
Impact of Different Fertilizer Sources under Supplemental Irrigation and Rainfed Conditions on Eco-Physiological Responses and Yield Characteristics of Dragon’s Head (Lallemantia iberica)
 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Results
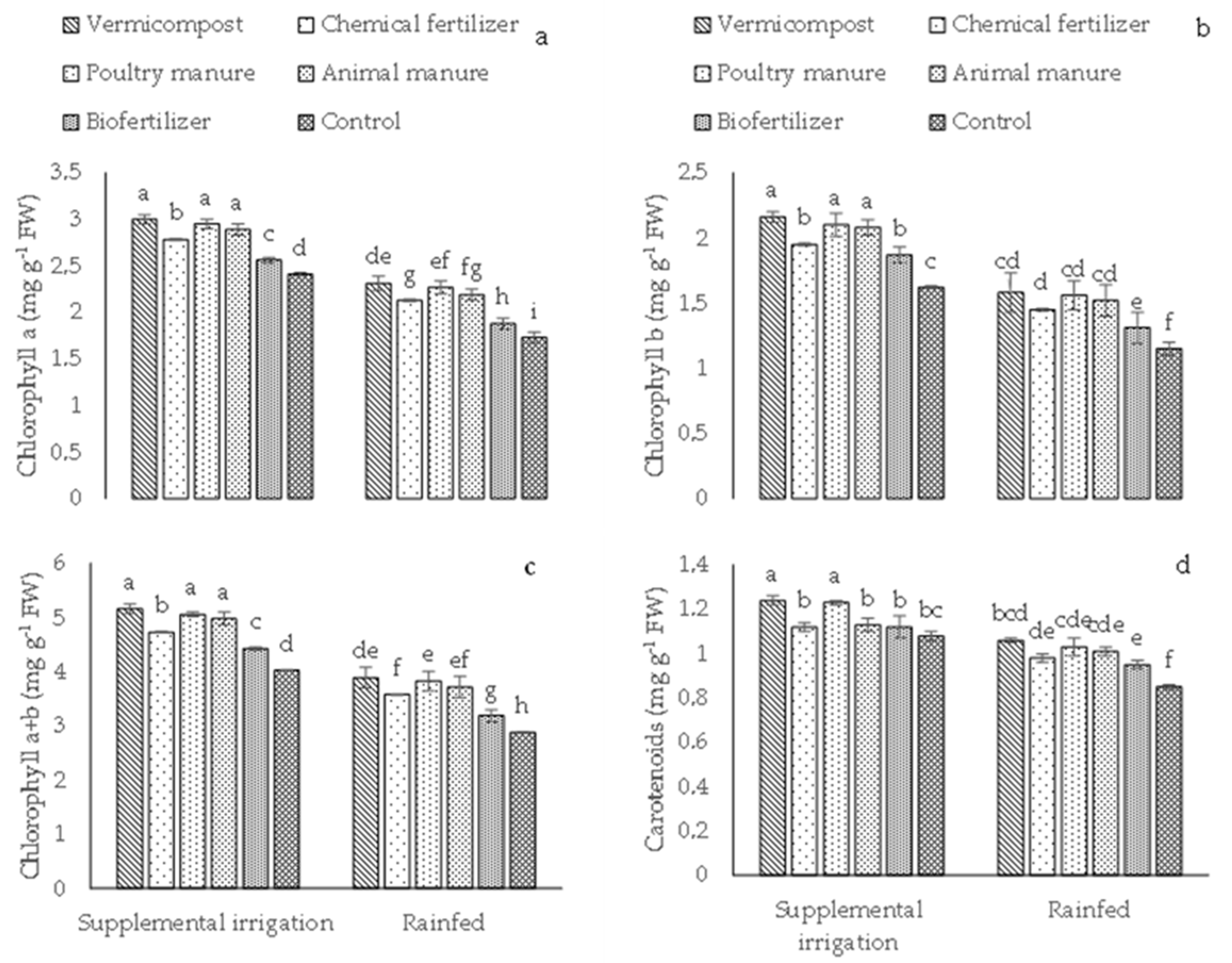
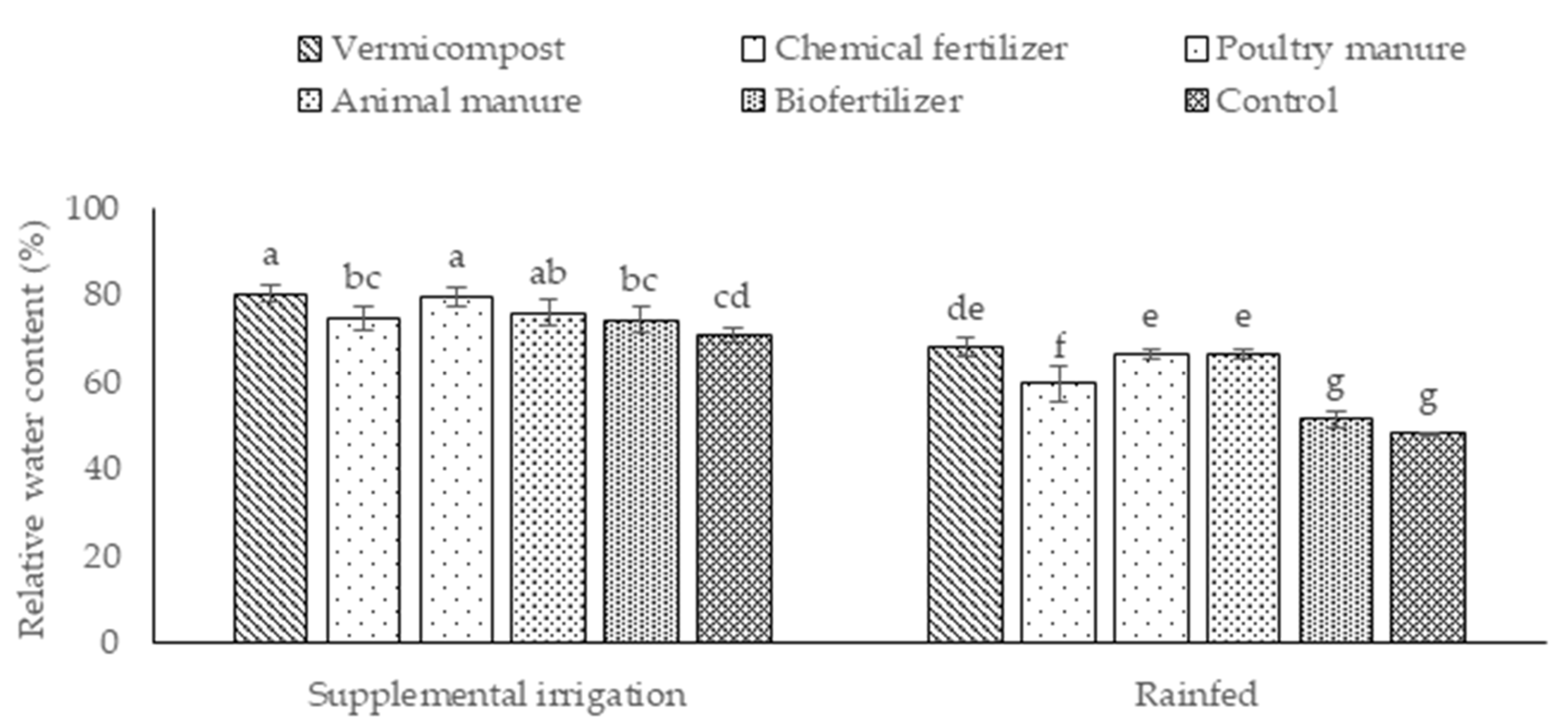
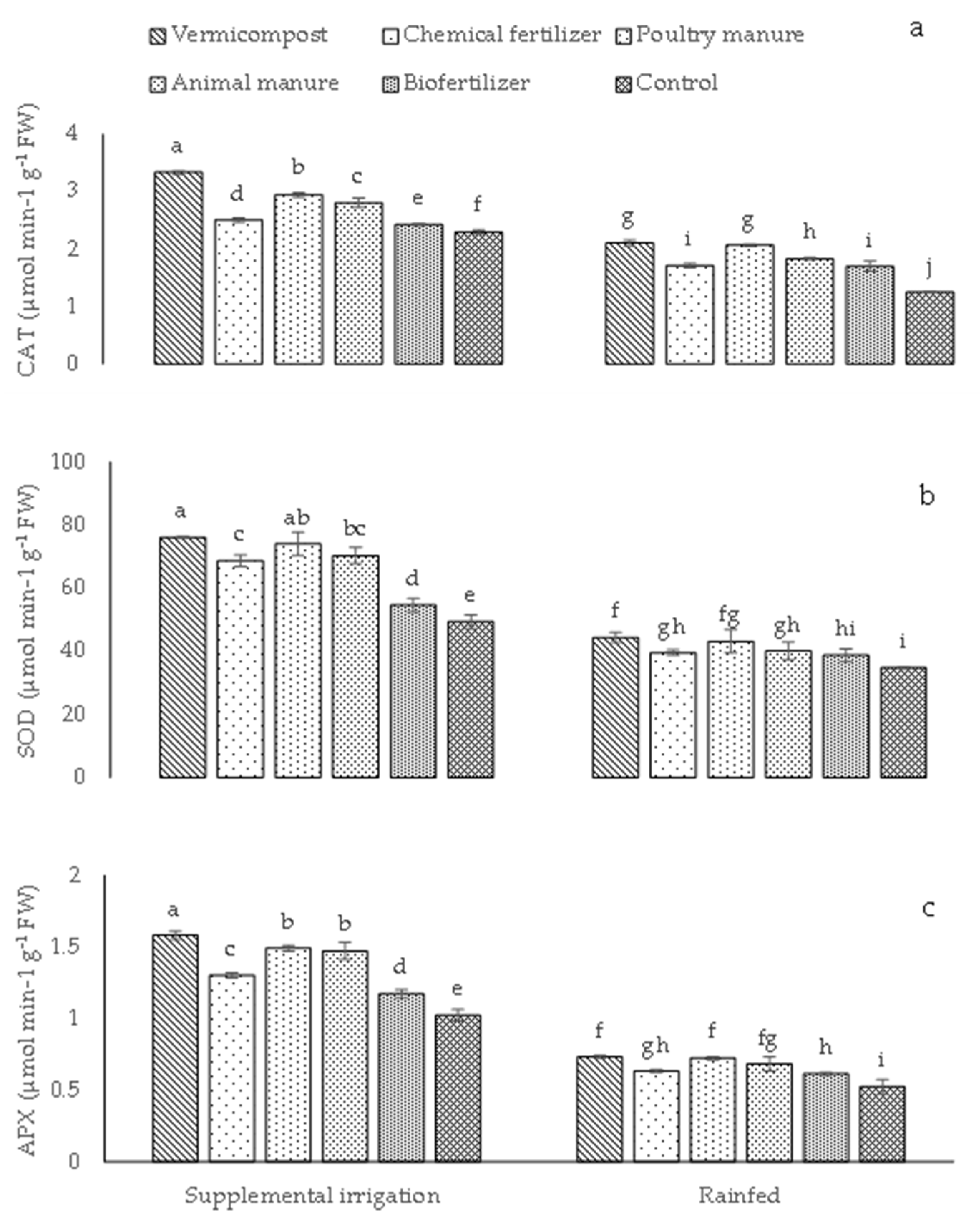
2.1. Photosynthetic Pigments, Relative Water Content (RWC), and Antioxidant Enzyme Activity
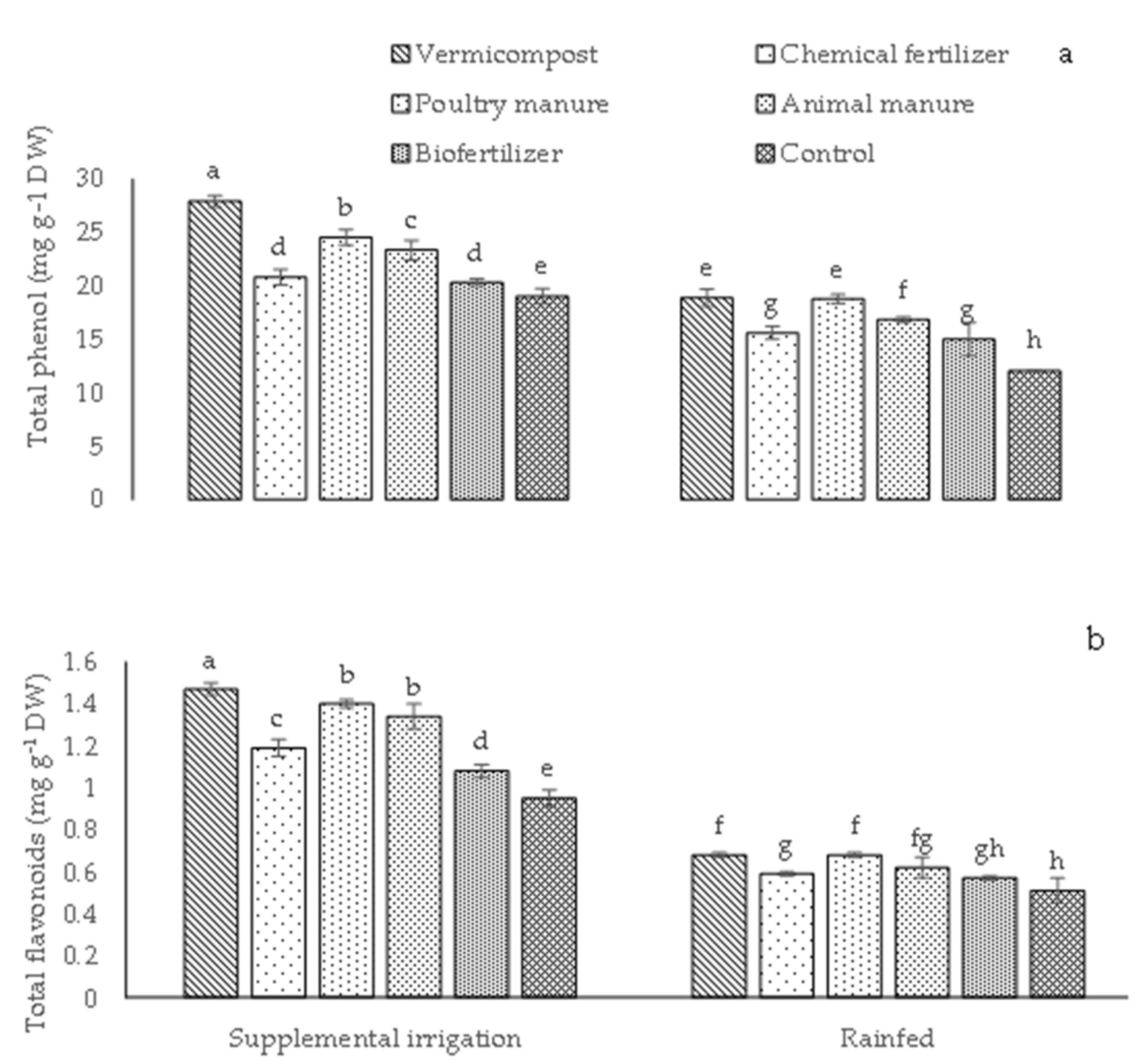
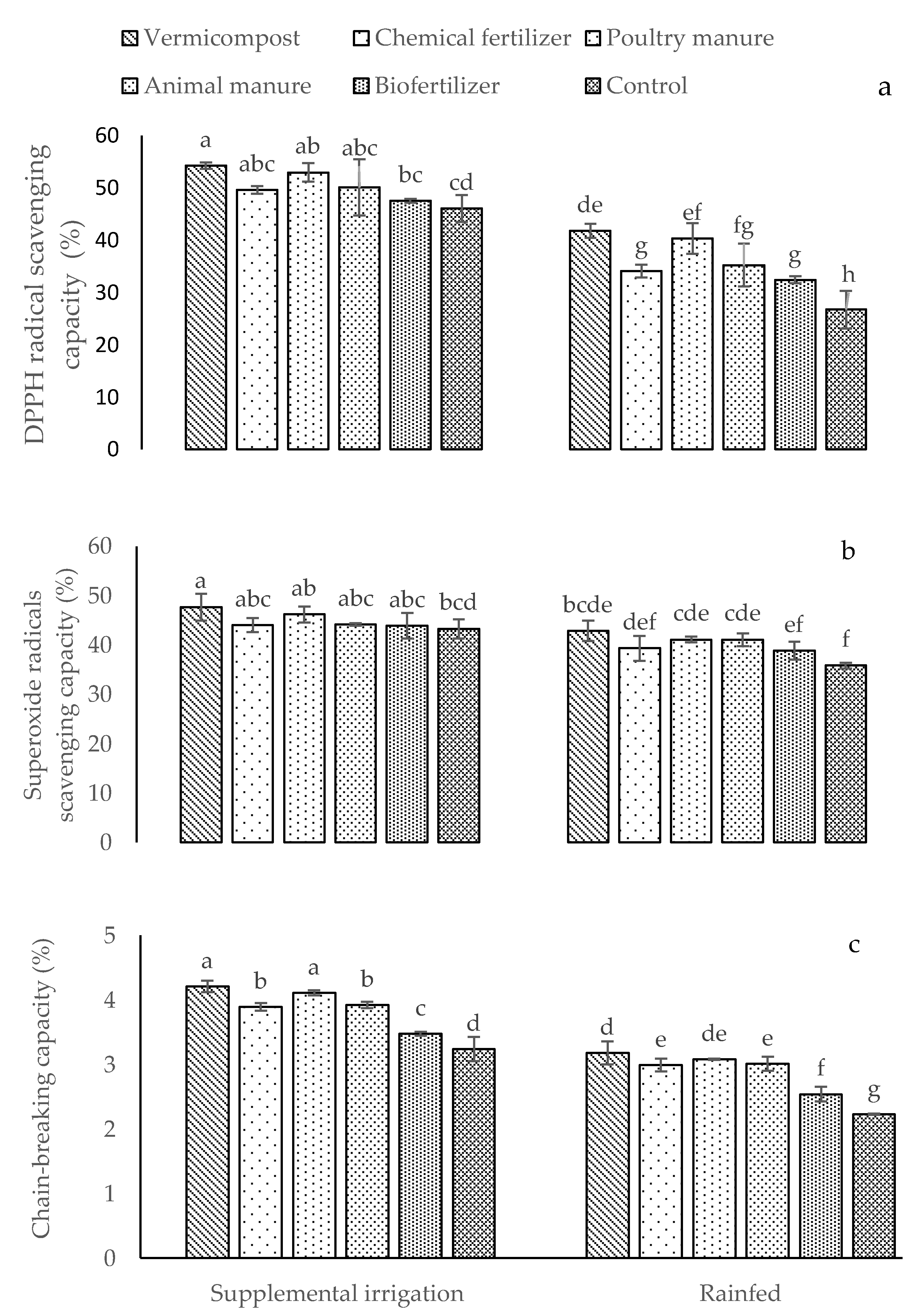
2.2. Total Flavonoids and Total Phenol Content, DPPH Radical Scavenging, Superoxide Radical Scavenging, and Chain-Breaking Capacity
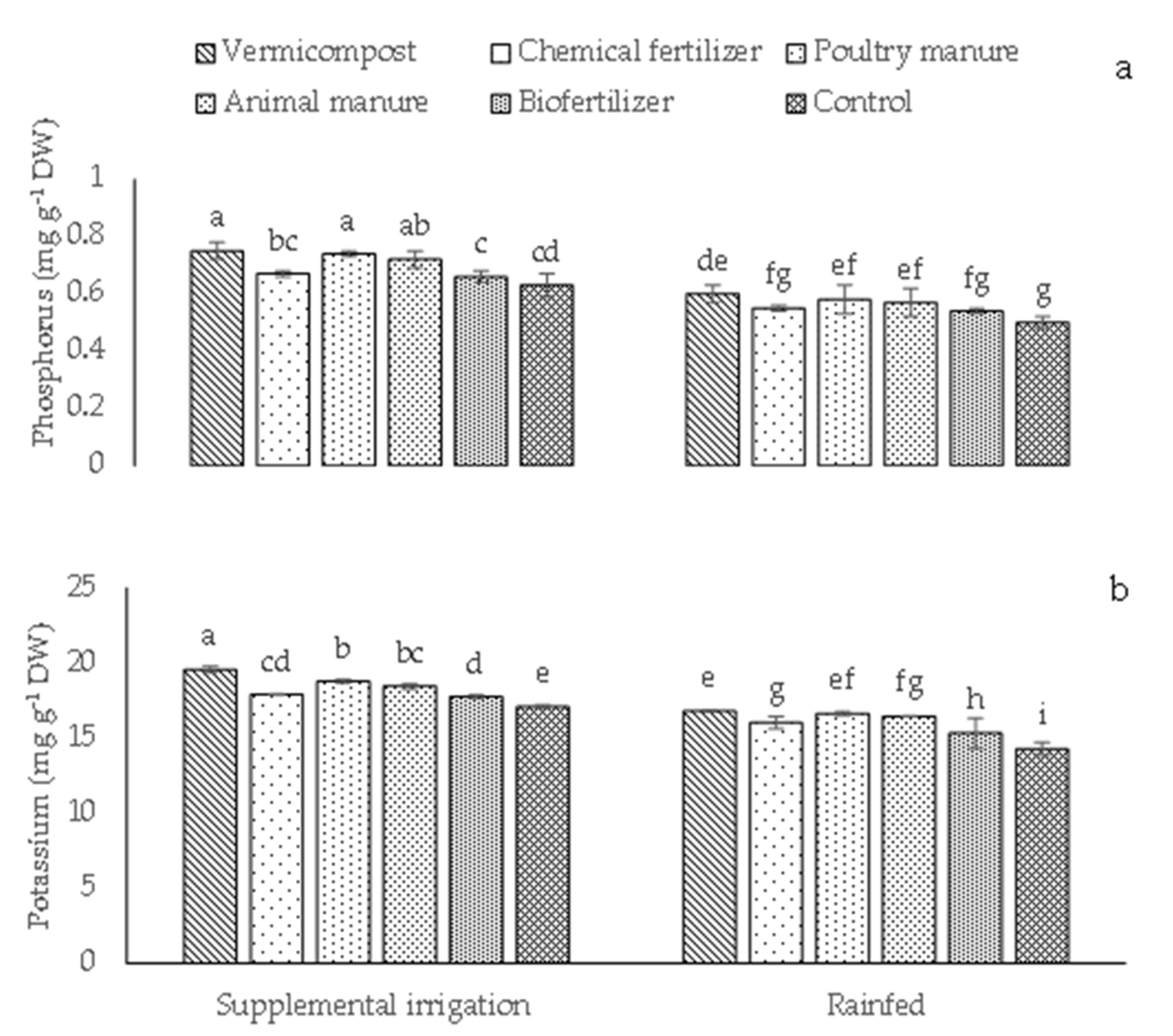
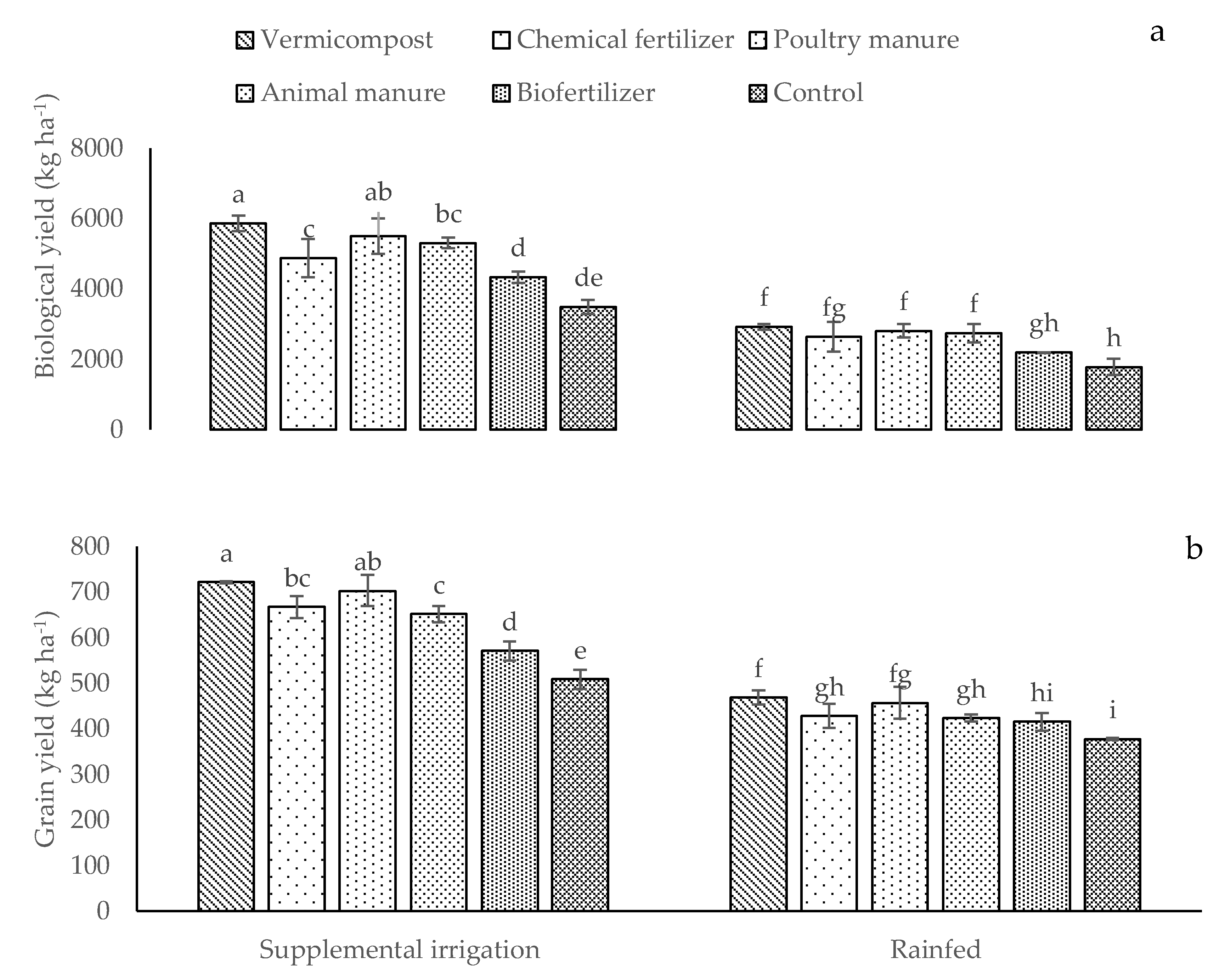
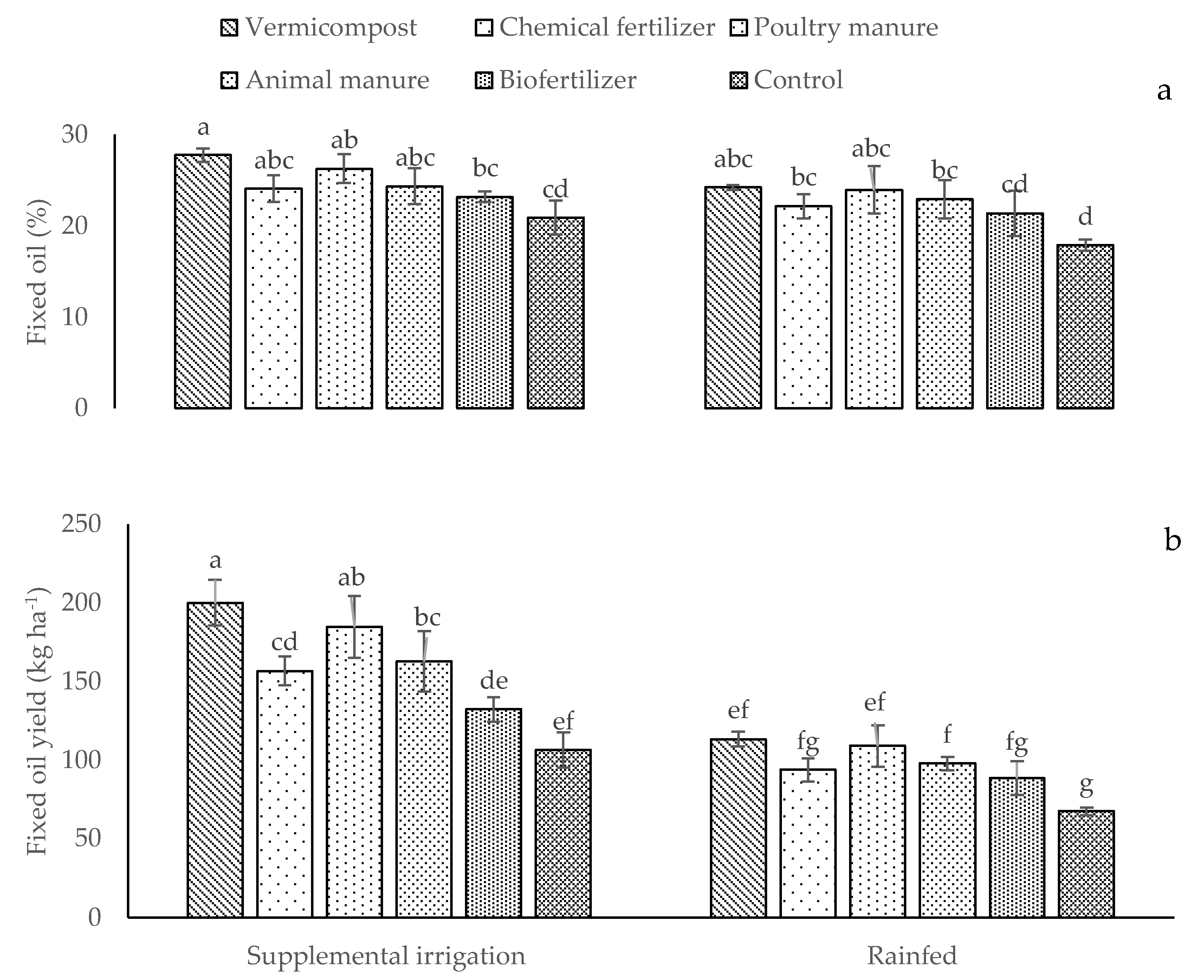
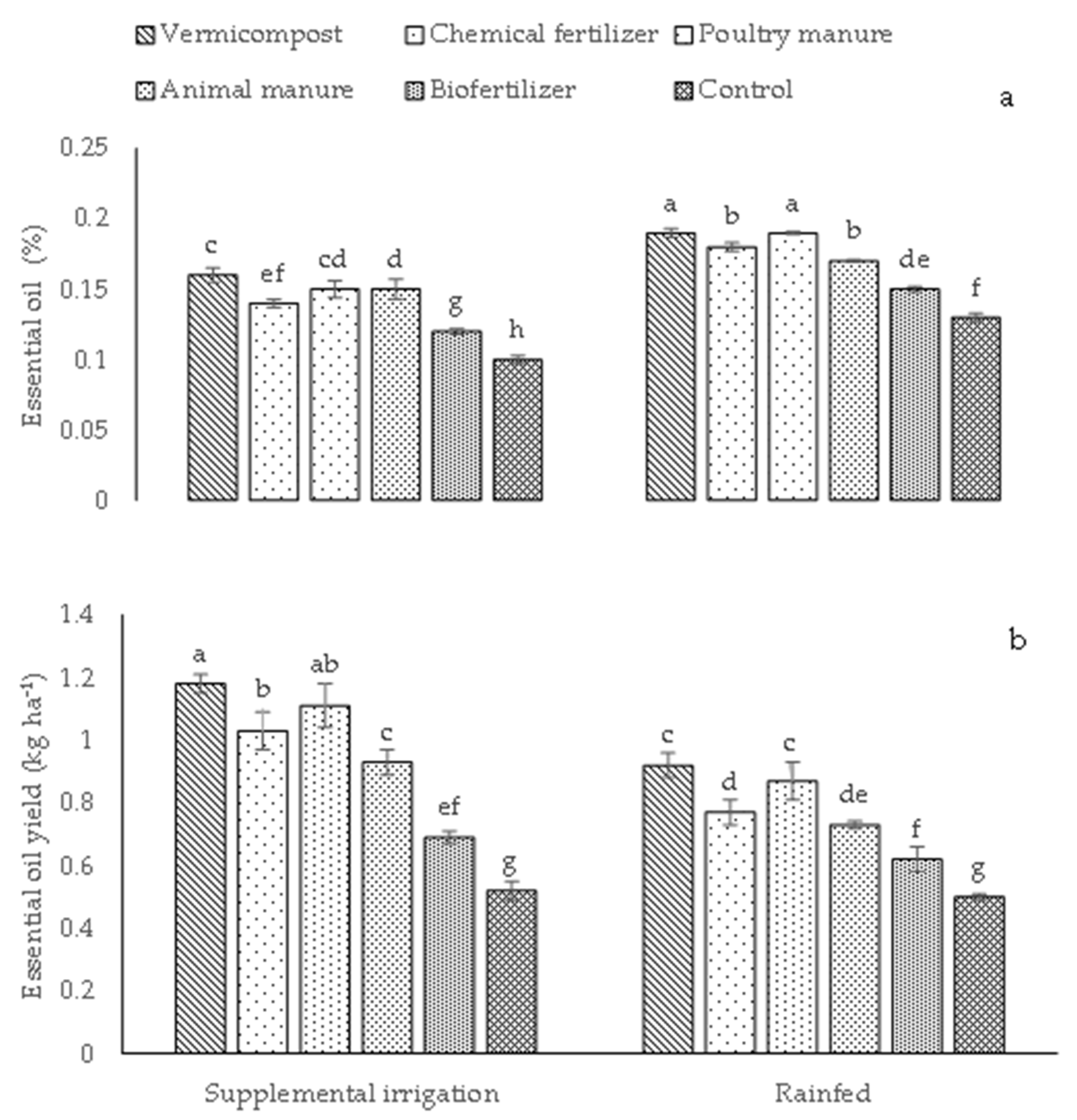
2.3. Content of Elements, Biological and Grain Yields, Fixed Oil Percentage and Fixed Oil Percentage Yield, and Essential Oil and Essential Oil Yield
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Plant Material
4.3. Measurements
4.3.1. Photosynthetic Pigment Content
4.3.2. Relative Water Content
4.3.3. Antioxidant Enzyme Extraction and Assays
Catalase (CAT)
Superoxide Dismutase (SOD)
Ascorbate Peroxidase (APX)
4.3.4. Antioxidant Compounds
Total Phenolic Content (TPC)
Total Flavonoid Content (TFC)
DPPH (2,2-diphenyl-1-picrylhydrazyl-hydrate) Radical Scavenging Activity
Superoxide Radical Scavenging Activity
Chain-Breaking Capacity
4.3.5. Nutrients of Potassium and Phosphorus
4.3.6. Seed and Biological Yield Characteristics
4.3.7. Content of Fixed Oil and Essential Oil
4.3.8. Data Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mirzapour, M.; Valizadeh, N.; Heydarzadeh, S.; Rahimi, A.; Gitari, H. Factors affecting the production of secondary metabolites in medicinal plants. In New Development on Medicinal and Aromatic Plants-II; Ozyazici, G., Ed.; Iksad Publishing House: Ankara, Turkey, 2022; pp. 3–32. [Google Scholar]
- Mohammadghasemi, V.; Moghaddam, S.S.; Rahimi, A.; Pourakbar, L.; Popović-Djordjević, J. The effect of winter sowing, chemical, and nano-fertilizer sources on oil content and fatty acids of dragon’s head (Lallemantia iberica Fischer & CA Meyrefeer. J. Plant Growth Regul. 2020, 40, 1714–1727. [Google Scholar]
- Mohammadghasemi, V.; Siavash Moghaddam, S.; Rahimi, A.; Pourakbar, L.; Popović-Djordjević, J. Winter cultivation and nano fertilizers improve yield components and antioxidant traits of dragon’s head (Lallemantia iberica (MB) Fischer & Meyer). Plants 2020, 9, 252. [Google Scholar]
- Nouri, M.; Homaeem, M.; Bannayan, M. Climate variability impacts on rainfed cereal yields in west and northwest Iran. Int. J. Biometeorol. 2017, 61, 1571–1583. [Google Scholar] [CrossRef] [PubMed]
- Heydarzadeh, S.; Jalilian, J.; Pirzad, A.; Jamei, R.; Petrussa, E. Fodder value and physiological aspects of rainfed smooth vetch affected by biofertilizers and supplementary irrigation in an agri-silviculture system. Agroforest Syst. 2022, 96, 221–232. [Google Scholar] [CrossRef]
- Heydari, S.; Pirzad, A. Efficiency of Funneliformis mosseae and Thiobacillus sp. on the secondary metabolites (essential oil, seed oil and mucilage) of Lallemantia iberica under salinity stress. J. Hortic. Sci. Biotechnol. 2020, 96, 249–259. [Google Scholar] [CrossRef]
- Rahimzadeh, S.; Pirzad, A. Arbuscular mycorrhizal fungi and Pseudomonas in reduce drought stress damage in flax (Linum usitatissimum L.): A field study. Mycorrhiza 2017, 27, 537–552. [Google Scholar] [CrossRef]
- Saadat, B.; Pirzad, A.; Jalilian, J. Yield-related biochemical response of understory mycorrhizal yellow sweet clover (Melilotus officinalis L.) to drought in agrisilviculture. Arch. Agron. Soil Sci. 2021, 67, 1603–1620. [Google Scholar] [CrossRef]
- Zheng, X.; Yu, Z.; Zhang, Y.; Shi, Y. Nitrogen supply modulates nitrogen remobilization and nitrogen use of wheat under sup-plemental irrigation in the North China Plain. Sci. Rep. 2020, 10, 3305. [Google Scholar] [CrossRef]
- Amirnia, R.; Ghiyasi, M.; Moghaddam, S.S.; Rahimi, A.; Damalas, C.A.; Heydarzadeh, S. (Nitrogen-fixing soil bacteria plus mycorrhizal fungi improve seed yield and quality traits of lentil (Lens culinaris Medik). J. Soil Sci. Plant Nutr. 2019, 19, 592–602. [Google Scholar] [CrossRef]
- Mehrabi, Y.; Movahhedi Dehnavi, M.; Salehi, A.; Mohatashami, R.; Hamidian, M. Improving physiological traits, yield and essential oil of cumin (Cuminum cyminum L.) via application of phosphorus bio-fertilizers and supplementary irrigation under dryland condition. Iran. J. Medic. Arom. Plants Res. 2020, 35, 1044–1057. [Google Scholar]
- Maddahi, S.; Rahimi, A.; Siavash Moghaddam, S.; Pourakbar, L.; Popović-Djordjević, J. Effects of sowing time and chemical, organic, and biological fertilizer sources on yield components and antioxidant properties of dragon’s head (Lallemantia iberica (M. Bieb.) Fisch. & CA Mey). J. Plant Growth Regul. 2021, 41, 1276–1290. [Google Scholar]
- Kahil, A.A.; Hassan, F.A.S.; Ali, E.F. Influence of bio-fertilizers on growth, yield and anthocyanin content of Hibiscus sabdariffa L. plant under Taif region conditions. Annu. Res. Biol. 2017, 17, 1–15. [Google Scholar] [CrossRef]
- Hassan, F.A.S.; Ali, E.F.; Mahfouz, S.A. Comparison between different fertilization sources, irrigation frequency and their combinations on the growth and yield of coriander plant. Aust. J. Basic. Appl. Sci. 2012, 6, 600–615. [Google Scholar]
- Elsayed, S.I.; Glala, A.A.; Abdalla, A.M.; Abd El Ghafour, A.; Darwish, M.A. Effect of biofertilizer and organic fertilization on growth, nutrient contents and fresh yield of dill (Anethum graveolens). Bull. Nat. Res. Cent. 2020, 44, 122. [Google Scholar] [CrossRef]
- Eliaspour, S.; Seyed Sharifi, R.; Shirkhani, A. Evaluation of interaction between Piriformospora indica, animal manure and NPK fertilizer on quantitative and qualitative yield and absorption of elements in sunflower. Food Sci. Nutr. 2020, 8, 2789–2797. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, A.; Siavash Moghaddam, S.; Ghiyasi, M.; Heydarzadeh, S.; Ghazizadeh, K.; Popović-Djordjević, J. The Influence of chemical, organic and biological fertilizers on agrobiological and antioxidant properties of Syrian cephalaria (Cephalaria Syriaca L.). Agriculture 2019, 9, 122. [Google Scholar] [CrossRef]
- Hosseinzadeh, M.H.; Ghalavand, A.; Boojar, M.M.A.; Modarres-Sanavy, S.A.M.; Mokhtassi-Bidgoli, A. Application of manure and biofertilizer to improve soil properties and increase grain yield, essential oil and ω3 of purslane (Portulaca oleracea L.) under drought stress. Soil Tillage Res. 2021, 205, 104633. [Google Scholar] [CrossRef]
- Ahmad, S.; Ghaffar, A.; Rahman, M.H.U.; Hussain, I.; Iqbal, R.; Haider, G.; Khan, M.A.; Ikram, R.M.; Hussnain, H.; Bashir, M.S. Effect of application of biochar, poultry and farmyard manures in combination with synthetic fertilizers on soil fertility and cotton productivity under arid environment. Commun. Soil Sci. Plant Anal. 2021, 52, 2018–2031. [Google Scholar] [CrossRef]
- Singh, A.; Karmegam, N.; Singh, G.S.; Bhadauria, T.; Chang, S.W.; Awasthi, M.K.; Sudhakar, S.; Arunachalam, K.D.; Biruntha, M.; Ravindran, B. Earthworms and vermicompost: An eco-friendly approach for repaying nature’s debt. Environ. Geochem. Health 2020, 42, 1617–1642. [Google Scholar] [CrossRef]
- Darakeh, S.A.S.S.; Weisany, W.; Diyanat, M.; Ebrahimi, R. Bio-organic fertilizers induce biochemical changes and affect seed oil fatty acids composition in black cumin (Nigella sativa Linn). Ind. Crops Prod. 2021, 164, 113383. [Google Scholar] [CrossRef]
- Ahmadian, A.; Tavassoli, A.; Amiri, E. The interaction effect of water stress and manure on yield components, essential oil and chemical compositions of cumin (Cuminum cyminum). Afr. J. Agric. Res. 2011, 6, 2309–2315. [Google Scholar] [CrossRef]
- Keshavarz, H.; Modarres-Sanavy, S.A.M.; Mahdipour Afra, M. Organic and chemical fertilizer affected yield and essential oil of two mint species. J. Essent. Oil-Bear. Plants. 2018, 21, 1674–1681. [Google Scholar] [CrossRef]
- Hessini, K.; Wasli, H.; Al-Yasi, H.M.; Ali, E.F.; Issa, A.A.; Hassan, F.A.; Siddique, K.H. Graded moisture deficit effect on secondary metabolites, antioxidant, and inhibitory enzyme activities in leaf extracts of Rosa damascena Mill. var. trigentipetala. Hortic 2022, 8, 177. [Google Scholar] [CrossRef]
- Baghbani-Arani, A.; Modarres-Sanavy, S.A.M.; Mashhadi-Akbar-Boojar, M.; Mokhtassi-Bidgoli, A. Towards improving the agronomic performance, chlorophyll fluorescence parameters and pigments in fenugreek using zeolite and vermicompost under deficit water stress. Ind Crops Prod. 2017, 109, 346–357. [Google Scholar] [CrossRef]
- Zhaoxiang, W.; Huihu, L.; Qiaoli, L.; Changyan, Y.; Faxin, Y. Application of bio-organic fertilizer, not biochar, in degraded red soil improves soil nutrients and plant growth. Rhizosphere 2020, 16, 100264. [Google Scholar] [CrossRef]
- Khorasaninejad, S.; Alizadeh Ahmadabadi, A.; Hemmati, K. The effect of humic acid on leaf morphophysiological and phytochemical properties of Echinacea purpurea L. under water deficit stress. Sci. Hortic. 2018, 239, 314–323. [Google Scholar] [CrossRef]
- Khan, A.U.; Ullah, F.; Khan, N.; Mehmood, S.; Fahad, S.; Datta, R.; Irshad, I.; Danish, S.; Saud, S.; Alaraidh, I.A.; et al. Production of organic fertilizers from rocket seed (Eruca Sativa L.), chicken peat and Moringa Oleifera leaves for growing linseed under water deficit stress. Sustainability 2021, 13, 59. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Souri, M.K.; Mousavi, A.; Sahebani, N. Biochar and vermicompost improve growth and physiological traits of eggplant (Solanum melongena L.) under deficit irrigation. Chem. Biol. Technol. Agric. 2021, 8, 19. [Google Scholar] [CrossRef]
- Hameed, A.; Bibi, N.; Akhter, J.; Iqbal, N. Differential changes in antioxidants, proteases, and lipid peroxidation in flag leaves of wheat genotypes under different levels of water deficit conditions. Plant Physiol. Biochem. 2011, 49, 178–185. [Google Scholar] [CrossRef]
- Mirzamohammadi, K.H.; Modarres-Sanavy, S.A.M.; Sefidkon, F.; Mokhtassi-Bidgoli, A.; Mirjalili, M.H. Irrigation and fertilizer treatments affecting rosmarinic acid accumulation, total phenolic content, antioxidant potential and correlation between them in peppermint (Mentha piperita L.). Irrig. Sci. 2021, 39, 671–683. [Google Scholar] [CrossRef]
- de Assis, R.M.A.; Carneiro, J.J.; Medeiros, A.P.R.; de Carvalho, A.A.; da Cunha Honorato, A.; Carneiro, M.A.C.; Bertolucci, S.K.V.; Pinto, J.E.B.P. Arbuscular mycorrhizal fungi and organic manure enhance growth and accumulation of citral, total phenols, and flavonoids in Melissa officinalis L. Ind. Crops Prod. 2020, 158, 112981. [Google Scholar] [CrossRef]
- Jayathilake, C.; Rizliya, V.; Liyanage, R. Antioxidant and free radical scavenging capacity of extensively used medicinal plants in Sri Lanka. Procedia Food. Sci. 2016, 6, 123–126. [Google Scholar] [CrossRef]
- Kumar, U.; Mishra, M.; Prakash, V. Assessment of antioxidant enzymes and free radical scavenging activity of selected medicinal plants. Free Radic. Antioxid. 2012, 2, 58–63. [Google Scholar] [CrossRef]
- Rahimi, A.; Mohammadi, M.M.; Moghadam, S.S.; Heydarzadeh, S.; Gitari, H. Effects of stress modifier biostimulants on vegetative growth, nutrients and antioxidants contents of garden thyme (Thymus vulgaris L.) under water stress. J. Plant Growth Regul. 2022, 42, 1–14. [Google Scholar] [CrossRef]
- Soratto, R.P.; Perdoná, M.J.; Parecido, R.J.; Pinotti, R.N.; Gitari, H.I. Turning biennial into biannual harvest: Long-term assessment of Arabica coffee–macadamia intercropping and irrigation synergism by biological and economic indices. Food Energy Secur. 2022, 11, 365. [Google Scholar] [CrossRef]
- Kulak, M.; Jorrín-Novo, J.V.; Romero-Rodriguez, M.C.; Yildirim, E.D.; Gul, F.; Karaman, S. Seed priming with salicylic acid on plant growth and essential oil composition in basil (Ocimum basilicum L.) plants grown under water stress conditions. Ind. Crops Prod. 2021, 161, 113235. [Google Scholar] [CrossRef]
- Sheshbahreh, M.J.; Dehnavi, M.M.; Salehi, A.; Bahreininejad, B. Effect of irrigation regimes and nitrogen sources on biomass production, water and nitrogen use efficiency and nutrients uptake in coneflower (Echinacea purpurea L.). Agric. Water Manag. 2019, 213, 358–367. [Google Scholar] [CrossRef]
- Mohamed, Y.; Mohamed, I.; Elsadek, M.; Ali, M.; Ghatas, Y. Improving growth, productivity, and chemical composition of Trachyspermum ammi L. by using organic and chemical fertilization in the presence of boron. Ind. Crops Prod. 2021, 169, 113637. [Google Scholar] [CrossRef]
- Mohammadzadeh, S.; Pirzad, A. Biochemical responses of mycorrhizal-inoculated Lamiaceae (Lavender, Rosemary and Thyme) plants to drought: A field study. J. Soil Sci. Plant Nutr. 2021, 67, 41–49. [Google Scholar] [CrossRef]
- Faridvand, S.; Rezaei-Chiyaneh, E.; Battaglia, M.; Gitari, H.; Raza, M.A.; Siddique, K.H.M. Application of bio and chemical fertilizers improves yield, and essential oil quantity and quality of Moldavian balm (Dracocephalum moldavica L.) intercropped with mung bean (Vigna radiata L.). Food Energy Secur. 2021, 11, 319. [Google Scholar] [CrossRef]
- Wani, S.P.; Gopalakrishnan, S. Plant growth-promoting microbes for sustainable agriculture. In Plant Growth Promoting Rhizobacteria (PGPR): Prospects for Sustainable Agriculture; Springer: Singapore, 2019; pp. 19–45. [Google Scholar]
- Benami, A.; Ofen, A. Irrigation engineering-sprinkler, trickle and surface irrigation: Principles, design and agricultural practices. In Irrigation Engineering Scientific Publications; Walker, W.R., Ed.; Irrigation Engineering Scientific Publications: Haifa, Israel, 1984; p. 257. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determination of total carotenoids and chlorophylls a and b of leaf in different solvents. Biochem. Soc. Trans. 1987, 11, 591–592. [Google Scholar] [CrossRef]
- Khosravi, S.A.; Rezaei Nejad, A.; Khandan Mirkohi, A.; Kalate Jari, S. Vermicompost and manure compost reduce water-deficit stress in pot marigold (Calendula officinalis L. cv. Candyman orange). Compost Sci. Land Util. 2019, 27, 61–68. [Google Scholar] [CrossRef]
- Tejera, G.N.A.; Olivera, M.; Iribarne, C.; Lluch, C. Partial purification and characterization of a non-specific acid phosphatase in leaves and root nodules of Phaseolus vulgaris. Plant Physiol. Biochem. 2004, 42, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Nakano, A.; Asada, K. Purification of ascorbate peroxidase in spinach chloroplasts: Its inactivation in ascorbate-depleted medium and reactivation by monodehydroascorbate radical. Plant Cell Physiol. 1987, 28, 131–140. [Google Scholar]
- Farnad, N.; Heidari, R.; Aslanipour, B. Phenolic composition and comparison of antioxidant activity of alcoholic extracts of peppermint (Mentha piperita). J. Food Meas. Charact. 2014, 8, 113–121. [Google Scholar] [CrossRef]
- Sakanaka, S.; Tachibana, Y.; Okada, Y. Preparation and antioxidant properties of extracts of Japanese persimmon leaf tea (kakinoha-cha). Food Chem. 2005, 89, 569–575. [Google Scholar] [CrossRef]
- Shin, Y.; Liu, R.H.; Nock, J.F.; Holliday, D.; Watkins, C.B. Temperature and relative humidity effects on quality, total ascorbic acid, phenolics and flavonoid concentrations, and antioxidant activity of strawberry. Postharvest Biol. Technol. 2007, 45, 349–357. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. Food Sci. Biotechnol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Ling, T.Y.; Zhao, X.Y. The improved pyrogallol method by using terminating agent for superoxide dismutase measurement. Progress Biochem. Biophys. 1995, 22, 84–86. [Google Scholar]
- Perkin, E. Analytical Methods for Atomic Absorbtion Spectrophotometry; Perkin-Elmer Inc.: Waltham, MA, USA, 1982. [Google Scholar]
- Waling, I.; Van Vark, W.; Houba, V.J.G.; Van der Lee, J.J. Soil and plant analysis, a series of syllabi: Part 7. In Plant Analysis Procedures Wageningen Agriculture University; Wageningen University: Wageningen, The Netherlands, 1989. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Joshi, N.L.; Mali, P.C.; Saxena, A. Effect of nitrogen and sulphur application on yield and fatty acid composition of mustard (Brassica juncea L.) oil. J. Agron. Crop Sci. 1998, 180, 59–63. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chl a | Chl b | Chl a+b | Car | RWC | CAT | SOD | APX | ||
|---|---|---|---|---|---|---|---|---|---|
| Irrigation conditions | |||||||||
| Supplemental irrigation | 2.76 a | 1.96 a | 4.73 a | 1.12 a | 70.19 a | 2.72 a | 65.47 a | 1.34 a | |
| Rainfed | 2.09 b | 1.43 b | 3.51 b | 1.01 b | 66.12 b | 1.78 b | 40.08b | 0.65 b | |
| Different fertilizer sources | |||||||||
| Vermicompost | 2.65 a | 1.87 a | 4.52 a | 1.19 a | 77.48 a | 2.72 a | 59.57 a | 1.15 a | |
| Chemical fertilizer | 2.46 c | 1.80 a | 4.24 b | 1.03 bc | 74.83 a | 2.11 d | 54.90 b | 0.96 c | |
| Poultry manure | 2.61 ab | 1.83 a | 4.44 a | 1.18 a | 76.89 a | 2.51 b | 57.07 ab | 1.11 b | |
| Animal manure | 2.54 b | 1.70 b | 4.26 b | 1.08 bc | 67.12 b | 2.32 c | 56.42 b | 1.07 b | |
| Biofertilizer | 2.22 d | 1.59 c | 3.81 c | 1.00 c | 62.84 c | 2.06 d | 46.63 c | 0.89 d | |
| Control | 2.07 e | 1.38 d | 3.46 d | 0.93 d | 49.46 d | 1.77 e | 42.03 d | 0.77 e | |
| Sources of variations | df | Mean Square | |||||||
| Block (Repetition) | 2 | 0.00004 | 0.02 | 0.01 | 0.002 | 13.59 | 0.0003 | 1.10 | 0.0001 |
| Irrigation (I) | 1 | 4.14 ** | 2.57 ** | 13.29 ** | 0.11 ** | 148.79 ** | 8.00 ** | 5804.66 ** | 4.26 ** |
| Fertilizer (F) | 5 | 0.32 ** | 0.19 ** | 1 ** | 0.06 ** | 690.54 ** | 0.69 ** | 282.99 ** | 0.12 ** |
| I × F | 5 | 0.0004 ns | 0.002 ns | 0.002 ns | 0.0008 ns | 6.95 ns | 0.04 ** | 110.07 ** | 0.02 ** |
| Error | 22 | 0.003 | 0.005 | 0.009 | 0.002 | 6.15 | 0.002 | 5.59 | 0.001 |
| C.V (%) | 2.44 | 4.42 | 2.32 | 4.62 | 3.63 | 2.14 | 4.48 | 3.44 | |
| TPC | TFC | DPPH RS | SRS | CBAC | ||
|---|---|---|---|---|---|---|
| Irrigation conditions | ||||||
| Supplemental irrigation | 22.08 a | 1.24 a | 48.75 a | 43.57 a | 3.78 a | |
| Rainfed | 16.76 b | 0.61 b | 38.76 b | 41.25 b | 2.86 b | |
| Different fertilizer sources | ||||||
| Vermicompost | 23.46 a | 1.07 a | 50.17 a | 45.97 a | 3.72 a | |
| Chemical fertilizer | 18.82c | 0.89 c | 45.20 bc | 42.24 bc | 3.46 b | |
| Poultry manure | 21.70 b | 1.04 a | 47.36 ab | 45.08 a | 3.64 a | |
| Animal manure | 21.06 b | 0.98 b | 44.92 bc | 43.49 ab | 3.44 b | |
| Biofertilizer | 17.66 d | 0.83 d | 42.47 c | 40.27 c | 3.01 c | |
| Control | 13.83 e | 0.73 e | 32.41 d | 37.39 d | 2.66 d | |
| Sources of variations | df | Mean Square | ||||
| Block (Repetition) | 2 | 1.10 | 0.0002 | 3.19 | 0.89 | 0.01 |
| Irrigation (I) | 1 | 255.30 ** | 2.54 ** | 898.40 ** | 48.51 ** | 7.54 ** |
| Fertilizer (F) | 5 | 70.75 ** | 0.24 ** | 225.44 ** | 60.91 ** | 1.00 ** |
| I × F | 5 | 5.48 ** | 0.12 ** | 2.07 ns | 1.08 ns | 0.002 ns |
| Error | 22 | 0.48 | 0.001 | 9.95 | 4.83 | 0.009 |
| C.V (%) | 3.57 | 4.37 | 4.58 | 5.18 | 2.88 | |
| BY | GY | FO | FOY | EO | EOY | P | K | ||
|---|---|---|---|---|---|---|---|---|---|
| Irrigation conditions | |||||||||
| Supplemental irrigation | 4629.15 a | 636.65 a | 24.40 a | 157.07 a | 0.14 b | 0.91 a | 0.66 a | 18.26 a | |
| Rainfed | 2518.45 b | 427.71 b | 22.08 b | 95.06 b | 0.17 a | 0.73 b | 0.59 b | 15.92 b | |
| Different fertilizer sources | |||||||||
| Vermicompost | 4394.90 a | 594.62 a | 25.97 a | 155.28 a | 0.18 a | 1.05 a | 0.74 a | 18.19 a | |
| Chemical fertilizer | 3757.20 c | 536.90 b | 23.24 bc | 131.98 b | 0.15 d | 0.83 d | 0.62 c | 16.96 a | |
| Poultry manure | 4153.60 ab | 579.45 a | 25.08 ab | 143.73 ab | 0.17 b | 0.99 b | 0.67 b | 17.71 b | |
| Animal manure | 4026.70 bc | 547.16 b | 23.49 bc | 128.13 bc | 0.16 c | 0.83 c | 0.59 cd | 17.45 b | |
| Biofertilizer | 2767.40 d | 492.41 c | 22.27 c | 110.39 c | 0.13 e | 0.66 e | 0.57 d | 16.53 d | |
| Control | 2343 e | 442.55 d | 19.39 d | 86.87 d | 0.12 f | 0.51 f | 0.53 e | 15.69 e | |
| Source of variation | df | Mean Square | |||||||
| Block (Repetition) | 2 | 31900.32 | 93.36 | 0.89 | 44.35 | 0.00006 | 0.001 | 0.0006 | 0.28 |
| Irrigation (I) | 1 | 10,023,862.05 ** | 310,044.80 ** | 48.51 ** | 34607.78 ** | 0.008 ** | 0.27 ** | 0.04 ** | 49 ** |
| Fertilizer (F) | 5 | 1,024,266.40 ** | 29,355.85 ** | 31.97 ** | 3584.23 ** | 0.003 ** | 0.25 ** | 0.03 ** | 4.81 ** |
| I × F | 5 | 232,976.53 ** | 13,046.31 ** | 1.08 ns | 676.56 ns | 0.0001 ns | 0.01 ** | 0.01 ns | 0.23 ns |
| Error | 22 | 20,479.44 | 503.32 | 4.83 | 256.57 | 0.00002 | 0.002 | 0.0007 | 0.10 |
| C.V(%) | 8.01 | 4.31 | 9.45 | 12.70 | 3.34 | 5.50 | 4.50 | 1.93 | |
| Characteristics of Fertilizer | C (%) | N (%) | P (%) | K (%) |
|---|---|---|---|---|
| Poultry manure | 4.5 | 1.75 | 2.8 | 2.15 |
| Animal manure | 2.9 | 2.27 | 0.94 | 1.25 |
| Vermicompost | 3.8 | 1.46 | 2.4 | 1.95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heydarzadeh, S.; Arena, C.; Vitale, E.; Rahimi, A.; Mirzapour, M.; Nasar, J.; Kisaka, O.; Sow, S.; Ranjan, S.; Gitari, H. Impact of Different Fertilizer Sources under Supplemental Irrigation and Rainfed Conditions on Eco-Physiological Responses and Yield Characteristics of Dragon’s Head (Lallemantia iberica). Plants 2023, 12, 1693. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12081693
Heydarzadeh S, Arena C, Vitale E, Rahimi A, Mirzapour M, Nasar J, Kisaka O, Sow S, Ranjan S, Gitari H. Impact of Different Fertilizer Sources under Supplemental Irrigation and Rainfed Conditions on Eco-Physiological Responses and Yield Characteristics of Dragon’s Head (Lallemantia iberica). Plants. 2023; 12(8):1693. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12081693
Chicago/Turabian StyleHeydarzadeh, Saeid, Carmen Arena, Ermenegilda Vitale, Amir Rahimi, Mohsen Mirzapour, Jamal Nasar, Oscar Kisaka, Sumit Sow, Shivani Ranjan, and Harun Gitari. 2023. "Impact of Different Fertilizer Sources under Supplemental Irrigation and Rainfed Conditions on Eco-Physiological Responses and Yield Characteristics of Dragon’s Head (Lallemantia iberica)" Plants 12, no. 8: 1693. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12081693