Alpha Glucosidase Inhibitory Activities of Plants with Focus on Common Vegetables

 ,
,  ,
,
Abstract
:1. Introduction
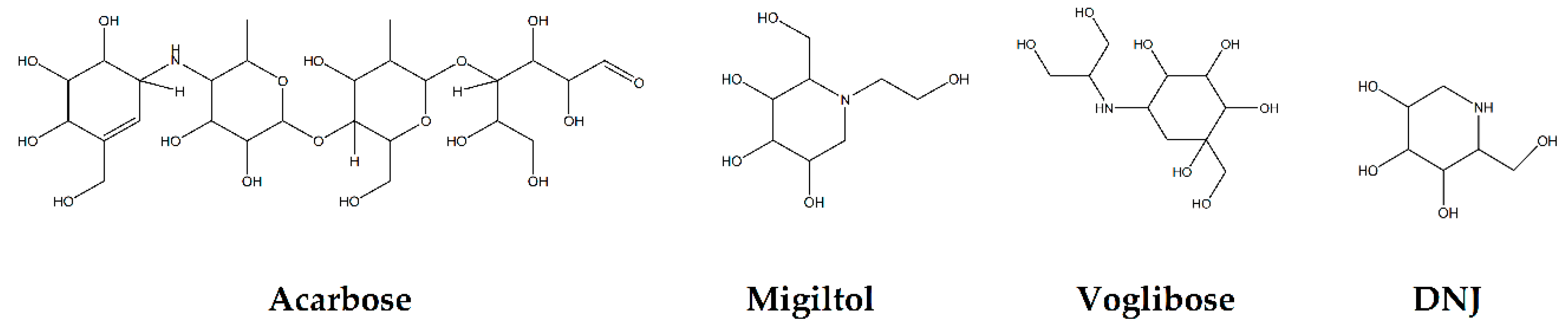
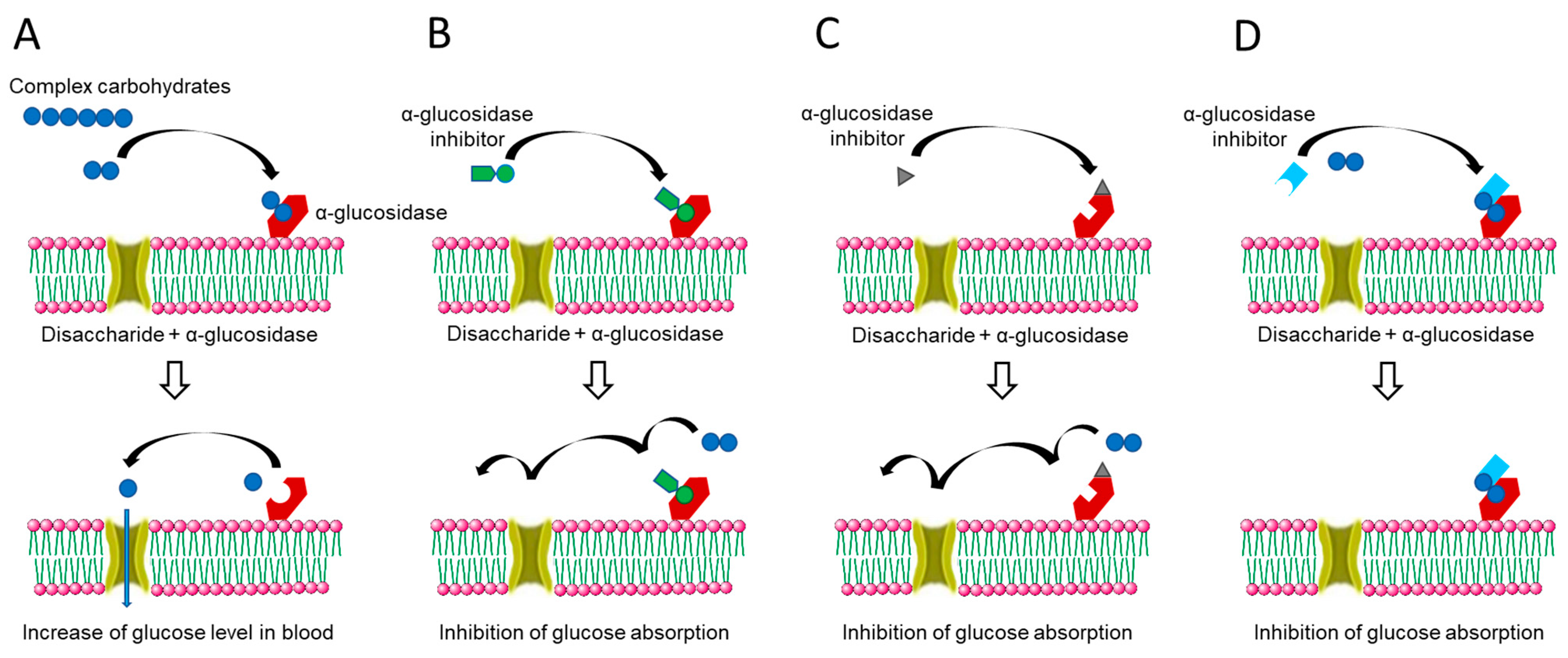
2. Natural Compounds as Potential Candidates for AGIs
3. Plants with AG Inhibitory Activity
4. Common Vegetables with AG Inhibitory Activities
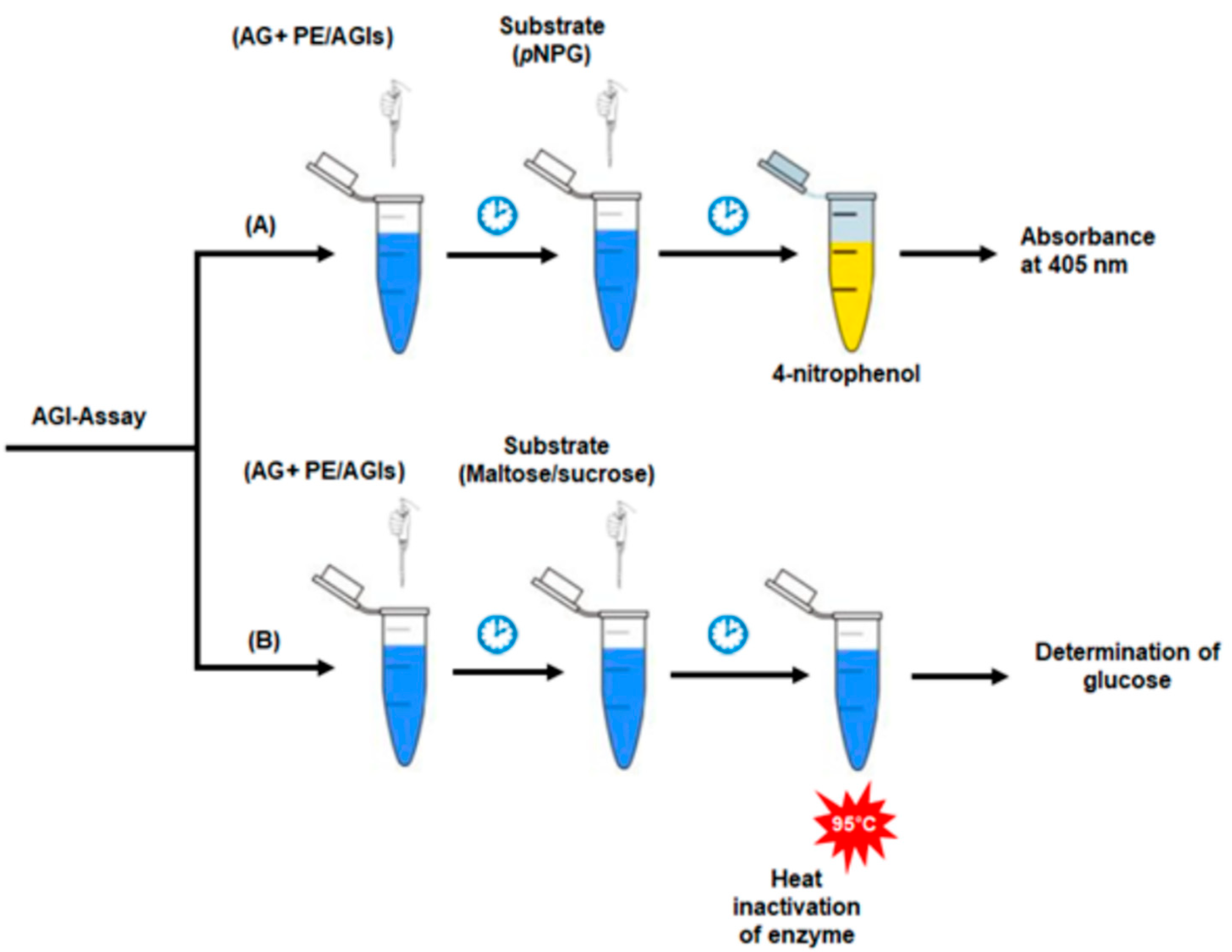
5. Analyses of AGI Activities in Plants
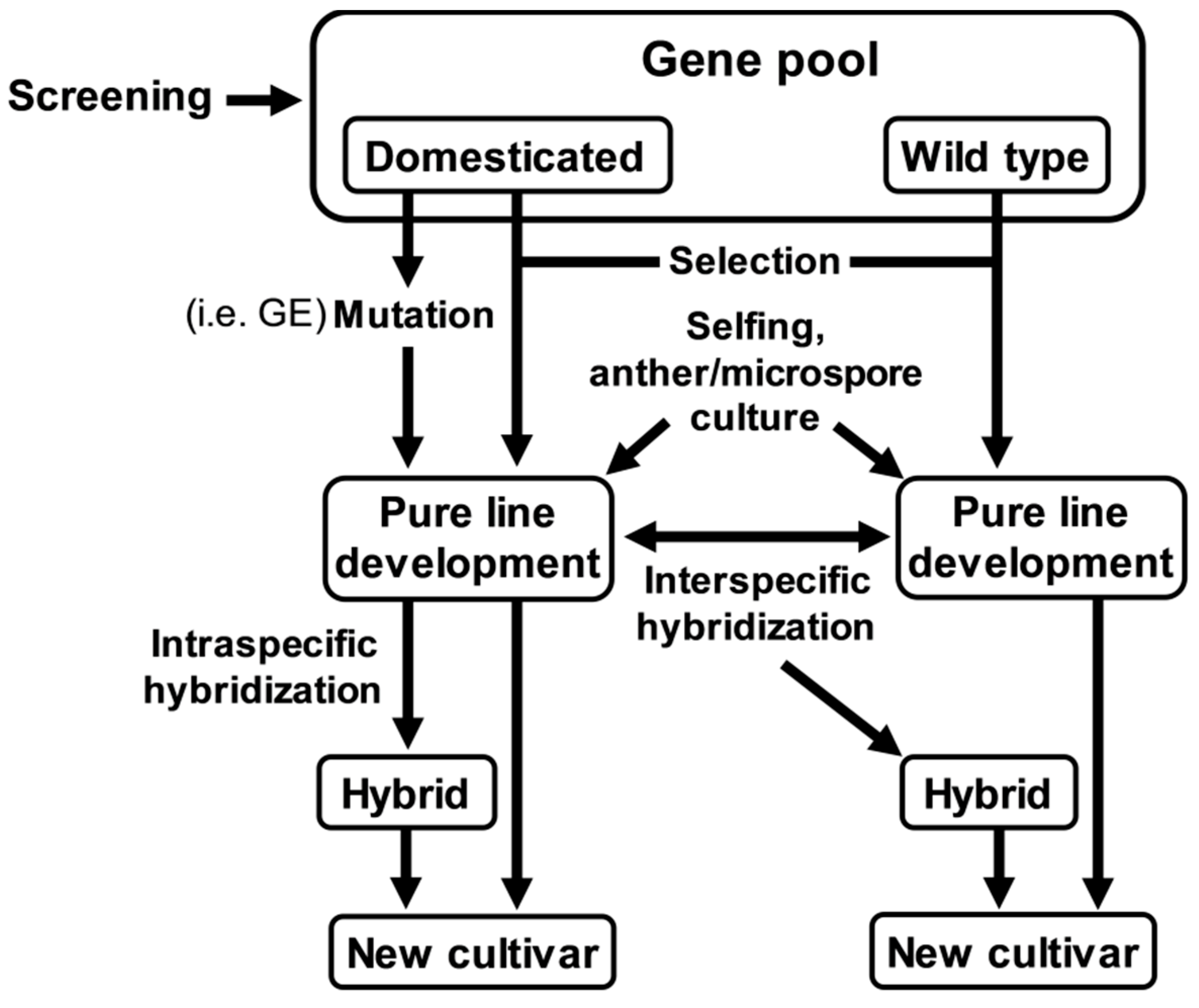
6. Production of Vegetables with Higher AGI Activity
7. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davies, G.J.; Gloster, T.M.; Henrissat, B. Recent structural insights into the expanding world of carbohydrate-active enzymes. Curr. Opin. Struct. Biol. 2005, 15, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Vocadlo, D.J.; Davies, G.J. Mechanistic insights into glycosidase chemistry. Curr. Opin. Chem. Biol. 2008, 12, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Kajimoto, T.; Node, M. Inhibitors against glycosidases as medicines. Curr. Top. Med. Chem. 2009, 9, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Stutz, A.E.; Wrodnigg, T.M. Imino sugars and glycosyl hydrolases: Historical context, current aspects, emerging trends. Adv. Carbohydr. Chem. Biochem. 2011, 66, 187–298. [Google Scholar] [PubMed]
- Bischoff, H. Pharmacology of alpha-glucosidase inhibition. Eur. J. Clin. Investig. 1994, 24, 3–10. [Google Scholar]
- Chiba, S. Molecular mechanism in alpha-glucosidase and glucoamylase. Biosci. Biotechnol. Biochem. 1997, 61, 1233–1239. [Google Scholar] [CrossRef]
- Meneilly, G.S.; Ryan, E.A.; Radziuk, J.; Lau, D.C.; Yale, J.F.; Morais, J.; Chiasson, J.L.; Rabasa-Lhoret, R.; Maheux, P.; Tessier, D.; et al. Effect of acarbose on insulin sensitivity in elderly patients with diabetes. Diabetes Care 2000, 23, 1162. [Google Scholar] [CrossRef] [Green Version]
- Truscheit, E.; Hillebrand, I.; Junge, B.; Müller, L.; Puls, W.; Schmidt, D. Microbial α-Glucosidase Inhibitors: Chemistry, Biochemistry, and Therapeutic Potential. In Drug Concentration Monitoring Microbial Alpha-Glucosidase Inhibitors Plasminogen Activators; Progress in Clinical Biochemistry and Medicine; Springer: Berlin/Heidelberg, Germany, 1988; Volume 7. [Google Scholar]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.T.; Sousa, J.L.C.; Tomé, S.M.; Ramos, M.J.; Silva, A.M.S.; Fernandes, P.A.; Fernandes, E. α-Glucosidase inhibition by flavonoids: An in vitro and in silico structure–activity relationship study. J. Enzym. Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef] [Green Version]
- Sugihara, H.; Nagao, M.; Harada, T.; Nakajima, Y.; Tanimura-Inagaki, K.; Okajima, F.; Tamura, H.; Inazawa, T.; Otonari, T.; Kawakami, M.; et al. Comparison of three α-glucosidase inhibitors for glycemic control and bodyweight reduction in Japanese patients with obese type 2 diabetes. J. Diabetes Investig. 2014, 5, 206–212. [Google Scholar] [CrossRef]
- Adefegha, S.A.; Oboh, G. Inhibition of key enzymes linked to type 2 diabetes and sodium nitroprusside-induced lipid peroxidation in rat pancreas by water extractable phytochemicals from some tropical spices. Pharm. Biol. 2012, 50, 857–865. [Google Scholar] [CrossRef] [Green Version]
- Udenigwe, C.C.; Adebiyi, A.P.; Doyen, A.; Bazinet, L.; Aluko, R.E. Low molecular weight flaxseed protein-derived arginine-containing peptides reduced blood pressure of spontaneously hypertensive rats faster than amino acid form of arginine and native flaxseed protein. Food Chem. 2012, 132, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Spiller, H.A.; Sawyer, T.S. Toxicology of oral antidiabetic medications. Am. J. Health Syst. Pharm. 2006, 63, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ma, S. Recent advances in synthetic α-glucosidase inhibitors. Chem. Med. Chem. 2017, 12, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Girdhar, S.; Girdhar, A.; Verma, S.K.; Lather, V.; Pandita, D. Plant derived alkaloids in major neurodegenerative diseases: From animal models to clinical trials. J. Ayurvedic Herb. Med. 2015, 1, 91–100. [Google Scholar]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An Overview of Plant Phenolic Compounds and Their Importance in Human Nutrition and Management of Type 2 Diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef] [PubMed]
- Falcone-Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [Green Version]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar]
- Roskar, I.; Molek, P.; Vodnik, M.; Stempelj, M.; Strukelj, B.; Lunder, M. Peptide modulators of alpha-glucosidase. J. Diabetes Investig. 2015, 6, 625–631. [Google Scholar] [CrossRef]
- Moorthy, N.S.; Ramos, M.J.; Fernandes, P.A. Studies on alpha-glucosidase inhibitors development: Magic molecules for the treatment of carbohydrate mediated diseases. Mini Rev. Med. Chem. 2012, 12, 713–720. [Google Scholar] [CrossRef]
- Zechel, D.L.; Boraston, A.B.; Gloster, T.; Boraston, C.M.; Macdonald, J.M.; Tilbrook, D.M.; Stick, R.V.; Davies, G.J. Iminosugar glycosidase inhibitors: Structural and thermodynamic dissection of the binding of isofagomine and 1-deoxynojirimycin to beta-glucosidases. J. Am. Chem. Soc. 2003, 125, 14313–14323. [Google Scholar] [CrossRef]
- Wang, R.W.; Qiu, X.L.; Bols, M.; Caballero, F.O.; Qing, F.L. Synthesis and biological evaluation of glycosidase inhibitors: Gem-Difluoromethylenated Nojirimycin analogues. J. Med. Chem. 2006, 49, 2989–2997. [Google Scholar] [CrossRef] [PubMed]
- Stiitz, E.A. Iminosugars as Glycosidase Inhibitors Nojirimycin and Beyond; Wiley-VCH Verlag GmbH: Weinheim, Germany, 1999. [Google Scholar]
- Gao, H.; Huang, Y.N.; Gao, B.; Li, P.; Inagaki, C.; Kawabata, J. Inhibitory effect on α-glucosidase by Adhatoda vasica Nees. Food Chem. 2008, 108, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Tabopda, T.K.; Ngoupayo, J.; Liu, J.; Mitaine-Offer, A.C.; Tanoli, S.A.; Khan, S.N.; Ali, M.S.; Ngadjui, B.T.; Tsamo, E.; Lacaille-Dubois, M.A.; et al. Bioactive aristolactams from Piper umbellatum. Phytochemistry 2008, 69, 1726–1731. [Google Scholar] [CrossRef] [PubMed]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of alpha-glucosidase and alpha-amylase by flavonoids. J. Nutr. Sci. Vitaminol. (Tokyo) 2006, 52, 149–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Narwal, S.; Kumar, V.; Prakash, O. Alpha-glucosidase inhibitors from plants: A natural approach to treat diabetes. Pharmacogn. Rev. 2011, 5, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, J.S.; Lauridsen, M.B.; Dragsted, L.O.; Nielsen, J.; Staerk, D. Development of a bioassay-coupled HPLC-SPE-ttNMR platform for identification of α-glucosidase inhibitors in apple peel (Malus x domestica Borkh.). Food Chem. 2012, 135, 1692–1699. [Google Scholar] [CrossRef]
- Thanakosaia, W.; Phuwapraisirisanb, P. First identification of α-glucosidase inhibitors from Okra (Abelmoschus esculentus) Seeds. Nat. Prod. Commun. 2013, 8, 1085–1088. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, J.; Hogan, S.; Chung, H.; Welbaum, G.E.; Zhou, K. Inhibitory effect of raspberries on starch digestive enzyme and their antioxidant properties and phenolic composition. Food Chem. 2010, 119, 592–599. [Google Scholar] [CrossRef]
- Chu, Y.H.; Wu, S.H.; Hsieh, J.F. Isolation and characterization of α-glucosidase inhibitory constituents from Rhodiola crenulata. Food Res. Int. 2014, 57, 8–14. [Google Scholar] [CrossRef]
- Wang, Z.B.; Jiang, H.; Xia, Y.G.; Yang, B.Y.; Kuang, H.X. Alpha-glucosidase inhibitory constituents from Acanthopanax senticosus harm leaves. Molecules 2012, 17, 6269–6276. [Google Scholar] [CrossRef]
- Kubínová, B.R.; Pořízková, R.; Navrátilová, A.; Farsa, O.; Hanáková, Z.; Bačinská, A.; Čížek, A.; Valentová, M. Antimicrobial and enzyme inhibitory activities of the constituents of Plectranthus madagascariensis (Pers). J. Enzym. Inhib. Med. Chem. 2014, 29, 749–752. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Ashok, P.; Tiwari, K.; Rao, R.R.; Madhusudhana, K.; Rao, V.R.; Ali, A.Z.; Babu, K.S.; Rao, J.M. New Labdane diterpenes as intestinal α-glucosidase inhibitor from anti hyperglycemic extract of Hedychium spicatum (Ham. Ex Smith) rhizomes. Bioorg. Med. Chem. Lett. 2009, 19, 2562–2565. [Google Scholar] [CrossRef] [PubMed]
- Ramu, R.; Shirahatti, P.S.; Zameer, F.; Ranganatha, L.V.; Prasad, M.N. Inhibitory effect of banana (Musa sp. var. Nanjangud rasa bale) flower extract and its constituents Umbelliferone and Lupeol on α-glucosidase, aldose reductase and glycation at multiple stages. S. Afr. J. Bot. 2014, 95, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Nile, S.H.; Park, S.W. Antioxidant, α-glucosidase and xanthine oxidase inhibitory activity of bioactive compounds from maize (Zea mays L.). Chem. Biol. Drug Des. 2014, 83, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tu, Z.; Yuan, T.; Wang, H.; Xie, X.; Fu, Z. Antioxidants and α-glucosidase inhibitors from Ipomoea batatas leaves identified by bioassay-guided approach and structure-activity relationships. Food Chem. 2016, 208, 61–67. [Google Scholar] [CrossRef]
- Zhang, S.S.; Wu, D.; Li, H.; Zhu, J.; Hu, W.; Lua, M.; Liu, X. Rapid identification of α-glucosidase inhibitors from Dioscorea opposita Thunb peel extract by enzyme functionalized Fe3O4 magnetic nanoparticles coupled with HPLC-MS/MS. Food Funct. 2017, 8, 3219–3227. [Google Scholar] [CrossRef]
- Jeong, E.Y.; Cho, K.S.; Lee, H.S. α-Amylase and α-Glucosidase Inhibitors Isolated from Triticum aestivum L. Sprouts. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 47–51. [Google Scholar] [CrossRef]
- Alonzi, D.S.; Scott, K.A.; Dwek, R.A.; Zitzmann, N. Iminosugar antivirals: The therapeutic sweet spot. Biochem. Soc. Trans. 2017, 45, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Zhang, W.; Feng, F.; Zhang, Y.; Kang, W. α-Glucosidase inhibitors isolated from medicinal plants. Food Sci. Hum. Wellness 2014, 3, 136–174. [Google Scholar] [CrossRef] [Green Version]
- Kawada, Y.; Miura, M.; Gomyo, T. Inhibitory Effect of Vegetables, Fruits and Herbs on α-Glucosidase in an Immobilized Enzyme Assay System. Food Sci. Technol. Res. 2006, 12, 275–277. [Google Scholar] [CrossRef] [Green Version]
- Oe, H.; Ozaki, S. Hypoglycemic Effect of 13-Membered Ring Thiocyclitol, a Novel α-Glucosidase Inhibitor from Kothala-himbutu (Salacia reticulata). Biosci. Biotechnol. Biochem. 2008, 72, 1962–1964. [Google Scholar] [CrossRef] [PubMed]
- Wafaa, B.; Said, B.; Mohamed, B. Antidiabetic medicinal plants as a source of alpha glucosidase inhibitors. Curr. Diabetes Rev. 2010, 6, 247–254. [Google Scholar]
- Hwang, S.H.; Li, H.M.; Lim, S.S.; Wang, Z.; Hong, J.S.; Huang, B. Evaluation of a standardized extract from Morus alba against α-glucosidase inhibitory effect and postprandial anti hyperglycemic in patients with impaired glucose tolerance: A randomized double-blind clinical trial. Evid. Based Complement. Altern. Med. 2016, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Yi, X.; Wang, Y.; Chen, G.; He, X. Benzophenones from Mango Leaves Exhibit α-Glucosidase and NO Inhibitory Activities. J. Agric. Food Chem. 2016, 64, 7475–7480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tu, Z.; Xie, X.; Wang, H.; Wang, H.; Wang, Z.; Sha, X.; Lua, Y. Jackfruit (Artocarpus heterophyllus Lam.) peel: A better source of antioxidants and α-glucosidase inhibitors than pulp, flake and seed, and phytochemical profile by HPLC-QTOF-MS/MS. Food Chem. 2017, 234, 303–313. [Google Scholar] [CrossRef]
- Musa, M.Y.; Griffith, A.M.; Michels, A.J.; Schneider, E.; Frei, B. Grape seed and tea extracts and catechin 3-gallates are potent inhibitors of α-amylase and α-glucosidase activity. J. Agric. Food Chem. 2012, 60, 8924–8929. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.I.; Apostolidis, E.; Shetty, K. Evaluation of pepper (Capsicum annuum) for management of diabetes and hypertension. J. Food Biochem. 2007, 31, 370–385. [Google Scholar] [CrossRef]
- Kim, Y.C.; Choi, D.; Lee, J.H.; Lee, S. Alpha-glucosidase inhibitory activity in different pepper cultivars (Capsicum annuum L.). Hortic. Sci. Technol. 2018, 36, 444–450. [Google Scholar]
- Park, M.S.; Zhu, Y.X.; Pae, H.O.; Park, S.H. In vitro and in vivo α-glucosidase and α-amylase inhibitory effects of the water extract of leaves of pepper (Capsicum annuum L. Cultivar Dangjo) and the active constituent luteolin 7-O-glucoside. J. Food Biochem. 2016, 40, 696–703. [Google Scholar] [CrossRef]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of α-glucosidase, α-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef]
- Nwanna, E.E.; Ibukun, E.O.; Oboh, G. Inhibitory effects of methanolic extracts of two eggplant species from South-western Nigeria on starch hydrolysing enzymes linked to type-2 diabetes. Afr. J. Pharm. Pharmacol. 2013, 7, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo-González, M.; Valentão, P.; Andrade, P.B. Tomato plant leaves: From by-products to the management ofenzymes in chronic diseases. Ind. Crops Prod. 2016, 94, 621–629. [Google Scholar] [CrossRef]
- Wu, H.; Xu, B. Inhibitory Effects of Onion against α-Glucosidase Activity and its Correlation with Phenolic Antioxidants. Int. J. Food Prop. 2014, 17, 599–609. [Google Scholar] [CrossRef]
- Kongstad, K.T.; Özdemir, C.; Barzak, A.; Wubshet, S.G.; Staerk, D. Combined use of high-resolution α-glucosidase inhibition profiling and high-performance liquid chromatography−high-resolution mass spectrometry−solid-phase extraction−nuclear magnetic resonance spectroscopy for investigation of antidiabetic principles in crude plant extracts. J. Agric. Food Chem. 2015, 63, 2257–2263. [Google Scholar] [PubMed]
- Matsui, T.; Ueda, T.; Oki, T.; Sugita, K.; Terahara, N.; Matsumoto, K. Alpha-glucosidase inhibitory action of natural acylated anthocyanins. 1. Survey of natural pigment with potent inhibitory activity. J. Agric. Food Chem. 2001, 49, 1948–1951. [Google Scholar] [CrossRef] [PubMed]
- Podsędek, A.; Majewska, I.; Kucharska, A.Z. Inhibitory potential of red cabbage against digestive enzymes linked to obesity and type 2 diabetes. J. Agric. Food Chem. 2017, 65, 7192–7199. [Google Scholar] [CrossRef]
- Baenas, N.; Piegholdt, S.; Schloesser, A.; Moreno, D.A.; Viguera, C.G.; Rimbach, G.; Wagner, A.E. Metabolic activity of radish sprouts derived isothiocyanates in Drosophila melanogaster. Int. J. Mol. Sci. 2016, 17, 251. [Google Scholar] [CrossRef] [Green Version]
- Gopal, S.S.; Lakshmi, M.J.; Sharavana, G.; Sathaiah, G.; Sreeramac, Y.N.; Baskaran, V. Lactucaxanthin—A potential anti-diabetic carotenoid from lettuce (Lactuca sativa) inhibits α-amylase and α-glucosidase activity in vitro and in diabetic rats. Food Funct. 2017, 8, 1124–1131. [Google Scholar] [CrossRef]
- Poovitha, S.; Parani, M. In vitro and in vivo α-amylase and α- glucosidase inhibiting activities of the protein extracts from two varieties of bitter gourd (Momordica charantia L.). BMC Complement. Altern. Med. 2016, 16 (Suppl. 1), 185. [Google Scholar] [CrossRef] [Green Version]
- Uebanso, T.; Arai, H.; Taketani, Y.; Fukaya, M.; Yamamoto, H.; Mizuno, A.; Uryu, K.; Hada, T.; Takeda, E. Extracts of Momordica charantia suppress postprandial hyperglycemia in rats. J. Nutr. Sci. Vitaminol. 2007, 53, 482–488. [Google Scholar] [CrossRef] [Green Version]
- Sulaiman, S.F.; Ooi, K.L. Antioxidant and α-glucosidase inhibitory activities of cucurbit fruit vegetables and identification of active and major constituents from phenolic-rich extracts of Lagenaria siceraria and Sechium edule. J. Agric. Food Chem. 2013, 61, 10080–10090. [Google Scholar] [CrossRef] [PubMed]
- Jibril, M.M.; Hamid, A.A.; Ghazali, H.M.; Dek, M.S.P.; Ramli, N.S.; Jaafar, A.H.; Karrupan, J.; Mohammed, A.S. Antidiabetic antioxidant and phytochemical profile of yellow-fleshed seeded watermelon (Citrullus Lanatus) extracts. J. Food Nut. Res. 2019, 7, 82–95. [Google Scholar]
- Sabitha, V.; Panneerselvam, K.; Ramachandran, S. In vitro α-glucosidase and α-amylase enzyme inhibitory effects in aqueous extracts of Abelmoscus esculentus (L.) Moench. Asian Pac. J. Trop. Biomed. 2012, 2, S162–S164. [Google Scholar] [CrossRef]
- Kim, Y.M.; Wang, M.H.; Rhee, H.I. A novel alpha-glucosidase inhibitor from pine bark. Carbohydr. Res. 2004, 339, 715–717. [Google Scholar] [CrossRef]
- Oki, T.; Matusi, T.; Osajima, Y. Inhibitory effect of alpha glucosidase inhibitors varies according to its origin. J. Agric. Food Chem. 1999, 47, 550–553. [Google Scholar] [CrossRef]
- Malunga, L.N.; Eck, P.; Beta, T. Inhibition of intestinal α-glucosidase and glucose absorption by feruloylated arabinoxylan Mono and oligosaccharides from corn bran and wheat aleurone. J. Nutr. Metab. 2016, 1932532. [Google Scholar] [CrossRef] [Green Version]
- Pyner, A.; Nyambe-silavwe, H.; Williamson, G. Inhibition of human and rat sucrase and maltase activities to assess antiglycemic potential: Optimization of the assay using acarbose and polyphenols. J. Agric. Food Chem. 2017, 65, 8643–8651. [Google Scholar] [CrossRef]
- Zhang, H.C.; Wang, G.; Beta, T.; Dong, J. Inhibitory properties of aqueous ethanol extracts of propolis on alpha-glucosidase. Evid. Based Complement. Altern. Med. 2015, 587383. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhao, S.; Yin, P.; Yan, L.; Han, J.; Shi, L.; Zhou, X.; Liu, Y.; Ma, C. α-Glucosidase inhibitory activity of polyphenols from the burs of Castanea mollissima blume. Molecules 2014, 19, 8373–8386. [Google Scholar] [CrossRef] [Green Version]
- Copeland, R.A. Mechanistic considerations in high-throughput screening. Anal. Biochem. 2003, 320, 1–12. [Google Scholar] [CrossRef]
- Acker, M.G.; Auld, D.S. Consideration for the design and reporting of enzyme assay in high-throughput screening applications. Perspect. Sci. 2014, 1, 56–73. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.C.; Yang, E.Y.; Rhee, H.I.; Chae, Y.S.; Jeong, H.B.; Chae, Y.; Pae, H.D.; Ko, D.K. Breeding of Functional pepper (Capsicum annuum) Variety ‘Wongi No. 1’. Kor. J. Hort. Sci. Technol. 2009, 5, 82–83. [Google Scholar]
- Kaushik, P.; Andújar, I.; Vilanova, S.; Plazas, M.; Gramazio, P.; Herraiz, F.J.; Brar, N.S.; Prohens, J. Breeding Vegetables with increased content in bioactive phenolic acids. Molecules 2015, 20, 18464–18481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plazas, M.; Prohens, J.; Cuñat, N.A.; Vilanova, S.; Gramazio, P.; Herraiz, J.F.; Andújar, I. Reducing capacity, chlorogenic acid content and biological activity in a collection of scarlet (Solanum aethiopicum) and gboma (S. macrocarpon) eggplants. Int. J. Mol. Sci. 2014, 15, 17221–17241. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.R.; Cho, M.C.; Lee, J.G. Genotypic variation in carotenoid, ascorbic acid, total phenolic, and flavonoid contents, and antioxidant activity in selected tomato breeding lines. Hortic. Environ. Biotechnol. 2016, 57, 440–452. [Google Scholar] [CrossRef]
- Gurnani, N.; Gupta, M.; Mehta, D.; Mehta, B.K. Chemical composition, total phenolic and flavonoid contents, and in vitro antimicrobial and antioxidant activities of crude extracts from red chilli seeds (Capsicum frutescens L.). J. Taibah Univ. Sci. 2016, 10, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Assefa, A.D.; Jeong, Y.J.; Kim, D.J.; Jeon, Y.A.; Lee, J.R.; Ko, H.C.; Baek, H.J.; Sung, J.S. Assessing phenolic content and antioxidant potential diversity in Allium plants using multivariate data analysis. Hortic. Environ. Biotechnol. 2018, 59, 759–773. [Google Scholar] [CrossRef]
- Marotti, M.; Piccagli, R. Characterization of flavonoids in different cultivars of Onion (Allium cepa L.). J. Food Sci. 2002, 67, 1229–1232. [Google Scholar] [CrossRef]
- Beato, V.M.; Orgaz, F.; Mansilla, F.; Montaño, A. Changes in phenolic compounds in garlic (Allium sativum L.) owing to the cultivar and location of growth. Plant Foods Hum. Nutr. 2011, 66, 218–223. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Yoon, M.K.; Kwak, J.H. Contents of phytochemical constituents and antioxidant activity of 19 garlic (Allium sativum L.) parental lines and cultivars. Hort. Environ. Biotechnol. 2014, 55, 138–147. [Google Scholar] [CrossRef]
- Hanson, P.M.; Yang, R.Y.; Tsou, S.C.S.; Ledesma, D.; Engle, L.; Lee, T.C. Diversity in eggplant (Solanum melongena) for superoxide scavenging activity, total phenolics, and ascorbic acid. J. Food Compos. Anal. 2006, 19, 594–600. [Google Scholar] [CrossRef]
- Kaur, C.; Nagal, S.; Nishad, J.; Kumar, R.; Sarika, J. Evaluating eggplant (Solanum melongena L.) genotypes for bioactive properties: A chemometric approach. Food Res. Int. 2014, 60, 205–211. [Google Scholar] [CrossRef]
- Ballester, A.R.; Tikunov, Y.; Molthoff, J.; Grandillo, S.; Viquez-Zamora, M.; de Vos, R.; de Maagd, R.A.; van Heusden, S.; Bovy, A.G. Identification of loci affecting accumulation of secondary metabolites in tomato fruit of a Solanum lycopersicum × Solanum chmielewskii introgression line population. Front. Plant Sci. 2016, 7, 1428. [Google Scholar] [CrossRef] [Green Version]
- Lim, W.; Li, J. Synergetic effect of the Onion CHI gene on the PAP1 regulatory gene for enhancing the flavonoid profile of tomato skin. Sci. Rep. 2017, 7, 12377. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classes of Compound | Chemical Structure | IC50 Value | Reference |
|---|---|---|---|
| Alkaloids | |||
| Vasicine |  | 125 µM (0.8 µM) | [24] |
| Piperumbellactam B |  | 43.8 µM (426 µM) | [25] |
| Piperumbellactam C |  | 29.6 µM (426 µM) | |
| Flavonoids | |||
| Quercetin |  | 7 µM | [26] |
| Luteolin |  | 21 µM | |
| Cyanidin |  | 4 µM | |
| Baicalein |  | 0.26 µM (0.02 µM)a | [27] |
| Quercitrin (quercetin-3-O-α-l-rhamnopyranoside |  | ~0.5 mM (0.90 mM) | [28] |
| Isoquercetin |  | 64.1 ± 3.3 µM (1.50 ± 0.14 µM) Maltase 42.5 ± 1.2 µM (2.38 ± 0.02 µM) Sucrase | [29] |
| Cyanidin-diglucoside |  | 14.7 µg/mL | [30] |
| Pelargonidin-3-rutinoside |  | 64.5 µg/mL | |
| Epicatechin-(4β,8)-Epicatechin gallate |  | 0.31 µM (5.3 µM) | [31] |
| Epicatechingallate |  | 0.71 µM (5.3 µM) | |
| Terpense | |||
| 22α-hydroxychiisanoside |  | 819.7 µM (788.6 µM) | [32] |
| 7β-acetoxy-6β-hydroxyroyleanone |  | 108.2 µM (131.2 µM) | [33] |
| Spicatanol |  | 34.1 µM (23.8 µM) | [34] |
| Lupeol |  | 7.18 µg/mL (9.68 µg/mL) | [35] |
| Phenols | |||
| p-hydroxycinnamic acid |  | 90.8 µg/mL (230.4 µg/mL) | [36] |
| Protocatechuic acid |  | 85.1 µg/mL (230.4 µg/mL) | |
| Trans-N-(p-Coumaroyl)tyramine |  | 4.47 µM (168.95 µM) | [37] |
| 2,4-dimethoxy-6,7-dihydroxyphenanthrene |  | 0.40 mM (3.52 mM) | [38] |
| Ferulic acid |  | 4.9 mM (1.7 mM) | [39] |
| Ellagic acid |  | 18.4 µg/mL | [30] |
| Umbelliferone |  | 7.08 µg/mL (9.68 µg/mL) | [34] |
| Iminosugars | |||
| N-(9′-methoxynonyl)-1-deoxynojirimycin |  | 0.015 µM | [40] |
| N-(6′-4″-azido-2″-nitrophenylamino) hexyl-1-deoxynojirimycin |  | 0.017 µM | |
| Vegetables | Phenolic Acids (g·kg−1) | Flavonoids (g·kg−1) | Reference |
|---|---|---|---|
| Tomato | 1.3–3.2 | 1.1–2.4 | [77] |
| Pepper | 7.95–26.15 | 4.64–12.84 | [78] |
| Onion | 3.43–22.19 | 0.0012–0.98 | [79,80] |
| Garlic | 3.4–10.8 | 0.1–0.22 | [81,82] |
| Eggplant (Solanum melongena) | 7.4–14.3 | 0.03–0.26 (fw) | [83,84] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assefa, S.T.; Yang, E.-Y.; Chae, S.-Y.; Song, M.; Lee, J.; Cho, M.-C.; Jang, S. Alpha Glucosidase Inhibitory Activities of Plants with Focus on Common Vegetables. Plants 2020, 9, 2. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9010002
Assefa ST, Yang E-Y, Chae S-Y, Song M, Lee J, Cho M-C, Jang S. Alpha Glucosidase Inhibitory Activities of Plants with Focus on Common Vegetables. Plants. 2020; 9(1):2. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9010002
Chicago/Turabian StyleAssefa, Samuel Tilahun, Eun-Young Yang, Soo-Young Chae, Mihye Song, Jundae Lee, Myeong-Cheoul Cho, and Seonghoe Jang. 2020. "Alpha Glucosidase Inhibitory Activities of Plants with Focus on Common Vegetables" Plants 9, no. 1: 2. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9010002