

Allelopathy and Allelochemicals of Solidago canadensis L. and S. altissima L. for Their Naturalization

Department of Applied Biological Science, Faculty of Agriculture, Kagawa University, Miki, Kagawa 761-0795, Japan
*
Author to whom correspondence should be addressed.
Plants 2022, 11(23), 3235; https://0-doi-org.brum.beds.ac.uk/10.3390/plants11233235
Submission received: 22 October 2022
/
Revised: 21 November 2022
/
Accepted: 23 November 2022
/
Published: 25 November 2022
(This article belongs to the Special Issue Plant Allelopathy: Mechanisms and Applications in Regenerative Agriculture)
Abstract
:Solidago canadensis L. and Solidago altissima L. are native to North America and have naturalized many other continents including Europa and Asia. Their species is an aggressive colonizer and forms thick monospecific stands. The evidence of the allelopathy for S. canadensis and S. altissima has accumulated in the literature since the late 20th century. The root exudates, extracts, essential oil and rhizosphere soil of S. canadensis suppressed the germination, growth and the arbuscular mycorrhizal colonization of several plants, including native plant species. Allelochemicals such as fatty acids, terpenes, flavonoids, polyphenols and their related compounds were identified in the extracts and essential oil of S. canadensis. The concentrations of total phenolics, total flavonoids and total saponins in the rhizosphere soil of S. canadensis obtained from the invasive ranges were greater than those from the native ranges. Allelochemicals such as terpenes, flavonoids, polyacetylene and phenols were also identified in the extracts, essential oil and the rhizosphere soil in S. altissima. Among the identified allelochemicals of S. altissima, the cis-dehydromatricaria ester may be involved in the allelopathy considering its growth inhibitory activity and its concentration in the rhizosphere soil. Therefore, the allelopathy of S. canadensis and S. altissima may support their invasiveness, naturalization and formation of thick monospecific stands. This is the first review article focusing on the allelopathy of both of S. canadensis and S. altissima.
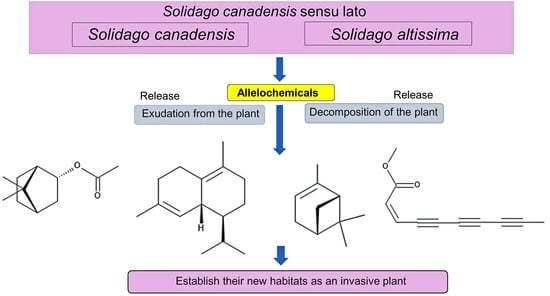
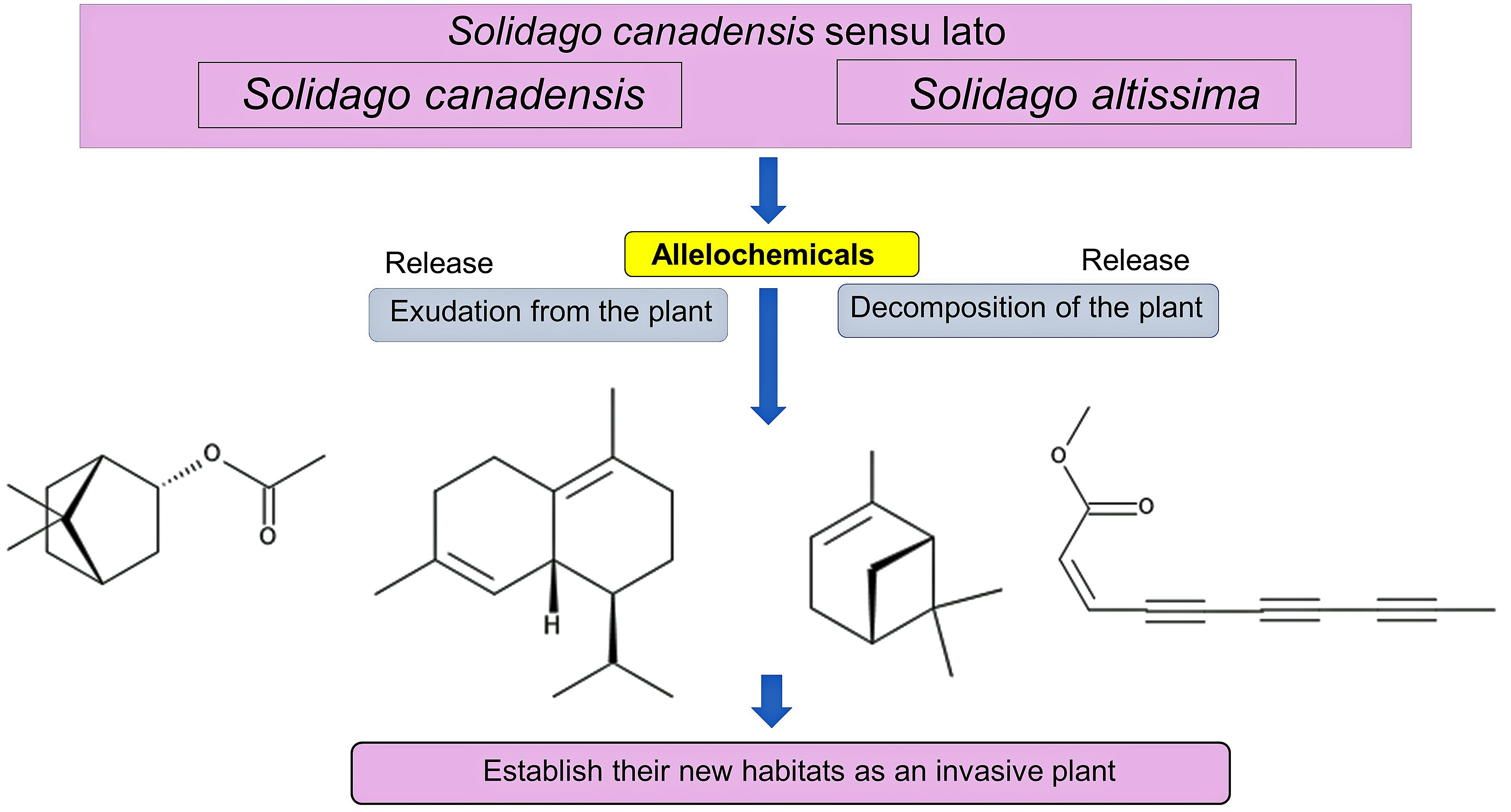
1. Introduction
Solidago canadensis sensu lato (s.l.), belonging Asteraceae, is an erect rhizomatous perennial plant, native to North America. S. canadensis s.l. was introduced to Europe as an ornamental plant in the 17th to 18th centuries. The species spread from the gardens to the natural environments, and has extended its habitats in Central and Eastern Europe. It expanded its habitat at a rate of 741 km2 per year in Europe [1]. The species has also been introduced and naturalized in many other countries such as Australia, Brazil, China, India, New Zealand and Japan [2,3,4,5,6].
The species expands its habitat though seed distribution and rhizome expansion. The rhizomes arise near the base of the shoots in autumn and produce aerial stems from their apex in the following spring. The stems are not branched, and bear triple-nerved, lanceolate, alternate leaves which are found along the stems and roots at the base of the shoots. The rhizome systems contribute to expanding the species’ community and to form thick monospecific stands [7,8]. Shoot density in the established stands of the species was reported to be 309 shoots per m2 [3]. In addition, oil-filled cavities, which contain terpenes and/or lipids, were randomly distributed in the rhizomes [9]. These compounds may have some biological functions such as allelopathy. The species is a prolific seed producer. Its inflorescence forms broad pyramidal panicles, which contain numerous florets (Figure 1). A single plant produces 1000–20,000 light-winged achenes which contain seeds. The achenes disperse easily by wind, water and human activities. The germination rate is 30–75%, depending on the conditions [7,8,10,11]. The seed distribution may contribute to establishing the populations of S. canadensis s.l. in new habitats.
This species has adapted to a wide range of soil fertility and water potential [8,11,12], as well as been colonized into disturbed areas such as abandoned fields, roadsides, riverbanks and forest edges [11,13,14]. The species established its population on an agriculture field in two years after the abandonment [15,16]. Once established, the population remained dominant over 30 years [8,17,18,19,20]. It was also reported that the species showed great impact on the native plant diversity in the introduced ranges [21,22,23]. Owing to the potential of the species for the rapid expansion and the formation of thick monospecific stands in the introduced ranges, as well as its impact on the environments, S. canadensis s.l. has been designated as a harmful invasive plant species [4,24].
The S. canadensis complex is a highly variable species. S. canadensis s.l. contains S. canadensis L. as S. canadensis subsp. canadensis (L.), and S. altissima L. as S. canadensis subsp. altissima (L.) O.Bolòs et Vigo [2,25,26]. S. canadensis and S. altissima are very similar taxa. The field experiments also showed that the competitive abilities of S. altissima and S. canadensis against other plant species was similar [27]. However, they could be distinguished by their morphological traits such as shoot length and flowering time [2,26,28,29]. The chromosome number between S. canadensis (diploid; 2n = 18) and S. altissima (hexaploid; 2n = 54) is also different [7,29,30]. The native ranges of both species in North America are not exactly the same [8,26]. In addition, the experimental crossing of S. canadensis and S. altissima could not bear viable seeds [30], which indicates a genetic barrier between both taxa. However, owing to the lack of consistency in the identification of the species, the separation of both species has been considered to be very problematic [2,28,29]. For example, most European populations of both species were described to be S. altissima [3,25], although macro-morphological analyses indicate that S. canadensis is a common species in Europa [29,31]. S. altissima was also mentioned as a synonym of S. canadensis [32,33]. There may have been a misidentification of the species.
It was reported that invasive plants are often allelopathic and inhibit the germination and growth of the native plant species in the invasive ranges through specific secondary metabolites defined as allelochemicals [34,35,36,37,38,39,40,41,42,43]. S. canadensis showed an allelopathic activity on the sugar maple seedlings in the field, greenhouse and laboratory conditions [44]. The cis-Dehydromatricaria ester was identified in the extracts of S. altissima as its allelochemical [45]. The evidence of the allelopathy of S. canadensis and S. altissima has been accumulated since the late 20th century, and their allelopathy was often implicated in the potential of their invasiveness and naturalization. However, there has been no review paper focusing on the allelopathy of both S. canadensis and S. altissima. This review provides an overview of the allelopathy and allelochemicals in S. canadensis and S. altissima, as well as a discussion on the involvement of allelopathy in the invasiveness and naturalization of the species. Despite the potential misidentification of S. canadensis and S. altissima described earlier, due to the fact that both species are very similar and the identification of the species is problematic [2,28,29], this paper followed the identification of the species in the publications because it is impossible to confirm their identification.
2. Allelopathy of S. canadensis
Allelopathy is the chemical interaction between donor plants and recipient plants through allelochemicals. Allelochemicals are produced in some plant parts and released into the vicinity of the donor plants, including their rhizosphere soil either by the root exudation, rainfall leachates, volatilization from the plant parts or decomposition processes of plant residues [46,47,48,49]. Several investigations in field conditions showed that S. canadensis reduced the number and biodiversity of the native plant community in its invaded ranges [50]. The invasion level of S. canadensis correlated negatively with the taxonomic diversity of the native plant community, and positively with the invasibility of the community [51]. Those observations may imply the involvement of allelopathy in the interaction between S. canadensis and native plant species to some extent. Many researchers have evaluated the allelopathic activity of the root exudates, rhizosphere soil, residues and plant extracts of S. canadensis (Table 1).
2.1. Allelopathy of Root Exudate and Plant Residue
Root exudates of S. canadensis, which were obtained from its aeroponic culture, significantly suppressed the growth of two Asian original plant species; Gnaphalium affine D.Don and Xanthium sibiricum Patrin ex Widder., two of America origin; Conyza canadensis (L.) Cronquist and Celosia argentea L., two of tropical origin; Aster subulatus Michx. and Sesbania cannabina (Renz.) Poir. and a cosmopolitan species; Eclipta prostrata (L.) L. The suppression rate was similar in all plant species [52]. Root exudates of S. canadensis also showed the growth inhibition of Arabidopsis thaliana (L.) Heynh. [53]. When the seeds of seven European native plant species were sown into the S. canadensis cultivated soils with or without activated carbon, the germination of five species such as Dactylis glomerata L., Lythrum salicaria L., Stachys officinalis (L.) Trevis. and Trifolium pratense L. were significantly suppressed in activated carbon-free plots than in activated carbon plots. Although the germination rate was not significantly different between both plots, the biomass of Arrhenatherum elatius (L.) P.Beauv. ex J. et C.Presl in the activated carbon plots after three months of sowing was two times greater than that in the activated carbon-free plots [53]. Activated carbon is a widely used material to investigate allelopathy because it adsorbs allelochemicals in the plant rhizosphere soil [34,54]. In addition, aqueous extracts of the rhizosphere soil of S. canadensis inhibited the germination and growth of Digitaria sanguinalis (L.) Scop. and Amaranthus retroflexus L., and the inhibitory activity was greater in the extracts of the soil obtained from the invasive ranges of S. canadensis (China) than that from its native ranges (USA) [55]. These observations suggest that certain allelochemicals, which may cause growth inhibition, would be released into the rhizosphere soil as root exudates of S. canadensis, and the released allelochemicals in the soil may be greater in the invasive ranges than those in the native ranges.
Crushed stems, leaves and rhizomes of S. canadensis were mixed with soil and water and kept at 20/15 °C (12/12 h light/dark condition), and the mixture was filtered after 45 days. The obtained filtrate suppressed the germination and growth of Raphanus sativus L. and Triticum aestivum L. [56]. This observation also suggests that certain allelochemicals may be released into the rhizosphere soil during the decomposition process of plant residues of S. canadensis.
2.2. Allelopathy of Plant Extract
Some plant tissues may contain allelochemicals, since allelochemicals are synthesized and stored in certain plant tissues until their release into the environment [46,47,48,49]. Many investigations on the allelopathic activity of the extracts from different plant parts of S. canadensis have been conducted. Aqueous extracts of the leaves of S. canadensis inhibited the germination and root growth of Raphanus sativus L. and Lactuca sativa L. [57], as well as those of Triticum aestivum L. and Setaria viridis (L.)P.Beauv. [58]. The extracts also suppressed the germination, growth and chlorophyll content of Trifolium pratense L. and Raphanus sativus L., and increased their electrolyte leakage from the cell membrane of the seedlings [59,60].
The fresh leaves and stems of S. canadensis were soaked in water for 48 h, and the obtained soaking water showed the inhibitory activity on the germination and growth of Raphanus sativus L. and Triticum aestivum L. [54]. Aqueous extracts of the above-ground parts of S. canadensis suppressed the germination and growth of Lactuca sativa L. [61], as well as those of Digitaria sanguinalis (L.) Scop. and Amaranthus retroflexus L. [55]. The inhibitory activity was greater in the plant extracts obtained from the heavily invaded stands than in those obtained from the lightly invaded stands [62], and in the plant extracts obtained from the invasive ranges than those from the native ranges [55].
Aqueous extracts of the above-ground parts and roots of S. canadensis inhibited the germination and growth of Zoysia japonica Steud, and the extracts of the above-ground parts significantly stimulated malondialdehyde and peroxidase activity [62]. The extracts of the stems, roots, blossoms and seeds of S. canadensis suppressed the germination and growth of Brassica napus L. and Lolium perenne L. [63], and the extracts of the roots and rhizomes of S. canadensis also inhibited the root growth of Raphanus sativus L. and Lactuca sativa L. [57].
Aqueous ethanol extracts of the roots and rhizomes of S. canadensis inhibited the germination and growth of Trifolium repens L., Trifolium pratense L., Medicago lupulina L., Suaeda glauca (Brunge) Brunge, Plantago virginica L., Kummerowia stipulacea (Maxium.) Makino, Festuca arundinacea Schreb., Ageratum conyzoides L., Portulaca oleracea L. and Amaranthus spinosus L. [64]. Aqueous ethanol extracts of the above- and below-ground parts of S. canadensis suppressed the germination of Kummerowia striata (Thunb.) Schindl., and the inhibitory activity was greater in the plant extracts collected from the invasive ranges of S. canadensis than those from its native ranges [65]. Aqueous and ethanol extracts of the leaves, stems and rhizomes of S. canadensis inhibited the germination and growth of Morus alba L., Pharbitis nil (L.) Roth, Triticum aestivum L. and Brassica campestris L., and the inhibition was grater in the ethanol extracts than in the aqueous extracts [66].
Investigations on the aqueous and ethanol extracts of every part of S. canadensis showed the allelopathic activity on the germination, growth, chlorophyll content, electrolyte leakage and/or some enzyme activities of several plant species, including the native plant species. The inhibitory activity was greater in the plant extracts obtained from the invasive ranges of S. canadensis than in those from its native ranges, and in the extracts collected from the heavily invaded stands than in those collected from the lightly invaded stands. These observations suggest that whole parts of S. canadensis may contain water and ethanol extractable allelochemicals, which may cause the inhibition. In addition, the plants grown in the invasive ranges and heavily invaded stands may contain more allelochemicals than the plants in the native ranges and lightly invaded stands.
2.3. Effects of the Extract on Arbuscular Mycorrhizal Fungi
The rhizomes of S. canadensis were soaked in water for 24 h, and the obtained soaking water caused the suppression of the arbuscular mycorrhizal colonization of Echinochloa crus-galli (L.) P.Beauv., Kummerowia striata (Thnb.) Schindl. and Ageratum conyzoides L. [67]. The field and greenhouse investigations also showed that S. canadensis altered the composition of the arbuscular mycorrhizal fungal population in its rhizosphere soil through the inhibition of some dominant species and the stimulation of other species. The established arbuscular mycorrhizal community increased the competitive ability and the biomass of S. canadensis [67,68,69,70,71]. This altered arbuscular mycorrhizal community also increased the mycorrhizal-mediated 15N uptake in S. canadensis, as well as decreased the 15N uptake in the native species Kummerowia statrica (Thunb.) Schindl. [72]. In addition, the aqueous ethanol extract of the roots and rhizomes of S. canadensis also suppressed the population of the soilborne pathogens, namely Pythium ultimum Trow and Rhizoctonia solani J.G. Kühn [73]. These observations indicate that the aqueous extracts of S. canadensis may alter the arbuscular mycorrhizal population and suppress the colonization of the native plant species. The established arbuscular mycorrhizal community enhanced the competitive ability of S. canadensis. Certain compounds in the extracts may be involved in the alteration of the arbuscular mycorrhizal community.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Allelopathic activities of exudates, rhizosphere soil, residues and plant extracts of S. canadensis.
Table 1.
Allelopathic activities of exudates, rhizosphere soil, residues and plant extracts of S. canadensis.
| Source | Inhibition | Target Plant Species | Reference | |||
|---|---|---|---|---|---|---|
| Germination | Growth | Chlorophyll | Mycorrhizal colonization | |||
| Root exudate | ✓ | Gnaphalium affine, Xanthium sibiricum, Conyza canadensis, Celosia argentea, Aster subulatus, Sesbania cannabina, Eclipta prostrata | [52] | |||
| ✓ | Arabidopsis thaliana, | [53] | ||||
| Rhizosphere soil | ✓ | ✓ | Dactylis glomerata, Lythrum salicaria, Stachys officinalis, Trifolium pratense | [53] | ||
| Soil extract | ✓ | ✓ | Digitaria sanguinalis, Amaranthus retroflexus | [55] | ||
| Residue | ✓ | ✓ | Raphanus sativus, Triticum aestivum | [56] | ||
| Plant extract | ||||||
| Whole part | ✓ | ✓ | Kummerowia striata | [65] | ||
| Leaf | ✓ | ✓ | Raphanus sativus, Lactuca sativa | [57] | ||
| ✓ | ✓ | Triticum aestivum, Setaria viridi | [58] | |||
| ✓ | Raphanus sativus | [59] | ||||
| ✓ | Trifolium pratense | [60] | ||||
| Leaf and stem | ✓ | ✓ | Raphanus sativus, Triticum aestivum | [56] | ||
| Above-ground part | ✓ | ✓ | Lactuca sativa | [61] | ||
| ✓ | ✓ | Digitaria sanguinalis (L.) Scop. and Amaranthus retroflexus | [55] | |||
| Above-ground part, root | ✓ | ✓ | Zoysia japonica | [62] | ||
| Stem, root, blossom, seed | ✓ | ✓ | Brassica napus, Lolium perenne | [63] | ||
| Leaf, stem, rhizome | ✓ | ✓ | Morus alba, Pharbitis nil, Triticum aestivum, Brassica campestris | [66] | ||
| Root, rhizome | ✓ | Raphanus sativus, Lactuca sativa | [57] | |||
| ✓ | ✓ | Trifolium repens, Trifolium pratense, Medicago lupulina, Suaeda glauca, Plantago virginica, Kummerowia stipulacea, Festuca arundinacea, Ageratum conyzoides, Portulaca oleracea, Amaranthus spinosus | [69] | |||
| Rhizome | ✓ | Echinochloa crus-galli, Kummerowia striata, Ageratum conyzoides | [67] |
2.4. Allelochemicals
As described above, the inhibitory activity of the extracts of the plants and rhizosphere soil of S. canadensis obtained from the invasive ranges was greater than that obtained from the native ranges [55,65]. The concentrations of total phenolics, total flavones and total saponins in S. canadensis and its rhizosphere soil obtained from the invasive ranges were also greater than those from the native ranges [55,65]. These concentrations in the soil obtained from S. canadensis-infested stands were also greater than those in the soil obtained from S. canadensis-free stands [74].
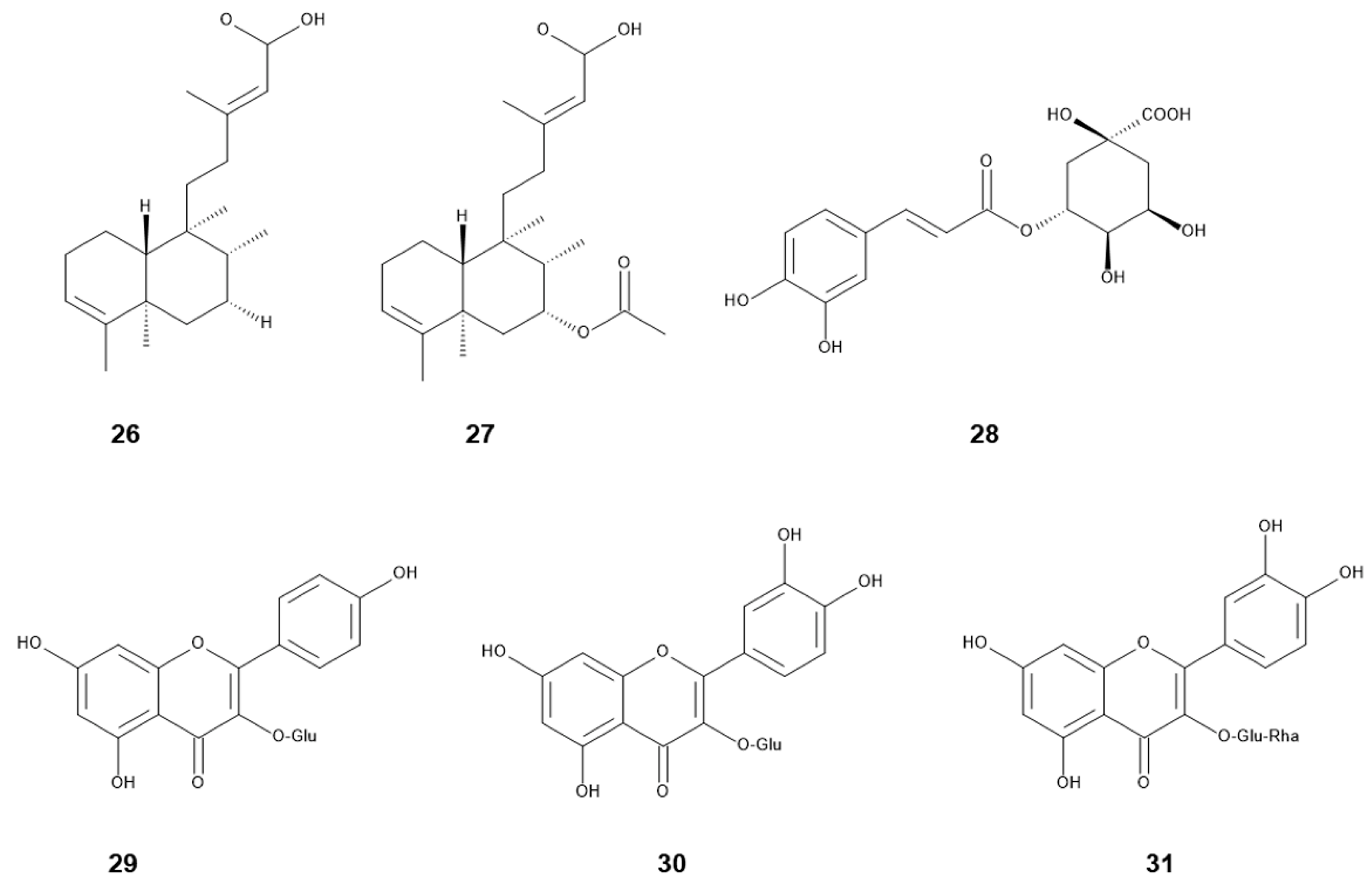
Major compounds identified in the aqueous methanol extracts of the leaves and inflorescences of S. canadensis were chlorogenic acid, quercitrin and rutin (quercetin-3-O-β-rutinoside) (Figure 2) [75]. A fatty acid, n-hexadecanonic acid, was isolated from the aqueous ethanol extract of the stems and leaves of S. canadensis as an allelopathic agent. n-Hexadecanonic acid significantly inhibited the growth of Triticum aestivum L. [56]. A flavonoid, kaempferol-3-O-d-glucoside, was isolated from the aqueous ethanol extract of the S. canadensis straw, and the compound inhibited the growth of Echinochloa colona (L.) Link [75]. In addition, the concentration of rutin in the leaves of S. canadensis was greater than that of other Solidago species [76,77]. Some flavonoids were also identified in the aerial parts of S. canadensis [78].
The essential oil of many plant species was reported to have several biological activities, and to comprise volatile compounds such as terpenoids [79]. These volatile compounds often exhibited several biological activities including allelopathic activity [80,81]. Therefore, the involvement of the essential oil of S. canadensis in allelopathy was also investigated. The production of the essential oil was greater in S. canadensis obtained from the heavily invaded stands than that from mildly invaded stands [82]. The main components of the essential oil in the leaves and inflorescences of S. canadensis were α-pinene, trans-verbenol, limonene, bornyl acetate and β-cubebene [83], and those in the aerial parts of S. canadensis were α-pinene, β-pinene, germacrene D, limonene, thymol, (+)epi-bicyclosesquiphellandrene, β-cadinene, γ-cadinene, δ-cadinene, α-muurolene, γ-muurolene, α-cubebene and β-elemene [84,85]. The essential oil in the inflorescences of S. canadensis showed antimicrobial activity, antioxidant activity and free-radical scavenging activity, and its main components were α-pinene, germacrene D and limonene [86]. α-Pinene and β-pinene were reported to disturb the cell division through the interference of DNA synthesis, and cause membrane peroxidation [87,88,89]. Several monoterpenes were reported to alter soil microflora [90]. It was also reported that the essential oil of the aerial parts of S. canadensis inhibited the germination and radical growth of Raphanus sativus L. and Lepidium sativum L. [84].
2.5. Contribution of Allelopathy of S. canadensis to Its Invasiveness
All parts of the extracts of S. canadensis, including its rhizosphere soils, the root exudates, residues and essential oil, inhibited the germination and growth of several plant species (Table 1). These observations suggest that S. canadensis produces and accumulates allelochemicals in its plant tissues, and that it releases those allelochemicals into the surrounding environments of S. canadensis, including its rhizosphere soils through the root exudation, decomposition process of plant residues and volatilization from its essential oil. Several allelochemicals such as fatty acids, terpenes, flavonoids and polyphenols (Table 2) were identified in the extracts and essential oil of S. canadensis. Some of these compounds were reported to inhibit the germination and growth of other plants species, including the native plant species.
The extracts of S. canadensis and its rhizosphere soil obtained from its invasive ranges had greater inhibitory activity compared to the extracts and rhizosphere soil obtained from its native ranges [55,65]. The total phenolics, flavones and saponins in S. canadensis in the invasive ranges were also greater than those in the native ranges [55,65,74]. These observations suggest that S. canadensis in the invasive ranges may enhance the competitive ability through the increased production of some of these compounds. The novel weapons hypothesis suggests that the competitive ability of the invasive plants is superior to that of the native plant species due to its allelochemicals. Those allelochemicals are new to the indigenous plant species, and those indigenous plants are susceptible to the compounds [34,91]. However, there has been no available information on the specific compounds of which the production is increased in S. canadensis in the invasive ranges, as well as the reasons for which those productions are increased in the invasive ranges.
S. canadensis showed the alteration of the composition of the arbuscular mycorrhizal population and suppressed the mycorrhizal colonization of the native plant species in the field and greenhouse experiments. The extracts of S. canadensis also suppressed the mycorrhizal colonization of the native plant species. These investigations suggest that certain compounds in the extracts may be involved in the suppression. The authors also indicated the involvement of allelochemicals in the suppression. Arbuscular mycorrhizal fungi are important mycorrhiza for the most territorial plants, and their colonization enhances the ability of the host plants to absorb mineral nutrients and water, as well as increase the defense functions against pathogen attacks and stress conditions [92,93,94]. The suppression of the colonization reduces the potential of the native plants for nutrient and water absorption, as well as the defense function, which may cause the reduction in growth and vigor of the native plant species.
All the literature described above indicates that S. canadensis may suppress the regeneration process of the native plant species directly by the suppression of their germination and growth, and indirectly by the degradation of the mycorrhizal fungal mutualism of the native plant species through its allelopathy. Therefore, the allelopathy of S. canadensis may contribute to its increasing competitive ability and make the plant invasive. However, it is necessary to identify the allelochemicals involved in the suppression of the fungal mutualism of the native plant species. Investigations into the specific inhibitory activity of the identified allelochemicals on the native plant species, as well as their concentrations in the rhizosphere soil and/or surrounding environments are necessary to evaluate the contribution of these allelochemicals to the invasiveness of S. canadensis.
3. Allelopathy of S. altissima
There has been limited information on the allelopathic activity of the root exudates, extracts and residues of S. altissima, unlike S. canadensis, as well as its allelopathy in the field conditions. The available information is the inhibitory effect of the n-hexane extracts of the underground parts of S. altissima on the growth of Lactuca sativa L. [95]. However, phytochemical investigations suggest that S. altissima contains polyacetylenes [96,97], monoterpene and sesquiterpenes [98], diterpenes [95,98,99,100,101,102], triterpenes [103,104], flavonoid glycosides [105,106] and polyphenols [107]. Some of them may act as allelopathic agents of the species.
3.1. Terpene
13E-7α-acetoxyl kolavenic acid (solidagonic acid) was isolated from the aqueous acetone extract of S. altissima [95,108], and the compound showed the growth inhibitory activity on the seedlings of Lactuca sativa L. and Lolium multiflorum Lam. [95,108]. 13E-kolavenic acid was also isolated from its n-hexane extracts [95]. Although 13E-kolavenic acid did not show any growth inhibitory activity, it exhibited antifungal activity and insect antifeedant activity [109,110]. These activities may help the species to protect itself from fungal pathogens and insect damage, as well as to establish its stands. It was reported that young rhizomes contained many more terpenes than old rhizomes, and α-pinene, β-pinene, limonene and germacrene D were identified in the rhizomes by GC-MS analysis [98]. α-Pinene, β-pinene, sabinene, myrcene, limonene, bornyl acetate and germacrene D were also found in the essential oil of S. altissima. However, the species was identified as a synonym of S. canadensis [33].
Table 2.
Allelochemicals identified in S. canadensis and S. canadensis and their sources.
| Chemical Class | Compound | Solidado canadensis | Reference | Solidado altissima | Reference | |||
|---|---|---|---|---|---|---|---|---|
| Extract | Essential oil | Extract | Soil | Essential oil | ||||
| Fatty acid | 1: n-Hexadecanonic acid | ✓ | [56] | |||||
| Polyacetylene | 2: cis-Dehydromatricaria ester | ✓ | ✓ | [45,96,98,104] | ||||
| 3: trans-Dehydromatricaria ester | ✓ | [45] | ||||||
| 4: (2Z,8Z)-10-Tigloyloxy matricaria ester | ✓ | [95] | ||||||
| 5: (2Z,8Z)-10-Angeloyloxy matricaria ester | ✓ | [95,96] | ||||||
| 6: Dehydromatricaria lactone | ✓ | [95,104,111] | ||||||
| 7: (4Z,8Z)-10-Trigloyloxy matricaria lactone | ✓ | [95,111] | ||||||
| 8: (4Z,8Z)-10-Angeloyloxy matricaria lactone | ✓ | [95] | ||||||
| Monoterpene | 9: α-Pinene | ✓ | [83,84,85,86] | ✓ | ✓ | [33,98] | ||
| 10: β-Pinene | ✓ | [84] | ✓ | ✓ | [33,98] | |||
| 11: trans-Verbenol | ✓ | [83] | ||||||
| 12: Limonene | ✓ | [83,84,85,86] | ✓ | ✓ | [33,98] | |||
| 13: Sabinene | ✓ | [33] | ||||||
| 14: Myrcene | ✓ | [33] | ||||||
| 15: Thymol | ✓ | [84] | ||||||
| 16: Bornyl acetate | ✓ | [83] | ✓ | [33] | ||||
| Sesquiterpene | 16: β-Elemene | ✓ | [84] | |||||
| 17: (+)epi-Bicyclosesquiphellandrene | ✓ | [84] | ||||||
| 18: β-Cadinene | ✓ | [84] | ||||||
| 19: γ-Cadinene | ✓ | [84] | ||||||
| 20: δ-Cadinene | ✓ | [84] | ||||||
| 21: α-muurolene | ✓ | [84] | ||||||
| 22: γ-muurolene | ✓ | [84] | ||||||
| 23: Germacrene D | ✓ | [84,86] | ✓ | ✓ | [33,98] | |||
| 24: α-cubebene | ✓ | [84] | ||||||
| 25: β-Cubebene | ✓ | [83,84] | ||||||
| Diterpene | 26: 13E-kolavenic acid | ✓ | [95] | |||||
| 27: 13E-7α-acetoxyl kolavenic acid | ✓ | [95,108] | ||||||
| Polyphenol | 28: Chlorogenic acid | ✓ | [76] | |||||
| Flavonoid | 29: Kaempferol-3-O-D-glucoside | ✓ | [75] | |||||
| 30: Quercitrin | ✓ | [76,77] | ||||||
| 31: Rutin | ✓ | [76] | ||||||
3.2. Polyacetylene
Several polyacetylenes, such as cis-dehydromatricaria ester (cis-DME), dehydromatricaria lactone, (2Z,8Z)-10-tigloyloxy matricaria ester, (4Z,8Z)-10-trigloyloxy matricaria lactone, (2Z,8Z)-10-angeloyloxy matricaria ester (methyl10-[(Z)-2-methyl-2-butenoyloxy]-(2Z,8Z)-2,8-decadiene-4,6-diynoate) and (4Z,8Z)-10-angeloyloxy matricaria lactone were isolated from the stems, roots and/or rhizomes of S. altissima [45,95,96,98,104,111]. Polyacetylenes in Asteraceae plants were reported to be synthesized from crepenynic acid, which is formed oleic acid with linoleic acid, introducing the first acetylene bond [95,112].
Among the polyacetylenes found in S. altissima, cis-DME, dehydromatricaria lactone and the (2Z,8Z)-10-angeloyloxy matricaria ester were reported to inhibit the coleoptile growth of Panicum crus-galli L. var. frumentaceum Trin. up to 77.4–93.5%, 81.7% and 80.0% that of the control, respectively, at the concentration of 1 ppm [96,111]. The (2Z,8Z)-10-Tigloyloxy matricaria ester also inhibited the growth of Lolium multiflorum Lam. [95]. However, cis-DME among those polyacetylenes was the most studied.
The bioactive concentration of cis-DME caused the growth inhibition, which was of 1–20 ppm in laboratory conditions [45,95,96,98,108,111,113]. cis-DME suppressed the germination of Asclepias syriaca L., Ambrosia artemisiifolia L. and Miscanthus sinensis Anderson; the germination and growth of Poa pratensis L. and Oryza sativa L.; and the growth of Lactuca sativa L. and Panicum crus-galli L. var. frumentaceum Trin. Their germination was inhibited up to 20–95% that of the control by 32–60 ppm cis-DME, and their growth was inhibited up to 10–56% that of the control by 48–100 ppm cis-DME (Table 3). Asclepias syriaca and Miscanthus sinensis, respectively, are competitive species with S. altissima in the native range of the USA and in the invasive range of Japan. Ambrosia artemisiifoli and Poa pratensis are the invasive plant species from North America, Europa and North Asia [45,98,114,115]. However, the effectiveness of cis-DME on those plant species was not evidently different.
The concentration of cis-DME was reported to be of 250–400 ppm in the roots of S. altissima, of 6.3 ppm in the soil under S. altissima [45] and of 0–17.3 ppm in the soil [116]. The concentrations in the soil varied depending on the soil properties and microbial activities [117,118,119]. The existence of cis-DME in the soil under S. altissima suggests that cis-DME may be released from S. altissima into the soils through the root exudation, rainfall leachates and/or the decomposition processes of plant residues. In addition, considering the bioactive concentration (1–20 ppm) described above and the soil concentration (0–17.3 ppm) of cis-DME, which is the same in laboratory conditions, the cis-DME in the soil may have some ecological function.
trans-DME was also in the soil under S. altissima, and its inhibitory activity was comparable of that of cis-DME [45]. When the ethanol solution (1%) of cis-DME was kept at 29 °C under 2000 lux for 2 days, 50% of cis-DME was isomerized into trans-DME, since the cis-form of the unsaturated ester is easily isomerized by light, pH or other conditions into the more stable trans-form [120]. In addition, trans-DME was not found in the roots and rhizomes of S. altissima [45]. Therefore, cis-DME may be isomerized by light and other environmental factors into trans-DME after the exudation from the plants into the soil. When we consider the involvement of DME in the allelopathy of S. altissima, the concentration of both cis-DME and trans-DME in the rhizosphere soil should be counted.
It was also reported that the aqueous extracts of the below-ground parts of S. altissima killed a pine wilt nematode (Bursaphelenchus lignicolous Mamiya and Kiyohara), and an active compound involved in it was determined as cis-DME. cis-DME at 10–11 ppm increased 50% of the molarity of the pine wilt nematode and a root-knot nematode (Meloidogyne incognita Kofild and White) [121]. The methanol extracts of the S. altissima rhizomes suppressed the hatching of fruit flies (Drosophila melanogaster Meigen), and an active compound was also determined as cis-DME [97]. The defense capacity of the invasive plants with natural enemies, such as pathogens, parasites and herbivores, contributes their establishment and naturalization into introduced ranges [37,122,123]. Therefore, the nematicidal and insecticidal activities of cis-DME may contribute to the establishment and naturalization of S. altissima in the introduced ranges.
3.3. Contribution of Allelopathy of S. altissima to Its Invasiveness
Unlike S. canadensis, the information on the allelopathic activity of S. altissima extracts, root exudates and residues is limited. However, polyacetylenes, monoterpene and sesquiterpenes, diterpenes, triterpenes and flavonoid glycosides were identified in its extracts and essential oil as allelopathic agents. Most potential compounds among them involved in the allelopathy may be cis-DME. cis-DME inhibited the germination and growth of several plant species at the concentration of 1–20 ppm [45,95,98,108,111,113]. Its concentration in the roots of S. altissima and in its rhizosphere soil was of 250–400 ppm and 0–17.3 ppm, respectively [45,116]. These observations suggest that cis-DME may be released into the rhizosphere soil by root exudation, rainfall leachates and/or decomposition processes of plant residues. trans-DME was also formed by the isomerization of cis-DME in the soil, and its inhibitory activity was the same as cis-DME. Although there has been no information of the concentration of trans-DME in the rhizosphere soil of S. altissima, the concentration of only cis-DME in several soils was over the concentration which was able to cause the growth inhibition [45,116]. In addition, cis-DME possesses nematicidal and insecticidal activities [97,113,121,124] Some flavonoids were also identified in the aerial parts of S. canadensis [78]. 13E-kolavenic acid also showed antifungal activity and insect antifeedant activity [109,110]. Therefore, these compounds including trans-DME may be involved in allelopathy and/or defense function against natural enemies, such as fungal pathogens, parasites and herbivores, and may contribute to their establishment and naturalization into the introduced ranges. However, it is necessary to determine the concentration of trans-DME in the rhizosphere soil of S. altissima, as well as to investigate the allelopathy of S. altissima in greenhouse and field conditions. It is also worth investigating the effect of S. altissima on the arbuscular mycorrhizal colonization of the native plant species due to the fact that S. altissima and S. canadensis are very close species.
4. Conclusions
S. canadensis and S. altissima are harmful invasive species naturalized in many countries, and form thick monospecific stands. Based on the literature, both species are allelopathic. The root exudate, residues, extracts, essential oil and rhizosphere soil of S. canadensis showed allelopathic activity on several plant species, including native plants, and suppressed the arbuscular mycorrhizal mutualism of the native plant species. Several allelochemicals were also identified in its extracts and essential oil. The concentrations of total phenolics, total flavones and total saponins in the rhizosphere soil of S. canadensis obtained from the invasive ranges were greater than those obtained from the native ranges. Therefore, the allelopathy of S. canadensis may contribute the interruption of the regeneration process of native plant species directly through the suppression of their germination and growth, and indirectly through the suppression of the mycorrhizal fungal mutualism of the native plant species. Several allelochemicals of S. altissima were also identified in its extracts and essential oil. Among them, cis-DME inhibited the germination and growth of several plant species at a concentration of 1–20 ppm, and its extractable concentration in the rhizosphere soil of S. altissima was of 0–17.3 ppm. cis-DME also exhibited nematicidal and insecticidal activities. trans-DME was formed by the isomerization of cis-DME in the soil, and its inhibitory activity was similar to that of cis-DME. Therefore, cis-DME, including trans-DME, may also cause the suppression of the regeneration process of the native plant species through the inhibition of their germination and growth. The allelopathy of S. canadensis and S. altissima may provide the species with competitive advantage against native plant species, and contribute to their invasiveness and naturalization.
Funding
This research received no external funding.
Institutional Review Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weber, E. The dynamics of plant invasions: A case study of three exotic goldenrod species (Solidago L.) in Europe. J. Biogeogr. 1998, 25, 147–154. [Google Scholar] [CrossRef]
- Weber, E. Morphological variation of the introduced perennial Solidago canadensis L. sensu lato (Asteraceae) in Europe. Bot. J. Linn. Soc. 1997, 123, 197–210. [Google Scholar] [CrossRef]
- Weber, E. Biological flora of Central Europe: Solidago altissima L. Flora 2000, 195, 123–134. [Google Scholar] [CrossRef]
- Invasive Species Compendium, Solidago canadensis. Available online: https://www.cabi.org/isc/datasheet/50599 (accessed on 12 September 2022).
- Royal Botanical Gardens, Kew, Solidago canadensis. Solidago altissima L. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:249454-1 (accessed on 12 September 2022).
- Royal Botanical Gardens, Kew, Solidago altissima L. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:285347-2 (accessed on 12 September 2022).
- Croat, T. Solidago canadensis complex of the great plains. Brittonia 1972, 24, 317–326. [Google Scholar] [CrossRef]
- Werner, P.A.; Bradbury, I.K.; Gross, R.S. The biology of Canadian weeds. 45. Solidago canadensis L. Can. J. Plant Sci. 1980, 60, 1393–1409. [Google Scholar] [CrossRef]
- Curtis, J.D.; Lersten, N.R. Oil reservorirs in stem, rhizomes, and root of Solidago canadensis (Asteraceas, tribe Astereae). Nor. J. Bot. 1990, 10, 443–449. [Google Scholar] [CrossRef]
- Meyer, A.H.; Schmid, B. Seed dynamics and seedling establishment in the invading perennial Solidago altissima under different experimental treatments. J. Ecol. 1999, 87, 28–41. [Google Scholar] [CrossRef]
- Cornelius, R. The strategies of Solidago canadensis L. in relation to urban habitats. I. Resource requirements. Acta Oecol. 1990, 11, 19–34. [Google Scholar]
- Huang, H.; Guo, S.; Chen, G. Reproductive biology in an invasive plant Solidago canadensis. Front. Biol. China 2007, 2, 196–204. [Google Scholar] [CrossRef]
- Follak, S.; Eberius, M.; Essl, F.; Fürdös, A.; Sedlacek, N.; Trognitz, F. Invasive alien plants along roadsides in Europe. EPPO Bull. 2018, 48, 256–265. [Google Scholar] [CrossRef]
- Park, J.S.; Choi, D.; Kim, Y. Potential Distribution of Goldenrod (Solidago altissima L.) during Climate Change in South Korea. Sustainability 2020, 12, 6710. [Google Scholar] [CrossRef]
- Kotowska, D.; Pärt, T.; Żmihorski, M. Evaluating google street view for tracking invasive alien plants along roads. Ecol. Indi. 2021, 121, 107020. [Google Scholar] [CrossRef]
- Shimoda, M.; Nakamoto, M. Vegetation and threatened plant dynamics of wet abandoned rice fields in Nakaikemi, Fukui, Prefecture, Japan. Jpn. J. Ecol. 2003, 53, 197–217. [Google Scholar]
- Shimoda, S.; Wagai, R. Ecosystem dynamics after abandonment of rice paddy fields; Does aaien plant invasion enhance carbon storage? Ecosystems 2020, 23, 617–629. [Google Scholar] [CrossRef]
- Newell, S.J.; Tramer, E.J. Reproductive strategies in herbaceous plant communities during succession. Ecology 1978, 59, 228–234. [Google Scholar] [CrossRef]
- Bakelaar, R.G.; Odum, E.P. Community and population level response to fertilization in an old-field ecosystem. Ecology 1978, 59, 660–665. [Google Scholar] [CrossRef]
- Maddox, G.D.; Cook, R.E.; Wimberger, P.H.; Gardescu, S. Clone structure in four Solidago altissima (Asteraceae) population: Rhizome connections within genotypes. Am. J. Bot. 1989, 76, 318–326. [Google Scholar] [CrossRef]
- Dong, L.J.; Yu, H.W.; He, W.M. What determines positive, neutral and negative impacts of Solidago canadensis invasion on native plant species richness? Sci. Rep. 2015, 5, 16804. [Google Scholar] [CrossRef] [Green Version]
- Fenesi, A.; Vágási, C.I.; Beldean, M.; Földesi, R.; Kolcsár, L.P.; Shapiro, J.T.; Török, E.; Kovács-Hostyánszki, A. Solidago canadensis impacts on native plant and pollinator communities in different-aged old fields. Basic Appl. Ecol. 2015, 16, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Zagurskaya, Y.V. Study issues of invasive species of the genus Solidago. Ecosyst. Transform. 2022, 5, 42–54. [Google Scholar] [CrossRef]
- EPPO. PQR database. Paris, France: European and Mediterranean Plant Protection Organization. Available online: https://gd.eppo.int/taxon/SOOCA (accessed on 12 September 2022).
- Weber, E.; Schmid, B. Latitudinal population differentiation in two species of Solidago (Asteraceae) introduced into Europe. Am. J. Bot. 1998, 85, 1110–1121. [Google Scholar] [CrossRef] [PubMed]
- iNatuarist, Identifying Solidago altissima & Solidago canadensis. Available online: https://www.inaturalist.org/posts/19288-identifying-solidago-altissima-solidago canadensis (accessed on 12 September 2022).
- Szymura, M.; Szymura, T.H. Interactions between alien goldenrods (Solidago and Euthamia species) and comparison with native species in Central Europe. Flora 2016, 218, 51–61. [Google Scholar] [CrossRef]
- Semple, J.C.; Rahman, H.; Bzovsky, S.; Sorour, M.K.; Kornobis, K.; Laphitz, R.L.; Tong, L. A multivariate morphometric study of the Solidago altissima complex and S. canadensis (Asteraceae: Astereae). Phytoneuron 2015, 10, 1–31. [Google Scholar]
- Verloove, F.; Zonneveld, B.J.M.; Semple, J.C.S. First evidence for the presence of invasive Solidago altissima (Asteraceae) in Europe. Willdenowia 2017, 47, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Melville, M.R.; Morton, J.K. A biosystematics study of the Solidago canadensis (Compositae) complex. I. The Ontario populations. Can. J. Bot. 1982, 60, 976–997. [Google Scholar] [CrossRef]
- Szymura, M.; Szymura, T.H.; Kreitschitz, A. Morphological and cytological diversity of goldenrods (Solidago L. and Euthamia Nutt.) from south-western Poland. Biodiv. Res. Conserv. 2015, 38, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Pisula, N.; Meiners, S.J. Allelopatyhic effcts of goldenrod species on turnover in successional communities. Am. Midl. Nat. 2010, 163, 161–172. [Google Scholar] [CrossRef]
- Lawson, S.K.; Sharp, L.G.; Powers, C.N.; McFeeters, R.L.; Satyal, P.; Setzer, W.N. Volatile Compositions and Antifungal Activities of Native American Medicinal Plants: Focus on the Asteraceae. Plants 2020, 9, 126. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 419–426. [Google Scholar] [CrossRef]
- Cappuccino, N.; Arnason, J.T. Novel chemistry of invasive exotic plants. Biol. Lett. 2006, 2, 189–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chengxu, W.; Mingxing, Z.; Xuhui, C.; Bo, Q. Review on allelopathy of exotic invasive plants. Procedia. Engin. 2011, 18, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Meiners, S.J.; Kong, C.H.; Ladwig, L.M.; Pisula, N.L.; Lang, K.A. Developing an ecological context for allelopathy. Plant. Ecol. 2012, 213, 1861–1867. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Involvement of allelopathy in the invasive potential of Tithonia diversifolia. Plants 2020, 9, 766. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy of Lantana camara as an invasive plant. Plants 2021, 10, 1028. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy of knotweeds as invasive plants. Plants 2022, 11, 3. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy and allelochemicals of Leucaena leucocephala as an invasive plant species. Plants 2022, 11, 1672. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy and Allelochemicals of Imperata cylindrica as an invasive plant species. Plants 2022, 11, 2551. [Google Scholar] [CrossRef]
- Fisher, R.F.; Woods, R.A.; Glavicic, M.R. Allelopathic effects of golderrod and aster on young sugar maple. Can. J. Forest Res. 1978, 8, 1–9. [Google Scholar] [CrossRef]
- Kobayashi, A.; Morimoto, S.; Shibata, Y.; Yamashita, K.; Numata, M. C10-Polyacetylenes as allelopathic substances in dominants in early stages of secondary succession. J. Chem. Ecol. 1980, 6, 119–131. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; pp. 1–422. [Google Scholar]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest. Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef] [PubMed]
- Bielecka, A.; Borkowska, L.; Królak, E. Environmental changes caused by the clonal invasive plant Solidago canadensis. Ann. Bot. Fennici. 2019, 57, 33–48. [Google Scholar] [CrossRef]
- Wang, C.; Cheng, H.; Wanga, S.; Weia, M.; Du, D. Plant community and the influence of plant taxonomic diversity on community stability and invasibility: A case study based on Solidago canadensis L. Sci. Total Environ. 2021, 768, 144518. [Google Scholar] [CrossRef]
- Yang, B.; Li, J. Phytotoxicity of toot exudates of invasive Solidago canadensis on co-occurring native and invasive plant species. Pak. J. Bot. 2022, 54, 1019–1024. [Google Scholar] [CrossRef]
- Abhilasha, D.; Quintana, N.; Vivanco, J.; Joshi, J. Do allelopathic compounds in invasive Solidago canadensis s.l. restrain the native European flora? J. Ecol. 2008, 96, 993–1001. [Google Scholar] [CrossRef]
- Weißhuhn, K.; Prati, D. ; Activated carbon may have undesired side effects for testing allelopathy in invasive plants. Basic Appl. Ecol. 2009, 10, 500–507. [Google Scholar] [CrossRef]
- Yang, X.; Cheng, J.; Yao, B.; Lu, H.; Zhang, Y.; Xu, J.; Song, X.; Qiang, S. Polyploidy-promoted phenolic metabolism confers the increased competitive ability of Solidago canadensis. Oikos 2021, 130, 1014–1025. [Google Scholar] [CrossRef]
- Zhang, D.; Ye, Y.; Li, L.; Domg, L. Allelopathic pathways, isolation and identification of an allelopathic substance from Solidago canadensis L. Allelopath. J. 2014, 33, 201–212. [Google Scholar]
- Butcko, V.M.; Jensen, R.J. Evidence of tissue-specific allelopathic activity in Euthamia graminifolia and Solidago canadensis (Asteraceae). Am. Midl. Nat. 2002, 148, 253–262. [Google Scholar] [CrossRef]
- Li, S.L.; Li, Z.H.; Wang, Y.F.; Xiao, R.; Pan, C.D.; Wang, Q. Preliminary study for the allelopathic effect of water extracts from Solidago canadensis leaves. Adv. Mater. Res. 2013, 699, 340–348. [Google Scholar] [CrossRef]
- Możdżeń, K.; Barabasz-Krasny, B.; Zandi, P.; Kliszcz, A.; Puła, J. Effect of aqueous extracts from Solidago canadensis L. Leaves on germination and early growth stages of three cultivars of Raphanus Sativus L. var. Radicula Pers. Plants 2020, 9, 1549. [Google Scholar] [CrossRef] [PubMed]
- Zandi, P.; Barabasz-Krasny, B.; Stachurska-Swakoń, A.; Puła, J.; Możdżeń, K. Allelopathic effect of invasive Canadian goldenrod (Solidago canadensis L.) on early growth of red clover (Trifolium pratense L.). Not. Bot. Horti. Agrobo. 2020, 48, 2060–2071. [Google Scholar] [CrossRef]
- Yu, Y.; Cheng, H.; Xu, Z.; Zhong, S.; Wang, C.; Guo, E. Invasion intensity modulates the allelopathic impact of Solidago canadensis L. leaves and roots against Lactuca sativa L. during germination and early seedling stage. Int. J. Environ. Res. 2022, 16, 48. [Google Scholar] [CrossRef]
- Sun, J.F.; Liang, Q.J.; Wu, N.; Javed, Q.; Huang, P.; Du, D.L. Allelopathic effects of aqueous extracts from different plant parts of Canada goldenrod (Solidago canadensis L.) on seed germination and seedling growth of Korean lawngrass (Zoysia japonica Steud.). Appl. Ecol. Environ. Res. 2022, 20, 1009–1022. [Google Scholar] [CrossRef]
- Baležentien, L. Secondary metabolite accumulation and phytotoxicity of invasive species Solidago canadensis L. during the growth period. Allelopath. J. 2015, 35, 217–226. [Google Scholar]
- Yang, R.Y.; Mei, L.X.; Tang, J.J.; Chen, X. Allelopathic effects of invasive Solidago canadensis L. on germination and growth of native Chinese plant species. Allelopath. J. 2007, 19, 241–248. [Google Scholar]
- Yuan, Y.; Wang, B.; Zhang, S.; Tang, J.; Tu, C.; Hu, S.; Yong, J.W.H.; Chen, X. Enhanced allelopathy and competitive ability of invasive plant Solidago canadensis in its introduced range. J. Plant Ecol. 2013, 6, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Sun, B.Y.; Tan, J.Z.; Wan, Z.G.; Gu, F.G.; Zhu, M.D. Allelopathic effects of extracts from Solidago canadensis L. against seed germination and seedling growth of some plants. J. Environ. Sci. 2006, 18, 304–309. [Google Scholar]
- Zhang, Q.; Yao, L.J.; Yang, R.Y.; Yang, X.Y.; Tang, J.J.; Chin, X. Potential allelopathic effects of an invasive species Solidago canadensis on the mycorrhizae of native plant species. Allelopath. J. 2007, 20, 71–78. [Google Scholar]
- Sun, Z.K.; He, W.M. Evidence for enhanced mutualism hypothesis: Solidago canadensis plants from regular soils perform better. PLoS ONE 2010, 5, e15418. [Google Scholar] [CrossRef]
- Betekhtina, A.A.; Mukhacheva, T.A.; Kovaleva, S.Y.; Gusevb, A.P.; Veselkin, D.V. Abundance and diversity of arbuscular mycorrhizal fungi in invasive Solidago canadensis and indigenous S. virgaurea. Russ. J. Ecol. 2016, 47, 575–579. [Google Scholar] [CrossRef]
- Dong, L.J.; Ma, L.N.; He, W.M. Arbuscular mycorrhizal fungi help explain invasion success of Solidago canadensis. Appl. Soil. Ecol. 2021, 167, 103763. [Google Scholar] [CrossRef]
- Řezáčová, V.; Řezáč, M.; Gryndler, M.; Hršelová, H.; Gryndlerová, H.; Michalová, T. Plant invasion alters community structure and decreases diversity of arbuscular mycorrhizal fungal communities. Appl. Soil. Ecol. 2021, 167, 104039. [Google Scholar] [CrossRef]
- Yang, R.; Zhou, G.; Zan, S.; Guo, F.; Su, N.; Li, J. Arbuscular mycorrhizal fungi facilitate the invasion of Solidago canadensis L. in southeastern China. Acta Oecol. 2014, 61, 74–77. [Google Scholar] [CrossRef]
- Zhang, S.; Jin, Y.; Tang, J.; Chen, X. The invasive plant Solidago canadensis L. Suppresses local soil pathogens through allelopathy. Appl. Soil. Ecol. 2009, 41, 215–222. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, W.; Wang, B.; Tang, J.; Chen, X. Secondary metabolites from the invasive Solidago canadensis L. Accumulation in soil and contribution to inhibition of soil pathogen Pythium ultimum. Appl. Soil. Ecol. 2011, 48, 280–286. [Google Scholar] [CrossRef]
- Jun, L.; Yonghao, Y.; Hongwu, H.; Liyao, D. Kaempferol-3-O-β-D-glucoside, a potential allelochemical isolated from Solidago canadensis. Allelopath. J. 2011, 28, 259–266. [Google Scholar]
- Radusiene, J.; Marska, M.; Ivanauskas, L.; Jakstas, J.; Karpaviciene, B. Assessment of phenolic compound accumulation in two widespread goldenrods. Ind. Crops Prod. 2015, 63, 158–166. [Google Scholar] [CrossRef]
- Likhanov, A.; Oliinyk, M.; Pashkevych, N.; Churilov, A.; Kozyr, M. The role of flavonoids in invasion strategy of Solidago canadensis L. Plants 2021, 10, 1748. [Google Scholar] [CrossRef]
- Shelepova, O.; Vinogradova, Y.; Vergun, O.; Grygorieva, O.; Brindza, J. Assessment of flavonoids and phenolic compound accumulation in invasive Solidago сanadensis L. in Slovakia. Slovak J. Food Sci. 2020, 14, 587–594. [Google Scholar]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological activities of essential oils: From plant chemoecology to traditional healing systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef] [PubMed]
- Jassbi, A.R.; Zare, S.; Firuzi, O.; Xiao, J. Bioactive phytochemicals from shoots and roots of Salvia species. Phytochem Rev. 2016, 15, 829–867. [Google Scholar] [CrossRef]
- Macías, F.A.; Mejías, F.J.R.; Molinillo, J.M.G. Recent advances in allelopathy for weed control: From knowledge to applications. Pest Manag. Sci. 2019, 75, 2413–2436. [Google Scholar] [CrossRef] [PubMed]
- Baranová, B.; Troščáková-Kerpčárová, E.; Grul’ová, D. Survey of the Solidago canadensis L. morphological traits and essential oil production: Aboveground biomass growth and abundance of the invasive goldenrod appears to be reciprocally enhanced within the invaded dtands. Plants 2022, 11, 535. [Google Scholar] [CrossRef]
- Radušienė, J.; Karpavičienė, B.; Marksa, M.; Ivanauskas, L.; Raudonė, L. Distribution patterns of essential oil terpenes in native and invasive Solidago species and their comparative assessment. Plants 2022, 11, 1159. [Google Scholar] [CrossRef]
- Grul’ová, D.; Baranová, B.; Ivanova, V.; De Martino, L.; Mancini, E.; De Feo, V. Composition and bio activity of essential oils of Solidago spp. and their Impact on radish and garden cress. Allelopath. J. 2016, 39, 129–141. [Google Scholar]
- Shelepova, O.; Vinogradova, Y.; Zaitchik, B.; Ruzhitsky, A.; Grygorieva, O.; Brindza, J. Constituents of the essential oil in Solidago canadensis L. from Eurasia. Slovak J. Food Sci. 2018, 12, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Marinas, I.C.; Oprea, E.; Buleandra, M.; Bleots, C.; Badea, I.R.; Anastasiu, P.; Lazar, V.; Gardus, I.D.; Chifiric, M.C. Chemical, antimicrobial, antioxidant and anti-proliferative features of the essential oil extracted from the invasive plant Solidago canadensis L. Rev. Chim. 2020, 71, 255–264. [Google Scholar] [CrossRef]
- Nishida, N.; Tamotsu, S.; Nagata, N.; Saito, C.; Sakai, A. Allelopathic effects of volatile monoterpenoids produced by Salvia leucophylla: Inhibition of cell proliferation and DNA synthesis in the root apical meristem of Brassica campestris seedlings. J. Chem. Ecol. 2005, 31, 1187–1203. [Google Scholar] [CrossRef]
- Chowhan, N.; Singh, H.P.; Batish, D.R.; Kohli, R.H. Phytotoxic effects of β-pinene on early growth and associated biochemical changes in rice. Acta Physiol. Plant. 2011, 33, 2369–2376. [Google Scholar] [CrossRef]
- Chowhan, N.; Singh, H.P.; Batish, D.R.; Kaur, S.; Ahuja, N.; Kohli, R.K. β-Pinene inhibited germination and early growth involves membrane peroxidation. Protoplasma 2013, 250, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Yun, K.W.; Kil, B.S.; Han, D.M. Phytotoxic and antimicrobial activity of volatile constituents of Artemisia princeps var. orientalis. J. Chem. Ecol. 1993, 19, 2757–2766. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: From molecules and genes to species interaction. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; pp. 1–815. [Google Scholar]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Tang, H.; Hassan, M.U.; Feng, L.; Nawaz, M.; Shah, A.N.; Qari, S.H.; Liu, Y.; Miao, J. The critical role of arbuscular mycorrhizal fungi to improve drought tolerance and nitrogen use efficiency in crops. Front. Plant Sci. 2022, 13, 919166. [Google Scholar] [CrossRef] [PubMed]
- Nishidono, Y.; Tanaka, K. Comprehensive characterization of polyacetylenes and diterpenes from the underground parts of Solidago altissima L. and their contribution to the overall allelopathic activity. Phytochemistry 2022, 193, 112986. [Google Scholar] [CrossRef]
- Ichihara, K.; Kawai, T.; Kaji, M.; Noda, M. A mew polyacetylene from Solidago altissima L. Agric. Biol. Chem. 1976, 40, 353–358. [Google Scholar] [CrossRef]
- Kawazu, K.; Ariwa, M.; Kii, Y. An ovicidal substance, cis-dehydromatricaria ester from Solidago altissima. Agric. Biol. Chem. 1977, 41, 223–224. [Google Scholar]
- Johnson, R.H.; Halitschke, R.; Kessler, A. Simultaneous analysis of tissue- and genotype-specific variation in Solidago altissima (Asteraceae) rhizome terpenoids, and the polyacetylene dehydromatricaria ester. Chemoecology 2010, 20, 255–264. [Google Scholar] [CrossRef]
- Nishino, C.; Manabe, S.; Kazui, M.; Matsuzaki, T. Piscicidal cis-clerodane diterpenes from Solidago altissima. L.: Absolute configurations of 5α, 10α-cis-clerodanes. Tetrahedron Lett. 1984, 25, 2809–2812. [Google Scholar] [CrossRef]
- Bohlmann, F.; Singh, P.; Singh, R.K.; Joshi, K.C.; Jakupovic, J. A diterpene with a new carbon skeleton from Solidago altissima. Phytochemistry 1985, 24, 1114–1115. [Google Scholar] [CrossRef]
- Tori, M.; Katto, A.; Sono, M. Nine new clerodane diterpenoids from rhizomes of Solidago altissima. Phytochemistry 1999, 52, 487–493. [Google Scholar] [CrossRef]
- Nishidono, Y.; Tanaka, K. New clerodane diterpenoids from Solidago altissima and stereochemical elucidation via 13C NMR chemical shift analysis. Tetrahedron 2022, 110, 132691. [Google Scholar] [CrossRef]
- Okano, A.; Nomura, Y.; Tezuka, T. Identification of bauerenol in Solidago altissima. J. Nat. Prod. 1983, 46, 750–751. [Google Scholar] [CrossRef]
- Sawabe, A.; Minemoto, K.; Minematsu, T.; Ouchi, S.; Okamoto, T.; Morita, M.; Ouchi, S.; Okamoto, T. Characterization of the Z and E isomers of dehydromatricaria lactones. J. Jpn. Oil Chem. Soc. 1997, 46, 413–417. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Tanaka, T.; Kouno, I.; Ishimaru, K. A new kaempferol trioside from Solidago altissima L. J. Nat. Med. 2007, 61, 351–354. [Google Scholar] [CrossRef]
- Wu, B.; Takahashi, T.; Kashiwagi, T.; Tebayashi, S.; Kim, C.S. New flavonoid glycosides from the leaves of Solidago altissima. Chem. Pharm. Bull. 2007, 55, 815–816. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Ogino, K.; Fujioka, T.; Yoshida, M.; Ishimaru, K. A new acylphloroglucinol glycoside from Solidago altissima L. J. Nat. Med. 2008, 62, 199–201. [Google Scholar] [CrossRef]
- Sawabe, A.; Minemoto, K.; Ouchi, S.; Okamoto, T. Effects of acetylenes and terpenoids from Solidago altissima L. on seed germination. J. Jpn. Oil Chem. Soc. 1999, 48, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Salah, M.A.; Bedir, E.; Toyang, N.J.; Khan, I.A.; Harries, M.D.; Wedge, D.E. Antifungal clerodane diterpenes from Macaranga monandra (L) Muell. et Arg. (Euphorbiaceae). J. Agric. Food Chem. 2003, 51, 7607–7610. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, M. Chemical defense against insects in Heterotheca subaxillaris and three Orobanchaceae species using exudates from trichomes. Pest Manag. Sci. 2019, 75, 2474–2481. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, K.; Kawai, T.; Noda, M. Polyacetylenes of Solidago altissima L. Agric. Biol. Chem. 1978, 42, 427–431. [Google Scholar] [CrossRef]
- Minto, R.E.; Blacklock, B.J. Biosynthesis and function of polyacetylenes and allied natural products. Prog. Lipid Res. 2008, 47, 233–306. [Google Scholar] [PubMed] [Green Version]
- Uesugi, A.; Kessler, K. Herbivore exclusion drives the evolution of plant competitiveness via increased allelopathy. New Phytol. 2013, 198, 916–924. [Google Scholar] [CrossRef]
- Carson, W.; Root, R. Herbivory and plant species coexistence: Community regulation by an outbreaking phytophagous insect. Ecol. Monogr. 2000, 70, 73–99. [Google Scholar] [CrossRef]
- Yamada, T. Miscanthus. In Industrial Crops. Handbook of Plant Breeding; Cruz, V.M.V., Dierig, D.A., Eds.; Springer: New York, NY, USA, 2015; Volume 9, pp. 43–66. [Google Scholar]
- Uesugi, A.; Johnson, R.; Kessler, K. Context-dependent induction of allelopathy in plants under competition. Oikos 2019, 128, 1492–1502. [Google Scholar] [CrossRef]
- Ito, I.; Kobayashi, K.; Yoneyama, T. Fate of dehydromatricaria ester added to soil and its implications for the allelopathic effect of Solidago altissima L. Ann. Bot. 1998, 82, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Kaur, R.; Kaur, S.; Baldwin, I.T.; Inderjit. Taking ecological function seriously: Soil microbial communities can obviate allelopathic effects of released metabolites. PLoS ONE 2004, 4, e4700. [Google Scholar] [CrossRef]
- Kobayashi, K.; Koyama, H.; Shim, I. Relationship between behaviour of dehydromatricaria ester in soil and the allelopathic activity of Solidago altissima L. in the laboratory. Plant Soil 2004, 259, 97–102. [Google Scholar] [CrossRef]
- Tsao, R.; Eto, M. Light-activated plant growth inhibitory activity of cis-dehydromatricaria ester, rose bengal and fluoren-9-one on lettuce (Lactuca sativa L.). Chemosphere 1996, 32, 1307–1317. [Google Scholar] [CrossRef]
- Sakai, H.; Yoneda, K. Possible dual roles of an allelopathic compound, cis-dehydromatricaria ester. J. Chem. Ecol. 1981, 8, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Mack, R.M. Predicting the identity and fate of plant invaders: Emergent and emerging approaches. Biol. Conserv. 1996, 78, 107–121. [Google Scholar] [CrossRef]
- Keane, R.M.; Crawley, M.J. Exotic plant invsions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–169. [Google Scholar] [CrossRef]
- Ismail, M.; Kowsar, A.; Javed, S.; Choudhary, M.I.; Khan, S.W.; Abbas, Q.; Tang, Y.; Wang, W. The antibacterial, insecticidal and nematocidal activities and toxicity studies of Tanacetum falconeri Hook.f. Turk. J. Pharm. Sci. 2021, 18, 744–751. [Google Scholar]
Figure 1.
Solidago canadensis s.l.

Figure 2.
Allelochemicals identified in S. canadensis and S. altissima. Compound number and name were listed in Table 2.
Figure 2.
Allelochemicals identified in S. canadensis and S. altissima. Compound number and name were listed in Table 2.


Table 3.
Growth inhibitory activity of cis-dehydromatricaria ester.
| cis-DME Concentration (ppm) | Target Plant | Germination | Growth | Reference |
|---|---|---|---|---|
| (% of Control) | ||||
| 32 | Asclepias syriaca | 5 | [98] | |
| 48 | Poa pratensis | 20 | 50 | [113] |
| 50 | Oryza sativa | 88 | 10–15 | [45] |
| 50 | Ambrosia artemisiifolia | 25 | [45] | |
| 50 | Miscanthus sinensis | 22 | [45] | |
| 100 | Lactuca sativa | 30–56 | [108] | |
| 100 | Panicum crus-galli | 18 | [111] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kato-Noguchi, H.; Kato, M. Allelopathy and Allelochemicals of Solidago canadensis L. and S. altissima L. for Their Naturalization. Plants 2022, 11, 3235. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11233235
AMA Style
Kato-Noguchi H, Kato M. Allelopathy and Allelochemicals of Solidago canadensis L. and S. altissima L. for Their Naturalization. Plants. 2022; 11(23):3235. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11233235
Chicago/Turabian StyleKato-Noguchi, Hisashi, and Midori Kato. 2022. "Allelopathy and Allelochemicals of Solidago canadensis L. and S. altissima L. for Their Naturalization" Plants 11, no. 23: 3235. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11233235
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.