Assessing the Correlations between Different Traits in Copper-Sensitive and Copper-Resistant Varieties of Jute (Corchorus capsularis L.)


 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions and Experimental Treatment
2.2. Growth and Morphological Traits
2.3. Determination of Oxidative Stress
2.4. Analysis of Antioxidant Enzyme Activities
2.5. Determination of Cu Concentration
2.6. Statistical Analysis
3. Results
3.1. Effects of Cu Toxicity on Seed Germination, Growth, and Chlorophyll Content
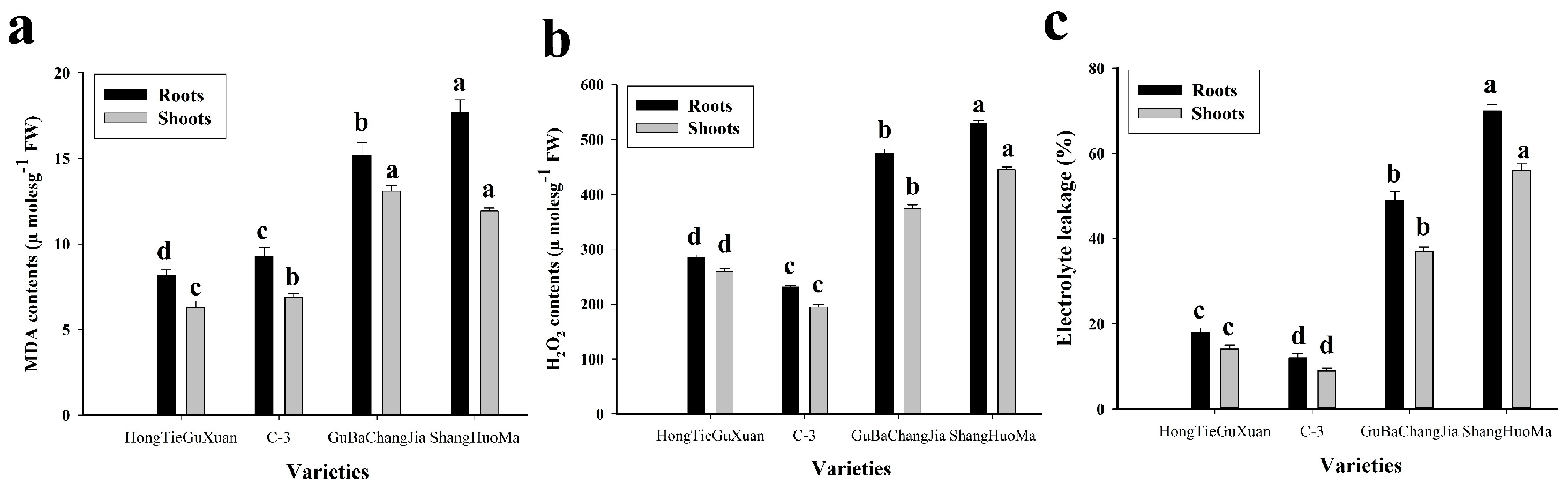
3.2. Effect of Cu Toxicity on Oxidative Stress
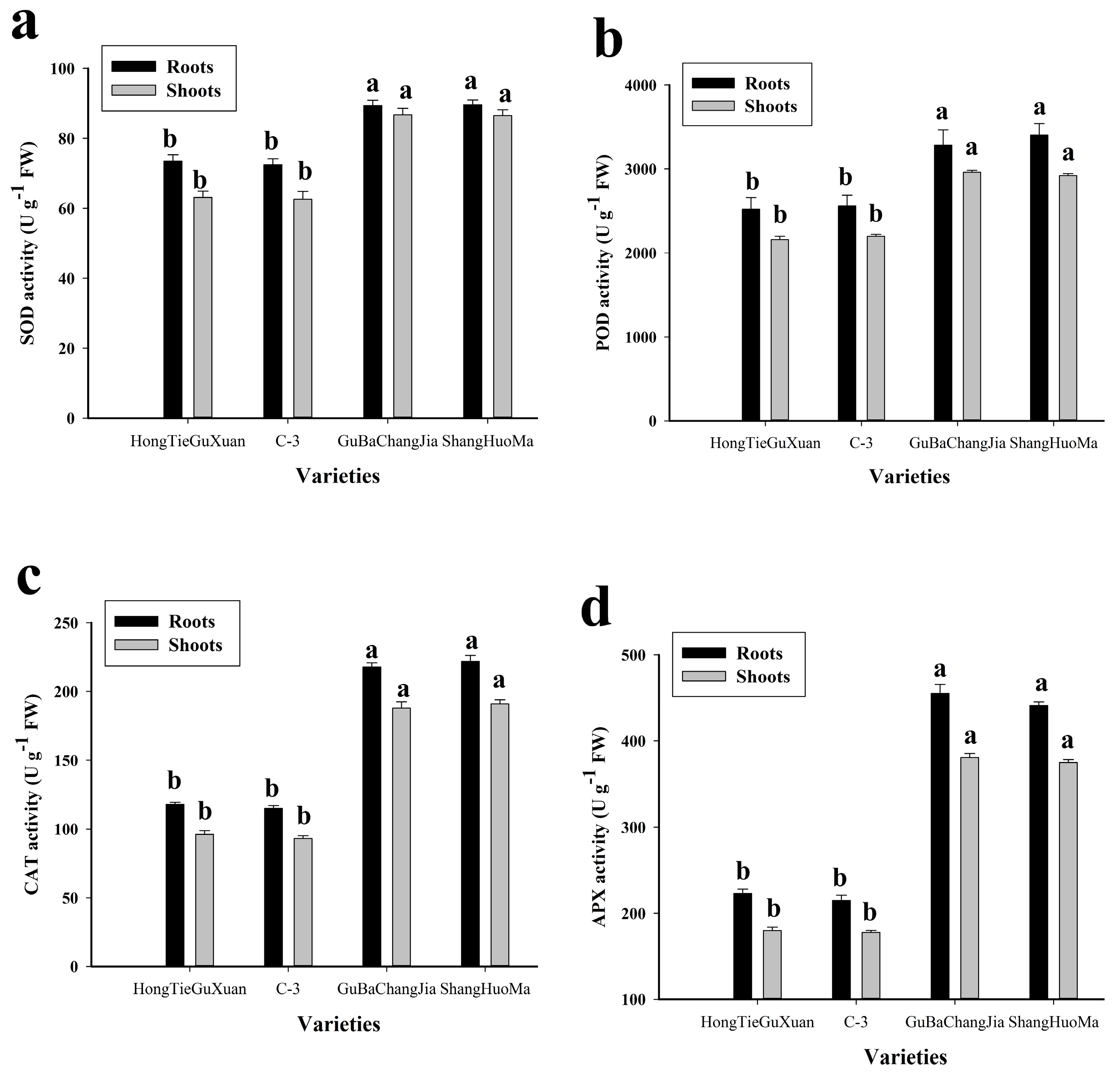
3.3. Effect of Cu Toxicity on Antioxidant Activities
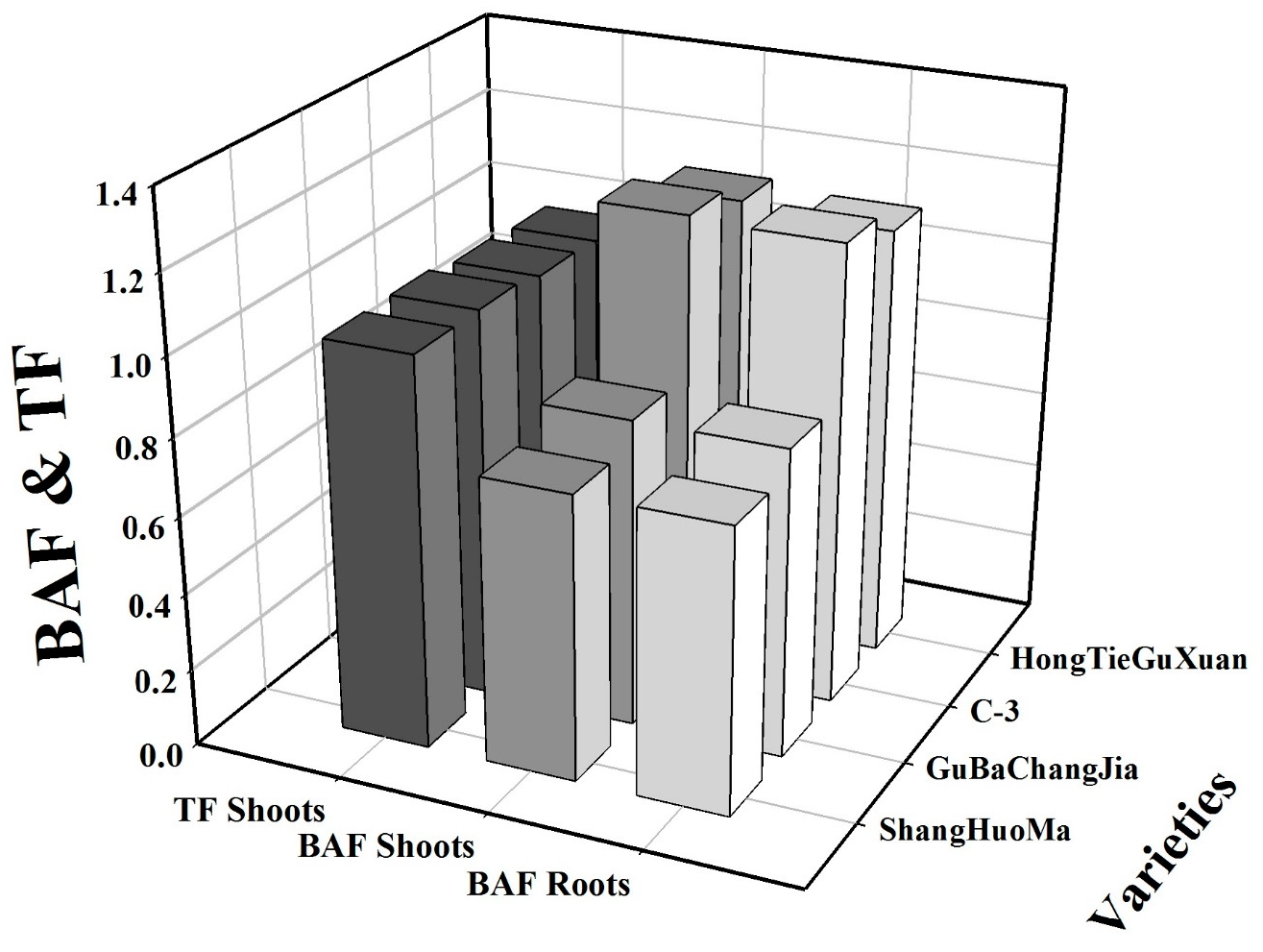
3.4. Cu Uptake and Bioaccumulation in the Roots and Shoots
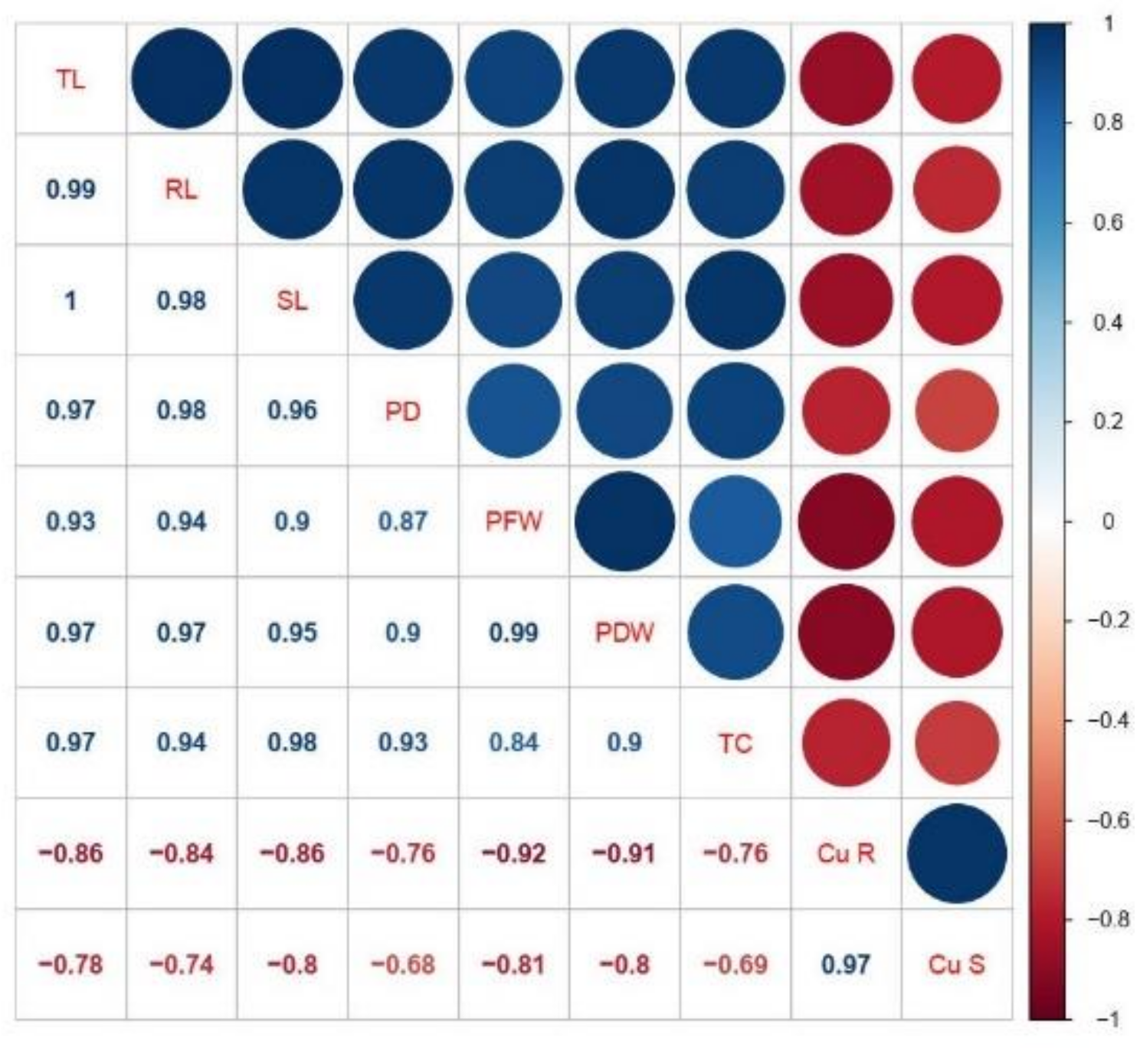
3.5. Correlation between Growth, Biomass, Total Chlorophyll Content, and Cu Uptake
4. Discussions
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vardhan, K.H.; Kumar, P.S.; Panda, R.C. A review on heavy metal pollution, toxicity and remedial measures: Current trends and future perspectives. J. Mol. Liq. 2019, 290, 111197. [Google Scholar] [CrossRef]
- Daud, M.K.; Ali, S.; Abbas, Z.; Zaheer, I.E.; Riaz, M.A.; Malik, A.; Hussain, A.; Rizwan, M.; Zia-ur-Rehman, M.; Zhu, S.J. Potential of duckweed (Lemna minor) for the Phytoremediation of Landfill Leachate. J. Chem. 2018, 2018. [Google Scholar] [CrossRef]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Let. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Rehman, M.; Liu, L.; Wang, Q.; Saleem, M.H.; Bashir, S.; Ullah, S.; Peng, D. Copper environmental toxicology, recent advances, and future outlook: A review. Environ. Sci. Pol. Res. 2019, 26, 18003–18016. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, K.; Gill, R.A.; Islam, F.; Farooq, M.A.; Wang, J.; Zhou, W. Ecotoxicological and Interactive Effects of Copper and Chromium on Physiochemical, Ultrastructural, and Molecular Profiling in Brassica napus L. BioMed Res. Inter. 2018, 2018. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Zia-ur-Rehman, M.; Irshad, M.K.; Bharwana, S.A. The effect of excess copper on growth and physiology of important food crops: A review. Environ. Sci. Pol. Res. 2015, 22, 8148–8162. [Google Scholar] [CrossRef]
- Rehman, M.; Maqbool, Z.; Peng, D.; Liu, L. Morpho-physiological traits, antioxidant capacity and phytoextraction of copper by ramie (Boehmeria nivea L.) grown as fodder in copper-contaminated soil. Environ. Sci. Pol. Res. 2019, 26, 5851–5861. [Google Scholar] [CrossRef]
- Chen, J.; Shafi, M.; Li, S.; Wang, Y.; Wu, J.; Ye, Z.; Peng, D.; Yan, W.; Liu, D. Copper induced oxidative stresses, antioxidant responses and phytoremediation potential of Moso bamboo (Phyllostachys pubescens). Sci. Rep. 2015, 5, 13554. [Google Scholar] [CrossRef]
- Liu, J.; Wang, J.; Lee, S.; Wen, R. Copper-caused oxidative stress triggers the activation of antioxidant enzymes via ZmMPK3 in maize leaves. PLoS ONE 2018, 13, e0203612. [Google Scholar] [CrossRef]
- Ahsan, N.; Lee, D.-G.; Lee, S.-H.; Kang, K.Y.; Lee, J.J.; Kim, P.J.; Yoon, H.-S.; Kim, J.-S.; Lee, B.-H. Excess copper induced physiological and proteomic changes in germinating rice seeds. Chemosphere 2007, 67, 1182–1193. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.; Liu, L.; Bashir, S.; Saleem, M.H.; Chen, C.; Peng, D.; Siddique, K.H. Influence of rice straw biochar on growth, antioxidant capacity and copper uptake in ramie (Boehmeria nivea L.) grown as forage in aged copper-contaminated soil. Plant Phy. Biochem. 2019, 138, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Liu, Y.; Li, X.; Feng, Z.; Xu, Z.; Wu, H.; Xu, J. Expected CO2-induced ocean acidification modulates copper toxicity in the green tide alga Ulva prolifera. Environ. Exp. Bot. 2017, 135, 63–72. [Google Scholar] [CrossRef]
- Habiba, U.; Ali, S.; Farid, M.; Shakoor, M.B.; Rizwan, M.; Ibrahim, M.; Abbasi, G.H.; Hayat, T.; Ali, B. EDTA enhanced plant growth, antioxidant defense system, and phytoextraction of copper by Brassica napus L. Environ. Sci. Pol. Res. 2015, 22, 1534–1544. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, I.E.; Ali, S.; Rizwan, M.; Farid, M.; Shakoor, M.B.; Gill, R.A.; Najeeb, U.; Iqbal, N.; Ahmad, R. Citric acid assisted phytoremediation of copper by Brassica napus L. Ecotox. Environ. Safe. 2015, 120, 310–317. [Google Scholar] [CrossRef]
- Nair, P.M.G.; Chung, I.M. Study on the correlation between copper oxide nanoparticles induced growth suppression and enhanced lignification in Indian mustard (Brassica juncea L.). Ecotox. Environ. Safe. 2015, 113, 302–313. [Google Scholar] [CrossRef]
- Ashagre, H.; Shelema, M.; Kedir, R.; Ebsa, S. Seed germination and seedling growth of haricot bean (Phaseolus vulgaris L.) cultivars as influenced by copper sulphate. World J. Agric. Sci. 2013, 1, 312–317. [Google Scholar]
- Farooq, M.A.; Li, L.; Ali, B.; Gill, R.A.; Wang, J.; Ali, S.; Gill, M.B.; Zhou, W. Oxidative injury and antioxidant enzymes regulation in arsenic-exposed seedlings of four Brassica napus L. cultivars. Environ. Sci. Pol. Res. 2015, 22, 10699–10712. [Google Scholar] [CrossRef]
- Quartacci, M.; Ranieri, A.; Sgherri, C. Antioxidative defence mechanisms in two grapevine (Vitis vinifera L.) cultivars grown under boron excess in the irrigation water. VITIS-J. Grape. Res. 2015, 54, 51–58. [Google Scholar]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Sun, B.-Y.; Kan, S.-H.; Zhang, Y.-Z.; Deng, S.-H.; Wu, J.; Yuan, H.; Qi, H.; Yang, G.; Li, L.; Zhang, X.-H. Certain antioxidant enzymes and lipid peroxidation of radish (Raphanus sativus L.) as early warning biomarkers of soil copper exposure. J. Hazard. Mater. 2010, 183, 833–838. [Google Scholar] [CrossRef]
- Ashraf, S.; Ali, Q.; Zahir, Z.A.; Ashraf, S.; Asghar, H.N. Phytoremediation: Environmentally sustainable way for reclamation of heavy metal polluted soils. Ecotox. Environ. Saf. 2019, 174, 714–727. [Google Scholar] [CrossRef] [PubMed]
- Lajayer, B.A.; Moghadam, N.K.; Maghsoodi, M.R.; Ghorbanpour, M.; Kariman, K. Phytoextraction of heavy metals from contaminated soil, water and atmosphere using ornamental plants: Mechanisms and efficiency improvement strategies. Environ. Sci. Pollut. Res. 2019, 26, 8468–8484. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Qi, J.; Zhang, G.; Lin, L.; Fang, P.; Tao, A.; Xu, J. Effect of cadmium stress on the growth, antioxidative enzymes and lipid peroxidation in two kenaf (Hibiscus cannabinus L.) plant seedlings. J. Integr. Agric. 2013, 12, 610–620. [Google Scholar] [CrossRef]
- Hosman, M.E.; El-Feky, S.S.; Mohamed, M.; Shaker, E.M. Mechanism of phytoremediation potential of flax (Linum usitatissimum L.) to Pb, Cd and Zn. Asian J. Plant Sci. Res. 2017, 7, 30–40. [Google Scholar]
- Ahmed, D.A.; Slima, D.F. Heavy metal accumulation by Corchorus olitorius L. irrigated with wastewater. Environ. Sci. Pollut. Res. 2018, 25, 14996–15005. [Google Scholar] [CrossRef]
- Ogoko, E. Accumulation of heavy metal in soil and their transfer to leafy vegetables with phytoremediation potential. Am. J. Chem. 2015, 5, 125–131. [Google Scholar]
- Abubakari, M.; Moomin, A.; Nyarko, G.; Dawuda, M. Heavy metals concentrations and risk assessment of roselle and jute mallow cultivated with three compost types. Ann. Agric. Sci. 2017, 62, 145–150. [Google Scholar] [CrossRef]
- Faruk, O.; Bledzki, A.K.; Fink, H.-P.; Sain, M. Biocomposites reinforced with natural fibers: 2000–2010. Prog. Polym. Sci. 2012, 37, 1552–1596. [Google Scholar] [CrossRef]
- Nath, K.; Saini, S.; Sharma, Y.K. Chromium in tannery industry effluent and its effect on plant metabolism and growth. J. Environ. Biol. 2005, 26, 197–204. [Google Scholar]
- Young, H.H.; Johnson, F.P. Practice of Urology: Based on a Study of 12,500 Cases; WB Saunders: Philadelphia, PA, USA, 1926. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Chen, C.-N.; Pan, S.-M. Assay of superoxide dismutase activity by combining electrophoresis and densitometry. Bot. Bull. Acad. Sin. 1996, 37, 107–111. [Google Scholar]
- Sakharov, I.Y.; Ardila, G.B. Variations of peroxidase activity in cocoa (Theobroma cacao L.) beans during their ripening, fermentation and drying. Food Chem. 1999, 65, 51–54. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; pp. 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Celis-Plá, P.S.; Brown, M.T.; Santillán-Sarmiento, A.; Korbee, N.; Sáez, C.A.; Figueroa, F.L. Ecophysiological and metabolic responses to interactive exposure to nutrients and copper excess in the brown macroalga Cystoseira tamariscifolia. Mar. Pollut. Bull. 2018, 128, 214–222. [Google Scholar] [CrossRef]
- Thounaojam, T.C.; Panda, P.; Mazumdar, P.; Kumar, D.; Sharma, G.; Sahoo, L.; Sanjib, P. Excess copper induced oxidative stress and response of antioxidants in rice. Plant Physiol. Biochem. 2012, 53, 33–39. [Google Scholar] [CrossRef]
- Kamran, M.; Malik, Z.; Parveen, A.; Zong, Y.; Abbasi, G.H.; Rafiq, M.T.; Shaaban, M.; Mustafa, A.; Bashir, S.; Rafay, M. Biochar alleviates Cd phytotoxicity by minimizing bioavailability and oxidative stress in pak choi (Brassica chinensis L.) cultivated in Cd-polluted soil. J. Environ. Manag. 2019, 250, 109500. [Google Scholar] [CrossRef]
- Kamran, M.; Malik, Z.; Parveen, A.; Huang, L.; Riaz, M.; Bashir, S.; Mustafa, A.; Abbasi, G.H.; Xue, B.; Ali, U. Ameliorative Effects of Biochar on Rapeseed (Brassica napus L.) Growth and Heavy Metal Immobilization in Soil Irrigated with Untreated Wastewater. J. Plant Growth Regul. 2019, 1–16. [Google Scholar] [CrossRef]
- Uddin Nizam, M.; Mokhlesur Rahman, M.; Kim, J.-E. Phytoremediation Potential of Kenaf (Hibiscus cannabinus L.), Mesta (Hibiscus sabdariffa L.), and Jute (Corchorus capsularis L.) in Arsenic-contaminated Soil. Korean J. Environ. Agric. 2016, 35, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Gang, A.; Vyas, A.; Vyas, H. Toxic effect of heavy metals on germination and seedling growth of wheat. J. Environ. Res. Dev. 2013, 8, 206. [Google Scholar]
- Singh, K.P.; Mohan, D.; Sinha, S.; Dalwani, R. Impact assessment of treated/untreated wastewater toxicants discharged by sewage treatment plants on health, agricultural, and environmental quality in the wastewater disposal area. Chemosphere 2004, 55, 227–255. [Google Scholar] [CrossRef] [PubMed]
- Akram, N.A.; Iqbal, M.; Muhammad, A.; Ashraf, M.; Al-Qurainy, F.; Shafiq, S. Aminolevulinic acid and nitric oxide regulate oxidative defense and secondary metabolisms in canola (Brassica napus L.) under drought stress. Protoplasma 2018, 255, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Schmidthoffer, I.; Szilák, L.; Molnár, P.; Csontos, P.; Skribanek, A. Drought tolerance of European barley (Hordeum vulgare L.) varieties. Agriculture (Pol’nohospodárstvo) 2018, 64, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.N.; Zhang, J.; Luo, T.; Liu, J.; Ni, F.; Rizwan, M.; Fahad, S.; Hu, L. Morpho-physiological and biochemical responses of tolerant and sensitive rapeseed cultivars to drought stress during early seedling growth stage. Acta Physiol. Plant. 2019, 41, 25. [Google Scholar] [CrossRef]
- Uddin, M.N.; Wahid-Uz-Zaman, M.; Rahman, M.M.; Islam, M.S.; Islam, M.S. Phytoremediation Potentiality of Lead from Contaminated Soils by Fibrous Crop Varieties. Am. J. Appl. Sci. Res. 2016, 2, 22. [Google Scholar] [CrossRef] [Green Version]
- Wodala, B.; Eitel, G.; Gyula, T.; Ördög, A.; Horváth, F. Monitoring moderate Cu and Cd toxicity by chlorophyll fluorescence and P 700 absorbance in pea leaves. Photosynthetica 2012, 50, 380–386. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, X.; Yang, L.; Tang, M.; Wang, K.; Wang, L.; Bai, L.; Song, C. Hydrogen peroxide plays an important role in PERK4-mediated abscisic acid-regulated root growth in Arabidopsis. Funct. Plant Biol. 2018, 46, 165–174. [Google Scholar] [CrossRef]
- Liu, H.; Guo, S.; Lu, M.; Zhang, Y.; Li, J.; Wang, W.; Wang, P.; Zhang, J.; Hu, Z.; Li, L.; et al. Biosynthesis of DHGA12 and its roles in Arabidopsis seedling establishment. Nat. Commun. 2019, 10, 1768. [Google Scholar] [CrossRef]
- Brahim, L.; Mohamed, M. Effects of copper stress on antioxidative enzymes, chlorophyll and protein content in Atriplex halimus. Afr. J. Biotech. 2011, 10, 10143–10148. [Google Scholar]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes; suppressed cadmium uptake and oxidative stress in cotton. Ecotoxicol. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qin, C.; Shu, X.; Chen, R.; Song, H.; Li, Q.; Xu, H. Effects of copper on induction of thiol-compounds and antioxidant enzymes by the fruiting body of Oudemansiella radicata. Ecotoxicol. Environ. Saf. 2015, 111, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Muszynska, E.; Hanus-Fajerska, E. Why are heavy metal hyperaccumulating plants so amazing? BioTechnol. J. Biotech. Comput. Biol. Bionanotech. 2015, 96, 265–271. [Google Scholar] [CrossRef]
- Saleem, M.H.; Fahad, S.; Khan, S.U.; Ahmar, S.; Khan, M.H.U.; Rehman, M.; Maqbool, Z.; Liu, L. Morpho-physiological traits, gaseous exchange attributes, and phytoremediation potential of jute (Corchorus capsularis L.) grown in different concentrations of copper-contaminated soil. Ecotoxicol. Environ. Safe. 2019. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Khan, M.A.; Asaf, S.; Lee, I.-J.; Kim, K.M. Metal resistant endophytic bacteria reduces cadmium, nickel toxicity, and enhances expression of metal stress related genes with improved growth of Oryza Sativa, via regulating its antioxidant machinery and endogenous hormones. Plants 2019, 8, 363. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Hou, M.; Cao, L.; Xia, Y.; Shen, Z.; Hu, Z.J.E. Glutathione S-transferases modulate Cu tolerance in Oryza sativa. Environ. Exp. Bot. 2018, 155, 313–320. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Plant Height | Plant Fresh Weight | Plant Dry Weight | Total Chlorophyll | Seed Germination |
|---|---|---|---|---|---|
| HongTieGuXuan | 3.72 ± 0.05 a | 0.255 ± 0.05 b | 0.103 ± 0.005 a | 2.64 ± 0.09 a | 100 |
| C-3 | 3.73 ± 0.85 a | 0.3 ± 0.01 a | 0.100 ± 0.01 a | 2.82 ± 0.03 a | 100 |
| GuBaChangaJia | 0.87 ± 0.81 b | 0.103 ± 0.003 c | 0.045 ± 0.015 b | 1.51 ± 0.03 b | 77.5 |
| ShangHuoMa | 0.61 ± 0.13 b | 0.105 ± 0.003 c | 0.040 ± 0.02 b | 1.47 ± 0.08 b | 75 |
| Varieties | Cu Concentration in Roots | Cu Concentration in Shoots |
|---|---|---|
| HongTieGuXuan | 56 ± 1.4 b | 57 ± 1.1 b |
| C-3 | 60 ± 0.8 a | 61 ± 0.9 a |
| GuBaChangaJia | 40 ± 0.8 c | 41 ± 0.6 c |
| ShangHuoMa | 37 ± 1 d | 38 ± 0.9 d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleem, M.H.; Ali, S.; Seleiman, M.F.; Rizwan, M.; Rehman, M.; Aisha Akram, N.; Liu, L.; Alotaibi, M.; Al-Ashkar, I.; Mubushar, M. Assessing the Correlations between Different Traits in Copper-Sensitive and Copper-Resistant Varieties of Jute (Corchorus capsularis L.). Plants 2019, 8, 545. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120545
Saleem MH, Ali S, Seleiman MF, Rizwan M, Rehman M, Aisha Akram N, Liu L, Alotaibi M, Al-Ashkar I, Mubushar M. Assessing the Correlations between Different Traits in Copper-Sensitive and Copper-Resistant Varieties of Jute (Corchorus capsularis L.). Plants. 2019; 8(12):545. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120545
Chicago/Turabian StyleSaleem, Muhammad Hamzah, Shafaqat Ali, Mahmoud F. Seleiman, Muhammad Rizwan, Muzammal Rehman, Nudrat Aisha Akram, Lijun Liu, Majed Alotaibi, Ibrahim Al-Ashkar, and Muhammad Mubushar. 2019. "Assessing the Correlations between Different Traits in Copper-Sensitive and Copper-Resistant Varieties of Jute (Corchorus capsularis L.)" Plants 8, no. 12: 545. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120545