens of temperate legumes with a focus on Botrytis spp. Front. Microbiol. 2019, 10, 1889. [Google Scholar] [CrossRef]
- Kisiel, A. Biological control as an alternative method of protecting crops against fungal pathogens. Rocz. Ochr. Srodowiska 2019, 21, 1366–1377. [Google Scholar]
- Chandler, D.; Bailey, A.; Tatchell, G.; Davidson, G.; Greaves, J.; Grant, W. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B—Biol. Sci. 2011, 366, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- Quiroz, R.; Maldonado, J.; Alanis, M.; Torres, J.; Saldivar, R. Fungi-based biopesticides: Shelf-life preservation technologies used in commercial products. J. Pest Sci. 2019, 92, 1003–1015. [Google Scholar] [CrossRef]
- Harveson, R. Improving yields and managing dry bean bacterial diseases in Nebraska with new copper-alternative chemicals. Plant Health Prog. 2019, 20, 14–19. [Google Scholar] [CrossRef]
- Chen, L.; Wu, Q.; He, T.; Lan, J.; Ding, L.; Liu, T.; Wu, Q.; Pan, Y.; Chen, T. Transcriptomic and metabolomic changes triggered by Fusarium solani in common bean (Phaseolus vulgaris L.). Genes 2020, 11, 177. [Google Scholar] [CrossRef] [Green Version]
- Mayo-Prieto, S.; Marra, R.; Vinale, F.; Rodríguez-González, Á.; Woo, S.L.; Lorito, M.; Gutiérrez, S.; Casquero, P.A. Effect of Trichoderma velutinum and Rhizoctonia solani on the metabolome of bean plants (Phaseolus vulgaris L.). Int. J. Mol. Sci. 2019, 20, 549. [Google Scholar] [CrossRef] [Green Version]
- Copley, T.; Aliferis, K.; Kliebenstein, D.; Jabaji, S. An integrated RNAseq-H-1 NMR metabolomics approach to understand soybean primary metabolism regulation in response to Rhizoctonia foliar blight disease. BMC Plant Biol. 2017, 17, 84. [Google Scholar] [CrossRef]
- Turetschek, R.; Desalegn, G.; Epple, T.; Kaul, H.; Wienkoop, S. Key metabolic traits of Pisum sativum maintain cell vitality during Didymella pinodes infection: Cultivar resistance and the microsymbionts’ influence. J. Proteom. 2017, 169, 189–201. [Google Scholar] [CrossRef]
- Pérez-Clemente, R.M.; Montoliu, A.; Vives-Peris, V.; Arbona, V.; Gómez-Cadenas, A. Hormonal and metabolic responses of mexican lime plants to CTV infection. J. Plant Physiol. 2019, 238, 40–52. [Google Scholar] [CrossRef]
- Srivastava, S.; Bisht, H.; Sidhu, O.; Srivastava, A.; Singh, P.; Pandey, R.; Raj, S.; Roy, R.; Nautiyal, C. Changes in the metabolome and histopathology of Amaranthus hypochondriacus L. in response to Ageratum enation Virus Infection. Phytochemistry 2012, 80, 8–16. [Google Scholar] [CrossRef]
- Mutuku, J.; Cui, S.; Hori, C.; Takeda, Y.; Tobimatsu, Y.; Nakabayashi, R.; Mori, T.; Saito, K.; Demura, T.; Umezawa, T. The structural integrity of lignin is crucial for resistance against Striga hermonthica parasitism in rice. Plant Physiol. 2019, 179, 1796–1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, W.; Zhu, X.; Wang, Y.; Chen, L.; Duan, Y. Transcriptomic and metabolomic analyses reveal that bacteria promote plant defense during infection of soybean cyst nematode in soybean. BMC Plant Biol. 2018, 18, 98. [Google Scholar] [CrossRef] [Green Version]
- Narula, K.; Elagamey, E.; Abdellatef, M.; Sinha, A.; Ghosh, S.; Chakraborty, N.; Chakraborty, S. Chitosan-triggered immunity to Fusarium in chickpea is associated with changes in the plant extracellular matrix architecture, stomatal closure and remodeling of the plant metabolome and proteome. Plant J. 2020, 103, 561–583. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Sawada, Y.; Ochiai, K.; Sato, M.; Inaba, J.; Hirai, M. Identification of a unique type of isoflavone O-methyltransferase, GmIOMT1, based on multi-Omics analysis of soybean under biotic stress. Plant Cell Physiol. 2020, 61, 1974–1985. [Google Scholar] [CrossRef] [PubMed]
- Sistani, N.; Kaul, H.; Desalegn, G.; Wienkoop, S. Rhizobium impacts on seed productivity, quality, and protection of Pisum sativum upon disease stress caused by Didymella pinodes: Phenotypic, proteomic, and metabolomic traits. Front. Plant Sci. 2017, 8, 1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makkouk, K.M. Plant pathogens which threaten food security: Viruses of chickpea and other cool season legumes in West Asia and North Africa. Food Secur. 2020, 12, 495–502. [Google Scholar] [CrossRef]
- Wood, C.; Pilkington, B.; Vaidya, P.; Biel, C.; Stinchcombel, J. Genetic conflict with a parasitic nematode disrupts the legume-rhizobia mutualism. Evol. Lett. 2018, 2, 233–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, E.L.; Mitchum, M.G. Nematodes. sophisticated parasites of legumes. Plant Physiol. 2005, 137, 1182–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abo, M.; Alegbejo, M. Strategies for sustainable control of viral diseases of some crops in Nigeria. J. Sustain. Agric. 1997, 10, 57–80. [Google Scholar] [CrossRef]
- Sauerborn, J.; Muller-Stover, D.; Hershenhorn, J. The role of biological control in managing parasitic weeds. Crop Prot. 2007, 26, 246–254. [Google Scholar] [CrossRef]
- Rodenburg, J.; Demont, M.; Zwart, S.; Bastiaans, L. Parasitic weed incidence and related economic losses in rice in Africa. Agric. Ecosyst. Environ. 2016, 235, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Hooper, A.; Hassanali, A.; Chamberlain, K.; Khan, Z.; Pickett, J. New genetic opportunities from legume intercrops for controlling Striga spp. parasitic weeds. Pest Manag. Sci. 2009, 65, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Perez-de-Luque, A.; Flores, F.; Rubiales, D. Differences in crenate broomrape parasitism dynamics on three legume crops using a thermal time model. Front. Plant Sci. 2016, 7, 1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubiales, D.; Flores, F.; Emeran, A.; Kharrat, M.; Amri, M.; Rojas-Molina, M.; Sillero, J. Identification and multi-environment validation of resistance against broomrapes (Orobanche crenata and Orobanche foetida) in faba bean (Vicia faba). Field Crops Res. 2014, 166, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Arora, P.; Yadav, R.; Dilbaghi, N.; Chaudhury, A. Biological nitrogen fixation: Host-rhizobium interaction. In Frontiers on Recent Developments in Plant Science; Goyal, A., Maheshwari, P., Eds.; Bentham Books: Sharjah, United Arab Emirates, 2012; Volume 1, pp. 39–59. ISBN 2589-1464. [Google Scholar]
- Hassan, S.; Behm, C.; Mathesius, U. Effectors of plant parasitic nematodes that re-program root cell development. Funct. Plant Biol. 2010, 37, 933–942. [Google Scholar] [CrossRef]
- Steinfath, M.; Strehmel, N.; Peters, R.; Schauer, N.; Groth, D.; Hummel, J.; Steup, M.; Selbig, J.; Kopka, J.; Geigenberger, P. Discovering plant metabolic biomarkers for phenotype prediction using an untargeted approach. Plant Biotechnol. J. 2010, 8, 900–911. [Google Scholar] [CrossRef]
- Hirayama, A.; Wakayama, M.; Soga, T. Metabolome analysis based on capillary electrophoresis-mass spectrometry. TrAC—Trends Anal. Chem. 2014, 61, 215–222. [Google Scholar] [CrossRef]
- Patel, M.; Pandey, S.; Kumar, M.; Haque, M.; Pal, S.; Yadav, N. Plants metabolome study: Emerging tools and techniques. Plants 2021, 10, 2409. [Google Scholar] [CrossRef]
- Junot, C.; Fenaille, F.; Colsch, B.; Becher, F. High resolution mass spectrometry based techniques at the crossroads of metabolic pathways. Mass Spectrom. Rev. 2014, 33, 471–500. [Google Scholar] [CrossRef]
- Sun, X.; Weckwerth, W. COVAIN: A toolbox for uni- and multivariate statistics, time-series and correlation network analysis and inverse estimation of the differential Jacobian from metabolomics covariance data. Metabolomics 2012, 8, 81–93. [Google Scholar] [CrossRef]
- Ghatak, A.; Chaturvedi, P.; Weckwerth, W. Metabolomics in plant stress physiology. In Plant Genetics and Molecular Biology; Varshney, R., Pandey, M., Chitikineni, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 164, pp. 187–236. ISBN 0724-6145. [Google Scholar]
- Fukushima, A.; Kusano, M. Recent progress in the development of metabolome databases for plant systems biology. Front. Plant Sci. 2013, 4, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, D.; Szymanski, J.; Erban, A.; Udvardi, M.; Kopka, J. Mining for robust transcriptional and metabolic responses to long-term salt stress: A case study on the model legume Lotus japonicus. Plant Cell Environ. 2010, 33, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, D.; Schwabe, F.; Erban, A.; Udvardi, M.; Kopka, J. Comparative metabolomics of drought acclimation in model and forage legumes. Plant Cell Environ. 2012, 35, 136–149. [Google Scholar] [CrossRef]
- Sanchez, D.; Pieckenstain, F.; Escaray, F.; Erban, A.; Kraemer, U.; Udvardi, M.; Kopka, J. Comparative ionomics and metabolomics in extremophile and glycophytic Lotus species under salt stress challenge the metabolic pre-adaptation hypothesis. Plant Cell Environ. 2011, 34, 605–617. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Flow diagram summarizing steps taken for metabolomic sample analysis in biotic stress experiments. Plant under biotic stress (A), samples from selected plant parts in a tube (B), snap freezing samples in liquid nitrogen and later stored in an ultra-freezer (C), extraction of metabolites in accordance with recommended protocols (D), metabolome analysis technologies (E), generation of raw spectral data (F).
Figure 1.
Flow diagram summarizing steps taken for metabolomic sample analysis in biotic stress experiments. Plant under biotic stress (A), samples from selected plant parts in a tube (B), snap freezing samples in liquid nitrogen and later stored in an ultra-freezer (C), extraction of metabolites in accordance with recommended protocols (D), metabolome analysis technologies (E), generation of raw spectral data (F).

Figure 2.
Flow diagram illustrating data handling steps for metabolomic experiments. After acquiring raw data, pre-processing, pre-treatment and statistical analysis are required prior to interpretation of results.
Figure 2.
Flow diagram illustrating data handling steps for metabolomic experiments. After acquiring raw data, pre-processing, pre-treatment and statistical analysis are required prior to interpretation of results.

Table 2.
Statistical tools and databases used for metabolome data processing and annotation in legume biotic stress studies.
Table 2.
Statistical tools and databases used for metabolome data processing and annotation in legume biotic stress studies.
Table 3.
Statistical tools and databases used for metabolome data processing and annotation in legume biotic stress studies.
Table 3.
Statistical tools and databases used for metabolome data processing and annotation in legume biotic stress studies.
| Legume | Statistical Tool/Database Name | Access Domain (URL, Accessed on 28 April 2022) | Function | Reference |
|---|---|---|---|---|
| L. japonicus | MapMan/PageMan | https://mapman.gabipd.org/mapman | Data processing Metabolite annotation | [108,109] |
| MeV | https://mev.tm4.org/#/about | Data processing Metabolite annotation | ||
| Microsoft Excel | https://www.microsoft.com/en-za/ | Data processing | ||
| MetaGeneAlyse | https://metagenealyse.mpimp-golm.mpg.de/ | Data processing Metabolite annotation | ||
| L. corniculatus L. creticus L. tenius L. burttii L. uligino L. filicaulis | GRaphPad (Prism) | https://www.graphpad.com/ | Data processing | [110] |
| MeV | https://mev.tm4.org/#/about | Data processing Metabolite annotation | ||
| MetaGeneAlyse | https://metagenealyse.mpimp-golm.mpg.de/ | Data processing Metabolite annotation | ||
| Microsoft Excel | https://www.microsoft.com/en-za/ | Data processing | ||
| Stylosanthes | Microsoft Excel | https://www.microsoft.com/en-za/ | Data processing | [26] |
| SPSS | https://www.ibm.com/products/spss-statistics | Data processing | ||
| R | https://www.r-project.org/ | Data processing | ||
| KEGG | https://www.genome.jp/kegg/kegg2.html | Metabolomic pathways | ||
| P. vulgaris | MapMan | https://mapman.gabipd.org/mapman | Data processing and Metabolite annotation | [80] |
| KEGG | https://www.genome.jp/kegg/kegg2.html | Metabolomic pathways |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Makhumbila, P.; Rauwane, M.; Muedi, H.; Figlan, S. Metabolome Profiling: A Breeding Prediction Tool for Legume Performance under Biotic Stress Conditions. Plants 2022, 11, 1756. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11131756
AMA Style
Makhumbila P, Rauwane M, Muedi H, Figlan S. Metabolome Profiling: A Breeding Prediction Tool for Legume Performance under Biotic Stress Conditions. Plants. 2022; 11(13):1756. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11131756
Chicago/Turabian StyleMakhumbila, Penny, Molemi Rauwane, Hangwani Muedi, and Sandiswa Figlan. 2022. "Metabolome Profiling: A Breeding Prediction Tool for Legume Performance under Biotic Stress Conditions" Plants 11, no. 13: 1756. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11131756
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.