Phytolith Formation in Plants: From Soil to Cell

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Phytolith Formation in Plants
2.1. Silicon Uptake and Transport
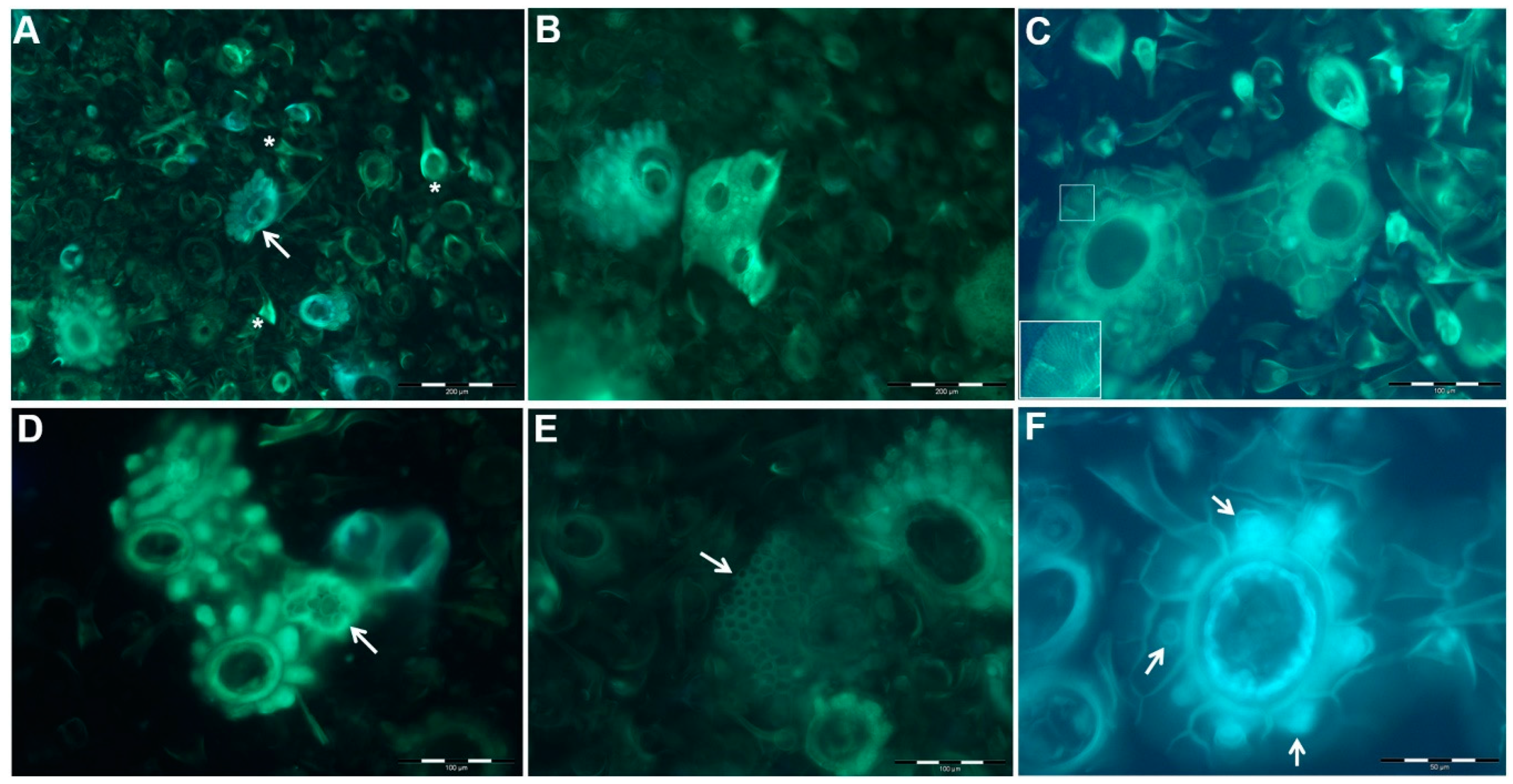
2.2. Si Deposition and the Formation of Phytoliths
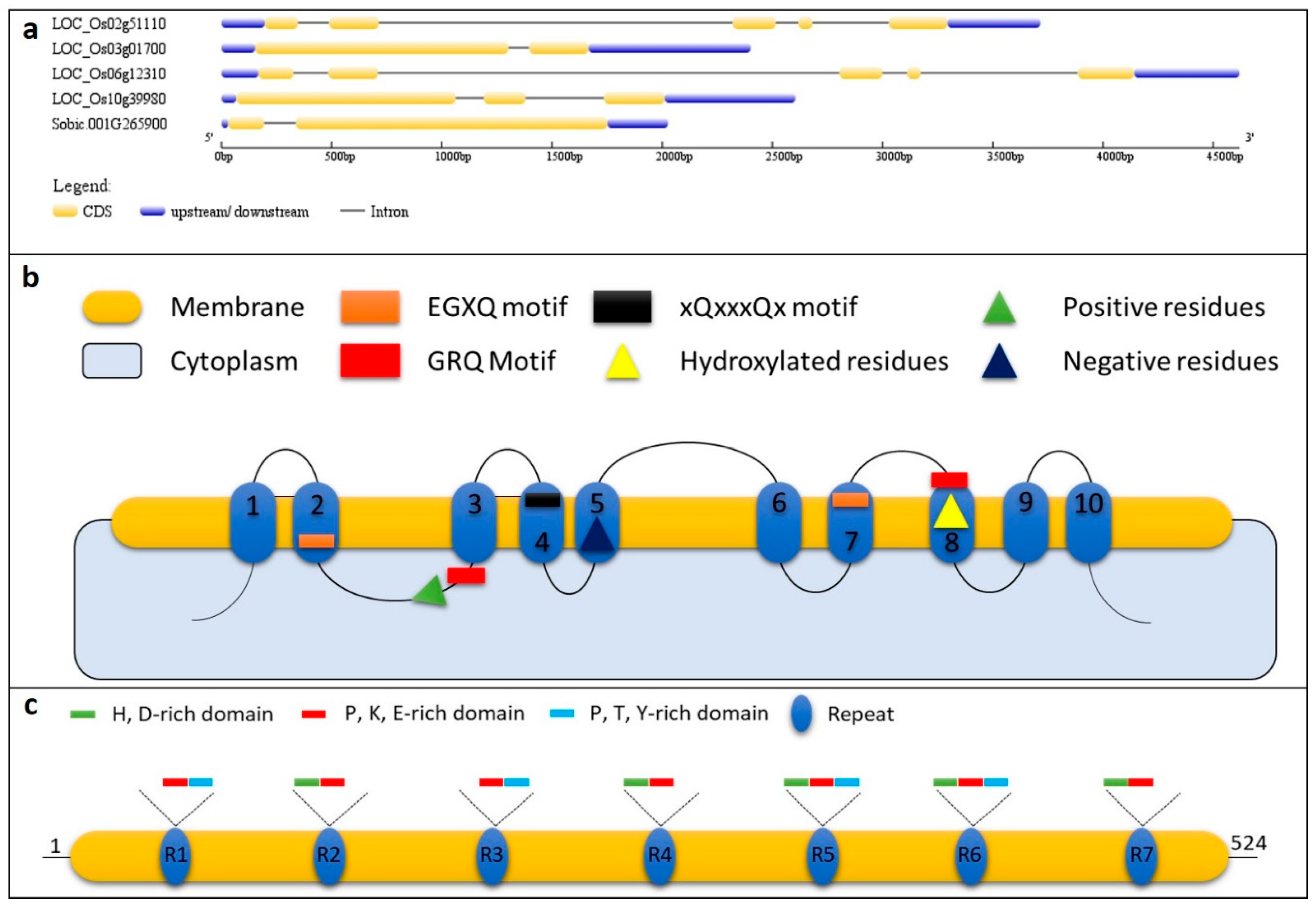
3. Proteins/Genes of Biosilicification in Plants
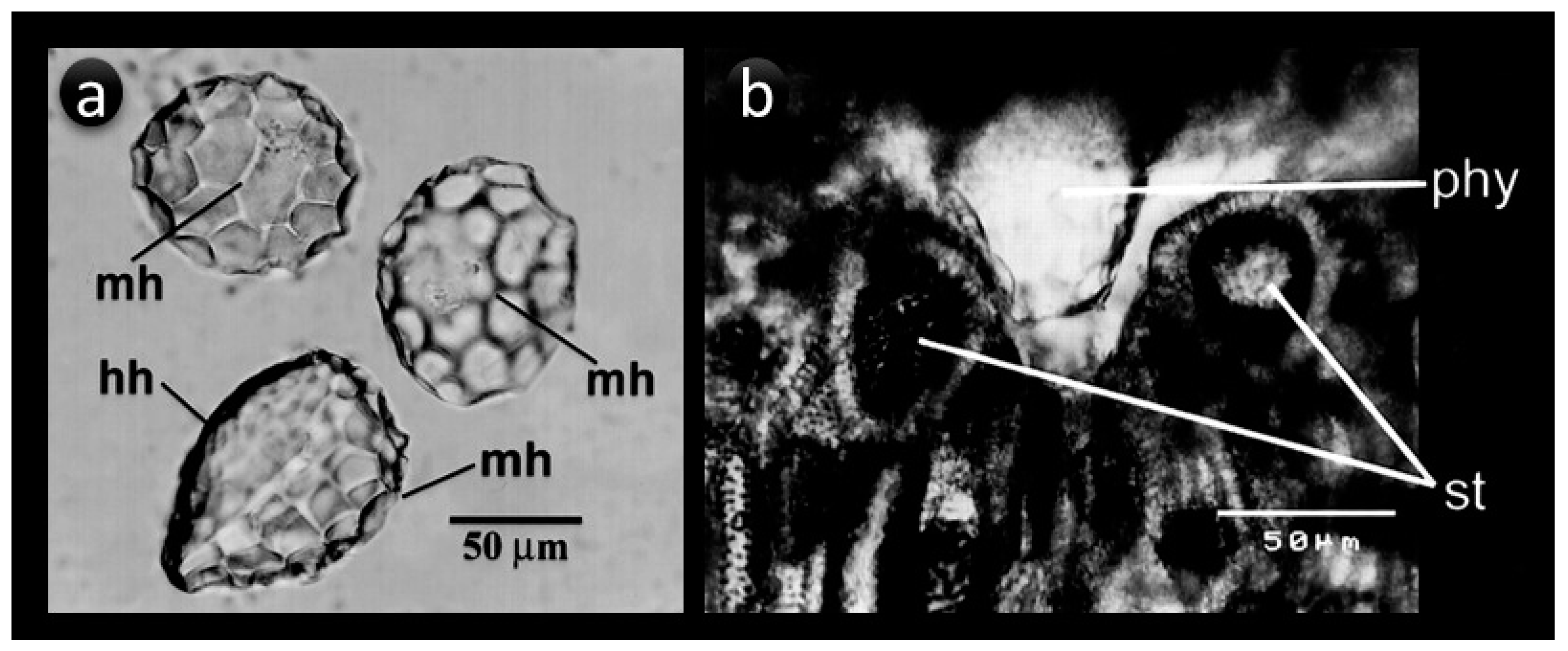
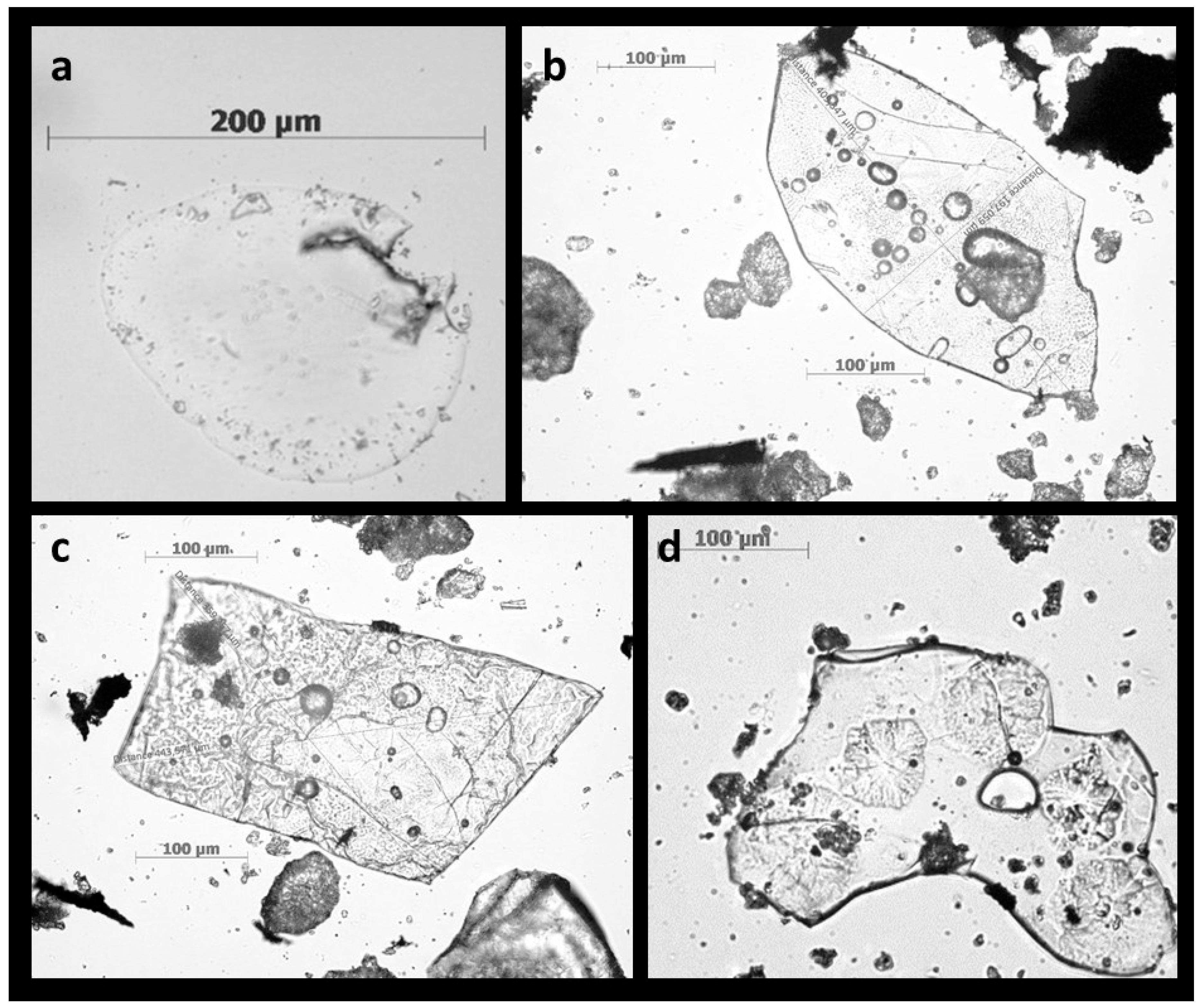
4. Shape, Size, and Chemistry of Phytoliths
5. Evolution of Biosilicification in Plants
6. Functions of Phytoliths
6.1. Biotic and Abiotic Stress Tolerance
6.2. Water Stress Tolerance
6.3. Salt and Metal Stress Tolerance
6.4. Tolerance to Pathogens
6.5. Role of Phytoliths against Herbivory
6.6. Role of Phytoliths as Mechanical Barriers
6.7. Phytoliths as Taxonomic Tools
6.7.1. Identification of Genus and Species
6.7.2. Differentiation between Wild and Cultivated Species
6.7.3. Historic Uses of Plants by Humans and Investigating Cereal Economies
6.7.4. Reconstruction of Ancient Floras, Landscapes, and Palaeoenvironments
6.7.5. Discrimination of Forests and Grass Covers and C4/C3 Grasslands
6.7.6. Limitations Associated with the Use of Phytoliths as Taxonomic Tools
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Piperno, D.R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists; Rowman Altamira: Lanham, MD, USA, 2006. [Google Scholar]
- Katz, O. Plant silicon and phytolith research and the Earth-life superdiscipline. Front. Plant Sci. 2018, 9, 1281. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Veneklaas, E.J.; Kuo, J.; Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant Sci. 2014, 19, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Hodson, M.J. The development of phytoliths in plants and its influence on their chemistry and isotopic composition. Implications for palaeoecology and archaeology. J. Archaeol. Sci. 2016, 68, 62–69. [Google Scholar] [CrossRef]
- Liu, L.; Jie, D.; Liu, H.; Gao, G.; Gao, Z.; Li, D.; Li, N.; Guo, J.; Qiao, Z. Assessing the importance of environmental factors to phytoliths of Phragmites communis in north-eastern China. Ecol. Indic. 2016, 69, 500–507. [Google Scholar] [CrossRef]
- Strömberg, C.A.; Dunn, R.E.; Crifò, C.; Harris, E.B. Phytoliths in paleoecology: Analytical considerations, current use, and future directions. In Methods in Paleoecology; Springer Nature Publisher: New York, NY, USA, 2018; pp. 235–287. [Google Scholar]
- Madella, M.; Alexandre, A.; Ball, T. International code for phytolith nomenclature 1.0. Ann. Bot. 2005, 96, 253–260. [Google Scholar] [CrossRef]
- Albert, R.M.; Ruiz, J.A.; Sans, A. PhytCore ODB: A new tool to improve efficiency in the management and exchange of information on phytoliths. J. Archaeol. Sci. 2016, 68, 98–105. [Google Scholar] [CrossRef]
- Raven, J.A. Cycling silicon–the role of accumulation in plants. New Phytol. 2003, 158, 419–421. [Google Scholar] [CrossRef]
- Golohvast, K.S.; Chaika, V.V.; Romashko, R.V.; Galkina, A.N.; Zemchenko, I.A.; Sergievich, A.A.; Artemenko, A.F.; Bulakh, E.M.; Dzubenko, E.V.; Seryodkin, I.V. Can Plants and Fungi Cure «The Wounds» Using Silicone Gel? Defect Diffus. Forum 2018, 386, 268–274. [Google Scholar] [CrossRef]
- Luyckx, M.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Impact of silicon in plant biomass production: Focus on bast fibres, hypotheses, and perspectives. Plants 2017, 6, 37. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. A cooperative system of silicon transport in plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Meunier, J.D.; Barboni, D.; Anwar-ul-Haq, M.; Levard, C.; Chaurand, P.; Vidal, V.; Grauby, O.; Huc, R.; Laffont-Schwob, I.; Rabier, J. Effect of phytoliths for mitigating water stress in durum wheat. New Phytol. 2017, 215, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conley, D.J.; Frings, P.J.; Fontorbe, G.; Clymans, W.; Stadmark, J.; Hendry, K.R.; Marron, A.O.; De La Rocha, C.L. Biosilicification drives a decline of dissolved Si in the oceans through geologic time. Front. Mar. Sci. 2017, 4, 397. [Google Scholar] [CrossRef]
- Cornelis, J.T.; Ranger, J.; Iserentant, A.; Delvaux, B. Tree species impact the terrestrial cycle of silicon through various uptakes. Biogeochemistry 2010, 97, 231–245. [Google Scholar] [CrossRef]
- Cornelis, J.T.; Delvaux, B. Soil processes drive the biological silicon feedback loop. Funct. Ecol. 2016, 30, 1298–1310. [Google Scholar] [CrossRef]
- Conley, D.J. Terrestrial ecosystems and the global biogeochemical silica cycle. Glob. Biogeochem. Cycles 2002, 16. [Google Scholar] [CrossRef]
- RAVEN, J.A. The transport and function of silicon in plants. Biol. Rev. 1983, 58, 179–207. [Google Scholar] [CrossRef]
- Currie, H.A.; Perry, C.C. Silica in Plants: Biological, Biochemical and Chemical Studies. Ann. Bot. 2007, 100. [Google Scholar] [CrossRef]
- Neethirajan, S.; Gordon, R.; Wang, L. Potential of silica bodies (phytoliths) for nanotechnology. Trends Biotechnol. 2009, 27, 461–467. [Google Scholar] [CrossRef]
- Haynes, R.J. A contemporary overview of silicon availability in agricultural soils. J. Plant Nutr. Soil Sci. 2014, 177, 831–844. [Google Scholar] [CrossRef]
- Gocke, M.; Liang, W.; Sommer, M.; Kuzyakov, Y. Silicon uptake by wheat: Effects of Si pools and pH. J. Plant Nutr. Soil Sci. 2013, 176, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Ma, J.F.; Goto, S.; Tamai, K.; Ichii, M. Role of root hairs and lateral roots in silicon uptake by rice. Plant Physiol. 2001, 127, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Miyake, Y.; Takahashi, E. Silicon as a beneficial element for crop plants. In Studies in Plant Science; Elsevier: Cambridge, MA, USA, 2001; Volume 8, pp. 17–39. [Google Scholar]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.K.; Sonah, H.; Belanger, R.R. Plant Aquaporins: Genome-Wide Identification, Transcriptomics, Proteomics, and Advanced Analytical Tools. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, E.; Ma, J.; Miyake, Y. The possibility of silicon as an essential element for higher plants. Comments Agric. Food Chem. 1990, 2, 99–102. [Google Scholar]
- Chiba, Y.; Mitani, N.; Yamaji, N.; Ma, J.F. HvLsi1 is a silicon influx transporter in barley. Plant J. 2009, 57, 810–818. [Google Scholar] [CrossRef]
- Sakurai, G.; Satake, A.; Yamaji, N.; Mitani-Ueno, N.; Yokozawa, M.; Feugier, F.G.; Ma, J.F. In silico simulation modeling reveals the importance of the Casparian strip for efficient silicon uptake in rice roots. Plant Cell Physiol. 2015, 56, 631–639. [Google Scholar] [CrossRef]
- Hu, J.; Cai, X.; Jeong, B.R. Silicon Affects Root Development, Tissue Mineral Content, and Expression of Silicon Transporter Genes in Poinsettia (Euphorbia pulcherrima Willd.) Cultivars. Plants 2019, 8, 180. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Vivancos, J.; Guerin, V.; Sonah, H.; Labbe, C.; Belzile, F.; Belanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Yamaji, N.; Mitatni, N.; Ma, J.F. A transporter regulating silicon distribution in rice shoots. Plant Cell 2008, 20, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Mitani, N.; Yamaji, N.; Ago, Y.; Iwasaki, K.; Ma, J.F. Isolation and functional characterization of an influx silicon transporter in two pumpkin cultivars contrasting in silicon accumulation. Plant J. 2011, 66, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Mitani-Ueno, N.; Yamaji, N.; Ma, J.F. Silicon efflux transporters isolated from two pumpkin cultivars contrasting in Si uptake. Plant Signal. Behav. 2011, 6, 991–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, N.; Chiba, Y.; Mitani-Ueno, N.; Ma, J.F. Functional Characterization of a Silicon Transporter Gene Implicated in Silicon Distribution in Barley. Plant Physiol. 2012, 160, 1491–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai-Ishikawa, J.; Murai-Hatano, M.; Hayashi, H.; Ahamed, A.; Fukushi, K.; Matsumoto, T.; Kitagawa, Y. Transpiration from shoots triggers diurnal changes in root aquaporin expression. Plant Cell Environ. 2011, 34, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- Casey, W.; Kinrade, S.; Knight, C.; Rains, D.; Epstein, E. Aqueous silicate complexes in wheat, Triticum aestivum L. Plantcell Environ. 2004, 27, 51–54. [Google Scholar] [CrossRef]
- Mitani, N.; Ma, J.F. Uptake system of silicon in different plant species. J. Exp. Bot. 2005, 56, 1255–1261. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, N.; Sakurai, G.; Mitani-Ueno, N.; Ma, J.F. Orchestration of three transporters and distinct vascular structures in node for intervascular transfer of silicon in rice. Proc. Natl. Acad. Sci. USA 2015, 112, 11401–11406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detmann, K.C.; Araújo, W.L.; Martins, S.C.; Sanglard, L.M.; Reis, J.V.; Detmann, E.; Rodrigues, F.Á.; Nunes-Nesi, A.; Fernie, A.R.; DaMatta, F.M. Silicon nutrition increases grain yield, which, in turn, exerts a feed-forward stimulation of photosynthetic rates via enhanced mesophyll conductance and alters primary metabolism in rice. New Phytol. 2012, 196, 752–762. [Google Scholar] [CrossRef]
- Jung, J.; Kim, J.; Jin, B.; Choi, Y.; Hong, C.O.; Lee, H.H.; Choi, Y.; Kang, J.; Park, Y. Characterization of the Lsi1 Homologs in Cucurbita moschata and C. ficifolia for Breeding of Stock Cultivars Used for Bloomless Cucumber Production. Hortic. Sci. Technol. 2017, 35, 333–343. [Google Scholar] [CrossRef]
- Wang, H.S.; Yu, C.; Fan, P.P.; Bao, B.F.; Li, T.; Zhu, Z.J. Identification of Two Cucumber Putative Silicon Transporter Genes in Cucumis sativus. J. Plant Growth Regul. 2015, 34, 332–338. [Google Scholar] [CrossRef]
- Montpetit, J.; Vivancos, J.; Mitani-Ueno, N.; Yamaji, N.; Remus-Borel, W.; Belzile, F.; Ma, J.F.; Belanger, R.R. Cloning, functional characterization and heterologous expression of TaLsi1, a wheat silicon transporter gene. Plant Mol. Biol. 2012, 79, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Bokor, B.; Ondos, S.; Vaculik, M.; Bokorova, S.; Weidinger, M.; Lichtscheidl, I.; Turna, J.; Lux, A. Expression of Genes for Si Uptake, Accumulation, and Correlation of Si with Other Elements in Ionome of Maize Kernel. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, G.; Yamaji, N.; Mitani-Ueno, N.; Yokozawa, M.; Ono, K.; Ma, J.F. A Model of Silicon Dynamics in Rice: An Analysis of the Investment Efficiency of Si Transporters. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, C.C.; Belton, D.; Shafran, K. Studies of biosilicas; structural aspects, chemicalprinciples, model studies and the future. In Silicon Biomineralization; Springer: Berlin, Germany, 2003; pp. 269–299. [Google Scholar]
- Exley, C. A possible mechanism of biological silicification in plants. Front. Plant Sci. 2015, 6, 853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, C.C.; Mann, S.; Williams, R.J.P. Structural and analytical studies of the silicified macrohairs from the lemma of the grass Phalaris canariensis L. Proc. R. Soc. London. Ser. B. Biol. Sci. 1984, 222, 427–438. [Google Scholar]
- Mann, S.; Perry, C. Structural aspects of biogenic silica. In Silicon the Biochemistry, CIBA Foundation Symposium; John Wiley and Sons: Chichester, UK, 1986; Volume 121. [Google Scholar]
- Perry, C.C.; Fraser, M. Silica deposition and ultrastructure in the cell wall of Equisetum arvense: The importance of cell wall structures and flow control in biosilicification? Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1991, 334, 149–157. [Google Scholar]
- Parry, D.W.; Smithson, F. Types of opaline silica depositions in the leaves of British grasses. Ann. Bot. 1964, 28, 169–185. [Google Scholar] [CrossRef]
- Sangster, A.; Hodson, M.; Tubb, H. Silicon deposition in higher plants. In Studies in Plant Science; Elsevier: Cambridge, MA, USA, 2001; Volume 8, pp. 85–113. [Google Scholar]
- Pamirsky, I.; Golokhvast, K. Silaffins of diatoms: From applied biotechnology to biomedicine. Mar. Drugs 2013, 11, 3155–3167. [Google Scholar] [CrossRef]
- Hildebrand, M.; Lerch, S.J.; Shrestha, R.P. Understanding Diatom Cell Wall Silicification-Moving Forward. Front. Mar. Sci. 2018, 5, 125. [Google Scholar] [CrossRef]
- Guerriero, G.; Deshmukh, R.; Sonah, H.; Sergeant, K.; Hausman, J.-F.; Lentzen, E.; Valle, N.; Siddiqui, K.S.; Exley, C. Identification of the aquaporin gene family in Cannabis sativa and evidence for the accumulation of silicon in its tissues. Plant Sci. 2019, 110167. [Google Scholar] [CrossRef]
- Schulz-Kornas, E.; Braune, C.; Winkler, D.E.; Kaiser, T.M. Does silica concentration and phytolith ultrastructure relate to phytolith hardness? Biosurf. Biotribol. 2017, 3, 135–143. [Google Scholar] [CrossRef]
- Kaufman, P.; Dayanandan, P.; Takeoka, Y.; Bigelow, W.; Jones, J.; Iler, R. Silica in shoots of higher plants. In Silicon and Siliceous Structures in Biological Systems; Springer Nature Publisher: New York, NY, USA, 1981; pp. 409–449. [Google Scholar]
- Kaufman, P.B.; Petering, L.B.; Smith, J.G. Ultrastructural development of cork-silica cell pairs in Avena internodal epidermis. Bot. Gaz. 1970, 131, 173–185. [Google Scholar] [CrossRef]
- Kumar, S.; Milstein, Y.; Brami, Y.; Elbaum, M.; Elbaum, R. Mechanism of silica deposition in sorghum silica cells. New Phytol. 2017, 213, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Soukup, M.; Elbaum, R. Silicification in grasses: Variation between different cell types. Front. Plant Sci. 2017, 8, 438. [Google Scholar] [CrossRef] [PubMed]
- Kauss, H.; Seehaus, K.; Franke, R.; Gilbert, S.; Dietrich, R.A.; Kröger, N. Silica deposition by a strongly cationic proline-rich protein from systemically resistant cucumber plants. Plant J. 2003, 33, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Motomura, H.; Mita, N.; Suzuki, M. Silica accumulation in long-lived leaves of Sasa veitchii (Carrière) Rehder (Poaceae–Bambusoideae). Ann. Bot. 2002, 90, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Motomura, H.; Fujii, T.; Suzuki, M. Silica deposition in abaxial epidermis before the opening of leaf blades of Pleioblastus chino (Poaceae, Bambusoideae). Ann. Bot. 2006, 97, 513–519. [Google Scholar] [CrossRef]
- Exley, C. Silicon in life: Whither biological silicification. In Biosilica in Evolution, Morphogenesis, and Nanobiotechnology; Springer: Berlin, Germany, 2009; pp. 173–184. [Google Scholar]
- Zhang, C.; Wang, L.; Zhang, W.; Zhang, F. Do lignification and silicification of the cell wall precede silicon deposition in the silica cell of the rice (Oryza sativa L.) leaf epidermis? Plant Soil 2013, 372, 137–149. [Google Scholar] [CrossRef]
- Kumar, S.; Elbaum, R. Interplay between silica deposition and viability during the life span of sorghum silica cells. New Phytol. 2018, 217, 1137–1145. [Google Scholar] [CrossRef]
- Kumar, S.; Adiram-Filiba, N.; Blum, S.; Sanchez-Lopez, J.A.; Tzfadia, O.; Omid, A.; Volpin, H.; Heifetz, Y.; Goobes, G.; Elbaum, R. Siliplant1 (SIp1) protein precipitates silica in sorghum silica cells. bioRxiv 2019, 518332. [Google Scholar] [CrossRef]
- Sangster, A.; Parry, D. Ultrastructure of silica deposits in higher plants. In Silicon and Siliceous Structures in Biological Systems; Springer: Berlin, Germany, 1981; pp. 383–407. [Google Scholar]
- Sangster, A.; Hodson, M. Silica in higher plants. In Silicon biochemistry; Elsevier: Cambridge, MA, USA, 1986; pp. 90–107. [Google Scholar]
- Hodson, M.; Sangster, A. X-ray microanalytical studies of mineral localization in the needles of white pine (Pinus strobus L.). Ann. Bot. 2002, 89, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Motomura, H.; Fujii, T.; Suzuki, M. Silica deposition in relation to ageing of leaf tissues in Sasa veitchii (Carriere) Rehder (Poaceae: Bambusoideae). Ann. Bot. 2004, 93, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Markovich, O.; Kumar, S.; Cohen, D.; Addadi, S.; Fridman, E.; Elbaum, R. Silicification in leaves of sorghum mutant with low silicon accumulation. Silicon 2015, 1–7. [Google Scholar] [CrossRef]
- Sakai, W.S.; Sanford, W. A developmental study of silicification in the abaxial epidermal cells of sugarcane leaf blades using scanning electron microscopy and energy dispersive X-ray analysis. Am. J. Bot. 1984, 71, 1315–1322. [Google Scholar] [CrossRef]
- Hodson, M.; Sangster, A.; Parry, D.W. An ultrastructural study on the development of silicified tissues in the lemma of Phalaris canariensis L. Proc. R. Soc. Lond. Ser. B. Biol. Sci. 1984, 222, 413–425. [Google Scholar]
- Pattathil, S.; Hahn, M.G.; Dale, B.E.; Chundawat, S.P. Insights into plant cell wall structure, architecture, and integrity using glycome profiling of native and AFEXTM-pre-treated biomass. J. Exp. Bot. 2015, 66, 4279–4294. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Rehman, H.M.; Imtiaz, M.; Baloch, F.S.; Lee, J.D.; Yang, S.H.; Lee, S.I.; Chung, G. Systems Identification and Characterization of Cell Wall Reassembly and Degradation Related Genes in Glycine max (L.) Merill, a Bioenergy Legume. Sci. Rep. 2017, 7, 10862. [Google Scholar] [CrossRef]
- Law, C.; Exley, C. New insight into silica deposition in horsetail (Equisetum arvense). BMC Plant Biol. 2011, 11, 112. [Google Scholar] [CrossRef]
- Pan, W.J.; Song, Z.L.; Liu, H.Y.; Van Zwieten, L.; Li, Y.T.; Yang, X.M.; Han, Y.; Liu, X.; Zhang, X.D.; Xu, Z.J.; et al. The accumulation of phytolith-occluded carbon in soils of different grasslands. J. Soils Sediments 2017, 17, 2420–2427. [Google Scholar] [CrossRef]
- Fry, S.C.; Nesselrode, B.H.; Miller, J.G.; Mewburn, B.R. Mixed-linkage (1→3, 1→4)-β-d-glucan is a major hemicellulose of Equisetum (horsetail) cell walls. New Phytol. 2008, 179, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Gidley, M.J.; Fincher, G.B. Heterogeneity in the chemistry, structure and function of plant cell walls. Nat. Chem. Biol. 2010, 6, 724. [Google Scholar] [CrossRef] [PubMed]
- Kido, N.; Yokoyama, R.; Yamamoto, T.; Furukawa, J.; Iwai, H.; Satoh, S.; Nishitani, K. The matrix polysaccharide (1; 3, 1; 4)-β-D-glucan is involved in silicon-dependent strengthening of rice cell wall. Plant Cell Physiol. 2014, 56, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zou, W.; Li, Y.; Feng, Y.; Zhang, H.; Wu, Z.; Tu, Y.; Wang, Y.; Cai, X.; Peng, L. Silica distinctively affects cell wall features and lignocellulosic saccharification with large enhancement on biomass production in rice. Plant Sci. 2015, 239, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Glazowska, S.; Baldwin, L.; Mravec, J.; Bukh, C.; Hansen, T.H.; Jensen, M.M.; Fangel, J.U.; Willats, W.G.T.; Glasius, M.; Felby, C.; et al. The impact of silicon on cell wall composition and enzymatic saccharification of Brachypodium distachyon. Biotechnol. Biofuels 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.Z.; Gan, X.H.; Mei, C.T.; Liang, Y.F. Structural analysis and transformation of biosilica during lignocellulose fractionation of rice straw. J. Mol. Struct. 2017, 1127, 575–582. [Google Scholar] [CrossRef]
- Maeda, H.; Nakajima, M.; Hagiwara, T.; Sawaguchi, T.; Yano, S. Bacterial cellulose/silica hybrid fabricated by mimicking biocomposites. J. Mater. Sci. 2006, 41, 5646–5656. [Google Scholar] [CrossRef]
- Golokhvast, K.S.; Pamirskiy, I.E.; Panichev, A.M. Homology of bacteria proteins, diatoms and sponges participating in biomineralization, and human proteins, and other animals. Pac. Sci. Rev. 2011, 13, 39–46. [Google Scholar]
- Otzen, D. The role of proteins in biosilicification. Scientifica 2012, 2012, 867562. [Google Scholar] [CrossRef]
- Hildebrand, M.; Volcani, B.E.; Gassmann, W.; Schroeder, J.I. A gene family of silicon transporters. Nature 1997, 385, 688. [Google Scholar] [CrossRef]
- Ma, J. Mechanism of Si uptake in plants. Fertilizer 2003, 94, 26–32. [Google Scholar]
- Wallace, I.S.; Roberts, D.M. Homology modeling of representative subfamilies of Arabidopsis major intrinsic proteins. Classification based on the aromatic/arginine selectivity filter. Plant Physiol. 2004, 135, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Mitani-Ueno, N.; Yamaji, N.; Zhao, F.J.; Ma, J.F. The aromatic/arginine selectivity filter of NIP aquaporins plays a critical role in substrate selectivity for silicon, boron, and arsenic. J. Exp. Bot. 2011, 62, 4391–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, R.K.; Vivancos, J.; Ramakrishnan, G.; Guerin, V.; Carpentier, G.; Sonah, H.; Labbe, C.; Isenring, P.; Belzile, F.J.; Belanger, R.R. A precise spacing between the NPA domains of aquaporins is essential for silicon permeability in plants. Plant J. 2015, 83, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, C.; Remus-Borel, W.; Vivancos, J.; Labbe, C.; Belzile, F.; Belanger, R.R. Discovery of a multigene family of aquaporin silicon transporters in the primitive plant Equisetum arvense. Plant J. 2012, 72, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Vatansever, R.; Ozyigit, I.I.; Filiz, E.; Gozukara, N. Genome-wide exploration of silicon (Si) transporter genes, Lsi1 and Lsi2 in plants; insights into Si-accumulation status/capacity of plants. BioMetals 2017, 30, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Sahebi, M.; Hanafi, M.M.; Abdullah, S.N.A.; Rafii, M.Y.; Azizi, P.; Nejat, N.; Idris, A.S. Isolation and Expression Analysis of Novel Silicon Absorption Gene from Roots of Mangrove (Rhizophora apiculata) via Suppression Subtractive Hybridization. Biomed Res. Int. 2014. [Google Scholar] [CrossRef]
- Song, J.Y.; Jeong, B.R. Effect of silicate source on growth of the Lsi2 transgenic plants of chrysanthemum ‘Brighton’. Hortic. Environ. Biotechnol. 2014, 55, 428–436. [Google Scholar] [CrossRef]
- Vivancos, J.; Deshmukh, R.; Gregoire, C.; Remus-Borel, W.; Belzile, F.; Belanger, R.R. Identification and characterization of silicon efflux transporters in horsetail (Equisetum arvense). J. Plant Physiol. 2016, 200, 82–89. [Google Scholar] [CrossRef]
- Marron, A.O.; Ratcliffe, S.; Wheeler, G.L.; Goldstein, R.E.; King, N.; Not, F.; de Vargas, C.; Richter, D.J. The Evolution of Silicon Transport in Eukaryotes. Mol. Biol. Evol. 2016, 33, 3226–3248. [Google Scholar] [CrossRef] [Green Version]
- Ball, T.; Vrydaghs, L.; Mercer, T.; Pearce, M.; Snyder, S.; Lisztes-Szabo, Z.; Peto, A. A morphometric study of variance in articulated dendritic phytolith wave lobes within selected species of Triticeae and Aveneae. Veg. Hist. Archaeobotany 2017, 26, 85–97. [Google Scholar] [CrossRef]
- Golokhvast, K.S.; Seryodkin, I.V.; Chaika, V.V.; Zakharenko, A.M.; Pamirsky, I.E. Phytoliths in taxonomy of phylogenetic domains of plants. Biomed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Piperno, D.R.; Holst, I.; Wessel-Beaver, L.; Andres, T.C. Evidence for the control of phytolith formation in Cucurbita fruits by the hard rind (Hr) genetic locus: Archaeological and ecological implications. Proc. Natl. Acad. Sci. USA 2002, 99, 10923–10928. [Google Scholar] [CrossRef] [PubMed]
- Huan, X.J.; Lu, H.Y.; Wang, C.; Tang, X.G.; Zuo, X.X.; Ge, Y.; He, K.Y. Bulliform Phytolith Research in Wild and Domesticated Rice Paddy Soil in South China. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Ball, T.B.; Ehlers, R.; Standing, M.D. Review of typologic and morphometric analysis of phytoliths produced by wheat and barley. Breed. Sci. 2009, 59, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.Y.; Liu, K.B. Phytoliths of common grasses in the coastal environments of southeastern USA. Estuar. Coast. Shelf Sci. 2003, 58, 587–600. [Google Scholar] [CrossRef]
- Carnelli, A.L.; Theurillat, J.P.; Madella, A. Phytolith types and type-frequencies in subalpine-alpine plant species of the European Alps. Rev. Palaeobot. Palynol. 2004, 129, 39–65. [Google Scholar] [CrossRef]
- Tubb, H.J.; Hodson, M.J.; Hodson, G.C. The inflorescence papillae of the Triticeae: A new tool for taxonomic and archaeological research. Ann. Bot. 1993, 72, 537–545. [Google Scholar] [CrossRef]
- Dorweiler, J.E.; Doebley, J. Developmental analysis of teosinte glume architecture1: A key locus in the evolution of maize (Poaceae). Am. J. Bot. 1997, 84, 1313–1322. [Google Scholar] [CrossRef]
- Guerriero, G.; Hausman, J.-F.; Legay, S. Silicon and the plant extracellular matrix. Front. Plant Sci. 2016, 7, 463. [Google Scholar] [CrossRef]
- Iriarte, J.; Paz, E.A. Phytolith analysis of selected native plants and modern soils from southeastern Uruguay and its implications for paleoenvironmental and archeological reconstruction. Quat. Int. 2009, 193, 99–123. [Google Scholar] [CrossRef]
- Golokhvast, K.; Chaika, V.; Ugay, S.; Seredkin, I.; Pamirsky, I. First data on the phytoliths of some medicinal plants of Primorye region, Russian Federation. Biol. Med. 2014, 6, BM-022-BM-022. [Google Scholar]
- An, X.-H. Morphological characteristics of phytoliths from representative conifers in China. Palaeoworld 2016, 25, 116–127. [Google Scholar] [CrossRef]
- Esteban, I.; Vlok, J.; Kotina, E.L.; Bamford, M.K.; Cowling, R.M.; Cabanes, D.; Albert, R.M. Phytoliths in plants from the south coast of the Greater Cape Floristic Region (South Africa). Rev. Palaeobot. Palynol. 2017, 245, 69–84. [Google Scholar] [CrossRef]
- Sommer, M.; Kaczorek, D.; Kuzyakov, Y.; Breuer, J. Silicon pools and fluxes in soils and landscapes—A review. J. Plant Nutr. Soil Sci. 2006, 169, 310–329. [Google Scholar] [CrossRef]
- Dayanandan, P.; Kaufman, P.; Franklin, C. Detection of silica in plants. Am. J. Bot. 1983, 70, 1079–1084. [Google Scholar] [CrossRef]
- Liu, H.Y.; Jie, D.M.; Liu, L.D.; Li, N.N.; Wang, T.; Qiao, Z.H. The shape factors of phytoliths in selected plants from the Changbai Mountains and their implications. Rev. Palaeobot. Palynol. 2016, 226, 44–53. [Google Scholar] [CrossRef]
- Li, B.; Feng, Y.Y.; Guo, J.X.; Jie, D.M.; Shi, L.X. Responses of phytolith in guinea grass (Leymus chinensis) leaves to simulated warming, nitrogen deposition and elevated CO2 concentration in Songnen grassland, Northeast China. Chin. Geogr. Sci. 2015, 25, 404–413. [Google Scholar] [CrossRef]
- Liu, L.D.; Jie, D.M.; Liu, H.Y.; Gao, Z.; Gao, G.Z.; Li, N.N.; Guo, J.X.; Qiao, Z.H. An orthogonal experimental study of phytolith size of Phragmites communis in northeast China. Boreas 2016, 45, 122–132. [Google Scholar] [CrossRef]
- Hongyan, L.; Dongmei, J.; Lidan, L.; Zhuo, G.; Guizai, G.; Lianxuan, S.; Jixun, G.; Zhihe, Q. The Research on Phytoliths Size Variation Characteristics in Phragmites communis Under Warming Conditions. Silicon 2018, 10, 445–454. [Google Scholar] [CrossRef]
- Li, R.C.; Fan, J.; Carter, J.; Jiang, N.; Gu, Y.S. Monthly variations of phytoliths in the leaves of the bamboo Dendrocalamus ronganensis (Poaceae: Bambusoideae). Rev. Palaeobot. Palynol. 2017, 246, 62–69. [Google Scholar] [CrossRef]
- Kistler, L.; Haney, J.M.; Newsom, L.A. Experimental investigation of pathogenic stress on phytolith formation in Cucurbita pepo var. texana (wild gourd). Veg. Hist. Archaeobotany 2013, 22, 165–170. [Google Scholar] [CrossRef]
- Rajendiran, S.; Coumar, M.V.; Kundu, S.; Dotaniya, A.M.; Rao, A.S. Role of phytolith occluded carbon of crop plants for enhancing soil carbon sequestration in agro-ecosystems. Curr. Sci. 2012, 103, 911–920. [Google Scholar]
- Qi, L.M.; Li, F.Y.; Huang, Z.T.; Jiang, P.K.; Baoyin, T.G.T.; Wang, H.L. Phytolith-occluded organic carbon as a mechanism for long-term carbon sequestration in a typical steppe: The predominant role of belowground productivity. Sci. Total Environ. 2017, 577, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, K.L.; Alfonso-Garcia, A.; Sanchez, J.; Potma, E.O.; Santos, G.M. Plant growth conditions alter phytolith carbon. Front. Plant Sci. 2015, 6, 753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.; Milne, A.; Sanders, J. Tabashir: An opal of plant origin. Science 1966, 151, 464–466. [Google Scholar] [CrossRef] [PubMed]
- Carnelli, A.L.; Madella, M.; Theurillat, J.P.; Ammann, B. Aluminum in the opal silica reticule of phytoliths: A new tool in palaeoecological studies. Am. J. Bot. 2002, 89, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Kameník, J.; Mizera, J.; Řanda, Z. Chemical composition of plant silica phytoliths. Environ. Chem. Lett. 2013, 11, 189–195. [Google Scholar] [CrossRef]
- Wyttenbach, A.; Tobler, L. Soil contamination in plant samples and in botanical reference materials: Signature, quantification and consequences. J. Radioanal. Nucl. Chem. 2002, 254, 165–174. [Google Scholar] [CrossRef]
- Kučera, J.; Mizera, J.; Řanda, Z.; Vávrová, M. Pollution of agricultural crops with lanthanides, thorium and uranium studied by instrumental and radiochemical neutron activation analysis. J. Radioanal. Nucl. Chem. 2007, 271, 581–587. [Google Scholar] [CrossRef]
- Bujan, E. Elemental composition of phytoliths in modern plants (Ericaceae). Quat. Int. 2013, 287, 114–120. [Google Scholar] [CrossRef]
- Nguyen, M.N.; Dultz, S.; Guggenberger, G. Effects of pretreatment and solution chemistry on solubility of rice-straw phytoliths. J. Plant Nutr. Soil Sci. 2014, 177, 349–359. [Google Scholar] [CrossRef]
- Cabanes, D.; Shahack-Gross, R. Understanding Fossil Phytolith Preservation: The Role of Partial Dissolution in Paleoecology and Archaeology. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Cabanes, D.; Weiner, S.; Shahack-Gross, R. Stability of phytoliths in the archaeological record: A dissolution study of modern and fossil phytoliths. J. Archaeol. Sci. 2011, 38, 2480–2490. [Google Scholar] [CrossRef]
- Guntzer, F.; Keller, C.; Poulton, P.R.; McGrath, S.P.; Meunier, J.D. Long-term removal of wheat straw decreases soil amorphous silica at Broadbalk, Rothamsted. Plant Soil 2012, 352, 173–184. [Google Scholar] [CrossRef]
- Elbaum, R.; Melamed-Bessudo, C.; Tuross, N.; Levy, A.A.; Weiner, S. New methods to isolate organic materials from silicified phytoliths reveal fragmented glycoproteins but no DNA. Quat. Int. 2009, 193, 11–19. [Google Scholar] [CrossRef]
- Baines, S.B.; Twining, B.S.; Brzezinski, M.A.; Krause, J.W.; Vogt, S.; Assael, D.; McDaniel, H. Significant silicon accumulation by marine picocyanobacteria. Nat. Geosci. 2012, 5, 886. [Google Scholar] [CrossRef]
- Marron, A.O.; Alston, M.J.; Heavens, D.; Akam, M.; Caccamo, M.; Holland, P.W.; Walker, G. A family of diatom-like silicon transporters in the siliceous loricate choanoflagellates. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122543. [Google Scholar] [CrossRef] [Green Version]
- Trembath-Reichert, E.; Wilson, J.P.; McGlynn, S.E.; Fischer, W.W. Four hundred million years of silica biomineralization in land plants. Proc. Natl. Acad. Sci. USA 2015, 112, 5449–5454. [Google Scholar] [CrossRef] [Green Version]
- Katz, O. Beyond grasses: The potential benefits of studying silicon accumulation in non-grass species. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef]
- Fortin, M.G.; Morrison, N.A.; Verma, D.P.S. Nodulin-26, a peribacteroid membrane nodulin is expressed independently of the development of the peribacteroid compartment. Nucleic Acids Res. 1987, 15, 813–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zardoya, R.; Ding, X.; Kitagawa, Y.; Chrispeels, M.J. Origin of plant glycerol transporters by horizontal gene transfer and functional recruitment. Proc. Natl. Acad. Sci. USA 2002, 99, 14893–14896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Zhu, Z. Functional divergence of the NIP III subgroup proteins involved altered selective constraints and positive selection. BMC Plant Biol. 2010, 10, 256. [Google Scholar] [CrossRef] [PubMed]
- Strömberg, C.A.; Di Stilio, V.S.; Song, Z. Functions of phytoliths in vascular plants: An evolutionary perspective. Funct. Ecol. 2016, 30, 1286–1297. [Google Scholar] [CrossRef]
- Alhousari, F.; Greger, M. Silicon and Mechanisms of Plant Resistance to Insect Pests. Plants 2018, 7, 33. [Google Scholar] [CrossRef]
- Katz, O. Silica phytoliths in angiosperms: Phylogeny and early evolutionary history. New Phytol. 2015, 208, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Sun, W.; Zhu, Y.-G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Datnoff, L.E.; Snyder, G.H.; Korndörfer, G.H. Silicon in agriculture; Elsevier: Cambridge, MA, USA, 2001; Volume 8. [Google Scholar]
- Luyckx, M.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Silicon and plants: Current knowledge and technological perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Maillard, A.; Hajirezaei, M.R.; Ali, N.; Schwarzenberg, A.; Jamois, F.; Yvin, J.-C. Induction of Barley Silicon Transporter HvLsi1 and HvLsi2, increased silicon concentration in the shoot and regulated Starch and ABA Homeostasis under Osmotic stress and Concomitant Potassium Deficiency. Front. Plant Sci. 2017, 8, 1359. [Google Scholar] [CrossRef] [Green Version]
- Samuels, A.; Glass, A.; Ehret, D.; Menzies, J. The effects of silicon supplementation on cucumber fruit: Changes in surface characteristics. Ann. Bot. 1993, 72, 433–440. [Google Scholar] [CrossRef]
- De Souza, P.V.; Machado, B.R.; da Silva, D.C.; Menezes, I.P.; Ara&ujo, M.S.; de Jesus, F.A. Effect of resistance and trichome inducers on attraction of Euschistus heros (Hemiptera: Pentatomidae) to soybeans. Afr. J. Agric. Res. 2014, 9, 889–894. [Google Scholar]
- Agarie, S.; Hanaoka, N.; Ueno, O.; Miyazaki, A.; Kubota, F.; Agata, W.; Kaufman, P.B. Effects of silicon on tolerance to water deficit and heat stress in rice plants (Oryza sativa L.), monitored by electrolyte leakage. Plant Prod. Sci. 1998, 1, 96–103. [Google Scholar] [CrossRef]
- Yoshida, S.; Ohnishi, Y.; Kitagishi, K. Histochemistry of silicon in rice plant: III. The presence of cuticle-silica double layer in the epidermal tissue. Soil Sci. Plant Nutr. 1962, 8, 1–5. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon improves water use efficiency in maize plants. J. Plant Nutr. 2005, 27, 1457–1470. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon decreases transpiration rate and conductance from stomata of maize plants. J. Plant Nutr. 2006, 29, 1637–1647. [Google Scholar] [CrossRef]
- Ma, J.F. Silicon uptake and translocation in plants. In Proceedings of the International Plant Nutrition Colloquium XVI; Department of Plant Sciences: Davis, CA, USA, 2009. [Google Scholar]
- Abdel-Haliem, M.E.; Hegazy, H.S.; Hassan, N.S.; Naguib, D.M. Effect of silica ions and nano silica on rice plants under salinity stress. Ecol. Eng. 2017, 99, 282–289. [Google Scholar] [CrossRef]
- Gong, H.; Randall, D.; Flowers, T. Silicon deposition in the root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plantcell Environ. 2006, 29, 1970–1979. [Google Scholar] [CrossRef]
- Tuna, A.L.; Kaya, C.; Higgs, D.; Murillo-Amador, B.; Aydemir, S.; Girgin, A.R. Silicon improves salinity tolerance in wheat plants. Environ. Exp. Bot. 2008, 62, 10–16. [Google Scholar] [CrossRef]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef]
- Gong, H.; Blackmore, D.; Clingeleffer, P.; Sykes, S.; Jha, D.; Tester, M.; Walker, R. Contrast in chloride exclusion between two grapevine genotypes and its variation in their hybrid progeny. J. Exp. Bot. 2010, 62, 989–999. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Naranjo, E.; Andrades-Moreno, L.; Davy, A.J. Silicon alleviates deleterious effects of high salinity on the halophytic grass Spartina densiflora. Plant Physiol. Biochem. 2013, 63, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Daoud, A.M.; Hemada, M.M.; El-Araby, A.A. Effect of silicon on the tolerance of wheat (Triticum aestivum L.) to salt stress at different growth stages: Case study for the management of irrigation water. Plants 2018, 7, 29. [Google Scholar]
- Kim, Y.H.; Khan, A.L.; Kim, D.H.; Lee, S.Y.; Kim, K.M.; Waqas, M.; Jung, H.Y.; Shin, J.H.; Kim, J.G.; Lee, I.J. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Ding, Y.; Xie, Y.; Sangari, S. Silicon mechanisms to ameliorate heavy metal stress in plants. Biomed Res. Int. 2018, 2018, 8492898. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef] [PubMed]
- Pontigo, S.; Godoy, K.; Jimenez, H.; Gutierrez-Moraga, A.; Mora, M.D.; Cartes, P. Silicon-Mediated Alleviation of Aluminum Toxicity by Modulation of Al/Si Uptake and Antioxidant Performance in Ryegrass Plants. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Cai, H.M.; He, C.W.; Zhang, W.J.; Wang, L.J. A hemicellulose-bound form of silicon inhibits cadmium ion uptake in rice (Oryza sativa) cells. New Phytol. 2015, 206, 1063–1074. [Google Scholar] [CrossRef]
- Dubey, A.K.; Kumar, N.; Ranjan, R.; Gautam, A.; Pande, V.; Sanyal, I.; Mallick, S. Application of glycine reduces arsenic accumulation and toxicity in Oryza sativa L. by reducing the expression of silicon transporter genes. Ecotoxicol. Environ. Saf. 2018, 148, 410–417. [Google Scholar] [CrossRef]
- Khan, E.; Gupta, M. Arsenic-silicon priming of rice (Oryza sativa L.) seeds influence mineral nutrient uptake and biochemical responses through modulation of Lsi-1, Lsi-2, Lsi-6 and nutrient transporter genes. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Greger, M.; Landberg, T.; Vaculík, M. Silicon influences soil availability and accumulation of mineral nutrients in various plant species. Plants 2018, 7, 41. [Google Scholar] [CrossRef]
- Hu, A.Y.; Che, J.; Shao, J.F.; Yokosho, K.; Zhao, X.Q.; Shen, R.F.; Ma, J.F. Silicon accumulated in the shoots results in down-regulation of phosphorus transporter gene expression and decrease of phosphorus uptake in rice. Plant Soil 2018, 423, 317–325. [Google Scholar] [CrossRef]
- Kabir, A.H.; Hossain, M.M.; Khatun, M.A.; Mandal, A.; Haider, S.A. Role of Silicon Counteracting Cadmium Toxicity in Alfalfa (Medicago sativa L.). Front. Plant Sci. 2016, 7, 1117. [Google Scholar] [CrossRef] [PubMed]
- Akcay, U.C.; Erkan, I.E. Silicon Induced Antioxidative Responses and Expression of BOR2 and Two PIP Family Aquaporin Genes in Barley Grown Under Boron Toxicity. Plant Mol. Biol. Rep. 2016, 34, 318–326. [Google Scholar] [CrossRef]
- Blaich, R.; Grundhöfer, H. Silicate incrusts induced by powdery mildew in cell walls of different plant species/Echter Mehltau induziert Silikateinlagerungen in Zellwänden unterschiedlichster Pflanzenarten. Z. Pflanzenkrankh. Und Pflanzenschutz/J. Plant Dis. Prot. 1998, 105, 114–120. [Google Scholar]
- Kim, S.G.; Kim, K.W.; Park, E.W.; Choi, D. Silicon-induced cell wall fortification of rice leaves: A possible cellular mechanism of enhanced host resistance to blast. Phytopathology 2002, 92, 1095–1103. [Google Scholar] [CrossRef]
- Fauteux, F.; Rémus-Borel, W.; Menzies, J.G.; Bélanger, R.R. Silicon and plant disease resistance against pathogenic fungi. FEMS Microbiol. Lett. 2005, 249, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Samuels, A.; Glass, A.; Ehret, D.; Menzies, J. Mobility and deposition of silicon in cucumber plants. Plant Cell Environ. 1991, 14, 485–492. [Google Scholar] [CrossRef]
- Chérif, M.; Asselin, A.; Bélanger, R. Defense responses induced by soluble silicon in cucumber roots infected by Pythium spp. Phytopathology 1994, 84, 236–242. [Google Scholar] [CrossRef]
- Fawe, A.; Abou-Zaid, M.; Menzies, J.; Bélanger, R. Silicon-mediated accumulation of flavonoid phytoalexins in cucumber. Phytopathology 1998, 88, 396–401. [Google Scholar] [CrossRef]
- Nakata, Y.; Uena, M.; Kihara, J.; Ichii, M.; Taketa, S.; Arase, S. Rice blast disease and susceptibility to pests in a silicon uptake-deficient mutant lsil of rice. Crop Prot. 2008, 27, 865–868. [Google Scholar] [CrossRef]
- Dallagnol, L.J.; Rodrigues, F.A.; Mielli, M.V.B.; Ma, J.F. Rice grain resistance to brown spot and yield are increased by silicon. Trop. Plant Pathol. 2014, 39, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.; Hoeck, H.N.; Perez, L. Microwear of mammalian teeth as an indicator of diet. Science 1978, 201, 908–910. [Google Scholar] [CrossRef] [PubMed]
- Baker, G.; Jones, L.; Wardrop, I. Cause of wear in sheeps’ teeth. Nature 1959, 184, 1583. [Google Scholar] [CrossRef] [PubMed]
- Sanson, G.D.; Kerr, S.A.; Gross, K.A. Do silica phytoliths really wear mammalian teeth? J. Archaeol. Sci. 2007, 34, 526–531. [Google Scholar] [CrossRef]
- Lucas, P.W.; van Casteren, A.; Al-Fadhalah, K.; Almusallam, A.S.; Henry, A.G.; Michael, S.; Watzke, J.; Reed, D.A.; Diekwisch, T.G.H.; Strait, D.S.; et al. The role of dust, grit and phytoliths in tooth wear. Ann. Zool. Fenn. 2014, 51, 143–152. [Google Scholar] [CrossRef]
- Reynolds, J.J.; Lambin, X.; Massey, F.P.; Reidinger, S.; Sherratt, J.A.; Smith, M.J.; White, A.; Hartley, S.E. Delayed induced silica defences in grasses and their potential for destabilising herbivore population dynamics. Oecologia 2012, 170, 445–456. [Google Scholar] [CrossRef]
- Hartley, S.E. Round and round in cycles? Silicon-based plant defences and vole population dynamics. Funct. Ecol. 2015, 29, 151–153. [Google Scholar] [CrossRef]
- Wieczorek, M.; Zub, K.; Szafrańska, P.A.; Książek, A.; Konarzewski, M. Plant–herbivore interactions: Silicon concentration in tussock sedges and population dynamics of root voles. Funct. Ecol. 2015, 29, 187–194. [Google Scholar] [CrossRef]
- Calandra, I.; Zub, K.; Szafranska, P.A.; Zalewski, A.; Merceron, G. Silicon-based plant defences, tooth wear and voles. J. Exp. Biol. 2016, 219, 501–507. [Google Scholar] [CrossRef] [Green Version]
- MacFadden, B.J. Terrestrial mammalian herbivore response to declining levels of atmospheric CO2 during the cenozoic: Evidence from North American fossil horses (family Equidae). In A History of Atmospheric CO2 and Its Effects on Plants, Animals, and Ecosystems; Springer: Berlin, Germany, 2005; pp. 273–292. [Google Scholar]
- Elger, A.; Lemoine, D.G.; Fenner, M.; Hanley, M.E. Plant ontogeny and chemical defence: Older seedlings are better defended. Oikos 2009, 118, 767–773. [Google Scholar] [CrossRef]
- Massey, F.P.; Hartley, S.E. Experimental demonstration of the antiherbivore effects of silica in grasses: Impacts on foliage digestibility and vole growth rates. Proc. R. Soc. B Biol. Sci. 2006, 273, 2299–2304. [Google Scholar] [CrossRef] [PubMed]
- Bazely, D.R.; Vicari, M.; Emmerich, S.; Filip, L.; Lin, D.; Inman, A. Interactions between herbivores and endophyte-infected Festuca rubra from the Scottish islands of St. Kilda, Benbecula and Rum. J. Appl. Ecol. 1997, 847–860. [Google Scholar] [CrossRef]
- Huitu, O.; Forbes, K.M.; Helander, M.; Julkunen-Tiitto, R.; Lambin, X.; Saikkonen, K.; Stuart, P.; Sulkama, S.; Hartley, S. Silicon, endophytes and secondary metabolites as grass defenses against mammalian herbivores. Front. Plant Sci. 2014, 5, 478. [Google Scholar] [CrossRef] [PubMed]
- Ookawa, T.; Inoue, K.; Matsuoka, M.; Ebitani, T.; Takarada, T.; Yamamoto, T.; Ueda, T.; Yokoyama, T.; Sugiyama, C.; Nakaba, S. Increased lodging resistance in long-culm, low-lignin gh2 rice for improved feed and bioenergy production. Sci. Rep. 2014, 4, 6567. [Google Scholar] [CrossRef] [Green Version]
- Schoelynck, J.; Bal, K.; Backx, H.; Okruszko, T.; Meire, P.; Struyf, E. Silica uptake in aquatic and wetland macrophytes: A strategic choice between silica, lignin and cellulose? New Phytol. 2010, 186, 385–391. [Google Scholar] [CrossRef]
- Soundararajan, P.; Sivanesan, I.; Jana, S.; Jeong, B.R. Influence of silicon supplementation on the growth and tolerance to high temperature in Salvia splendens. Hortic. Environ. Biotechnol. 2014, 55, 271–279. [Google Scholar] [CrossRef]
- Vulavala, V.K.; Elbaum, R.; Yermiyahu, U.; Fogelman, E.; Kumar, A.; Ginzberg, I. Silicon fertilization of potato: Expression of putative transporters and tuber skin quality. Planta 2016, 243, 217–229. [Google Scholar] [CrossRef]
- Yamamoto, T.; Nakamura, A.; Iwai, H.; Ishii, T.; Ma, J.F.; Yokoyama, R.; Nishitani, K.; Satoh, S.; Furukawa, J. Effect of silicon deficiency on secondary cell wall synthesis in rice leaf. J. Plant Res. 2012, 125, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Motomura, H.; Hikosaka, K.; Suzuki, M. Relationships between photosynthetic activity and silica accumulation with ages of leaf in Sasa veitchii (Poaceae, Bambusoideae). Ann. Bot. 2007, 101, 463–468. [Google Scholar] [CrossRef]
- Dorairaj, D.; Ismail, M.R. Distribution of Silicified Microstructures, Regulation of Cinnamyl Alcohol Dehydrogenase and Lodging Resistance in Silicon and Paclobutrazol Mediated Oryza sativa. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef]
- Cao, H.S.; Zhuo, L.; Su, Y.; Sun, L.X.; Wang, X.M. Non-specific phospholipase C1 affects silicon distribution and mechanical strength in stem nodes of rice. Plant J. 2016, 86, 308–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehghanipoodeh, S.; Ghobadi, C.; Baninasab, B.; Gheysari, M.; Shiranibidabadi, S. Effect of Silicon on Growth and Development of Strawberry under Water Deficit Conditions. Hortic. Plant J. 2018, 4, 226–232. [Google Scholar] [CrossRef]
- Goto, M.; Ehara, H.; Karita, S.; Takabe, K.; Ogawa, N.; Yamada, Y.; Ogawa, S.; Yahaya, M.S.; Morita, O. Protective effect of silicon on phenolic biosynthesis and ultraviolet spectral stress in rice crop. Plant Sci. 2003, 164, 349–356. [Google Scholar] [CrossRef]
- Yao, X.; Chu, J.; Cai, K.; Liu, L.; Shi, J.; Geng, W. Silicon improves the tolerance of wheat seedlings to ultraviolet-B stress. Biol. Trace Elem. Res. 2011, 143, 507–517. [Google Scholar] [CrossRef] [PubMed]
- KAUFMAN, P.B.; Dayanandan, P.; Franklin, C.; Takeoka, Y. Structure and function of silica bodies in the epidermal system of grass shoots. Ann. Bot. 1985, 55, 487–507. [Google Scholar] [CrossRef]
- Kostic, L.; Nikolic, N.; Bosnic, D.; Samardzic, J.; Nikolic, M. Silicon increases phosphorus (P) uptake by wheat under low P acid soil conditions. Plant Soil 2017, 419, 447–455. [Google Scholar] [CrossRef]
- Haddad, C.; Arkoun, M.; Jamois, F.; Schwarzenberg, A.; Yvin, J.C.; Etienne, P.; Laine, P. Silicon Promotes Growth of Brassica napus L. and Delays Leaf Senescence Induced by Nitrogen Starvation. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Markovich, O.; Steiner, E.; Kouřil, Š.; Tarkowski, P.; Aharoni, A.; Elbaum, R. Silicon promotes cytokinin biosynthesis and delays senescence in Arabidopsis and Sorghum. Plantcell Environ. 2017, 40, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Maillard, A.; Ali, N.; Schwarzenberg, A.; Jamois, F.; Yvin, J.C.; Hosseini, S.A. Silicon transcriptionally regulates sulfur and ABA metabolism and delays leaf senescence in barley under combined sulfur deficiency and osmotic stress. Environ. Exp. Bot. 2018, 155, 394–410. [Google Scholar] [CrossRef]
- Nikolic, D.B.; Nesic, S.; Bosnic, D.; Kostic, L.; Nikolic, M.; Samardzic, J.T. Silicon Alleviates Iron Deficiency in Barley by Enhancing Expression of Strategy II Genes and Metal Redistribution. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Cooke, J.; Leishman, M.R. Silicon concentration and leaf longevity: Is silicon a player in the leaf dry mass spectrum? Funct. Ecol. 2011, 25, 1181–1188. [Google Scholar] [CrossRef]
- Neumann, K.; Chevalier, A.; Vrydaghs, L. Phytoliths in Archaeology: Recent Advances; Springer: Berlin, Germany, 2017. [Google Scholar]
- Piperno, D.R. Phytolith analysis and tropical paleo-ecology: Production and taxonomic significance of siliceous forms in New World plant domesticates and wild species. Rev. Palaeobot. Palynol. 1985, 45, 185–228. [Google Scholar] [CrossRef]
- Kealhofer, L.; Piperno, D.R. Opal phytoliths in Southeast Asian flora; Smithsonian Institution Press: Washington, DC, USA, 1998. [Google Scholar]
- Bowdery, D. An enigma revisited: Identification of palm phytoliths extracted from the 1983 Rapa Nui, Rano Kao2 core. Veg. Hist. Archaeobotany 2015, 24, 455–466. [Google Scholar] [CrossRef]
- Morcote-Ríos, G.; Bernal, R.; Raz, L. Phytoliths as a tool for archaeobotanical, palaeobotanical and palaeoecological studies in Amazonian palms. Bot. J. Linn. Soc. 2016, 182, 348–360. [Google Scholar] [CrossRef] [Green Version]
- Patterer, N.I. Análisis fitolíticos de las principales especies de palmeras (Arecaceae) presentes en regiones subtropicales de América del Sur. Boletín De La Soc. Argent. De Botánica 2014, 49, 491–502. [Google Scholar]
- Jattisha, P.; Sabu, M. Phytoliths as a tool for the identification of some chloridoideae grasses in Kerala. Isrn Bot. 2012, 2012, 246057. [Google Scholar] [CrossRef]
- Ball, T.B.; Gardner, J.S.; Anderson, N. Identifying inflorescence phytoliths from selected species of wheat (Triticum monococcum, T. dicoccon, T. dicoccoides, and T. aestivum) and barley (Hordeum vulgare and H. spontaneum)(Gramineae). Am. J. Bot. 1999, 86, 1615–1623. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, J.; Wu, N.; Liu, K.-b.; Xu, D.; Li, Q. Phytoliths analysis for the discrimination of foxtail millet (Setaria italica) and common millet (Panicum miliaceum). PLoS ONE 2009, 4, e4448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lu, H.; Wu, N.; Yang, X.; Diao, X. Phytolith analysis for differentiating between foxtail millet (Setaria italica) and green foxtail (Setaria viridis). PLoS ONE 2011, 6, e19726. [Google Scholar] [CrossRef]
- Hart, T.C. Phytoliths: The storytelling stones inside plants. Am. Sci. 2015, 103, 136. [Google Scholar] [CrossRef]
- Vaughan, D.; Balazs, E.; Heslop-Harrison, J. From crop domestication to super-domestication. Ann. Bot. 2007, 100, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Ball, T.; Chandler-Ezell, K.; Dickau, R.; Duncan, N.; Hart, T.C.; Iriarte, J.; Lentfer, C.; Logan, A.; Lu, H.; Madella, M. Phytoliths as a tool for investigations of agricultural origins and dispersals around the world. J. Archaeol. Sci. 2016, 68, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Weisskopf, A.; Harvey, E.; Kingwell-Banham, E.; Kajale, M.; Mohanty, R.; Fuller, D.Q. Archaeobotanical implications of phytolith assemblages from cultivated rice systems, wild rice stands and macro-regional patterns. J. Archaeol. Sci. 2014, 51, 43–53. [Google Scholar] [CrossRef]
- Huan, X.; Lu, H.; Zhang, J.; Wang, C. Phytolith assemblage analysis for the identification of rice paddy. Sci. Rep. 2018, 8, 10932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radomski, K.U.; Neumann, K. Grasses and Grinding Stones: Inflorrescence Phytoliths from tvioclern West African Poaceae and Archaeological Stone artefacts. Windows Afr. Past Curr. Approaches Afr. Archaeobot. 2011, 3, 153. [Google Scholar]
- Dal Corso, M.; Nicosia, C.; Balista, C.; Cupitò, M.; Dalla Longa, E.; Leonardi, G.; Kirleis, W. Bronze Age crop processing evidence in the phytolith assemblages from the ditch and fen around Fondo Paviani, northern Italy. Veg. Hist. Archaeobot. 2017, 26, 5–24. [Google Scholar] [CrossRef]
- Dal Corso, M.; Out, W.A.; Ohlrau, R.; Hofmann, R.; Dreibrodt, S.; Videiko, M.; Müller, J.; Kirleis, W. Where are the cereals? Contribution of phytolith analysis to the study of subsistence economy at the Trypillia site Maidanetske (ca. 3900-3650 BCE), central Ukraine. J. Arid Environ. 2018, 157, 137–148. [Google Scholar] [CrossRef]
- Zuo, X.; Lu, H.; Jiang, L.; Zhang, J.; Yang, X.; Huan, X.; He, K.; Wang, C.; Wu, N. Dating rice remains through phytolith carbon-14 study reveals domestication at the beginning of the Holocene. Proc. Natl. Acad. Sci. USA 2017, 114, 6486–6491. [Google Scholar] [CrossRef] [Green Version]
- Tsartsidou, G.; Lev-Yadun, S.; Efstratiou, N.; Weiner, S. Use of space in a Neolithic village in Greece (Makri): Phytolith analysis and comparison of phytolith assemblages from an ethnographic setting in the same area. J. Archaeol. Sci. 2009, 36, 2342–2352. [Google Scholar] [CrossRef]
- Lancelotti, C.; Balbo, A.L.; Madella, M.; Iriarte, E.; Rojo-Guerra, M.; Royo, J.I.; Tejedor, C.; Garrido, R.; García, I.; Arcusa, H. The missing crop: Investigating the use of grasses at Els Trocs, a Neolithic cave site in the Pyrenees (1564 m asl). J. Archaeol. Sci. 2014, 42, 456–466. [Google Scholar] [CrossRef]
- Rosen, A.M.; Weiner, S. Identifying ancient irrigation: A new method using opaline phytoliths from emmer wheat. J. Archaeol. Sci. 1994, 21, 125–132. [Google Scholar] [CrossRef]
- Madella, M.; Jones, M.; Echlin, P.; Powers-Jones, A.; Moore, M. Plant water availability and analytical microscopy of phytoliths: Implications for ancient irrigation in arid zones. Quat. Int. 2009, 193, 32–40. [Google Scholar] [CrossRef]
- Jenkins, E.; Jamjoum, K.; Nuimat, S.; Stafford, R.; Nortcliff, S.; Mithen, S. Identifying ancient water availability through phytolith analysis: An experimental approach. J. Archaeol. Sci. 2016, 73, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Castillo, C.; Fuller, D. Bananas: The Spread of a Tropical Forest Fruit as an Agricultural Staple; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Wallis, L.A. Environmental history of northwest Australia based on phytolith analysis at Carpenter’s Gap 1. Quat. Int. 2001, 83, 103–117. [Google Scholar] [CrossRef]
- Ollendorf, A.L. Archaeological implications of a phytolith study at Tel Miqne (Ekron), Israel. J. Field Archaeol. 1987, 14, 453–463. [Google Scholar]
- Esteban, I.; Marean, C.W.; Fisher, E.C.; Karkanas, P.; Cabanes, D.; Albert, R.M. Phytoliths as an indicator of early modern humans plant gathering strategies, fire fuel and site occupation intensity during the Middle Stone Age at Pinnacle Point 5-6 (south coast, South Africa). PLoS ONE 2018, 13, e0198558. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.K.; Thompson, J.C.; Schilt, F.; Cohen, A.S.; Choi, J.-H.; Mercader, J.; Nightingale, S.; Miller, C.E.; Mentzer, S.M.; Walde, D. Approaches to Middle Stone Age landscape archaeology in tropical Africa. J. Archaeol. Sci. 2017, 77, 64–77. [Google Scholar] [CrossRef] [Green Version]
- Tsartsidou, G.; Karkanas, P.; Marshall, G.; Kyparissi-Apostolika, N. Palaeoenvironmental reconstruction and flora exploitation at the Palaeolithic cave of Theopetra, central Greece: The evidence from phytolith analysis. Archaeol. Anthropol. Sci. 2015, 7, 169–185. [Google Scholar] [CrossRef]
- Fearn, M.L. Phytoliths in sediment as indicators of grass pollen source. Rev. Palaeobot. Palynol. 1998, 103, 75–81. [Google Scholar] [CrossRef]
- Watling, J.; Iriarte, J. Phytoliths from the coastal savannas of French Guiana. Quat. Int. 2013, 287, 162–180. [Google Scholar] [CrossRef]
- Chendev, Y.; Khokhlova, O.; Ponomarenko, E.; Ershova, E.; Alexandrovskiy, A.; Myakshina, T. Holocene Environmental and Anthropogenic Changes of Soils and Vegetation in the Central Russian Upland: The Case Study in the “Belogorie” Natural Reserve. Geosciences 2018, 8, 473. [Google Scholar] [CrossRef]
- Gavrilov, D.; Loiko, S.; Klimova, N. Holocene Soil Evolution in South Siberia Based on Phytolith Records and Genetic Soil Analysis (Russia). Geosciences 2018, 8, 402. [Google Scholar] [CrossRef]
- Albert, R.M.; Bamford, M.K.; Esteban, I. Reconstruction of ancient palm vegetation landscapes using a phytolith approach. Quat. Int. 2015, 369, 51–66. [Google Scholar] [CrossRef]
- Strömberg, C.A. Using phytolith assemblages to reconstruct the origin and spread of grass-dominated habitats in the great plains of North America during the late Eocene to early Miocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 207, 239–275. [Google Scholar] [CrossRef]
- Barboni, D.; Ashley, G.M.; Dominguez-Rodrigo, M.; Bunn, H.T.; Mabulla, A.Z.; Baquedano, E. Phytoliths infer locally dense and heterogeneous paleovegetation at FLK North and surrounding localities during upper Bed I time, Olduvai Gorge, Tanzania. Quat. Res. 2010, 74, 344–354. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Cerling, T.E.; Helliker, B.R. C 4 photosynthesis, atmospheric CO2, and climate. Oecologia 1997, 112, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Fredlund, G.G.; Tieszen, L.T. Modern phytolith assemblages from the North American great plains. J. Biogeogr. 1994, 21, 321–335. [Google Scholar] [CrossRef]
- Baker, R.G.; Fredlund, G.G.; Mandel, R.D.; Bettis III, E.A. Holocene environments of the central Great Plains: Multi-proxy evidence from alluvial sequences, southeastern Nebraska. Quat. Int. 2000, 67, 75–88. [Google Scholar] [CrossRef]
- Blinnikov, M.; Busacca, A.; Whitlock, C. Reconstruction of the late Pleistocene grassland of the Columbia basin, Washington, USA, based on phytolith records in loess. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 177, 77–101. [Google Scholar] [CrossRef]
- Scott, L. Grassland development under glacial and interglacial conditions in southern Africa: Review of pollen, phytolith and isotope evidence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 177, 47–57. [Google Scholar] [CrossRef]
- Strömberg, C.A. The origin and spread of grass-dominated ecosystems in the late Tertiary of North America: Preliminary results concerning the evolution of hypsodonty. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 177, 59–75. [Google Scholar] [CrossRef]
- Abrantes, F. A 340,000 year continental climate record from tropical Africa–news from opal phytoliths from the equatorial Atlantic. Earth Planet. Sci. Lett. 2003, 209, 165–179. [Google Scholar] [CrossRef]
- Alexandré, A.; Meunier, J.-D.; Lézine, A.-M.; Vincens, A.; Schwartz, D. Phytoliths: Indicators of grassland dynamics during the late Holocene in intertropical Africa. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 136, 213–229. [Google Scholar] [CrossRef]
- Jiang, Q.; Piperno, D.R. Environmental and archaeological implications of a Late Quaternary palynological sequence, Poyang Lake, Southern China. Quat. Res. 1999, 52, 250–258. [Google Scholar] [CrossRef]
- McInerney, F.A.; Strömberg, C.A.; White, J.W. The Neogene transition from C 3 to C 4 grasslands in North America: Stable carbon isotope ratios of fossil phytoliths. Paleobiology 2011, 37, 23–49. [Google Scholar] [CrossRef]
- Strömberg, C.A.; McInerney, F.A. The Neogene transition from C 3 to C 4 grasslands in North America: Assemblage analysis of fossil phytoliths. Paleobiology 2011, 37, 50–71. [Google Scholar] [CrossRef]
- Rashid, I.; Mir, S.H.; Zurro, D.; Dar, R.A.; Reshi, Z.A. Phytoliths as proxies of the past. Earth-Sci. Rev. 2019, 194, 234–250. [Google Scholar] [CrossRef]
- Cabanes, D.; Gadot, Y.; Cabanes, M.; Finkelstein, I.; Weiner, S.; Shahack-Gross, R. Human impact around settlement sites: A phytolith and mineralogical study for assessing site boundaries, phytolith preservation, and implications for spatial reconstructions using plant remains. J. Archaeol. Sci. 2012, 39, 2697–2705. [Google Scholar] [CrossRef]
- Calegari, M.R.; Madella, M.; Vidal-Torrado, P.; Pessenda, L.C.R.; Marques, F.A. Combining phytoliths and delta C-13 matter in Holocene palaeoenvironmental studies of tropical soils: An example of an Oxisol in Brazil. Quat. Int. 2013, 287, 47–55. [Google Scholar] [CrossRef]
- Webb, E.A.; Longstaffe, F.J. Limitations on the climatic and ecological signals provided by the delta C-13 values of phytoliths from a C-4 North American prairie grass. Geochim. Et Cosmochim. Acta 2010, 74, 3041–3050. [Google Scholar] [CrossRef]
- Corbineau, R.; Reyerson, P.E.; Alexandre, A.; Santos, G.M. Towards producing pure phytolith concentrates from plants that are suitable for carbon isotopic analysis. Rev. Palaeobot. Palynol. 2013, 197, 179–185. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Silicon Transporter (Acc. No./Gene ID) | Tissue Localization | Sub-Cellular Localization | Expression Pattern (Si treatment effect) | Spatial Expression | Reference(s) |
|---|---|---|---|---|---|---|
| Channel Type Si-Transporters | ||||||
| Orzya sativa | OsLsi1NSCR (Os02g0745100) | Root (Exodermis and endodermis) | Plasma membrane | Downregulated | Mature roots | [26] |
| Hordium vulgare | HvLsi1 (100301576) | Root (Epidermal, hypodermal and cortex cells) | Plasma membrane | Unaffected | Mature roots | [30] |
| Zea mays L. | ZmLsi1 (542643) | Roots (Epidermal, hypodermal and cortex cells) Polar Localization at distal size Kernel (pericarp, embryo, endosperm) | Plasma membrane | Unaffected | Immature and Mature roots Immature and Mature Kernels | [14,46] |
| Triticum aestivum | TaLsi1 (ADM47602) | All tissues of roots and shoots | Plasma membrane | Unaffected | Central Cylinder within the root differentiation zone | [45] |
| Cucumus sativus | CsiT-1 (Csa017389)CR CsiT-2 (Csa017390)CR | Roots Leaves Stem Flower | Plasma membrane | Upregulated | Both mature and young leaves | [44] |
| Cucurbita moschata Duch. | CmLsi1(B+ ) (LOC111441250) | Roots (all root cells) | Plasma membrane | Upregulated | -- | [36] |
| CmLsi1(B-) | Shoot | Endoplasmic reticulum | ||||
| O. sativa | OsLsi6 (Os06g0228200) | Root (All root cells) Shoot | Plasma membrane | Downregulated | Immature Root Tip | [34] |
| H. vulgare | HvLsi6 (AB447484) | Root (Epidermis and cortex cells) Shoot Leaf blades and sheaths (Parenchyma cells and vascular bundle) | Plasma membrane | Upregulated | Root Tip (Vegetative growth stage) Nodes (Reproductive growth stage) | [37] |
| Z. mays L | ZmLsi6 (AMQ98973) | Root (All root cells) Shoot (Xylem parenchyma cells) No polar localizationLeaf sheaths and blades (PL) | Plasma membrane | Unaffected | [14] | |
| Equistium arvense L. | EaLsi1 (NIP3;1) | Root | Plasma membrane | [95] | ||
| Sorghum bicolor | SbLsi1 | Leaf blade (epidermal cells) | Plasma membrane | [74] | ||
| Cannabis sativa | CsaNIP2-1s PKNIP2-1 (PK09456.1) FNNIP2-1 (FN14156.1) | Aerial organs (flowers and shoot) | Plasma membrane | [57] | ||
| Efflux Silicon transporters | ||||||
| O. sativa | OsLsi2 (Os03g0107300) | Root Node (Bundle Sheeth Cells) PL | Plasma membrane | Upregulated | [28] | |
| H. vulgare | HvLsi2 (100502546) | Root Node (Parenchyma Cells) | Plasma membrane | [37] | ||
| Z. mays L. | ZmLsi2 (541884) | Root | Plasma membrane | [14] | ||
| C. moschata | CmLsi2-1 (AB551951) CmLsi2-2 (AB551952) | Root Shoot | Plasma membrane | Young roots | [35] | |
| O. sativa | OsLsi3 (Os10g0547500) | Node (Parenchyma tissue) | Plasma membrane | Upregulated | [77] | |
| Chrysanthemum (Chrysanthemum X morifolium) | CmLsi2 | [98] | ||||
| E. arvense | EaLsi2-1 | RootShoot | Plasma membrane | [99] | ||
| Glycine max | GmNIP2-1 (Glyma09G238200) | Root | Plasma membrane | Downregulated | -- | [33] |
| G. max | GmNIP2-2 (Glyma18G259500) | Shoot | Plasma membrane | Downregulated | ||
| C. sativus | CsaLsi21 (PK18630.1/FN08935.1) CsaLsi2-2 (PK08860.1/FN07382.1) CsaLsi2-3 (PK00413.1/FN13891.1) | Variable expression across plant tissues | Plasma membrane | [57] | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawaz, M.A.; Zakharenko, A.M.; Zemchenko, I.V.; Haider, M.S.; Ali, M.A.; Imtiaz, M.; Chung, G.; Tsatsakis, A.; Sun, S.; Golokhvast, K.S. Phytolith Formation in Plants: From Soil to Cell. Plants 2019, 8, 249. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8080249
Nawaz MA, Zakharenko AM, Zemchenko IV, Haider MS, Ali MA, Imtiaz M, Chung G, Tsatsakis A, Sun S, Golokhvast KS. Phytolith Formation in Plants: From Soil to Cell. Plants. 2019; 8(8):249. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8080249
Chicago/Turabian StyleNawaz, Muhammad Amjad, Alexander Mikhailovich Zakharenko, Ivan Vladimirovich Zemchenko, Muhammad Sajjad Haider, Muhammad Amjad Ali, Muhammad Imtiaz, Gyuhwa Chung, Aristides Tsatsakis, Sangmi Sun, and Kirill Sergeyevich Golokhvast. 2019. "Phytolith Formation in Plants: From Soil to Cell" Plants 8, no. 8: 249. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8080249