Using Morphogenic Genes to Improve Recovery and Regeneration of Transgenic Plants

 , ,
, ,
Abstract
:1. Introduction
2. Phenotypic Responses from Ectopic Overexpression of Morphogenic Genes
2.1. Enhancing the Somatic Embryogenic Response
2.2. Ectopic Formation of Somatic Embryos or Meristems
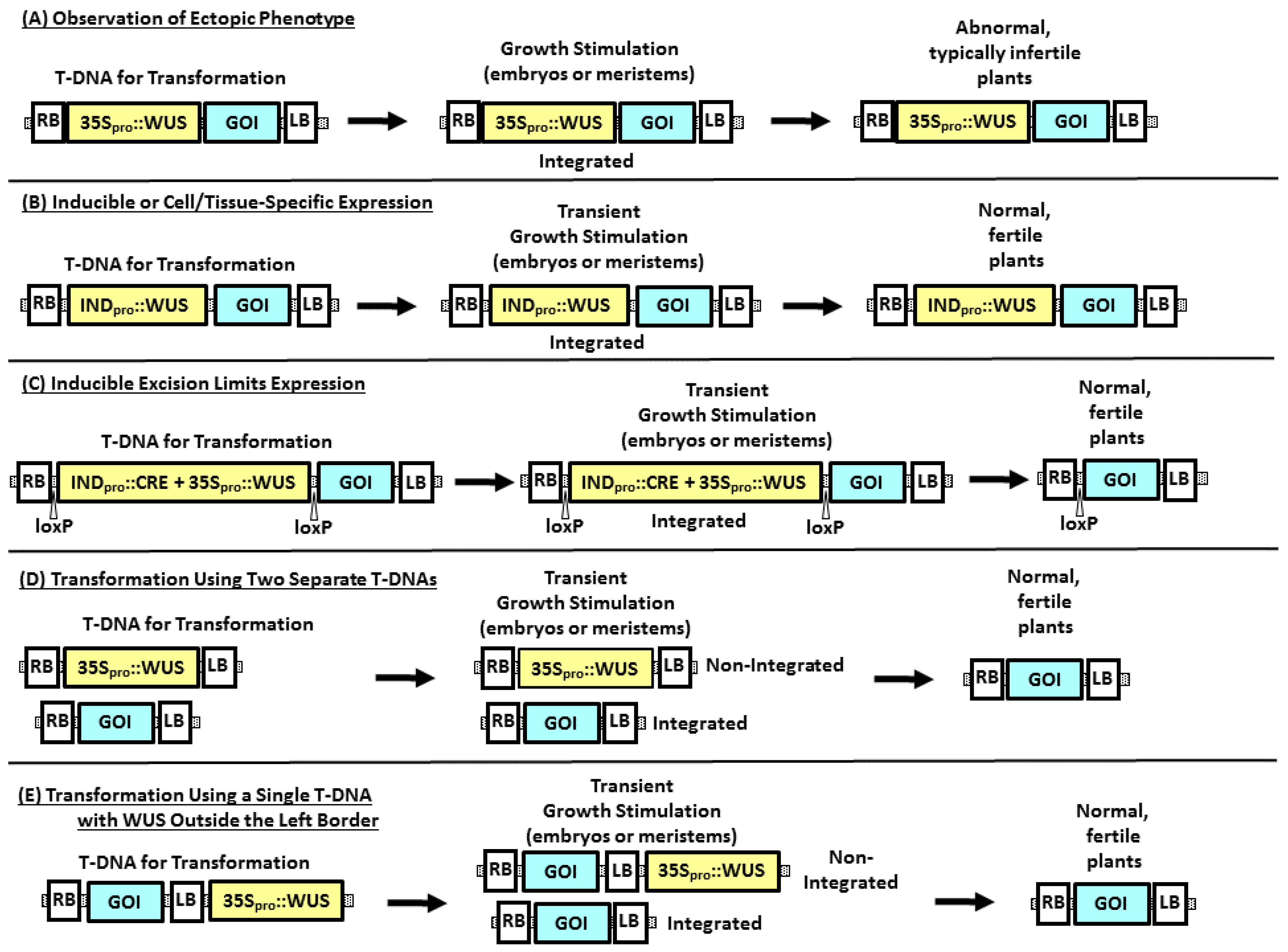
3. Strategies to Improve Transformation Using Morphogenic Genes
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Altpeter, F.; Springer, N.M.; Bartley, L.E.; Blechl, A.E.; Brutnell, T.P.; Citovsky, V.; Conrad, L.J.; Gelvin, S.B.; Jackson, D.P.; Kausch, A.P.; et al. Advancing Crop Transformation in the Era of Genome Editing. Plant Cell 2016, 28, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Smertenko, A.; Bozhkov, P.V. Somatic embryogenesis: Life and death processes during apical-basal patterning. J. Exp. Bot. 2014, 65, 1343–1360. [Google Scholar] [CrossRef] [PubMed]
- Snipes, S.A.; Rodriguez, K.; DeVries, A.E.; Miyawaki, K.N.; Perales, M.; Xie, M.; Reddy, G.V. Cytokinin stabilizes WUSCHEL by acting on the protein domains required for nuclear enrichment and transcription. PLOS Genet. 2018, 14, e1007351. [Google Scholar] [CrossRef] [PubMed]
- Grienenberger, E.; Fletcher, J.C. Polypeptide signaling molecules in plant development. Curr. Opin. Plant Biol. 2015, 23, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.K. Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Dev. Biol. 2010, 341, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef]
- Skoog, F.; Miller, C.O. Chemical regulation of growth and organ formation in plant tissues cultured. In Vitro Symp. Soc. Exp. Biol. 1957, 11, 118–130. [Google Scholar]
- Murashige, T. Plant propagation through tissue cultures. Ann. Rev. Plant Physiol. 1974, 25, 135–166. [Google Scholar] [CrossRef]
- Smigocki, A.C.; Owens, L.D. Cytokinin gene fused with a strong promoter enhances shoot organogenesis and zeatin levels in transformed plant cells. Proc. Natl. Acad. Sci. USA 1998, 85, 5131–5135. [Google Scholar] [CrossRef]
- Ondřej, M.; Macháčková, I.; Čatský, J.; Eder, J.; Hrouda, M.; Pospíšilová, J.; Synková, H. Potato transformation by T-DNA cytokinin synthesis gene. Biol. Plant. 1990, 32, 401–406. [Google Scholar] [CrossRef]
- Casanova, E.; Valdes, A.E.; Zuker, A.; Frenandez, B.; Vainstein, A.; Trillas, M.I. rolC-transgenic carnation plants: Adventitious organogenesis and levels of endogenous auxin and cytokinins. Plant Sci. 2004, 167, 551–560. [Google Scholar] [CrossRef]
- Gorpenchenko, T.Y.; Kiselev, K.V.; Bulgakov, V.P.; Tchernoded, G.K.; Bragina, E.A.; Khodakovskaya, M.V.; Koren, O.G.; Batygina, T.B.; Zhuravlev, Y.N. The Agrobacterium rhizogenes rolC-gene-induced somatic embryogenesis and shoot organogenesis in Panax ginseng transformed calluses. Planta 2006, 223, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Otten, L. The Agrobacterium Phenotypic Plasticity (Plast) Genes. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Roeckel, P.; Oancia, T.; Drevet, J.R. Phenotypic alterations and component analysis of seed yield in transgenic Brassica napus plants expressing the tzs gene. Physiol. Plant. 1998, 102, 243–249. [Google Scholar] [CrossRef]
- Choi, Y.I.; Noh, E.W.; Choi, K.S. Low level expression of prokaryotic tzs gene enhances growth performance of transgenic poplars. Trees 2009, 23, 7441–7750. [Google Scholar] [CrossRef]
- Gordon-Kamm, W.; Dilkes, B.P.; Lowe, K.; Hoerster, G.; Sun, X.; Ross, M.; Church, L.; Bunde, C.; Farrell, J.; Hill, P.; et al. Stimulation of the cell cycle and maize transformation by disruption of the plant retinoblastoma pathway. Proc. Nat. Acad. Sci. USA 2002, 99, 11975–11980. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic Regulators Baby boom and Wuschel Improve Monocot Transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.; Velle-Calzada, J.P.; Hartog, M.V.; Schmidt, E.D.; Boutilier, K.; Grossniklaus, U.; de Vries, S.C. The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE 1 gene is expressed in developing ovules and embryos and enhances embryogenic competence in culture. Plant Physiol. 2001, 127, 803–816. [Google Scholar] [CrossRef]
- Pérez-Pascual, D.; Jiménez-Guillen, D.; Villanueva-Alonzo, H.; Souza-Perera, R.; Godoy-Hernández, G.; Zúñiga-Aguilar, J.J. Ectopic expression of the Coffea canephora SERK1 homolog-induced differential transcription of genes involved in auxin metabolism and in the developmental control of embryogenesis. Physiol. Plant. 2018, 163, 530–551. [Google Scholar] [CrossRef]
- Harding, E.W.; Tang, W.; Nichols, K.W.; Fernandez, D.E.; Perry, S.E. Expression and maintenance of embryogenic potential is enhanced through constitutive expression of AGAMOUS-LIKE15. Plant Physiol. 2003, 133, 653–663. [Google Scholar] [CrossRef]
- Thakare, D.; Tang, W.; Hill, K.; Perry, S.E. The MADS-domain transcriptional regulator AGAMOUS-LIKE15 promotes somatic embryo development in Arabidopsis and soybean. Plant Physiol. 2008, 146, 1663–1672. [Google Scholar] [CrossRef]
- Yang, Z.; Li, C.; Wang, Y.; Zhang, C.; Wu, Z.; Zhang, X.; Liu, C.; Li, F. GhAGL15s, preferentially expressed during somatic embryogenesis, promote embryogenic callus formation in cotton (Gossypium hirsutum L.). Mol. Genet. Genom. 2014, 289, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Herrera, A.; Gonzalez, A.K.; Moo, R.C.; Quiroz-Figueroa, F.R.; Loyola-Vargas, V.M.; Rodriguez-Zapata, L.C.; Burgeff D′Hondt, C.; Suárez-Solís, V.M.; Castaño, E. Expression of WUSCHEL in Coffea canephora causes ectopic morphogenesis and increases somatic embryogenesis. Plant Cell Tissue Organ Cult. 2008, 94, 171–180. [Google Scholar] [CrossRef]
- Elhiti, M.; Tahir, M.; Gulden, R.H.; Khamiss, K.; Stasolla, C. Modulation of embryo-forming capacity in culture through the expression of Brassica genes involved in the regulation of the shoot apical meristem. J. Exp. Bot. 2010, 61, 4069–4085. [Google Scholar] [CrossRef] [PubMed]
- Bouchabké-Coussa, O.; Obellianne, M.; Linderme, D.; Montes, E.; Maia-Grondard, A.; Vilaine, F.; Pannetier, C. Wuschel overexpression promotes somatic embryogenesis and induces organogenesis in cotton (Gossypium hirsutum L.) tissues cultured in vitro. Plant Cell Rep. 2013, 32, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.M.; van Lammeren, A.A.; Miki, B.L.; et al. Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, C.; Liu, Z.; Heidmann, I.; Supena, E.D.; Fukuoka, H.; Joosen, R.; Lambalk, J.; Angenent, G.; Scorza, R.; Custers, J.B.; et al. Heterologous expression of the BABY BOOM AP2/ERF transcription factor enhances the regeneration capacity of tobacco (Nicotiana tabacum L.). Planta 2007, 225, 341–351. [Google Scholar] [CrossRef]
- El Ouakfaoui, S.; Schnell, J.; Abdeen, A.; Colville, A.; Labbé, H.; Han, S.; Baum, B.; Laberge, S.; Miki, B. Control of somatic embryogenesis and embryo development by AP2 transcription factors. Plant Mol. Biol. 2010, 74, 313–326. [Google Scholar] [CrossRef]
- 29 Florez, S.L.; Erwin, R.L.; Maximova, S.N.; Guiltinan, M.J.; Curtis, W.R. Enhanced somatic embryogenesis in Theobroma cacao using the homologous BABY BOOM transcription factor. BMC Plant Biol. 2015, 16, 121. [Google Scholar] [CrossRef]
- Morcillo, F.; Gallard, A.; Pillot, M.; Jouannic, S.; Aberlenc-Bertossi, F.; Collin, M.; Verdeil, J.L.; Tregear, J.W. EgAP2-1, an AINTEGUMENTA-like (AIL) gene expressed in meristematic and proliferating tissues of embryos in oil palm. Planta 2007, 226, 1353–1362. [Google Scholar] [CrossRef]
- Tsuwamoto, R.; Yokoi, S.; Takahata, Y. Arabidopsis EMBRYOMAKER encoding an AP2 domain transcription factor plays a key role in developmental change from vegetative to embryonic phase. Plant Mol. Biol. 2010, 73, 481–492. [Google Scholar] [CrossRef]
- Mursyanti, E.; Purwantoro, A.; Moeljopawiro, S.; Semiarti, E. Induction of Somatic Embryogenesis through Overexpression of ATRKD4 Genes in Phalaenopsis “Sogo Vivien”. Indones. J. Biotechnol. 2015, 20, 42–53. [Google Scholar] [CrossRef]
- Lotan, T.; Ohto, M.; Yee, K.M.; West, M.A.; Lo, R.; Kwong, R.W.; Yamagishi, K.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells. Cell 1998, 93, 1195–1205. [Google Scholar] [CrossRef]
- Zhu, S.-P.; Wang, J.; Ye, J.-L.; Zhu, A.-D.; Guo, W.-W.; Deng, X.-X. Isolation and characterization of LEAFY COTYLEDON 1-LIKE gene related to embryogenic competence in Citrus sinensis. Plant Cell Tiss. Organ Cult. 2014, 119, 1–13. [Google Scholar] [CrossRef]
- Uddenberg, D.; Abrahamsson, M.; von Arnold, S. Overexpression of PaHAP3A stimulates differentiation of ectopic embryos from maturing somatic embryos of Norway spruce. Tree Genet. Genomes 2016, 12, 18. [Google Scholar] [CrossRef]
- Gazzarrini, S.; Tsuchiya, Y.; Lumba, S.; Okamoto, M.; McCourt, P. The transcription factor FUSCA3 controls developmental timing in Arabidopsis through the hormones gibberellin and abscisic acid. Dev. Cell 2004, 7, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Kwong, L.W.; Yee, K.M.; Pelletier, J.; Lepiniec, L.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON2 encodes a B3 domain transcription factor that induces embryo development. Proc. Natl. Acad. Sci. USA 2001, 98, 11806–11811. [Google Scholar] [CrossRef]
- Zuo, J.; Niu, Q.W.; Frugis, G.; Chua, N.H. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. Plant J. 2002, 30, 349–359. [Google Scholar] [CrossRef]
- Gallois, J.L.; Woodward, C.; Reddy, G.V.; Sablowski, R. Combined SHOOT MERISTEMLESS and WUSCHEL trigger ectopic organogenesis in Arabidopsis. Development 2002, 129, 3207–3217. [Google Scholar]
- Gallois, J.L.; Nora, F.R.; Mizukami, Y.; Sablowski, R. WUSCHEL induces shoot stem cell activity and developmental plasticity in the root meristem. Genes Dev. 2004, 18, 375–380. [Google Scholar] [CrossRef]
- Rashid, S.Z.; Yamaji, N.; Kyo, M. Shoot formation from root tip region: A developmental alteration by WUS in transgenic tobacco. Plant Cell Rep. 2007, 26, 1449–1455. [Google Scholar] [CrossRef]
- Rashid, S.Z.; Kyo, M. Ectopic Expression of WOX5 Dramatically Alters Root-tip Morphology in Transgenic Tobacco. Transgenic Plant J. 2009, 3, 92–96. [Google Scholar]
- Luo, K.; Zheng, X.; Chen, Y.; Xiao, Y.; Zhao, D.; McAvoy, R.; Pei, Y.; Li, Y. The maize Knotted1 gene is an effective positive selectable marker gene for Agrobacterium-mediated tobacco transformation. Plant Cell Rep. 2006, 25, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Tamaoki, M.; Sakamoto, T.; Matsuoka, M. Over-Expression of Tobacco knotted 1-Type Classl Homeobox Genes Alters Various Leaf Morphology. Plant Cell Physiol. 2000, 41, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Daimon, Y.; Takabe, K.; Tasaka, M. The CUP-SHAPED COTYLEDON genes promote adventitious shoot formation on calli. Plant Cell. Physiol. 2003, 44, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Wójcikowska, B.; Jaskóła, K.; Gąsiorek, P.; Meus, M.; Nowak, K.; Gaj, M.D. LEAFY COTYLEDON2 (LEC2) promotes embryogenic induction in somatic tissues of Arabidopsis, via YUCCA-mediated auxin biosynthesis. Planta 2013, 238, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Banno, H.; Ikeda, Y.; Niu, Q.W.; Chua, N.H. Overexpression of Arabidopsis ESR1 induces initiation of shoot regeneration. Plant Cell 2001, 13, 2609–2618. [Google Scholar] [CrossRef]
- Ikeda, Y.; Banno, H.; Niu, Q.W.; Howell, S.H.; Chua, N.H. The ENHANCER OF SHOOT REGENERATION 2 gene in Arabidopsis regulates CUP-SHAPED COTYLEDON 1 at the transcriptional level and controls cotyledon development. Plant Cell Physiol. 2006, 47, 1443–1456. [Google Scholar] [CrossRef]
- Ckurshumova, W.; Smirnova, T.; Marcos, D.; Zayed, Y.; Berleth, T. Irrepressible MONOPTEROS/ARF5 promotes de novo shoot formation. New Phytol. 2014, 204, 556–566. [Google Scholar] [CrossRef]
- Heidmann, I.; de Lange, B.; Lambalk, J.; Angenent, G.C.; Boutilier, K. Efficient sweet pepper transformation mediated by the BABY BOOM transcription factor. Plant Cell Rep. 2011, 30, 1107–1115. [Google Scholar] [CrossRef]
- Lutz, K.A.; Martin, C.; Khairzada, S.; Maliga, P. Steroid-inducible BABY BOOM system for development of fertile Arabidopsis thaliana plants after prolonged tissue culture. Plant Cell Rep. 2015, 34, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Niu, Q.-W.; Teng, C.; Li, C.; Mu, J.; Chua, N.-H.; Zuo, J. Overexpression of PGA37/MYB118 and MYB115 promotes vegetative-to-embryonic transition in Arabidopsis. Cell Res. 2009, 19, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Shires, M.E.; Florez, S.L.; Lai, T.S.; Curtis, W.R. Inducible somatic embryogenesis in Theobroma cacao achieved using the DEX-activatable transcription factor-glucocorticoid receptor fusion. Biotechnol. Lett. 2017, 39, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Kyo, M.; Maida, K.; Nishioka, Y.; Matsui, K. Coexpression of WUSCHEL related homeobox (WOX) 2 with WOX8 or WOX9 promotes regeneration from leaf segments and free cells in Nicotiana tabacum L. Plant Biotechnol. 2018, 35, 23–30. [Google Scholar] [CrossRef]
- Deng, W.; Li, Z.; Luo, K.; Yang, Y. A novel method for induction of plant regeneration via somatic embryogenesis. Plant Sci. 2009, 177, 43–48. [Google Scholar] [CrossRef]
- Mookkan, M.; Nelson-Vasilchik, K.; Hague, J.; Zhang, Z.J.; Kausch, A.P. Selectable marker independent transformation of recalcitrant maize inbred B73 and sorghum P898012 mediated by morphogenic regulators BABY BOOM and WUSCHEL2. Plant Cell Rep. 2017, 36, 1477–1491. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; La Rota, M.; Hoerster, G.; Hastings, C.; Wang, N.; Chamberlin, M.; Wu, E.; Jones, T.; Gordon-Kamm, W. Rapid genotype “independent” Zea mays L. (maize) transformation via direct somatic embryogenesis. In Vitro Cell Dev. Biol. Plant. 2018, 54, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Karlova, R.; Boeren, S.; Russinova, E.; Aker, J.; Vervoort, J.; de Vries, S. The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE1 protein complex includes brassinosteroid-insensitive1. Plant Cell 2006, 18, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Gou, X.; Yin, H.; He, K.; Du, J.; Yi, J.; Xu, S.; Lin, H.; Clouse, S.D. Genetic Evidence for an Indispensable Role of Somatic Embryogenesis Receptor Kinases in Brassinosteroid Signaling. PLOS Genet. 2012, 8, e1002452. [Google Scholar] [CrossRef]
- Santiago, J.; Henzler, C.; Hothorn, M. Molecular mechanism for plant steroid receptor activation by somatic embryogenesis co-receptor kinases. Science 2013, 341, 889–892. [Google Scholar] [CrossRef]
- Hohmann, U.; Lau, K.; Hothorn, M. The structural basis of ligand perception and signal activation by receptor kinases. Ann. Rev. Plant Biol. 2017, 68, 109–137. [Google Scholar] [CrossRef]
- Laux, T.; Mayer, K.F.; Berger, J.; Jurgens, G. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar] [PubMed]
- Long, J.A.; Moan, E.I.; Medford, J.I.; Barton, M.K. A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA 2/Ethylene Responsive Factor (AP 2/ERF) transcription factors: Mediators of stress responses and developmental programs. New Phytol. 2013, 199, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Waki, T.; Kiki, T.; Watanabe, R. The Arabidopsis RWP-RK Protein RKD4 Triggers Gene Expression and Pattern Formation in Early Embryogenesis. Curr. Biol. 2011, 21, 1277–1281. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Palmer, T.M.; Lukowitz, W. The RWP-RK Factor GROUNDED Promotes Embryonic Polarity by Facilitating YODA MAP Kinase Signaling. Curr. Biol. 2011, 21, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Santos Mendoza, M.; Dubreucq, B.; Miquel, M.; Caboche, M.; Lepiniec, L. LEAFY COTYLEDON 2 activation is sufficient to trigger the accumulation of oil and seed specific mRNAs in Arabidopsis leaves. FEBS Lett. 2005, 579, 4666–4670. [Google Scholar] [CrossRef] [PubMed]
- Weigel, D.; Alvarez, J.; Smyth, D.R.; Yanofsky, M.F.; Meyerowitz, E.M. LEAFY controls floral meristem identity in Arabidopsis. Cell 1992, 69, 843–859. [Google Scholar] [CrossRef]
- Nagle, M.; Déjardin, A.; Pilate, G.; Strauss, S.H. Opportunities for innovation in genetic transformation of forest trees. Front. Plant Sci. 2018, 9, 1443. [Google Scholar] [CrossRef]
- Miller, M.; Tagliani, L.; Wang, N.; Berka, B.; Bidney, D.; Zhao, Z.-Y. High Efficiency Transgene Segregation in Co-Transformed Maize Plants using an Agrobacterium Tumefaciens 2 T-DNA Binary System. Transgenic Res. 2002, 11, 381–396. [Google Scholar] [CrossRef]
- Kononov, M.E.; Bassuner, B.; Gelvin, S.B. Integration of T-DNA binary vector ‘backbone’ sequences into the tobacco genome: Evidence for multiple complex patterns of integration. Plant J. 1997, 11, 945–957. [Google Scholar] [CrossRef]
- Hanson, B.; Engler, D.; Moy, Y.; Newman, B.; Ralston, E.; Gutterson, N. A simple method to enrich an Agrobacterium-transformed population for plants containing only T-DNA sequences. Plant J. 1999, 19, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Lee, J.; Jun, S.H.; Park, S.; Kang, H.G.; Kwon, S.; An, G. Transgene structures in TDNA-inserted rice plants. Plant Mol. Biol. 2003, 52, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Podevin, N.; De Buck, S.; De Wilde, C.; Depicker, A. Insights into recognition of the T-DNA border repeats as termination sites for T-strand synthesis by Agrobacterium tumefaciens. Transgenic Res. 2006, 15, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Richael, C.; Kalyeava, M.; Chretien, R.C.; Rommens, C.M. Cytokinin vectors mediate marker-free and backbone-free plant transformation. Transgenic Res. 2008, 17, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; Hoerster, G.; Anand, A.; Arling, M.; Wang, N.; McBride, K.; Gordon-Kamm, W. Transient expression of morphogenic genes positioned outside the T-DNA borders results in rapid formation of somatic embryos and fertile transgenic cereal plants. Unpublished; manuscript in preparation.
- Ananiev, E.V.; Wu, C.; Chamberlin, M.A.; Svitashev, S.; Schwartz, C.; Gordon-Kamm, W.; Tingey, S. Artificial chromosome formation in maize (Zea mays L.). Chromosoma 2009, 118, 157–177. [Google Scholar] [CrossRef] [PubMed]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted Mutagenesis, Precise Gene Editing, and Site-Specific Gene Insertion in Maize Using Cas9 and Guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Cigan, A.M. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Examples | |||||
|---|---|---|---|---|---|
| Strategy | CDS | Promoter for Transgene | Transformed Species | Variety * | Ref. |
| (A) Enhance pre-existing somatic embryogenic culture response | AtSERK1 | 35S | Arabidopsis thaliana | Ws | [18] |
| CcSERK1 (or RNAi) | 35S and Inducible | Coffea canephora | cv. Robusta | [19] | |
| AtAGL15 | 35S | A. thaliana | Ws | [20] | |
| GmAGL15 | 35S | Glycine max | “Jack” | [21] | |
| GhAGL15 | 35S | Gossypium hirsutum | cv. CRI24 | [22] | |
| AtWUS | Inducible | C. canephora | cv. Robusta | [23] | |
| BnSTM, BoSTM | 35S | A. thaliana | Col | [24] | |
| BnSTM, BoSTM | 35S | Brassica napus | cv. Topas | [24] | |
| BrSTM | 35S | A. thaliana | Col | [24] | |
| BrSTM | 35S | B. napus | cv. Topas | [24] | |
| AtWUS | 35S | G. hirsutum | var. Coker 310 | [25] | |
| (B) Ectopic formation of somatic embryos or meristems | BnBBM | 35S | A. thaliana | Col and C24 | [26] |
| AtBBM~GR | Inducible | Nicotiana tabacum | Wisconsin 38 | [27] | |
| AtBBM~GR | Inducible | N. tabacum | Petit Havana SR1 | [27] | |
| GmBBM | 35S | A. thaliana | not specified | [28] | |
| TcBBM | 35S | Theobroma cacao | Scavina-6 (SCA6) | [29] | |
| EgBBM | 35S | A. thaliana | Col | [30] | |
| AtEMK | 35S | A. thaliana | Col-0 | [31] | |
| AtRKD4 | Inducible | P.sp (Orchid) | “Sogo Vivian” | [32] | |
| AtLEC1 | 35S | A. thaliana | Ws-0 | [33] | |
| CsL1L | 35S | Clonorchis sinensis | cv. “Olinda” | [34] | |
| PaHAP3A | Inducible | Picea abies | cell lines 88 and 61 | [35] | |
| AtFUS3 | AtML1 | A. thaliana | Col | [36] | |
| AtLEC2 | 35S | A. thaliana | Ws-0 | [37] | |
| AtWUS | Inducible | A. thaliana | Col, Ws, Ler | [38] | |
| AtWUS, AtSTM | Both inducible | A. thaliana | Ler | [39] | |
| AtWUS | Activated | A. thaliana | Ler | [40] | |
| AtWUS | Inducible | N. tabacum | cv. Samsun | [41] | |
| AtWOX5 | Inducible | N. tabacum | cv. Samsun | [42] | |
| ZmKN1 | 35S | N. tabacum | cv. Xanthi | [43] | |
| NtKN1 | 35S | N. tabacum | cv. Samsun | [44] | |
| AtCUC1, AtCUC2 | 35S | A. thaliana | Ler | [45] | |
| AtLEC2 | Inducible | A. thaliana | Col-0 | [46] | |
| AtESR1 | Inducible | A. thaliana | Ws | [47] | |
| AtESR2 | Inducible | A. thaliana | Ler and Ws | [48] | |
| AtMPΔ | MP Promoter | A. thaliana | Col-0 | [49] | |
| (C) Restrict morphogenic response to enable recovery of normal plants | BnBBM | Inducible | Capsicum annuum | Three hybrids b | [50] |
| AtBBM | Inducible | A. thaliana | RDL and Ler | [51] | |
| AtPGA37 | Inducible | A. thaliana | Col-0, Ws, Ler | [52] | |
| AtLEC2 | Inducible | T. cacao | var. SCA6 | [53] | |
| AtWOX2 WOX8 WOX9 | Inducible | N. tabacum | cv. Samsun | [54] | |
| BcBBMa | 35S | Populus tomentosa | not specified | [55] | |
| ZmBBM/ZmWUS2a | Ubi + NOS | Zea mays | 4 Pioneer Inbreds c | [17] | |
| ZmBBM/ZmWUS2a | Ubi + NOS | Oryza sativa | (indica) cv. IRV95 | [17] | |
| ZmBBM/ZmWUS2 a | Ubi + NOS | Sorghum bicolor | var. Tx430 | [17] | |
| ZmBBM/ZmWUS2 a | Ubi + NOS | Salvia officianalis | var. CP01-1372 | [17] | |
| ZmBBM/ZmWUS2 a | Ubi + NOS | Z. mays | public inbred B73 | [56] | |
| ZmBBM/ZmWUS2 a | Ubi + NOS | S. bicolor | var. P898012 | [56] | |
| ZmBBM/ZmWUS2 | PLTP + AXIG1 | Z. mays | maize inbreds d | [57] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordon-Kamm, B.; Sardesai, N.; Arling, M.; Lowe, K.; Hoerster, G.; Betts, S.; Jones, T. Using Morphogenic Genes to Improve Recovery and Regeneration of Transgenic Plants. Plants 2019, 8, 38. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8020038
Gordon-Kamm B, Sardesai N, Arling M, Lowe K, Hoerster G, Betts S, Jones T. Using Morphogenic Genes to Improve Recovery and Regeneration of Transgenic Plants. Plants. 2019; 8(2):38. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8020038
Chicago/Turabian StyleGordon-Kamm, Bill, Nagesh Sardesai, Maren Arling, Keith Lowe, George Hoerster, Scott Betts, and Todd Jones. 2019. "Using Morphogenic Genes to Improve Recovery and Regeneration of Transgenic Plants" Plants 8, no. 2: 38. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8020038