
Pepper Plants Harboring L Resistance Alleles Showed Tolerance toward Manifestations of Tomato Brown Rugose Fruit Virus Disease


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
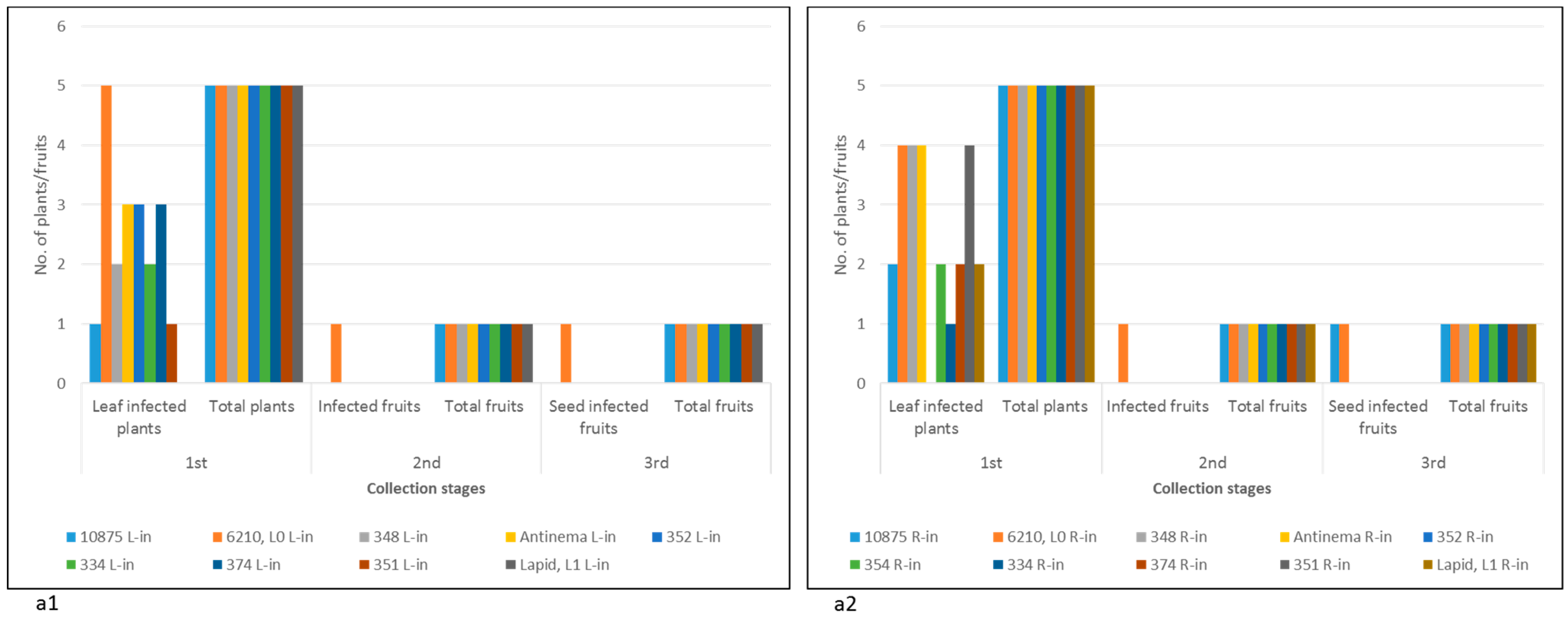
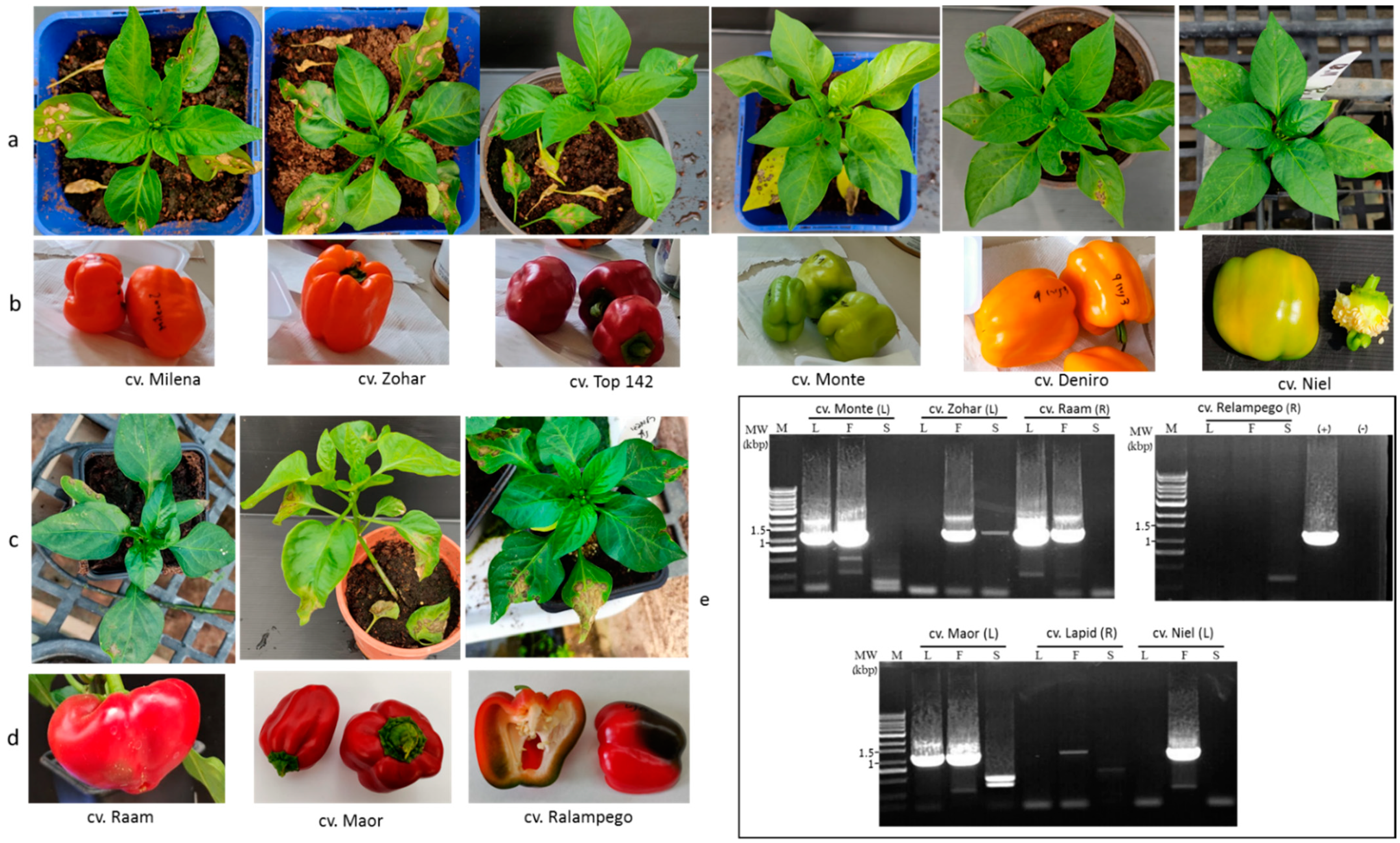
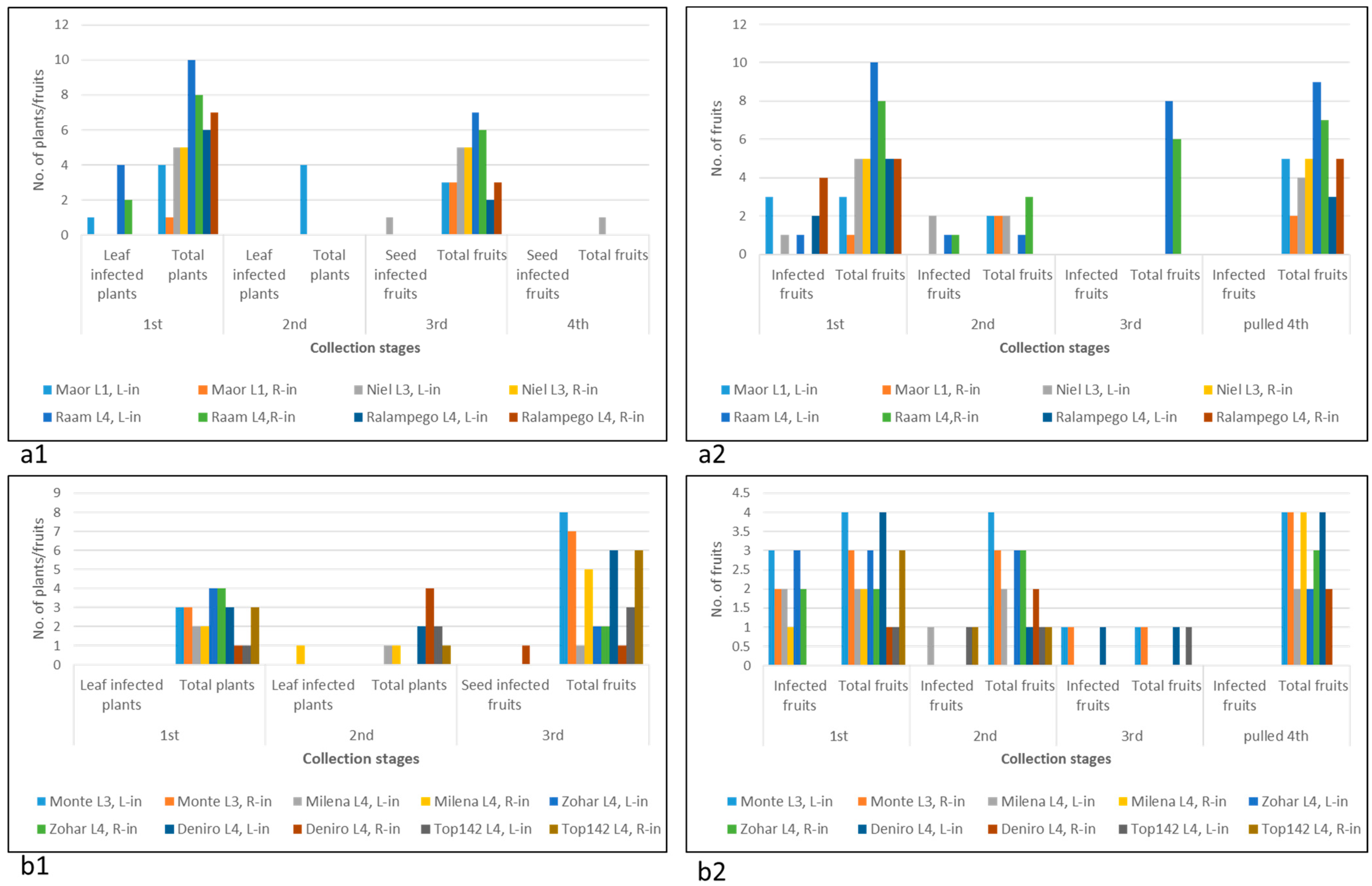
2.1. Susceptibility of Multiple Pepper Plant Varieties to ToBRFV Infection Was Not Tightly Associated with Disease Manifestations
2.2. Pepper Plants Harboring L1, L3, or L4 Resistance Alleles That Manifest HRs Are Susceptible to ToBRFV Systemic Infection but Show Tolerance toward the Disease
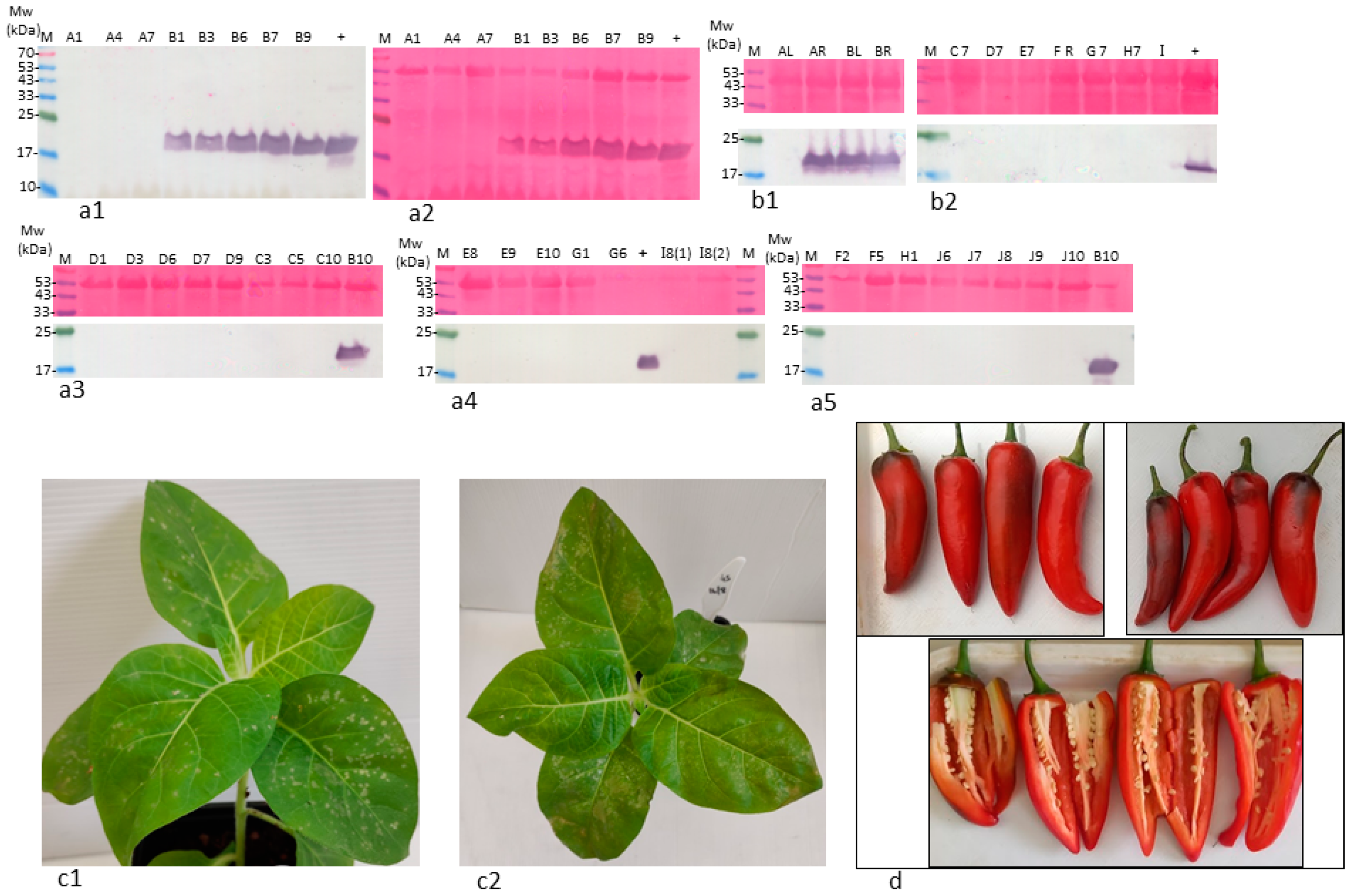
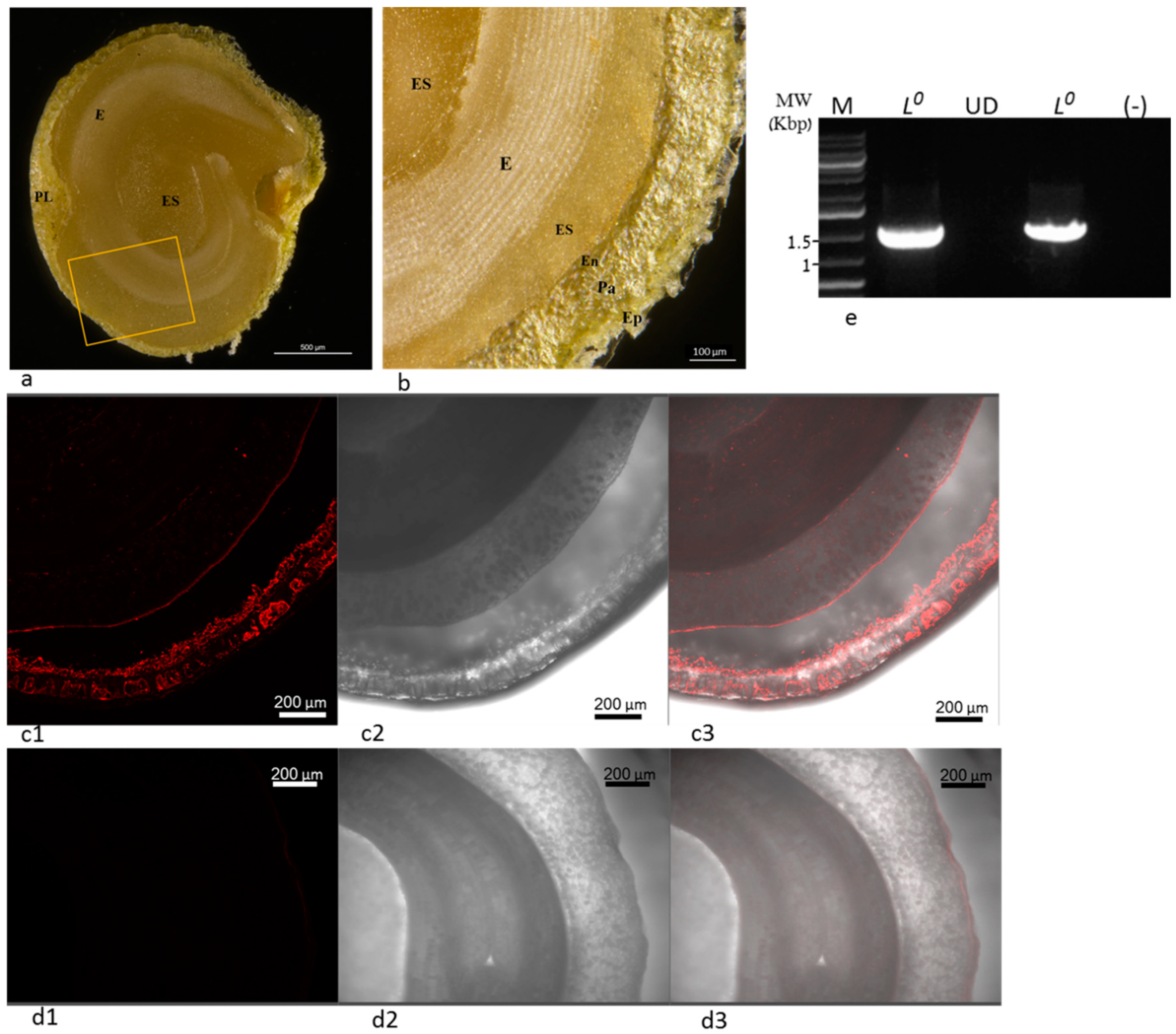
2.3. ToBRFV in Stably Infected L0 Pepper Plants cv. 6210 Was Infectious
3. Discussion
4. Materials and Methods
4.1. Tested Plants, ToBRFV Inoculations, and Biological Assays for Infectious Virus
4.2. Indirect Enzyme-Linked Immunosorbent Assay (ELISA)
4.3. Western Blot Analysis
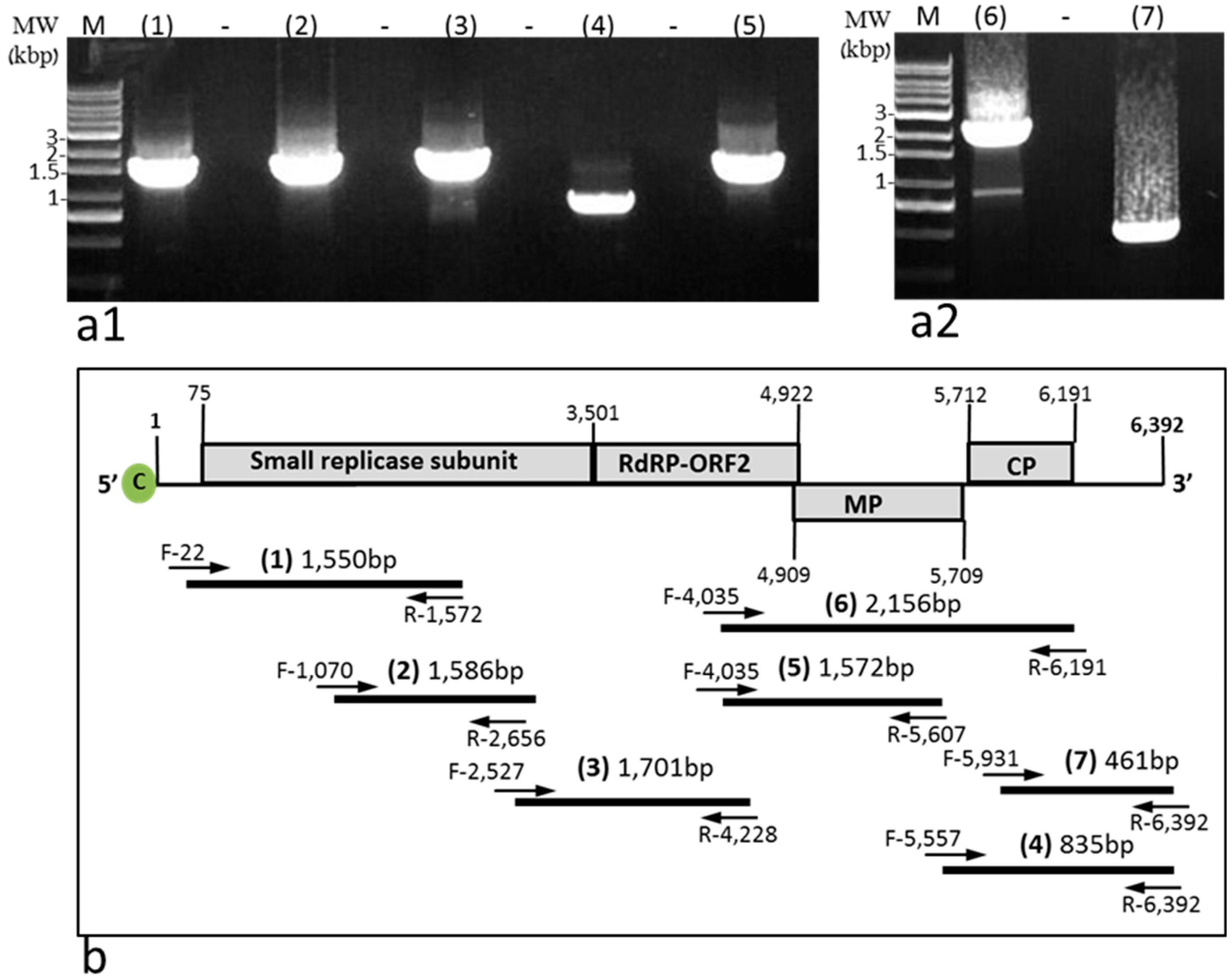
4.4. Reverse Transcription (RT)-PCR
4.5. In Situ Immunofluorescence
5. Conclusions
- Both foliar- and root-inoculated plants harboring the L alleles were susceptible to a transient ToBRFV infection, which was not associated with symptomatic fruits.
- A severe cell death response and wilting occurred in one foliar-inoculated undefined cultivar (cv. 354) that showed systemic necrosis and did not reach the fruiting stage. The root-inoculated plants of this cultivar had asymptomatic fruits.
- A stable systemic ToBRFV infection occurred in defined L0 pepper plants, in which the virus was localized to the maternal origin of the seed tissues (epidermis, parenchyma, and endothelium). ToBRFV in the L0 plants was infectious and highly similar to the original ToBRFV sequence from tomato plants, indicating low constraints were imposed on ToBRFV by the L0 pepper host. Similar to L gene-resistant plants, there was a dissociation between ToBRFV systemic infection of the L0 plants and disease symptoms on fruits.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balint-Kurti, P. The plant hypersensitive response: Concepts, control and consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef] [PubMed]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Niu, F.; Liu, W.-Z.; Yang, B.; Zhang, J.; Ma, J.; Cheng, H.; Han, F.; Jiang, Y.-Q. Identification, cloning and characterization of R2R3-MYB gene family in canola (Brassica napus L.) identify a novel member modulating ROS accumulation and hypersensitive-like cell death. DNA Res. 2016, 23, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Jockusch, H. The role of host genes, temperature and polyphenoloxidase in the necrotization of TMV infected tobacco tissue. J. Phytopathol. 1966, 55, 185–192. [Google Scholar] [CrossRef]
- Whitham, S.; McCormick, S.; Baker, B. The N gene of tobacco confers resistance to tobacco mosaic virus in transgenic tomato. Proc. Natl. Acad. Sci. USA 1996, 93, 8776–8781. [Google Scholar] [CrossRef]
- Lanfermeijer, F.C.; Jiang, G.; Ferwerda, M.A.; Dijkhuis, J.; de Haan, P.; Yang, R.; Hille, J. The durable resistance gene Tm-22 from tomato confers resistance against ToMV in tobacco and preserves its viral specificity. Plant Sci. 2004, 167, 687–692. [Google Scholar] [CrossRef]
- Nagai, A.; Duarte, L.M.; Chaves, A.L.; Peres, L.E.; dos Santos, D.Y. Tomato mottle mosaic virus in Brazil and its relationship with Tm-22 gene. Eur. J. Plant Pathol. 2019, 155, 353–359. [Google Scholar] [CrossRef]
- Salem, N.; Mansour, A.; Ciuffo, M.; Falk, B.; Turina, M. A new tobamovirus infecting tomato crops in Jordan. Arch. Virol. 2016, 161, 503–506. [Google Scholar] [CrossRef]
- Luria, N.; Smith, E.; Reingold, V.; Bekelman, I.; Lapidot, M.; Levin, I.; Elad, N.; Tam, Y.; Sela, N.; Abu-Ras, A.; et al. A New Israeli Tobamovirus Isolate Infects Tomato Plants Harboring Tm-22 Resistance Genes. PLoS ONE 2017, 12, e0170429. [Google Scholar] [CrossRef]
- Zhang, S.; Griffiths, J.S.; Marchand, G.; Bernards, M.A.; Wang, A. Tomato brown rugose fruit virus: An emerging and rapidly spreading plant RNA virus that threatens tomato production worldwide. Mol. Plant Pathol. 2022, 23, 1262–1277. [Google Scholar] [CrossRef]
- Vogler, H.; Akbergenov, R.; Shivaprasad, P.V.; Dang, V.; Fasler, M.; Kwon, M.-O.; Zhanybekova, S.; Hohn, T.; Heinlein, M. Modification of small RNAs associated with suppression of RNA silencing by tobamovirus replicase protein. J. Virol. 2007, 81, 10379–10388. [Google Scholar] [CrossRef] [PubMed]
- Panno, S.; Caruso, A.; Blanco, G.; Davino, S. First report of Tomato brown rugose fruit virus infecting sweet pepper in Italy. New Dis. Rep. 2020, 41, 20. [Google Scholar] [CrossRef]
- Rizzo, D.; Da Lio, D.; Panattoni, A.; Salemi, C.; Cappellini, G.; Bartolini, L.; Parrella, G. Rapid and sensitive detection of tomato brown rugose fruit virus in tomato and pepper seeds by reverse transcription loop-mediated isothermal amplification assays (real time and visual) and comparison with RT-PCR end-point and RT-qPCR methods. Front. Microbiol. 2021, 12, 640932–640944. [Google Scholar] [CrossRef] [PubMed]
- Fidan, H.; Sarikaya, P.; Yildiz, K.; Topkaya, B.; Erkis, G.; Calis, O. Robust molecular detection of the new Tomato brown rugose fruit virus in infected tomato and pepper plants from Turkey. J. Integr. Agric. 2021, 20, 2170–2179. [Google Scholar] [CrossRef]
- Abou Kubaa, R.; Choueiri, E.; Heinoun, K.; Cillo, F.; Saponari, M. First report of tomato brown rugose fruit virus infecting sweet pepper in Syria and Lebanon. J. Plant Pathol. 2022, 104, 425. [Google Scholar] [CrossRef]
- Tomita, R.; Sekine, K.-T.; Mizumoto, H.; Sakamoto, M.; Murai, J.; Kiba, A.; Hikichi, Y.; Suzuki, K.; Kobayashi, K. Genetic basis for the hierarchical interaction between Tobamovirus spp. and L resistance gene alleles from different pepper species. Mol. Plant-Microbe Interact. 2011, 24, 108–117. [Google Scholar] [CrossRef]
- Sawada, H.; Takeuchi, S.; Hamada, H.; Kiba, A.; Matsumoto, M.; Hikichi, Y. A new tobamovirus-resistance gene, L1a, of sweet pepper (Capsicum annuum L.). J. Jpn. Soc. Hortic. Sci. 2004, 73, 552–557. [Google Scholar] [CrossRef]
- Matsumoto, K.; Sawada, H.; Matsumoto, K.; Hamada, H.; Yoshimoto, E.; Ito, T.; Takeuchi, S.; Tsuda, S.; Suzuki, K.; Kobayashi, K. The coat protein gene of tobamovirus P0 pathotype is a determinant for activation of temperature-insensitive L 1a-gene-mediated resistance in Capsicum plants. Arch. Virol. 2008, 153, 645–650. [Google Scholar] [CrossRef]
- Chanda, B.; Gilliard, A.; Jaiswal, N.; Ling, K.-S. Comparative analysis of host range, ability to infect tomato cultivars with Tm-22 gene, and real-time reverse transcription PCR detection of tomato brown rugose fruit virus. Plant Dis. 2021, 105, 3643–3652. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Mor, N.; Gantz, S.; Lachman, O.; Smith, E. Disinfection Efficacy of Tobamovirus-Contaminated Soil in Greenhouse-Grown Crops. Horticulturae 2022, 8, 563. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Molnar, A.; Bassett, A.; Baulcombe, D.C. Mobile 24 nt small RNAs direct transcriptional gene silencing in the root meristems of Arabidopsis thaliana. Curr. Biol. 2011, 21, 1678–1683. [Google Scholar] [CrossRef] [PubMed]
- Andika, I.B.; Kondo, H.; Sun, L. Interplays between soil-borne plant viruses and RNA silencing-mediated antiviral defense in roots. Front. Microbiol. 2016, 7, 1458–1470. [Google Scholar] [CrossRef]
- Genda, Y.; Sato, K.; Nunomura, O.; Hirabayashi, T.; Ohnishi, J.; Tsuda, S. Immunolocalization of Pepper mild mottle virus in Capsicum annuum seeds. J. Gen. Plant Pathol. 2005, 71, 238–242. [Google Scholar] [CrossRef]
- Pagán, I.; García-Arenal, F. Tolerance to plant pathogens: Theory and experimental evidence. Int. J. Mol. Sci. 2018, 19, 810. [Google Scholar] [CrossRef] [PubMed]
- Keen, N.; Ersek, T.; Long, M.; Bruegger, B.; Holliday, M. Inhibition of the hypersensitive reaction of soybean leaves to incompatible Pseudomonas spp. by blasticidin S, streptomycin or elevated temperature. Physiol. Plant Pathol. 1981, 18, 325–337. [Google Scholar] [CrossRef]
- Harder, D.; Samborski, D.; Rohringer, R.; Rimmer, S.; Kim, W.; Chong, J. Electron microscopy of susceptible and resistant near-isogenic (sr6/Sr6) lines of wheat infected by Puccinia graminis tritici. III. Ultrastructure of incompatible interact. Can. J. Bot. 1979, 57, 2626–2634. [Google Scholar] [CrossRef]
- Widana Gamage, S.M.; McGrath, D.J.; Persley, D.M.; Dietzgen, R.G. Transcriptome analysis of Capsicum chlorosis virus-induced hypersensitive resistance response in bell Capsicum. PLoS ONE 2016, 11, e0159085. [Google Scholar] [CrossRef]
- Tsai, W.-A.; Shafiei-Peters, J.R.; Mitter, N.; Dietzgen, R.G. Effects of Elevated Temperature on the Susceptibility of Capsicum Plants to Capsicum Chlorosis Virus Infection. Pathogens 2022, 11, 200. [Google Scholar] [CrossRef]
- Shivaprasad, P.V.; Chen, H.-M.; Patel, K.; Bond, D.M.; Santos, B.A.; Baulcombe, D.C. A microRNA superfamily regulates nucleotide binding site–leucine-rich repeats and other mRNAs. Plant Cell 2012, 24, 859–874. [Google Scholar] [CrossRef]
- Leonetti, P.; Stuttmann, J.; Pantaleo, V. Regulation of plant antiviral defense genes via host RNA-silencing mechanisms. Virol. J. 2021, 18, 194. [Google Scholar] [CrossRef]
- Lopez-Gomollon, S.; Baulcombe, D.C. Roles of RNA silencing in viral and non-viral plant immunity and in the crosstalk between disease resistance systems. Nat. Rev. Mol. Cell Biol. 2022, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Klap, C.; Luria, N.; Smith, E.; Bakelman, E.; Belausov, E.; Laskar, O.; Lachman, O.; Gal-On, A.; Dombrovsky, A. The Potential Risk of Plant-Virus Disease Initiation by Infected Tomatoes. Plants 2020, 9, 623–637. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eldan, O.; Ofir, A.; Luria, N.; Klap, C.; Lachman, O.; Bakelman, E.; Belausov, E.; Smith, E.; Dombrovsky, A. Pepper Plants Harboring L Resistance Alleles Showed Tolerance toward Manifestations of Tomato Brown Rugose Fruit Virus Disease. Plants 2022, 11, 2378. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11182378
Eldan O, Ofir A, Luria N, Klap C, Lachman O, Bakelman E, Belausov E, Smith E, Dombrovsky A. Pepper Plants Harboring L Resistance Alleles Showed Tolerance toward Manifestations of Tomato Brown Rugose Fruit Virus Disease. Plants. 2022; 11(18):2378. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11182378
Chicago/Turabian StyleEldan, Or, Arie Ofir, Neta Luria, Chen Klap, Oded Lachman, Elena Bakelman, Eduard Belausov, Elisheva Smith, and Aviv Dombrovsky. 2022. "Pepper Plants Harboring L Resistance Alleles Showed Tolerance toward Manifestations of Tomato Brown Rugose Fruit Virus Disease" Plants 11, no. 18: 2378. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11182378