An Exploration of Common Greenhouse Gas Emissions by the Cyanobiont of the Azolla–Nostoc Symbiosis and Clues as to Nod Factors in Cyanobacteria

Department of Botany, University of Sri Jayewardenepura, Gangodawila 10250, Sri Lanka
Plants 2019, 8(12), 587; https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120587
Submission received: 21 November 2019
/
Revised: 30 November 2019
/
Accepted: 4 December 2019
/
Published: 10 December 2019
(This article belongs to the Special Issue Nitrogen-Fixing Plants )
Abstract
:Azolla is a genus of aquatic ferns that engages in a unique symbiosis with a cyanobiont that is resistant to cultivation. Azolla spp. are earmarked as a possible candidate to mitigate greenhouse gases, in particular, carbon dioxide. That opinion is underlined here in this paper to show the broader impact of Azolla spp. on greenhouse gas mitigation by revealing the enzyme catalogue in the Nostoc cyanobiont to be a poor contributor to climate change. First, regarding carbon assimilation, it was inferred that the carboxylation activity of the Rubisco enzyme of Azolla plants is able to quench carbon dioxide on par with other C3 plants and fellow aquatic free-floating macrophytes, with the cyanobiont contributing on average ~18% of the carboxylation load. Additionally, the author demonstrates here, using bioinformatics and past literature, that the Nostoc cyanobiont of Azolla does not contain nitric oxide reductase, a key enzyme that emanates nitrous oxide. In fact, all Nostoc species, both symbiotic and nonsymbiotic, are deficient in nitric oxide reductases. Furthermore, the Azolla cyanobiont is negative for methanogenic enzymes that use coenzyme conjugates to emit methane. With the absence of nitrous oxide and methane release, and the potential ability to convert ambient nitrous oxide into nitrogen gas, it is safe to say that the Azolla cyanobiont has a myriad of features that are poor contributors to climate change, which on top of carbon dioxide quenching by the Calvin cycle in Azolla plants, makes it an efficient holistic candidate to be developed as a force for climate change mitigation, especially in irrigated urea-fed rice fields. The author also shows that Nostoc cyanobionts are theoretically capable of Nod factor synthesis, similar to Rhizobia and some Frankia species, which is a new horizon to explore in the future.

1. Introduction
Three gases—carbon dioxide, methane (~34 times greater global warming potential than carbon dioxide), and nitrous oxide (a ~298-fold higher global warming potential than carbon dioxide)—are the chief contributors to global warming and, consequently, climate change [1,2]. The cultivation of crops or agriculture is one of the main causes of climate change, because it leads to the emission of all three of these gases. Heavy machinery and the Haber process, which is used to produce ammonia fertilizers, have high carbon footprints. Farming of ruminants and rice cultivation, which promote anaerobic decomposition, are both emitters of methane. Finally, the use of synthetic nitrogen fertilizers potentiates the denitrification of nitrogen compounds, which leads to the emission of nitrous oxide as a by-product. Therefore, it is necessary to reduce the effect of agriculture on climate change by promoting the use of biological alternatives to nitrogen fertilizers (biofertilizers), i.e., bacterial/cyanobacterial organisms that possess the remarkable ability to fix nitrogen gas and to furnish the nitrogen requirements of plants [3]. Nitrogen fixation is biologically viable, does not pollute or emit greenhouse gases, and is cost effective and user friendly, which makes this biological process a potent player in mitigating future climate change. Notably, among such alternative solutions is a plant, the aquatic fern Azolla sp., that can act as a biofertilizer by the fortification of a plant biological system with a nitrogen-fixing cyanobiont that churns out ammonia in abundance [3].
Azolla spp. can quench ample carbon dioxide due to their rapid proliferation, doubling their body weight in 2–5 days [4]. Azolla spp. are grown in paddy fields, where it has been measured that the yields of rice are significantly elevated due to dual cropping [5,6], while the lowering of the urea footprint is also a positive outcome based on the growth of Azolla spp. in irrigated lands. Urea has been shown to drastically improve nitrous oxide production while being a lesser candidate for the emission of methane; however, it increases the carbon dioxide footprint, which means that using biofertilizers as surrogates to urea diminishes climate change in three combined mechanisms [3,7]. At an ambient CO2 concentration of 800 ppm, Azolla spp., in a small-scale production system, showed a strong increase in biomass production, i.e., 36–47%, which establishes the potential of Azolla spp. as a solution to elevated atmospheric carbon dioxide levels [8]. Azolla spp. are C3 plants, and based on the first assimilation step by the action of the Rubisco carboxylase, they provide a heightened form of assimilation which would be negligible in C4 plants, where the first step is a concentrating/enrichment mechanism [9,10,11].
The production of copious amounts of ammonia via nitrogen fixation, which is then transported as amino acids such as glutamine, gives aquatic ferns abundant amounts of the Rubisco enzyme, which is the workhorse of plants carboxylating ribulose-1,5-disphosphate into carbon skeletons made of six carbon atoms [12]. In plants, Rubisco is said to compose ~30% of the total protein content, which may be even higher for an ammonium-churning engine such as the Azolla superorganism. Although rightfully considered a carbon quencher, this benefit would be made irrelevant by the production of more potent greenhouse gases such as nitrous oxide, methane, and halocarbons from the microbiome associated with the aquatic Azolla fern, mainly, the commonly-found cyanobiont which is popularly known as Nostoc azollae. It is therefore imperative to investigate the potential of both (1) the Azolla sp. (the host organism) and (2) the Nostoc major cyanobiont as contributors to greenhouse gas reductions. The major cyanobiont is known by three generic names, Nostoc, Anabaena, and Trichormus [13], but for the sake of simplicity and consistency, I have employed the name N. azollae throughout the article.
This study focused mostly on the cyanobiont N. azollae and its paucity of de novo production of greenhouse gases, with the exception of carbon dioxide, which can be soaked/negated conceptually by the Azolla aquatic fern. I also shed light on the nature of putative signaling between the cyanobiont and host plant system using bioinformatics. To my knowledge, this is the first time that cyanobacteria have been shown (at the genome level) to possess putative Nod-factor-modifying enzyme encoding genes.
This study only showcased the absence of key proteins for greenhouse gas production in the cyanobiont harbored inside Azolla spp. by using simple bioinformatics tools. The results suggest that Azolla spp. can be particularly useful for climate change mitigation, and this potential should be harnessed. This study will hopefully trigger physiological and climate change mitigation studies on the carbon fixation potential of Azolla species.
2. Results and Discussion
2.1. Carbon Dioxide Emissions by the Cyanobiont N. azollae
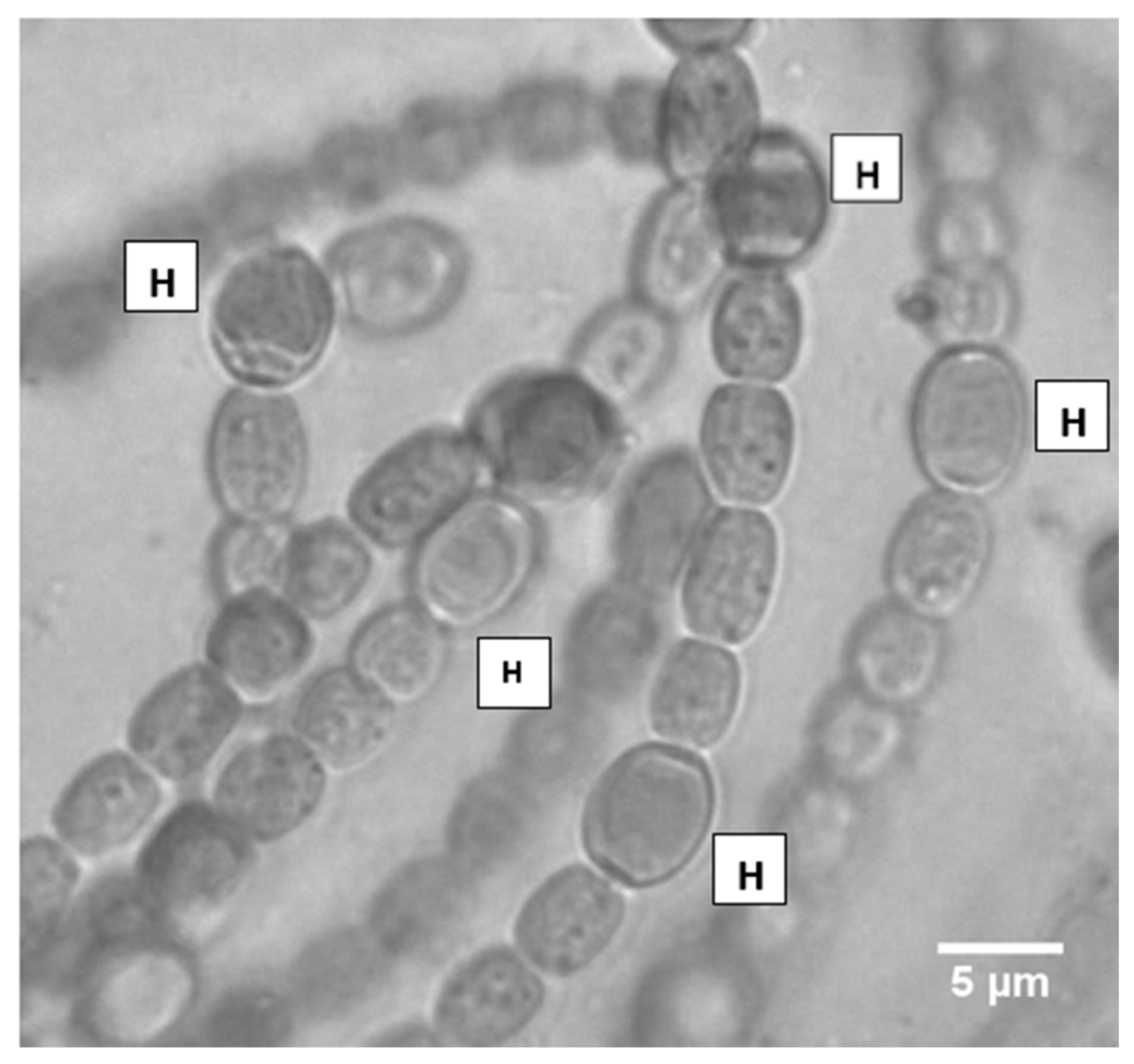
Due to the widespread presence of N. azollae inside Azolla fronds, it is unclear how much of the carbon dioxide that is emanated by Nostoc colonies is quenched by Azolla plants. It is known that in heterocysts, where nitrogen fixation occurs, there are strong respiration rates to maintain a low stream of oxygen at the active site of the nitrogenase enzyme, which means that heightened carbon dioxide emissions are a common occurrence from Nostoc filaments, especially in the dark. In this study on N. azollae, heterocysts were found as close as two cells apart, and were commonly spaced every 2–10 cells (Figure 1). The presence of a high number of heterocysts, compared with other Nostoc species (Table 1), means that N. azollae could be a potent contributor to greenhouse gas emissions and, consequently, climate change, unless the emanated carbon dioxide is quenched by photosynthesis (Calvin cycle) taking place in the Azolla fronds.
Due to the high production of ammonium by nitrogen fixation in the filaments of N. azollae, Azolla plants, which benefit from the copious amounts of fixed nitrogen, can grow more rapidly, and quickly double their biomass. Ironically, the compartment (heterocyst) that synthesizes the nitrogen for Rubisco also produces high amounts of heterocystous carbon dioxide, which means that both the enzyme Rubisco and the substrate carbon dioxide emanate as physiological/biochemical products from the cyanobiont.
Azolla cyanobionts have 5-fold less Rubisco, as quantified by their transcript numbers, compared with their free-living counterparts, and cyanobionts only have a partial/depleted carbon dioxide fixing ability [14]. This puts the responsibility of counteracting the carbon dioxide emanated by heterocysts—and overall, by Nostoc filaments—on the shoulders of the plant-type Rubisco enzyme.
Rubisco is known to be present at around 30% of a plant’s catalogue of proteins. Here, however, I was unable to quantify the level of Rubisco in Azolla plants, which has not been indicated in the literature. It has been shown that carbon dioxide assimilation takes place at 90–100 µmol CO2/mg Chl.h for Azolla symbiosis and 75–80 µmol CO2/mg Chl.h for Azolla lacking the cyanobiont [15], which shows that photosynthesis is affected when the symbiosis is absent. Therefore, for maximum carbon assimilation, the cyanobiont is a critical component of the Azolla system.
Furthermore, in C3 plants, there is 15–35 mg CO2/(dm2·h), while in C4 plants, there is 40–80 mg CO2/(dm2·h), which hints that the rates of carbon capture are relatively conservative in the Azolla–Nostoc system, even without quantifying the cyanobiont’s contribution [16]. On average, there is 3–5 mg of chlorophyll in 1 dm2 of leaf tissue, which is between 12–22 mg CO2/(dm2·h) for Azolla–cyanobiont, a conservative measurement for the plant system, being low to moderate among C3 counterparts, but similar to other floating aquatic plants that assimilate CO2 over the water surface at 20–23 mg CO2/(dm2·h) [17]. Still, this value may be higher, considering that the CO2 in the atmosphere is beyond 415 ppm now, and the aquatic fern’s carboxylation potential was measured in a study 40 years ago.
2.2. Oxides of Nitrogen
In this bioinformatics exploration, I examined the potential of the cyanobiont of Azolla spp. to be a nitrous oxide, methane, and halocarbon producer, compared to other members of the order Nostocales. I hypothesized that greenhouse-gas-producing enzymes were made redundant at the genome level in the Azolla cyanobiont, as it lost its independence and was put under the protection of the carbon nutrition of the host organism, which is a case of “genomic erosion” [20].
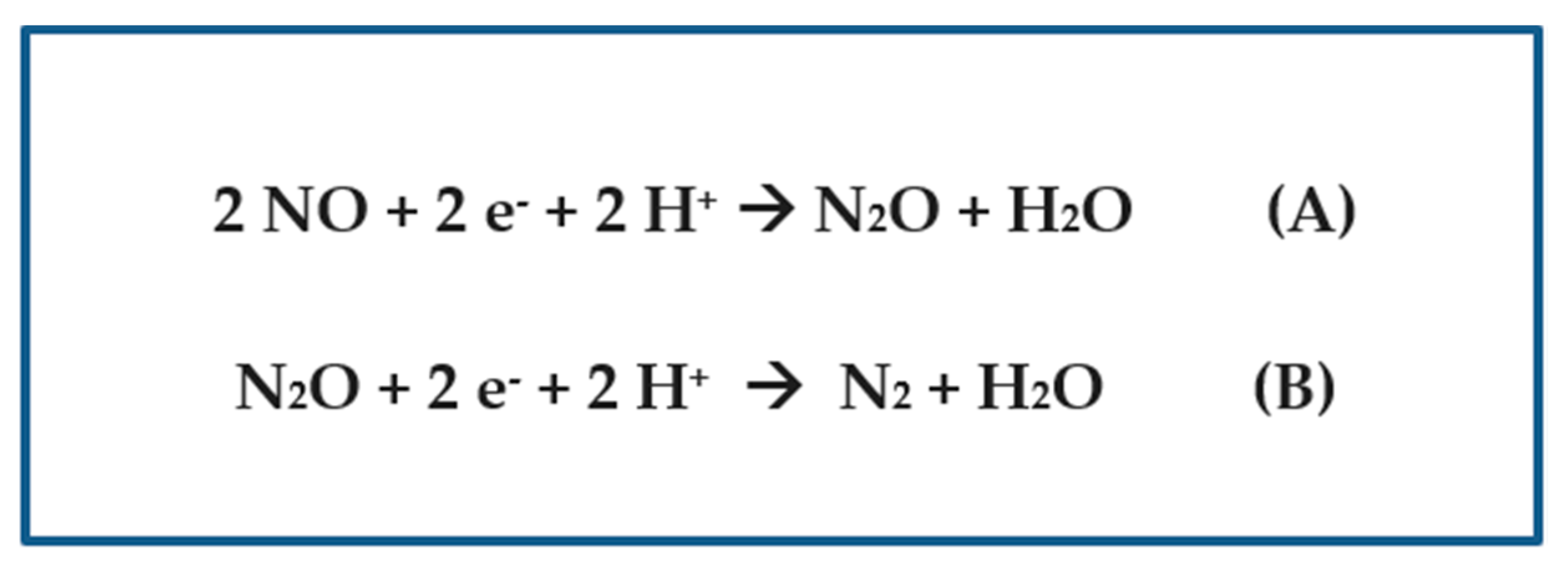
Regarding nitrous oxide production, I focused first on the nitric oxide reductase (GenBank: AFZ55274.1, belonging to Cyanobacterium aponinum PCC 10605) (Table 2) and the nitrous oxide reductase (RCJ21339.1, belonging to Nostoc sp. ATCC 43529) (Table 2), which catalyze sequential steps in the mechanism of denitrification, by using the PSI-BLAST search tool to unveil their presence (or absence) in their respective proteomes [21]. The reactions that these enzymes catalyze are shown in Figure 2. Surprisingly, the genus Nostoc (as well as the sibling Anabaena) was found not to possess any nitric oxide reductases that catalyze the production of the greenhouse gas nitrous oxide, but it does contain putative nitrous oxide reductases that produce dinitrogen gas (Table 3).
It seems, theoretically, that Nostoc spp. have an evolutionary adaptation to convert nitrous oxide in the ambient environment into dinitrogen gas, which can be utilized in subsequent nitrogen fixation, which is an extremely clever biological process. The same enzyme (a putative nitrous oxide reductase) was found to be present in the proteome of the cyanobiont of Azolla spp. at 31% identity and 55% coverage (Table 3), showcasing the value of growing Azolla in rice fields, where urea is used abundantly as a chemical fertilizer, to negate nitrous oxide emissions.
On the other hand, nitric oxide synthases, which synthesize NO, were found in two filamentous subsections, and are available in plant cyanobionts such as Nostoc cycadae. However, nitric oxide synthases are absent in the Azolla cyanobiont’s genome/proteome (Table 3). Nevertheless, it is interesting that there is an ad hoc nature to the presence of the nitric oxide synthases, being found in only a few species of Nostoc and Anabaena, which are highly abundant species of the order Nostocales. Interestingly, the Nostoc sp. HK-1 (which establishes a symbiosis with Cycas plants), which is a drought-tolerant species of Nostoc that has been suggested to be a contender that can survive in extraterrestrial environments, such as on Mars. However, in the absence of lab-based evidence, the possibility of the role of NO in cellular signaling and in the dialogue between plants and microbes cannot be eliminated.
The accumulation of NO, which takes place in Nostoc species, appears to be a signaling event between plants and cyanobacteria. Still, there is another enzymatic contender for the production of NO, namely, the family of nitrite reductases. Table 3 presents a summary of the representation of subsections IV and V of cyanobacteria and their complement of nitrogen-based enzymes, in particular, denitrifying proteins. Of note, N. azollae also had a low homology match (22% identity and 30% coverage) to nitrite reductases, which requires experimental data to convincingly demonstrate that it is an authentic nitrite reductase (Figure S1).
It is noteworthy that, in the past, scientists questioned whether NO is more important for plants or plant pathogens, since it is exuded by the latter [22]. In a similar context, the presence of NO, but not N2O, is a measure of the use of NO in Nostoc biology. It is known that microorganisms that are capable of parasitism utilize NO in highly efficient, constitutive, and inducible ways [22]. Therefore, the presence of NO could analogously be a windfall for the Nostoc cyanobacteria, especially those capable of symbiotic unions. Thus, similar to being a virulence factor in plant pathogens [22], NO could well be a symbiotic factor in cyanobacteria capable of symbioses. Current knowledge related to this is strictly limited in showcasing all the biological phenomena associated with NO [22].
2.3. Cyanobacterial Signaling with Host: New In Silico-Based Evidence on Nod Factors
The key finding of this study is the gap in the denitrification process in Nostoc species (Table 3). Still, the presence of nitrite reductases in some Nostoc species and nitric oxide synthases in others presents a picture of accumulating NO levels that are not utilized downstream due to the absence of nitric oxide reductases, which prevents the production of nitrous oxide by any Nostoc species (Table 3).
Furthermore, I hypothesize that the presence and accumulation of nitric oxide may have a role to play in the establishment of symbioses with host plants. Fascinatingly, (Ns) H-NOX is a hemeprotein found in symbiotic cyanobacteria that is markedly similar to the β subunit of soluble guanylyl cyclases (sGC), which indicates a NO-sensing function in cyanobacteria that may use plant-derived NO for various processes [23]. However, it does not explain the de novo synthesis of NO in such cyanobacteria using nitric oxide synthases and nitrite reductases, although the emanation of NO by cyanobacterium can be a mechanism for drawing the root closer toward the cyanobacterium, aided by the elongation of the root.
Considering the similarities between the genera Frankia and Nostoc, in that both form symbiotic unions with land plants while producing an organ such as a root nodule and possessing the ability to differentiate into a distinct cell/vesicle to fix nitrogen, I checked for the presence of nodulation protein NodA, nodulation protein H, and NodB polysaccharide deacetylase from “Candidatus Frankia californiensis” [24] as the search query using the PSI-BLAST search tool. Table 4 clearly shows the presence of NodB polysaccharide deacetylases in cyanobacterial genomes, including that of Nostoc spp., which draws a link between two filamentous/mycelial types in two divergent plant systems. What is most interesting is that the NodB polysaccharide deacetylase family is strongly conserved in several cyanobacterial species, especially those of the order Nostocales.
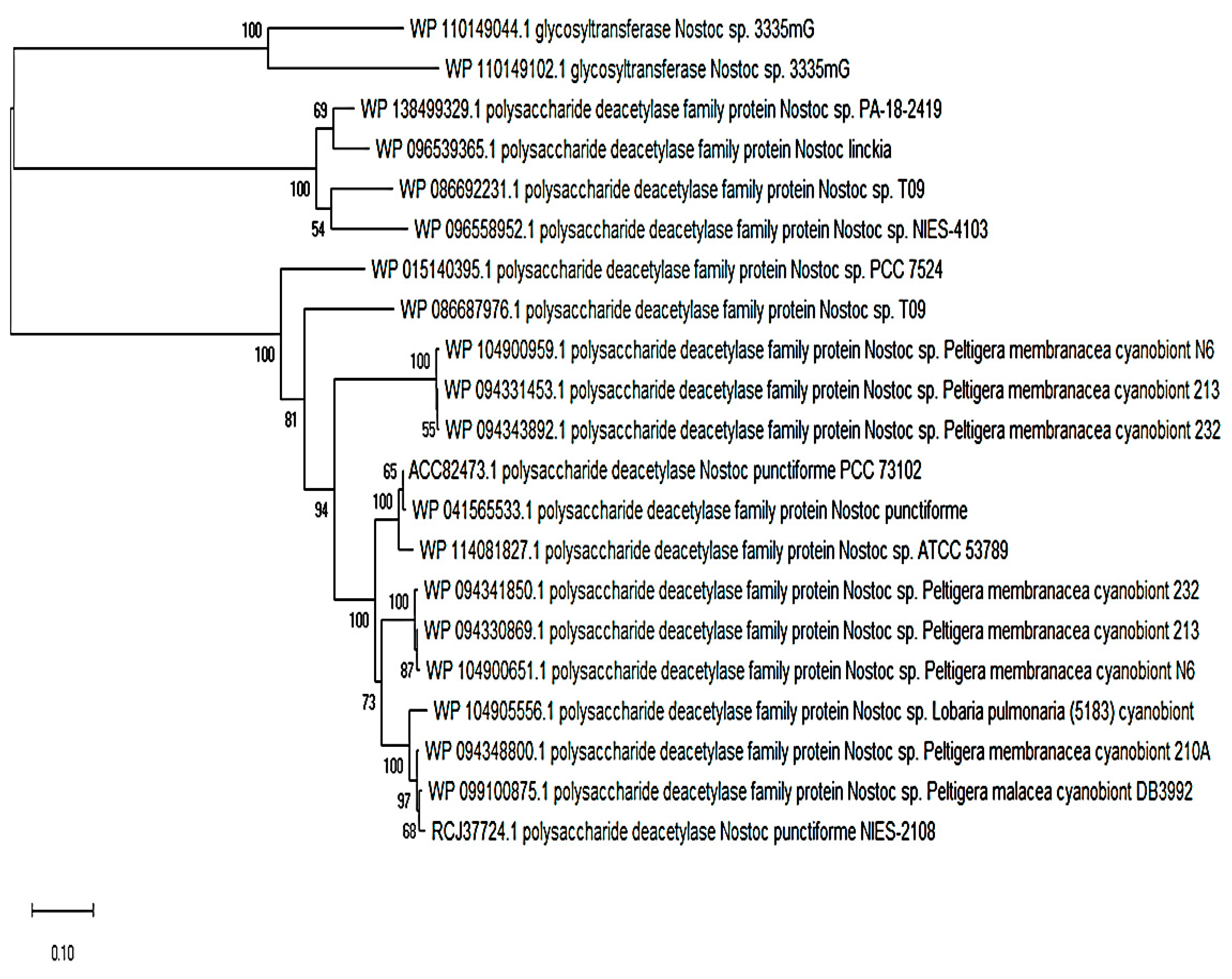
I also further searched the genus Nostoc with a NodB polysaccharide deacetylase protein from a rhizobial strain (Rhizobium leucaenae USDA 9039) using the PSI-BLAST search tool. I found a myriad of Nostoc species that possess homologues of >30% identity and 80% or over sequence coverage. The first 25 hits on the PSI-BLAST search were mostly symbiotically-inclined cyanobionts representing hosts such as lichens, lower plants, gymnosperms, and angiosperms (Figure 3). However, there were NodB polysaccharide deacetylases found in noncyanobionts.
It is worth considering what made the NodB family of polysaccharide deacetylases important for communication by cyanobacteria, as revealed by their widespread conservation (Figure 3). It seems that when it comes to partnerships such as Rhizobia—legumes, actinorhizal-plants—Frankia, and host—cyanobacterium symbiotic systems, there is a role to be played by NodB polysaccharide deacetylases, which is clearly shown by the level of sequence conservation in cyanobionts. This family of proteins (carbohydrate esterase family 4 (CE4)) may perform its function by cleaving an N-acetyl moiety from the nonreducing end of the chitin oligosaccharide (CO) molecule [25].
Surprisingly, even NodC nodulation proteins (glycosyltransferases) were present in PSI-BLAST queries of a NodC protein (R. leucaenae USDA 9039), with the best match being 55% coverage/35% identity (Figure S2). NodC proteins are N-acetylglucosaminyltransferases forming chitin oligosaccharide molecules. Therefore, two sequential steps of the rhizobial nod factor design pathway (NodCand NodB nodulation proteins) are putatively conserved in many Nostoc species.
Surprisingly, the NodB polysaccharide deacetylase protein is found in the cyanobiont of Azolla spp. at 90% query cover and 35% sequence identity to the Rhizobium query sequence, while the NodC acetylglucosaminyltransferase is found at 34.7% coverage but only 22% identity when searched with the respective proteins of R. leucaenae USDA 9039 (Table 5). Whether or not the activities of synthesis and deacetylation of the chitin oligosaccharide molecules take place in Azolla spp. is dependent on showing experimentally that they have a function in the symbiosis. This changes our perception of the subject; that is, N. azollae, contrary to the notion that it is incapable of horizontal transmission, is perhaps capable of infection, or once had such a capability.
Upon searching the Azolla plant proteome for the presence of the LysM receptor-like kinase (Nod factor perception protein), which mediates Nod factor recognition, I found that they were absent in the genus Azolla. Perhaps contrary to the belief that it is the cyanobiont that lost its capacity to infect, I hypothesize here that the Azolla plant does not possess LysM receptor-like kinases for Nod factor perception. I suggest that the Nod factors in N. azollae are involved in a different, unknown context to mere recognition and infection, which is also supported by the presence of NodB nodulation proteins in noncyanobionts incapable of symbioses.
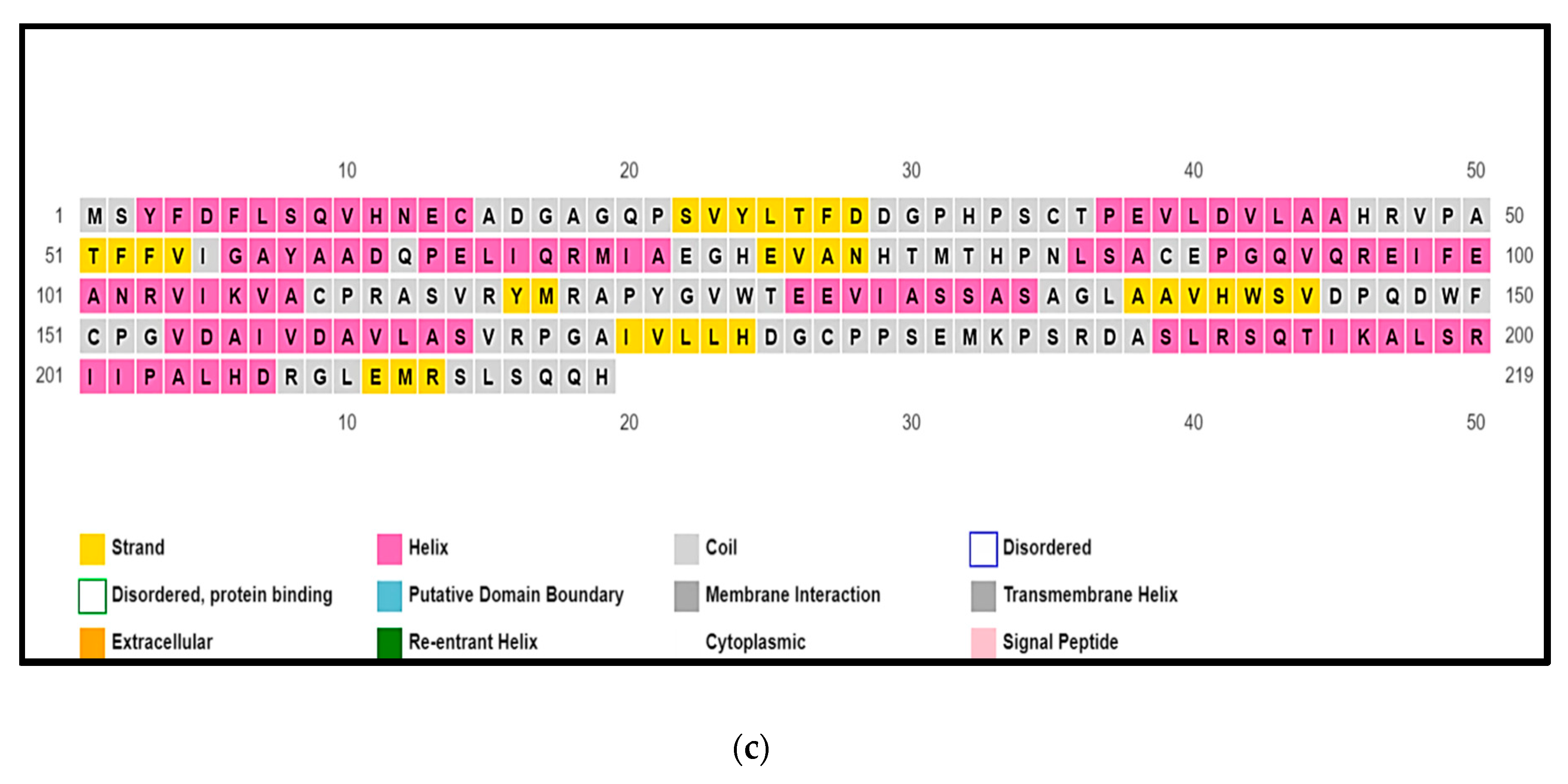
Using ClustalW, I aligned four sequences of putative NodB polysaccharide deacetylases from an actinorhizal symbiont, a species of Rhizobium, and homologues from N. azollae and Nostoc punctiforme to examine the sequences in detail (Figure 4). The NodB domain was found mostly within the globular fold of the NodB proteins (Figure 4b), which aligned well with its role as the catalytically-competent area of the protein. However, the NodB domain was interrupted (a deleted region) in both cyanobacterial sequences after the fifth conserved motif/patch (Figure 4a), which highlighted the fact that the substrate of cyanobacterial NodB proteins may perhaps be different than the bacterial counterparts in their substrate profiles, or such cyanobacterial proteins are inactivated or pseudogenized at the genome level. The deleted region (Figure 4a) corresponded to a coil region (Figure 4c) when the secondary structural motifs were predicted of the AFJ42532.1 NodB protein (Mesorhizobium plurifarium).
2.4. Methane Emissions
Cyanobacteria have been shown to produce methane from aquatic and terrestrial ecosystems [26]. It is a stark reality that biogenic methane production by cyanobacteria affects the recent and future methane budgets, and is also part of a protracted time frame (3.5 billion years) since their evolutionary beginnings, where they would have furnished methane to the ambient environment [26]. It is hypothesized that the increased rates of hydrogen production in diazotrophs, such as the cyanobiont N. azollae, promote the production of methane through the transfer of hydrogen gas into hydrogenotrophic methanogenic bacteria in the ambient environment [26]. Still, the cultivation of Azolla spp. in methane-emanating rice paddies, which are hot spots for methanogenic phenomena, has been shown to alleviate methane emissions in dual cropping systems [27]. The supposed explanation of this observation is that the dissolved oxygen in irrigation systems and the soil redox potential become favorable under Azolla cultivation [27].
I searched cyanobacteria for the presence of methanogenic reactions using the hypothetical protein CEN44_02070 (Fischerella muscicola CCMEE 5323), which can synthesize methane from methyl-coenzyme M and coenzyme B through a methyl-coenzyme M reductase (MCR) gamma subunit domain, which is abundant in methanogenic organisms, including cyanobacteria. Again, no homologues of the abovementioned protein were found in the proteome of the Azolla cyanobiont (Table 3). I then constructed a phylogenetic tree with some of the best aligned sequences to the hypothetical protein CEN44_02070 (F. muscicola CCMEE 5323), where I found an enrichment/abundance of sequences from cyanobionts, from several sources, compared with those of free-living cyanobacteria. In particular, there was a strong representation of the Nostoc partners of Peltigera lichens in the phylogenetic reconstruction (Figure 5).
In addition, it was recently shown that the wild-type iron–iron (Fe-only) nitrogenase from the bacterium Rhodopseudomonas palustris can produce hydrogen, ammonia, and methane in a single enzymatic reaction [28]. Therefore, I also checked this using PSI-BLAST, considering the unlikely possibility of N. azollae possessing this rare enzyme (iron–iron nitrogenase), but I failed to identify a candidate enzyme in the database.
2.5. Halocarbon Emissions
I searched for vanadium-dependent haloperoxidases that can synthesize halocarbons as by-products from the oxidation of halides, which can damage the ozone layer and contribute to global warming [29]. It is interesting that almost all cyanobionts of plants and lichens have a vanadium nitrogenase as a component of their two-prong nitrogen fixation systems, which makes the partitioning of vanadium to the nitrogenase and haloperoxidases a critical task, when it is present [30,31].
Chloroperoxidases and bromoperoxidases have highly different protein sequences, but are conserved in their catalytic mechanisms, proving that enzyme evolution takes place by modifying the substrate binding site but keeping the coordination chemistry of the active site [32]. I searched the vanadium chloroperoxidase and bromoperoxidase from Nostoc sp. PCC 7120 and Nostoc commune NIES-4072, respectively, against the N. azollae proteome. The former (vanadium chloroperoxidase) yielded the best match at 84% coverage and 48% sequence identity (Table 3), while the latter was not found, showing that vanadium chloroperoxidases may be a potential source of greenhouse gases in Azolla spp.
3. Conclusions
This bioinformatics study demonstrated the paucity of greenhouse gas emissions by the Azolla—cyanobiont symbiosis, namely, nitrous oxide and methane, with the exception of some forms of halocarbons and carbon dioxide, which are emanated by N. azollae. It was also shown that the released carbon dioxide can be quenched by the fronds of Azolla spp. at the same efficiency and range as other aquatic floating plants and C3 counterparts. It was further demonstrated, based on computational biology, that there is nitrous oxide quenching and transformation into dinitrogen gas by the Azolla cyanobiont, offering a near-ideal biological system for greenhouse gas mitigation for rice cultivation with extensive urea usage. Cyanobacteria also possess the genomic ability to produce Nod factors, which are, surprisingly, found in N. azollae as well.
4. The Future
During the Azolla event 49 million years ago, Azolla blooms in the Arctic Ocean were able to trap carbon dioxide and become sediments on the ocean floor, which changed the earth from a greenhouse into an icehouse [33]. This one event by itself demonstrates the importance of Azolla spp. in a future world impacted by climate change, where they could play a crucial role in reversing greenhouse effects and the resulting threat of climate change, which has uprooted lives in island nations such as Sri Lanka. Such places have already witnessed changing weather patterns and the degradation of the overall quality of life. In that stark horizon, there lies a role for the genus Azolla to be a frontrunner to combat climate change.
5. Materials and Methods
5.1. Sequence Queries
Proteins were identified from past literature and searched using the NCBI protein database (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/protein) using word-based queries. The most crucial word-based protein searches are provided in Table 2.
I assessed the top ~100 sequences derived from PSI-BLAST searches and downloaded nonredundant sequences for phylogenetic tree construction. Phylogenetic trees played two roles in this study, i.e., providing a visual/nominal identity, and showing the phylogeny of cyanobionts against noncyanobionts. I attempted, when possible, to employ a balance between free-living cyanobacteria and cyanobionts, while eliminating redundant sequences.
The PSI-BLAST query focused on the umbrella taxon “cyanobacteria” using the FASTA sequence of the sequence query to run the search tool (provided in Table 2). Furthermore, for specific queries such as nitric oxide reductases, I employed a “Nostocales”-only PSI-BLAST search, which failed to yield any Nostoc or Anabaena species, even at a low sequence homology level, demonstrating their absence in such genera. This search, in particular, was conducted to ensure that both the genera Nostoc and Anabaena were without nitric oxide reductases, a key finding of this study. Similarly, more specific searches were also conducted at the genus and species levels according to the impending requirements.
The threshold/cut-off of 30% sequence identity was used as an indicator of members of a single family of proteins according to the Structural Classification of Proteins (SCOP) rules [34]. The 30% coverage was more of an arbitrary value, considering the spans of sequence conservation.
5.2. Phylogenetic Reconstructions
The nonredundant downloaded amino acid sequences (as FASTA files) from each query were first aligned with the ClustalW algorithm using MEGA version X (default parameters) [35], which were converted to the MEGA sequence format, and phylogenetic reconstruction was performed using the neighborhood joining/maximum parsimony methods (again in MEGA version X) with support from 500 bootstrap replications. There was no assignment of outgroups, since the phylogenetic trees were used here to showcase the topmost hits (the IDs) and, to a lesser extent, to show phylogeny between cyanobiont sequence clusters.
5.3. Secondary Structure Prediction
The secondary structure prediction service PSIPRED 4.0 (http://bioinf.cs.ucl.ac.uk/psipred/) was used to showcase the helixes, beta strands, and coils [36].
5.4. Globular Domain Prediction
Globular domains were predicted using the bioinformatics server at http://globplot.embl.de/ [37].
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2223-7747/8/12/587/s1, Figure S1: Screenshot of PSI-BLAST result for Nitrite Reductases in N. azollae., Figure S2: Screenshot of PSI-BLAST query result, when a single NodC nodulation protein from R. leucaenae USDA 9039 was searched specifically against cyanobacteria.
Funding
The research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Beaulieu, J.J.; DelSontro, T.; Downing, J.A. Eutrophication will increase methane emissions from lakes and impoundments during the 21st century. Nat. Commun. 2019, 10, 1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassia, R.; Nocioni, M.; Correa-Aragunde, N.; Lamattina, L. Climate Change and the Impact of Greenhouse Gasses: CO2 and NO, Friends and Foes of Plant Oxidative Stress. Front. Plant Sci. 2018, 9, 273. [Google Scholar] [CrossRef] [PubMed]
- Kollah, B.; Patra, A.K.; Mohanty, S.R. Aquatic microphylla Azolla: A perspective paradigm for sustainable agriculture, environment and global climate change. Environ. Sci. Pollut. Res. Int. 2016, 23, 4358–4369. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Song, U. Microcosm investigation of growth and phytoremediation potential of Azolla japonica along nitrogen gradients. Int. J. Phytoremediat. 2017, 19, 863–869. [Google Scholar] [CrossRef]
- Khumairoh, U.; Lantinga, E.A.; Schulte, R.P.; Suprayogo, D.; Groot, J.C. Complex rice systems to improve rice yield and yield stability in the face of variable weather conditions. Sci. Rep. 2018, 8, 14746. [Google Scholar] [CrossRef]
- Khumairoh, U.; Groot, J.C.; Lantinga, E.A. Complex agro-ecosystems for food security in a changing climate. Ecol. Evol. 2012, 2, 1696–1704. [Google Scholar] [CrossRef]
- Banger, K.; Tian, H.; Lu, C. Do nitrogen fertilizers stimulate or inhibit methane emissions from rice fields? Glob. Chang. Biol. 2012, 18, 3259–3267. [Google Scholar] [CrossRef]
- Brouwer, P.; Schluepmann, H.; Nierop, K.G.; Elderson, J.; Bijl, P.K.; van der Meer, I.; de Visser, W.; Reichart, G.-J.; Smeekens, S.; van der Werf, A. Growing Azolla to produce sustainable protein feed: The effect of differing species and CO2 concentrations on biomass productivity and chemical composition. J. Sci. Food Agric. 2018, 98, 4759–4768. [Google Scholar] [CrossRef] [Green Version]
- Wolf, J.; Ziska, L. Comment on “Unexpected reversal of C3 versus C4 grass response to elevated CO2 during a 20-year field experiment”. Science 2018, 361, 6402. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H. Growth temperature can alter the temperature dependent stimulation of photosynthesis by elevated carbon dioxide in Albutilon theophrasti. Physiol. Plant 2001, 111, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H.; Hogan, K.P.; Smith, A.P.; Drake, B.G. Growth and photosynthetic response of nine tropical species with long-term exposure to elevated carbon dioxide. Oecologia 1991, 86, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Erb, T.J.; Zarzycki, J. A short history of RubisCO: The rise and fall (?) of Nature’s predominant CO2 fixing enzyme. Curr. Opin. Biotechnol. 2018, 49, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.L.; Vasconcelos, V. Classification and phylogeny of the cyanobiont Anabaena azollae Strasburger: An answered question? Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 6, 1830–1840. [Google Scholar] [CrossRef] [Green Version]
- Madan, A.P.; Nierzwicki-Bauer, S.A. In situ detection of transcripts for ribulose-1,5-bisphosphate carboxylase in cyanobacterial heterocysts. J. Bacteriol. 1993, 175, 7301–7306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, T.B.; Mayne, B.C.; Toia, R.E.; Peters, G.A. Azolla-Anabaena Relationship: VIII. Photosynthetic Characterization of the Association and Individual Partners. Plant Physiol. 1979, 64, 791–795. [Google Scholar] [CrossRef] [Green Version]
- Ruan, C.J.; Shao, H.B.; da Silva, J.A.T. A critical review on the improvement of photosynthetic carbon assimilation in C3 plants using genetic engineering. Crit. Rev. Biotechnol. 2012, 32, 1–21. [Google Scholar] [CrossRef]
- Sale, P.J.M.; Orr, P.T.; Shell, G.S.; Erskine, D.J.C. Photosynthesis and growth rates in Salvinia molesta and Eichhornia crassipes. J. Appl. Ecol. 1985, 22, 125–137. [Google Scholar] [CrossRef]
- Park, J.J.; Lechno-Yossef, S.; Wolk, C.P.; Vieille, C. Cell-specific gene expression in Anabaena variabilis grown phototrophically, mixotrophically, and heterotrophically. BMC Genom. 2013, 14, 759. [Google Scholar] [CrossRef] [Green Version]
- Silverman, S. Morphological and Isotopic Changes of Anabaena cylindrica PCC 7122 in Response to N2 Partial Pressure. Bachelor’s Thesis, University of Colorado, Boulder, CO, USA, 2017. Available online: https://scholar.colorado.edu/honr_theses/1444 (accessed on 10 December 2019).
- Ran, L.; Larsson, J.; Vigil-Stenman, T.; Nylander, J.A.; Ininbergs, K.; Zheng, W.W.; Lapidus, A.; Lowry, S.; Haselkorn, R.; Bergman, B. Genome erosion in a nitrogen-fixing vertically transmitted endosymbiotic multicellular cyanobacterium. PLoS ONE 2010, 5, e11486. [Google Scholar] [CrossRef]
- Al-Attar, S.; de Vries, S. An electrogenic nitric oxide reductase. FEBS Lett. 2015, 589, 2050–2057. [Google Scholar] [CrossRef] [Green Version]
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J. Nitric oxide: An effective weapon of the plant or the pathogen? Mol. Plant Pathol. 2014, 15, 406–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, A.L.; Berka, V.; Martin, F.; Ma, X.; van den Akker, F.; Fabian, M.; Olson, J.S. Is Nostoc H-NOX a NO sensor or redox switch? Biochemistry 2010, 49, 6587–6599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persson, T.; Battenberg, K.; Demina, I.V.; Vigil-Stenman, T.; Heuvel, B.V.; Pujic, P.; Facciotti, M.T.; Wilbanks, E.G.; O’Brien, A.; Fournier, P.; et al. Candidatus Frankia Datiscae Dg1, the Actinobacterial Microsymbiont of Datisca glomerata, Expresses the Canonical nod Genes nodABC in Symbiosis with Its Host Plant. PLoS ONE 2015, 10, e0127630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambon, R.; Pradeau, S.; Fort, S.; Cottaz, S.; Armand, S. High yield production of Rhizobium NodB chitin deacetylase and its use for in vitro synthesis of lipo-chitinoligosaccharide precursors. Carbohydr. Res. 2017, 442, 25–30. [Google Scholar] [CrossRef]
- Bizic-Ionescu, M.; Klintzch, T.; Ionescu, D.; Hindiyeh, M.Y.; Guenthel, M.; Pastor, A.M.M.; Keppler, F.; Grossart, H.P. Widespread formation of methane by Cyanobacteria in aquatic and terrestrial environments. BioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xu, H.; Jiang, Y.; Zhang, K.; Hu, Y.; Zeng, Z. Methane Emissions and Microbial Communities as Influenced by Dual Cropping of Azolla along with Early Rice. Sci. Rep. 2017, 7, 40635. [Google Scholar] [CrossRef]
- Zheng, Y.; Harris, D.F.; Yu, Z.; Fu, Y.; Poudel, S.; Ledbetter, R.N.; Fixen, K.R.; Yang, Z.-Y.; Boyd, E.S.; Lidstrom, M.E.; et al. A pathway for biological methane production using bacterial iron-only nitrogenase. Nat. Microbiol. 2018, 3, 281–286. [Google Scholar] [CrossRef]
- Baharum, H.; Chu, W.C.; Teo, S.S.; Ng, K.Y.; Rahim, R.A.; Ho, C.L. Molecular cloning, homology modeling and site-directed mutagenesis of vanadium-dependent bromoperoxidase (GcVBPO1) from Gracilaria changii (Rhodophyta). Phytochemistry 2013, 92, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Nelson, J.M.; Hauser, D.A.; Gudiño, J.A.; Guadalupe, Y.A.; Meeks, J.C.; Allen, N.S.; Villarreal, J.C.; Li, F.W. Complete Genomes of Symbiotic Cyanobacteria Clarify the Evolution of Vanadium-Nitrogenase. Genome Biol. Evol. 2019, 11, 1959–1964. [Google Scholar] [CrossRef]
- Pushpakumar, B.L.D.U.; Gunawardana, D. Preliminary data on the presence of an alternate vanadium nitrogenase in a culturable cyanobiont of Azolla pinnata R. Brown: Implications on Chronic Kidney Disease of an unknown etiology (CKDu). Data Brief 2018, 21, 2590–2597. [Google Scholar] [CrossRef]
- McLauchlan, C.C.; Murakami, H.A.; Wallace, C.A.; Crans, D.C. Coordination environment changes of the vanadium-dependent haloperoxidase enzymes. J. Inorg. Biochem. 2018, 186, 267–279. [Google Scholar] [CrossRef]
- Neville, L.A.; Grasby, S.E.; McNeil, D.H. Limited freshwater cap in the Eocene Arctic Ocean. Sci. Rep. 2019, 9, 4226. [Google Scholar] [CrossRef] [Green Version]
- Lo Conte, L.; Ailey, B.; Hubbard, T.J.; Brenner, S.E.; Murzin, A.G.; Chothia, C. SCOP: A structural classification of proteins database. Nucleic Acids Res. 2000, 28, 257–259. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Jones, D.T. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Linding, R.; Russell, R.B.; Neduva, V.; Gibson, T.J. GlobPlot: Exploring protein sequences for globularity and disorder. Nucleic Acids Res. 2003, 31, 3701–3708. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Heterocysts (H) of Nostoc azollae, where they are shown to have subtle differences in size, shape, and the smoothness/roughness of contours, and are spaced as close as four cells apart. Heterocysts are a major site of cellular respiration.
Figure 1.
Heterocysts (H) of Nostoc azollae, where they are shown to have subtle differences in size, shape, and the smoothness/roughness of contours, and are spaced as close as four cells apart. Heterocysts are a major site of cellular respiration.

Figure 2.
(A) The equation for the enzymatic conversion of nitric oxide into nitrous oxide. (B) The equation for the enzymatic conversion of nitrous oxide into elemental dinitrogen gas.
Figure 2.
(A) The equation for the enzymatic conversion of nitric oxide into nitrous oxide. (B) The equation for the enzymatic conversion of nitrous oxide into elemental dinitrogen gas.

Figure 3.
The downloaded amino acid sequences of NodB nodulation proteins that were aligned with the ClustalW algorithm using MEGA version X and the phylogenetic reconstruction performed using the neighborhood joining method with support from 500 bootstrap replications. NodB nodulation proteins are found in symbiotic cyanobacteria (namely, Nostoc), but are also available in noncyanobionts
Figure 3.
The downloaded amino acid sequences of NodB nodulation proteins that were aligned with the ClustalW algorithm using MEGA version X and the phylogenetic reconstruction performed using the neighborhood joining method with support from 500 bootstrap replications. NodB nodulation proteins are found in symbiotic cyanobacteria (namely, Nostoc), but are also available in noncyanobionts

Figure 4.
(a): ClustalW sequence alignment of four NodB deacetylase proteins: [>WP_131767784.1 chitooligosaccharide deacetylase NodB [Frankia symbiont of Datisca glomerata]; [>AFJ42532.1 NodB nodulation protein [M. plurifarium]; [>RCJ39328.1 polysaccharide deacetylase [Nostoc punctiforme NIES-2108]; [>ADI63120.1 polysaccharide deacetylase [Nostoc azollae 0708]. Note: Shown in green are the 3/5 motifs that form key sites for enzymatic activity. Yellow indicates the interruptions (deletions) in the NodB domain in sequences from cyanobacteria. (b) Prediction of the Globular Domain of sequence AFJ42532.1 (NodB nodulation protein from M. plurifarium) using http://globplot.embl.de/. The amino acids in green indicate the globular domain, which contains the majority of the catalytic site. (c) Output of the secondary structure prediction of sequence AFJ42532.1 (NodB nodulation protein from M. plurifarium) using the service PSIPRED 4.0, showcasing helixes, beta strands, and coils.
Figure 4.
(a): ClustalW sequence alignment of four NodB deacetylase proteins: [>WP_131767784.1 chitooligosaccharide deacetylase NodB [Frankia symbiont of Datisca glomerata]; [>AFJ42532.1 NodB nodulation protein [M. plurifarium]; [>RCJ39328.1 polysaccharide deacetylase [Nostoc punctiforme NIES-2108]; [>ADI63120.1 polysaccharide deacetylase [Nostoc azollae 0708]. Note: Shown in green are the 3/5 motifs that form key sites for enzymatic activity. Yellow indicates the interruptions (deletions) in the NodB domain in sequences from cyanobacteria. (b) Prediction of the Globular Domain of sequence AFJ42532.1 (NodB nodulation protein from M. plurifarium) using http://globplot.embl.de/. The amino acids in green indicate the globular domain, which contains the majority of the catalytic site. (c) Output of the secondary structure prediction of sequence AFJ42532.1 (NodB nodulation protein from M. plurifarium) using the service PSIPRED 4.0, showcasing helixes, beta strands, and coils.


Figure 5.
The downloaded amino acid sequences of the methane production catalyzing proteins that were aligned with the ClustalW algorithm using MEGA version X and the phylogenetic reconstruction performed using the maximum parsimony method with support from 500 bootstrap replications. There was no assignment of outgroups. The majority cyanobionts cluster (mixed with noncyanobionts) is shown in a bracket.
Figure 5.
The downloaded amino acid sequences of the methane production catalyzing proteins that were aligned with the ClustalW algorithm using MEGA version X and the phylogenetic reconstruction performed using the maximum parsimony method with support from 500 bootstrap replications. There was no assignment of outgroups. The majority cyanobionts cluster (mixed with noncyanobionts) is shown in a bracket.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The spacing of heterocysts in cyanobacteria of different pedigrees. N. punctiforme is a horizontally transferred cyanobiont, Anabaena variabilis possesses a two-prong nitrogenase system (inclusive of a vanadium nitrogenase), and Anabaena cylindrica PCC 7122 is a free-living cyanobacterium. The following were obtained from [18,19] and this study.
Table 1.
The spacing of heterocysts in cyanobacteria of different pedigrees. N. punctiforme is a horizontally transferred cyanobiont, Anabaena variabilis possesses a two-prong nitrogenase system (inclusive of a vanadium nitrogenase), and Anabaena cylindrica PCC 7122 is a free-living cyanobacterium. The following were obtained from [18,19] and this study.
| Cell Type | Organism | Spacing/Percentage |
|---|---|---|
| Heterocyst | N. azollae | 2–10 cells apart |
| Heterocyst | N. punctiforme | 3–10% of the total cells |
| Heterocyst | A. cylindrica PCC 7122 | 8–15 cells apart |
| Heterocyst | A. variabilis | 5–10% of cells |
Table 2.
Query and the PSI-BLAST query employed in this study.
| Word Query | PSI-BLAST Query | Type of Cyanobacteria/Bacteria |
|---|---|---|
| Nitric Oxide reductase | AFZ55274.1, belonging to Cyanobacterium aponinum PCC 10605 | Freshwater, Unicellular |
| Nitrous Oxide reductase | RCJ21339.1, belonging to Nostoc sp. ATCC 43529 | Subsection IV, Filamentous Cyanobacteria |
| Methanogenic enzymes | CEN44_02070 (Fischerella muscicola CCMEE 5323) | Subsection V, Filamentous Cyanobacteria |
| NodB protein | CAA67138.1—NodB polysaccharide deacetylase (Rhizobium leucaenae USDA 9039) | Bacterial, Nodular |
| NodC protein | CAA67139.1—N-acetylglucosaminyltransferases (R. leucaenae USDA 9039) | Bacterial, Nodular |
| Chloroperoxidases | CAA04998.1—Nostoc sp. PCC 7120 | Subsection IV, Filamentous |
| Bromoperoxidases | GBG19525.1—Nostoc commune NIES-4072 | Subsection IV, Filamentous |
Table 3.
The enzymes that were employed for bioinformatics explorations and their occurrences/distribution. (Yes—sequence identity is >30% and sequence coverage of >30%; No—identity is <30% or sequence coverage <30%).
Table 3.
The enzymes that were employed for bioinformatics explorations and their occurrences/distribution. (Yes—sequence identity is >30% and sequence coverage of >30%; No—identity is <30% or sequence coverage <30%).
| Protein | Key Enzyme-Derived Product | Subsection V Cyanobacteria | Subsection IV Cyanobacteria | Azolla Major Cyanobiont |
|---|---|---|---|---|
| Nitrite reductase | Nitric Oxide | YES | YES | NO |
| Nitric Oxide (NO) synthase | Nitric Oxide | NO | YES | NO |
| Nitric Oxide (NO) reductase | Nitrous Oxide | YES | NO | NO |
| Nitrous Oxide (N2O) reductase | Nitrogen gas | YES | YES | YES |
| Methanogenic enzyme | Methane | YES | YES | NO |
| Vanadium dependent Chloroperoxidase | Halocarbons | YES | YES | YES |
| Vanadium dependent Bromoperoxidase | Halocarbons | YES | YES | NO |
Table 4.
Search results of Nod-factor-producing/modifying proteins in all cyanobacteria and in the genus Nostoc, using “Candidatus Frankia californiensis” Nod-factor-related sequences as queries.
Table 4.
Search results of Nod-factor-producing/modifying proteins in all cyanobacteria and in the genus Nostoc, using “Candidatus Frankia californiensis” Nod-factor-related sequences as queries.
| Protein | Best Match in Cyanobacteria /Coverage/Sequence Identity | Best Match in Nostoc/Coverage/Sequence Identity |
|---|---|---|
| Nodulation protein A | RsmB/NOP family class I SAM-dependent RNA methyltransferase [Scytonema tolypothrichoides]; 41%; 24.69% | transcriptional regulator [Nostoc sp. MBR 210]; 40%; 26.25% |
| Nodulation protein H | type I polyketide synthase [Calothrix brevissima]; 22%; 31.75% | glycosyl transferase [Nostoc sp. ATCC 53789]; 17%; 34.09% |
| NodB polysaccharide deacetylases | Polysaccharide Deacetylase family protein [Fischerella sp. PCC 9605]; 86%; 35.8% | Deacteylase NodB [Nostoc sp. 3335mG]; 81%; 37.44% |
Table 5.
Search results of Nod-factor-producing/modifying proteins in the Azolla cyanobiont’s proteome using the relevant proteins from R. leucaenae USDA 9039 as the query proteins.
Table 5.
Search results of Nod-factor-producing/modifying proteins in the Azolla cyanobiont’s proteome using the relevant proteins from R. leucaenae USDA 9039 as the query proteins.
| Azolla Cyanobiont’s Protein Type | Sequence Identity to the Homolog in R. leucaenae USDA 9039 | Coverage |
|---|---|---|
| NodB polysaccharide deacetylase | 35% | 90% |
| NodC acetylglucosaminyltransferase | 34.7% | 22% |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gunawardana, D. An Exploration of Common Greenhouse Gas Emissions by the Cyanobiont of the Azolla–Nostoc Symbiosis and Clues as to Nod Factors in Cyanobacteria. Plants 2019, 8, 587. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120587
AMA Style
Gunawardana D. An Exploration of Common Greenhouse Gas Emissions by the Cyanobiont of the Azolla–Nostoc Symbiosis and Clues as to Nod Factors in Cyanobacteria. Plants. 2019; 8(12):587. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120587
Chicago/Turabian StyleGunawardana, Dilantha. 2019. "An Exploration of Common Greenhouse Gas Emissions by the Cyanobiont of the Azolla–Nostoc Symbiosis and Clues as to Nod Factors in Cyanobacteria" Plants 8, no. 12: 587. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120587
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.