Effect of Indigenous and Introduced Arbuscular Mycorrhizal Fungi on Growth and Phytochemical Content of Vegetatively Propagated Prunus africana (Hook. f.) Kalkman Provenances


Abstract
:1. Introduction
2. Materials and Methods
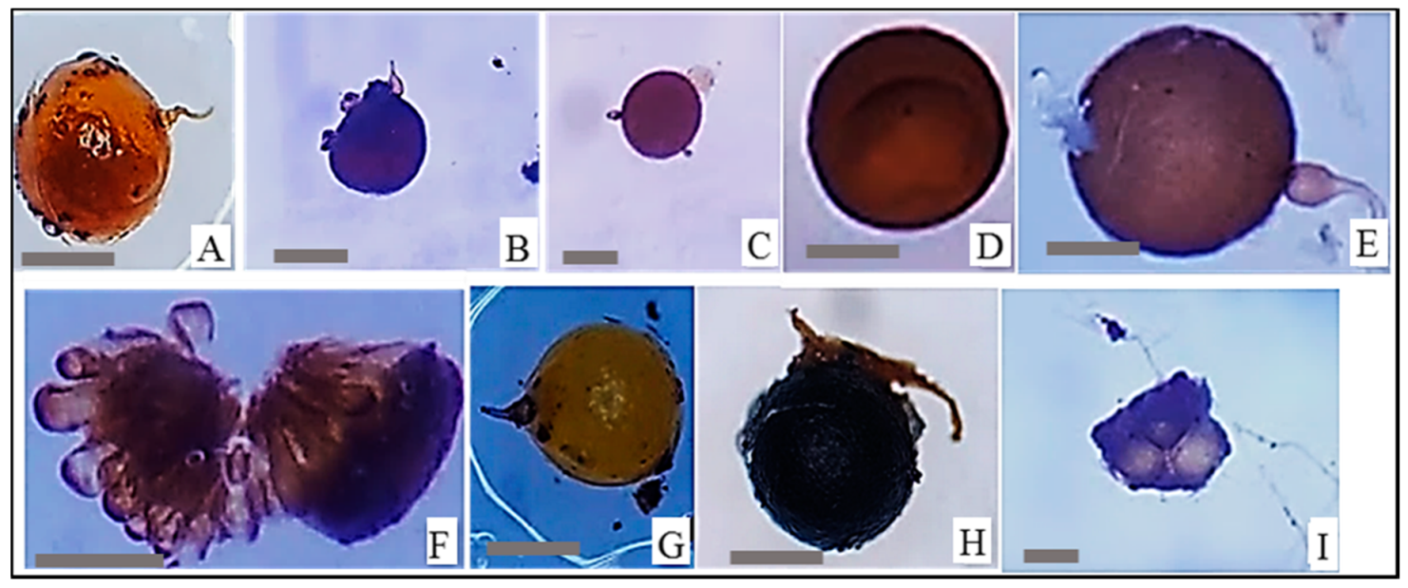
2.1. Soil Samples, Trap Cultures and AMF Inoculum Production
2.2. Production of Vegetatively Propagated P. Africana
2.3. AMF Inoculation Procedure
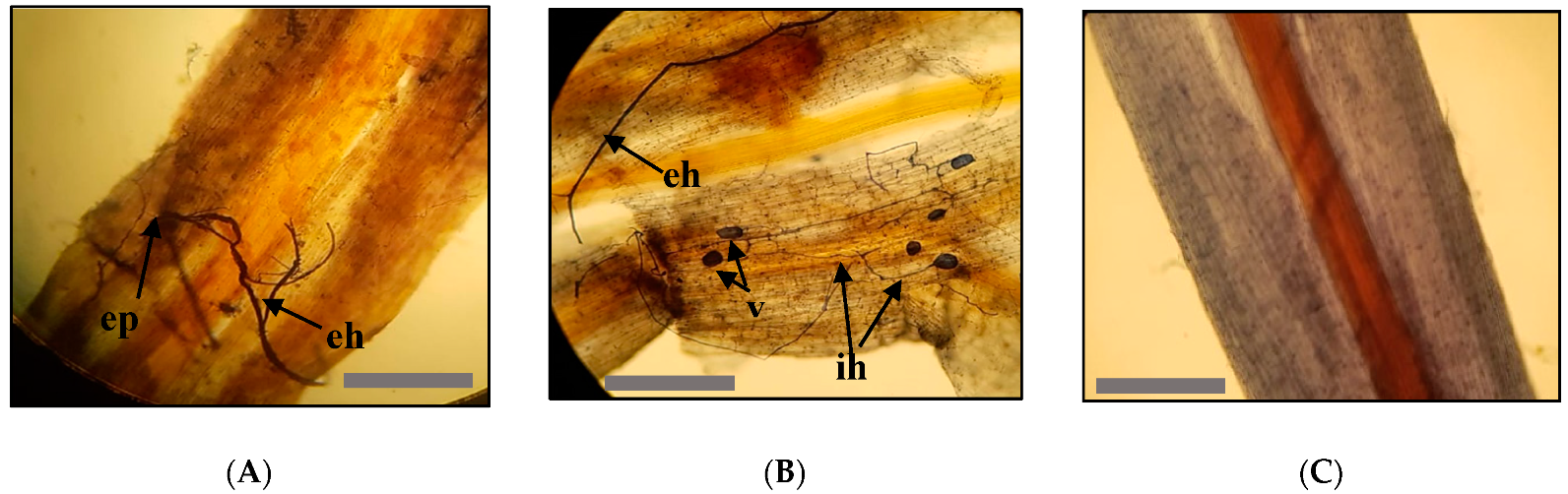
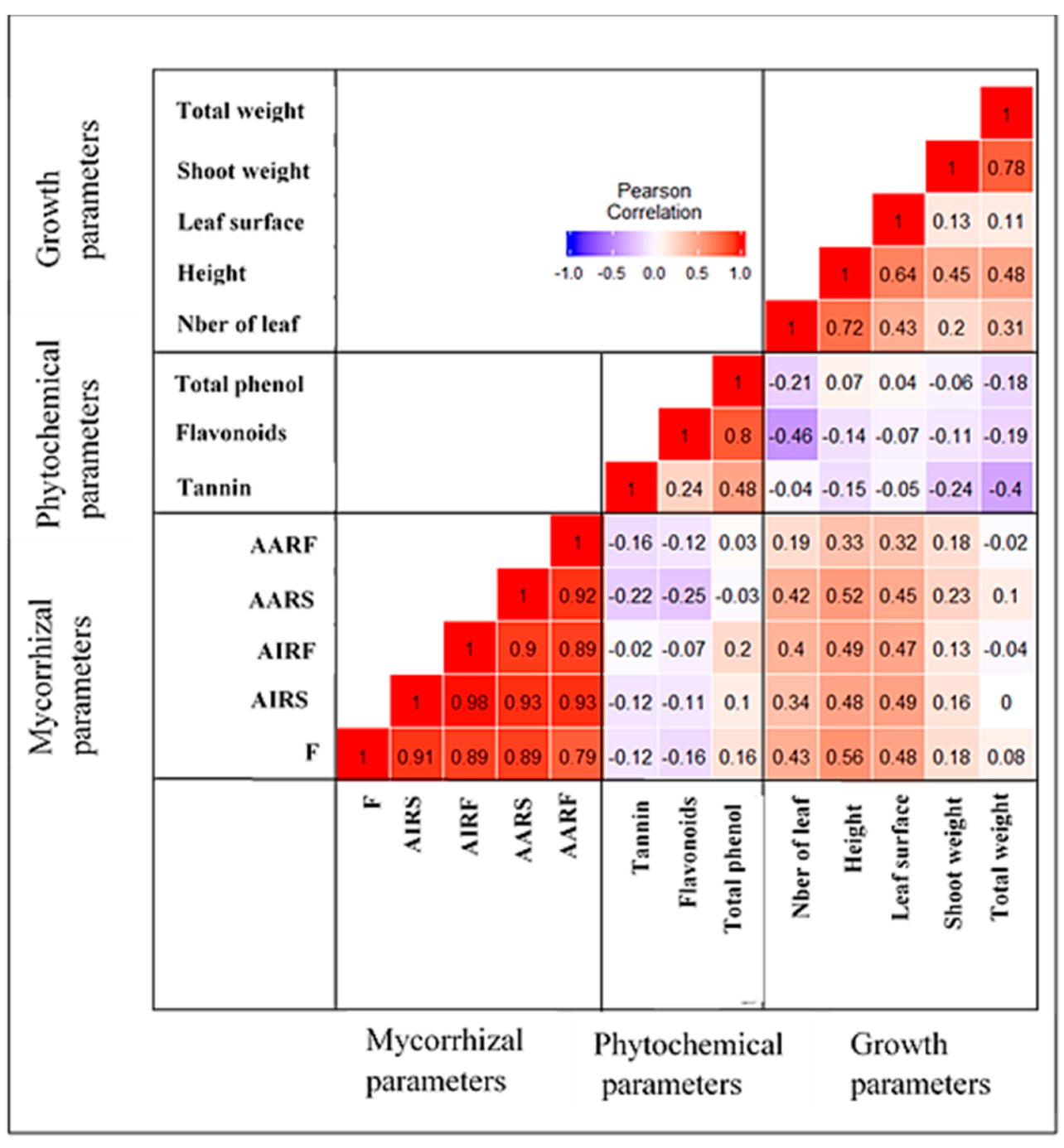
2.4. Measurement of Mycorrhizal Colonization Parameters, Growth and Phytochemical Contents Parameters
2.5. Statistical Analyses
3. Results
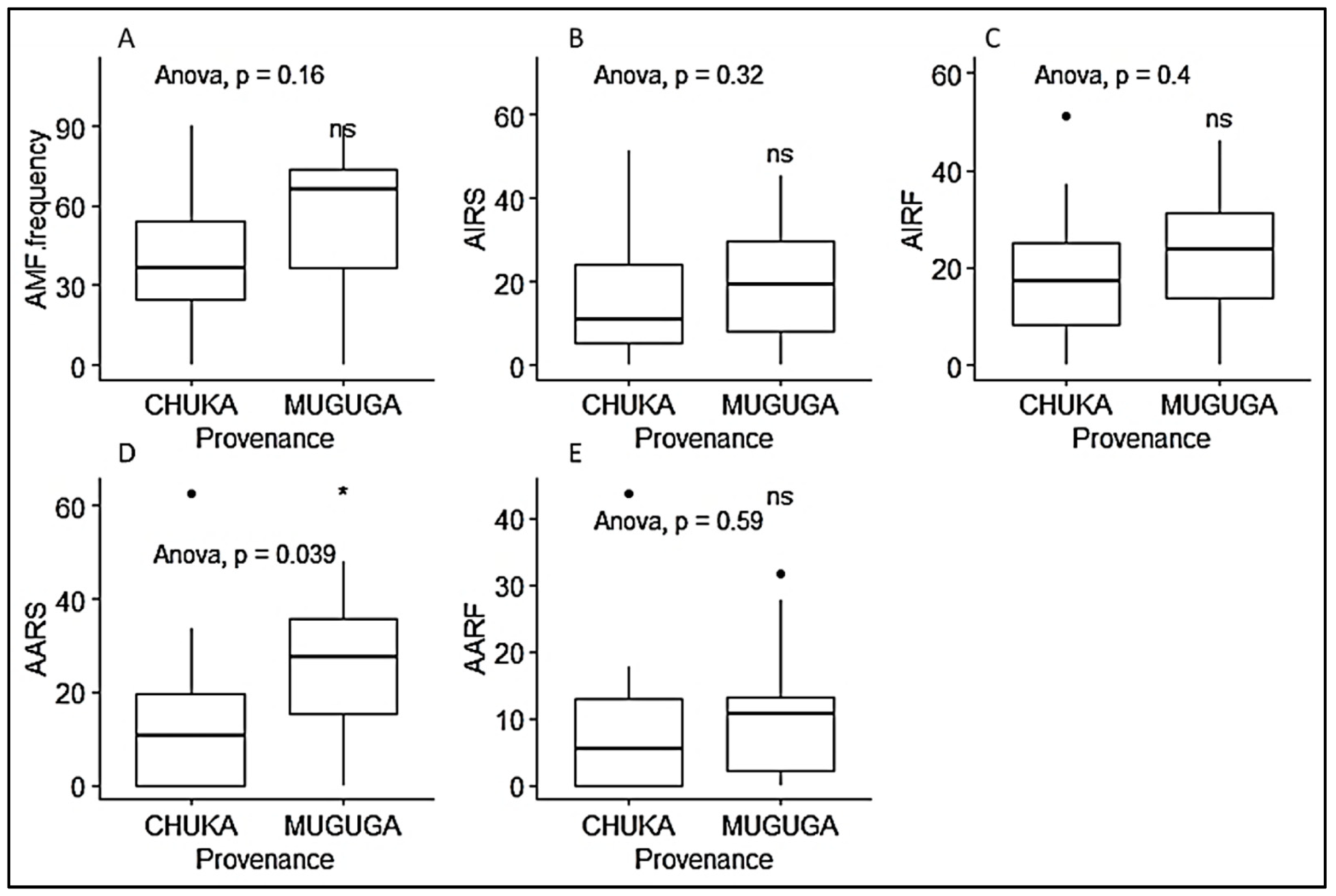
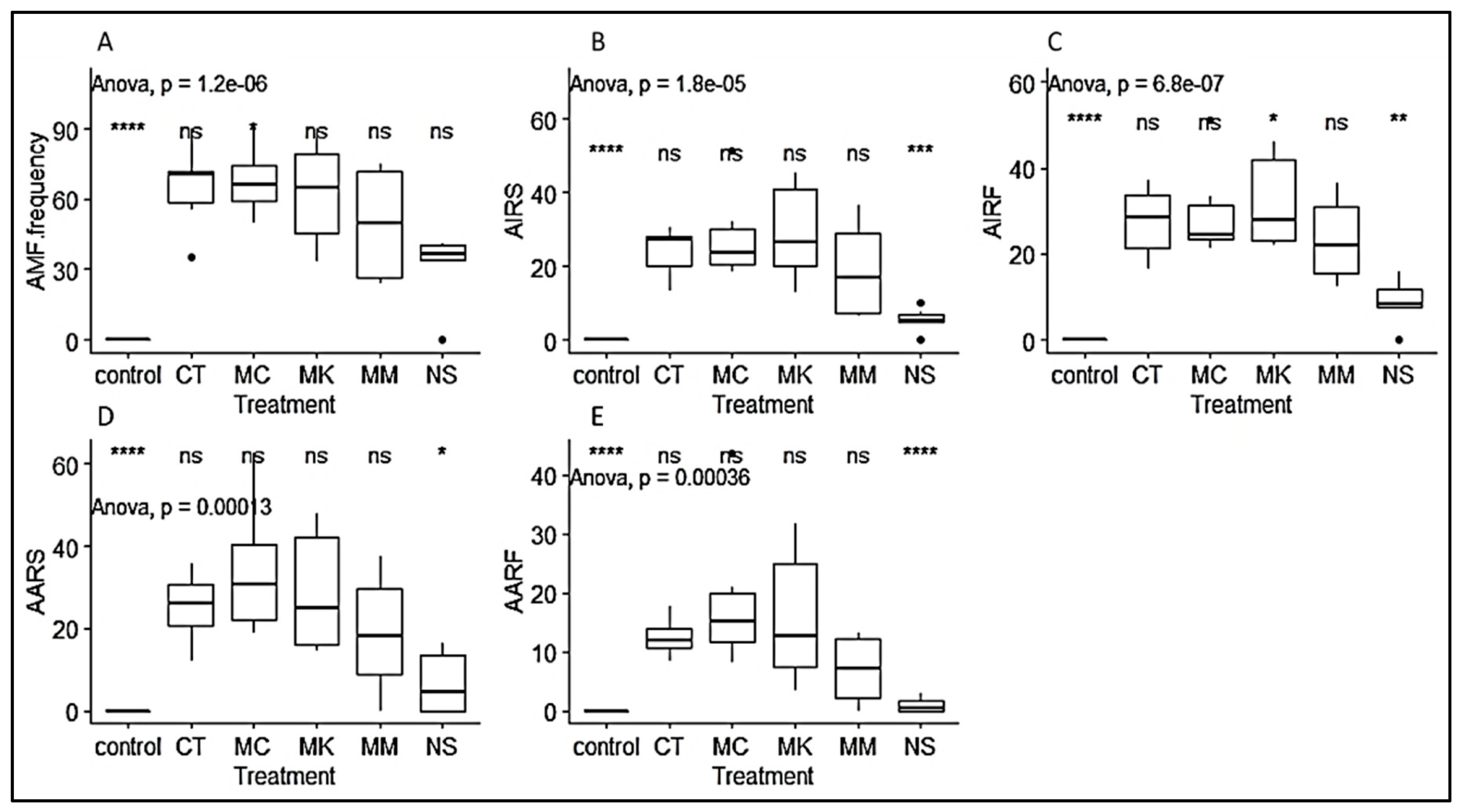
3.1. Mycorrhizal Colonization Parameters
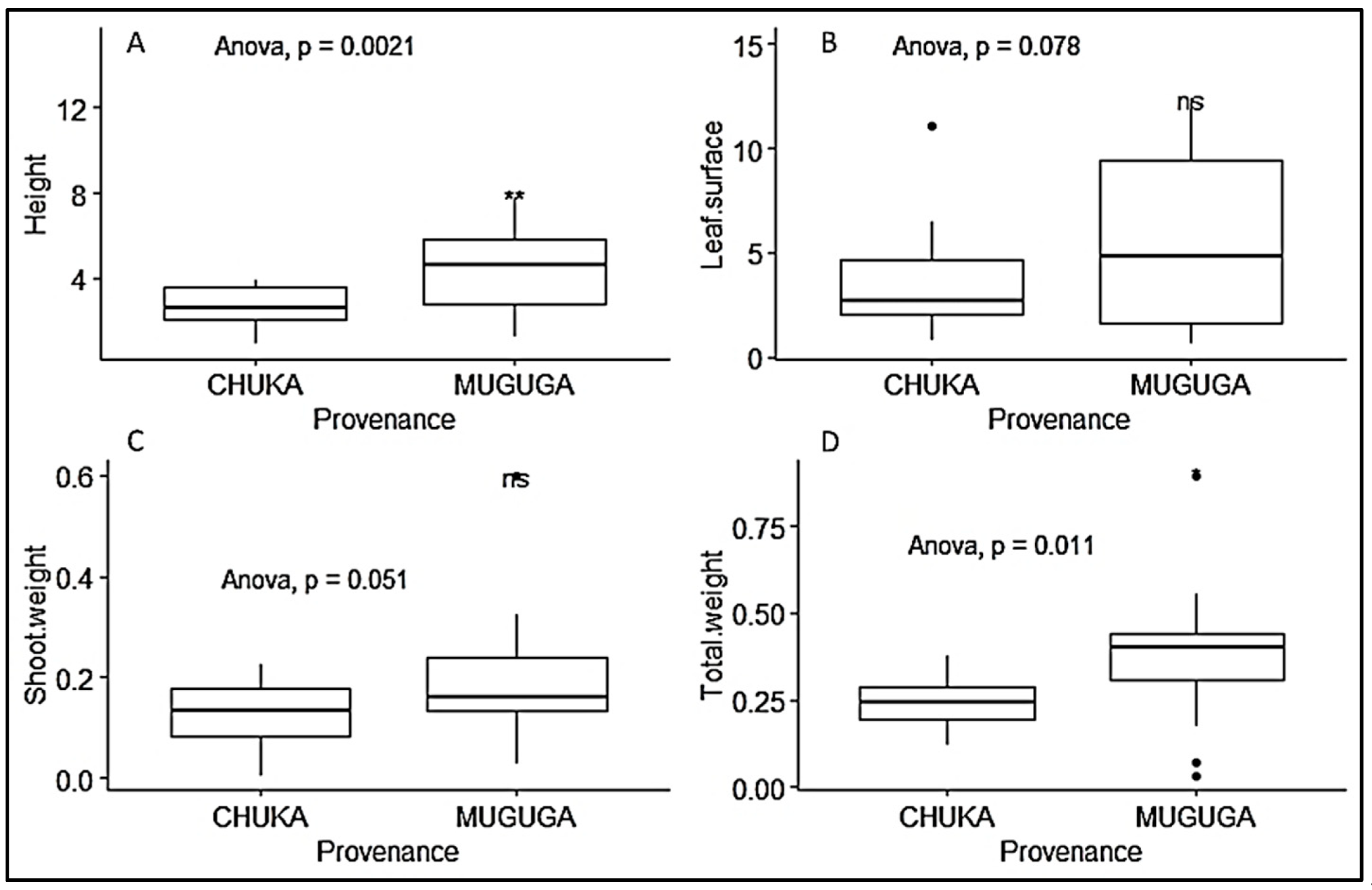
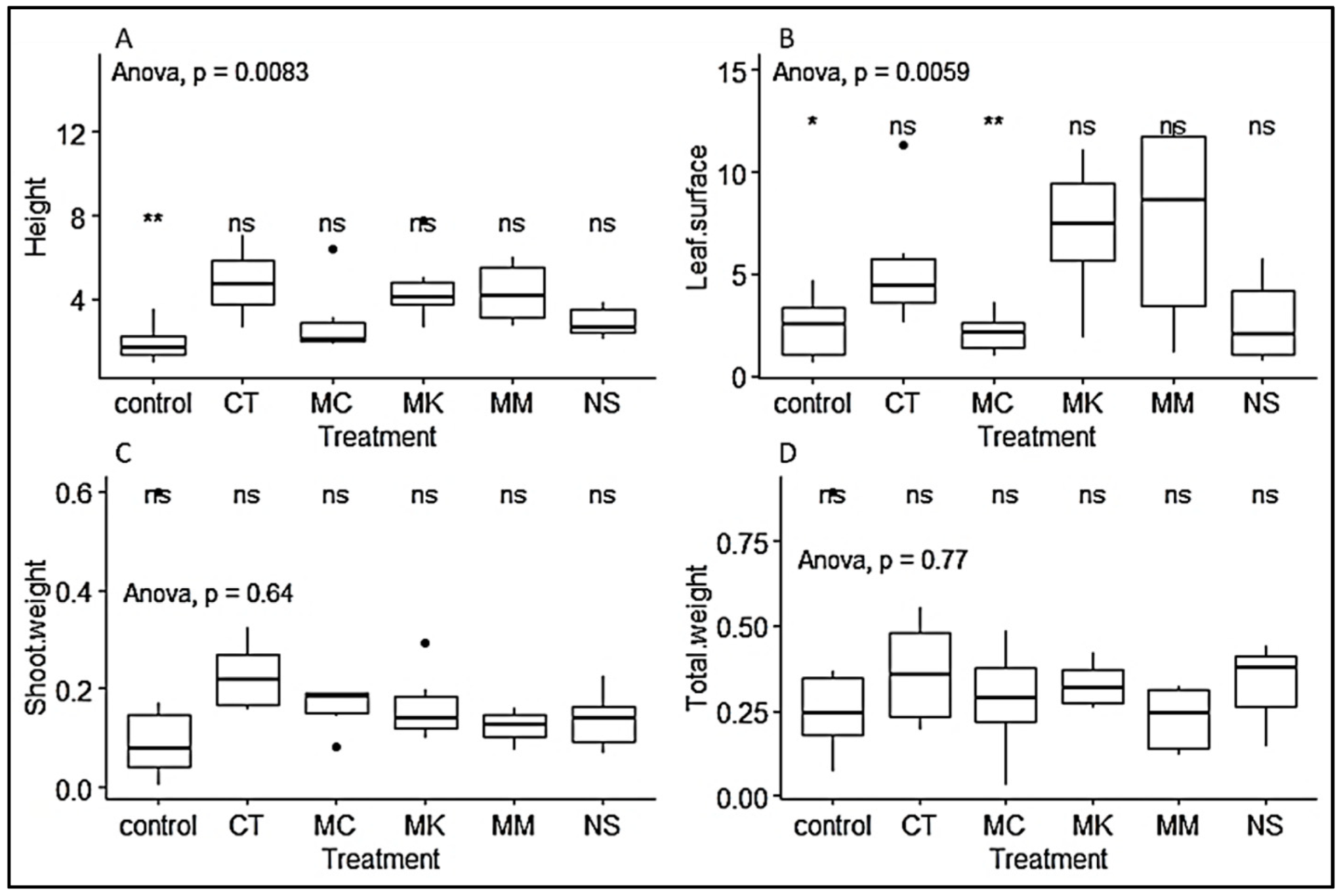
3.2. Growth Parameters of P. Africana Seedlings
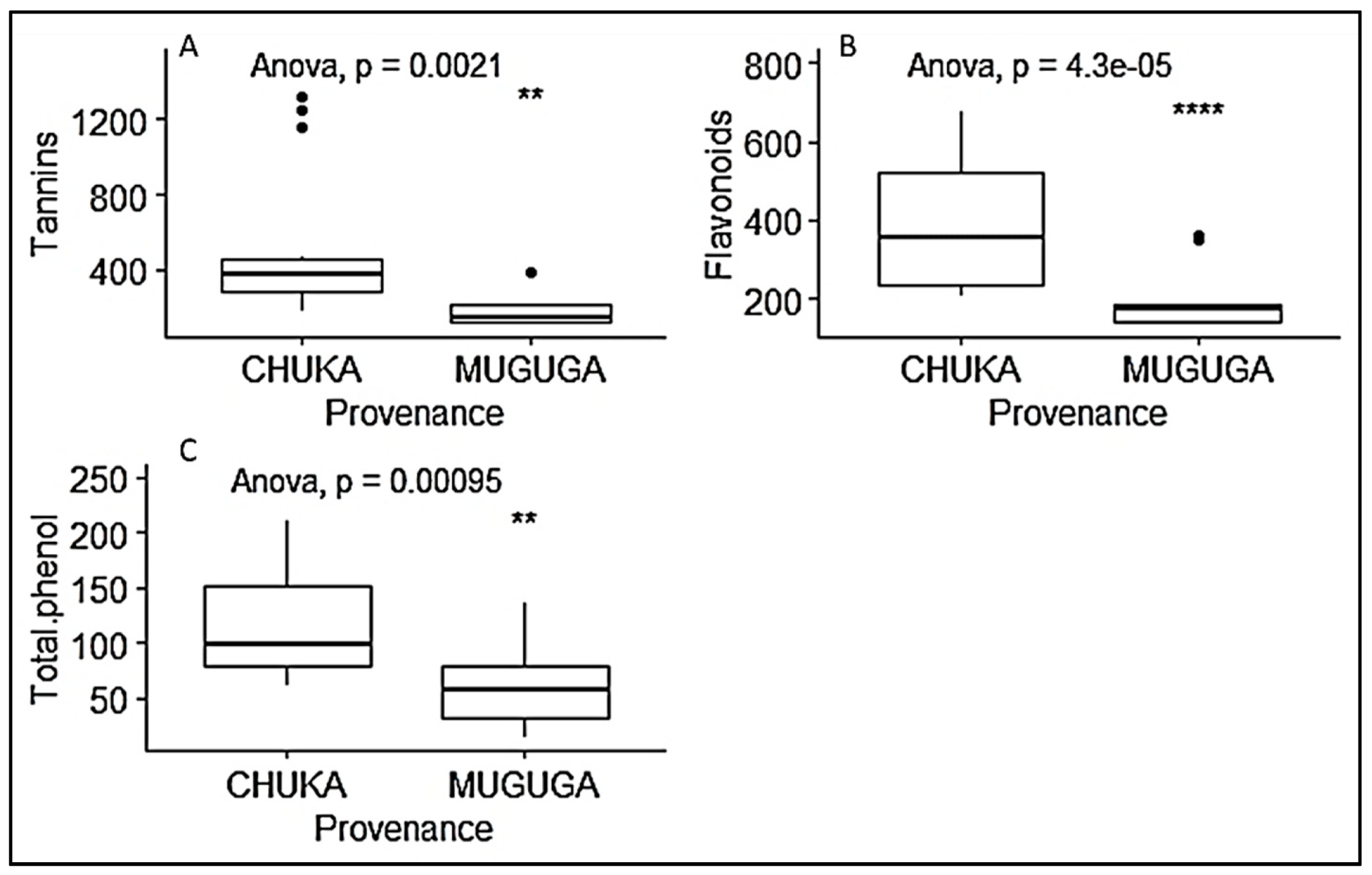
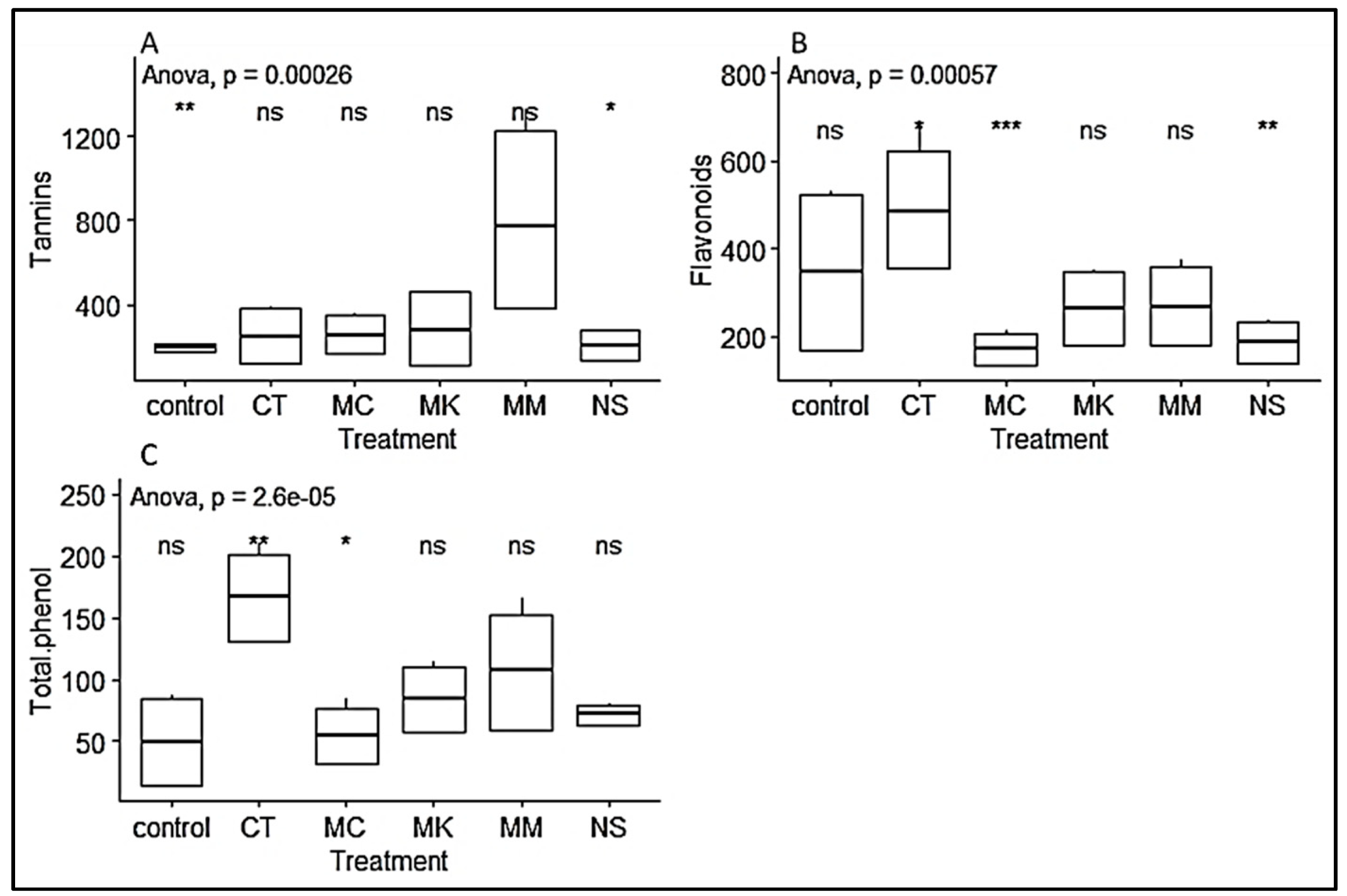
3.3. Phytochemical Content of P. Africana Seedlings
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kadu, C.A.C.; Schueler, S.; Konrad, H.; Muluvi, G.M.M.; Eyog-Matig, O.; Muchugi, A.; Williams, V.L.; Ramamonjisoa, L.; Kapinga, C.; Foahom, B.; et al. Phylogeography of the Afromontane Prunus africana reveals a former migration corridor between East and West African highlands. Mol. Ecol. 2011, 20, 165–178. [Google Scholar] [CrossRef]
- Kotina, E.L.; Oskolski, A.A.; Tilney, P.M.; Wyk, B.V. Bark and wood structure of Prunus africana (Rosaceae), an important African medicinal plant. S. Afr. J. Bot. 2016, 106, 89–95. [Google Scholar] [CrossRef]
- Cunningham, A.; Anoncho, V.F.; Sunderland, T. Power, policy and the Prunus africana bark trade, 1972–2015. J. Ethnopharmacol. 2016, 178, 323–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koros, H.; Konje, M.M.; Wambua, M.; Chesire, C.K.; Odeny, D.; Malombe, I.B. Population Status And Conservation Hotspots of Population Status and Conservation Hotspots Of Prunus africana (Hook. F.) Kalkman in South Nandi Forest, Western Kenya. Res. J. For. 2016, 3, 1–13. [Google Scholar]
- Jena, A.K.; Vasisht, K.; Kaur, R.; Sachdeva, M.; Karan, M. Amelioration of testosterone induced benign prostatic hyperplasia by Prunus species. J. Ethnopharmacol. 2016, 190, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Jena, A.K.; Vasisht, K.; Karan, M. Therapeutic Management of Benign Prostatic Hyperplasia: From Synthetics to Naturals. Annu. Res. Rev. Biol. 2017, 17, 1–34. [Google Scholar] [CrossRef]
- Komakech, R.; Kang, Y.; Lee, J.H.; Omujal, F. A review of the potential of phytochemicals from Prunus africana (Hook f.) kalkman stem bark for chemoprevention and chemotherapy of prostate cancer. Evid. Based Complement. Alternat. Med. 2017, 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, K.M. The African cherry (Prunus africana): Can lessons be learned from an over-exploited medicinal tree? J. Ethnopharmacol. 2003, 89, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Tchoundjeu, Z.; Avana, M.L.; Leakey, R.R.B.; Simons, A.J.; Assah, E.; Duguma, B.; Bell, J.M. Vegetative propagation of Prunus africana: Effects of rooting medium, auxin concentrations and leaf area. Agrofor. Syst. 2002, 54, 183–192. [Google Scholar] [CrossRef]
- Avana, T.L.M. Domestication de Prunus africana (Hook F.) Kalkam: Etude de la Germination et du Bouturage. Ph.D. Thesis, The University of Yaounde, Yaounde, Cameroon, 2006. [Google Scholar]
- Kebede, M.; Hulten, H.; Balcha, G. Vegetative Propagation of Juvenile Leafy Stem Cuttings of Prunus africana (Hook.f.) Kalkm and Syzygium guineense (Willd.) DC. Int. J. Bot. 2013, 9, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Wubet, T.; Weiß, M.; Kottke, I.; Teketay, D.; Franz, O. Molecular diversity of arbuscular mycorrhizal fungi in Prunus an endangered medicinal tree species in dry Afromontane forests of Ethiopia. New Phytol. 2003, 161, 517–528. [Google Scholar] [CrossRef]
- Tchiechoua, Y.H. Étude Morphologique et Moléculaire des Champignons Mycorhiziens à Arbuscules (CMA) Associés à Prunus africana (Hook.f.) Kalkman Dans Deux Zones Agroécologiques du Cameroun; University of Yaounde: Yaounde, Cameroon, 2012. [Google Scholar]
- Brundrett, M.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Spatafora, J.W.; Stajich, J.E.; Benny, G.L.; Smith, M.E.; Berbee, M.L.; Corradi, N.; Grigoriev, I.; James, T.Y.; Donnell, K.O.; Roberson, R.W.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, C.; Carvalho, M.; Brígido, C.; Goss, M.J.; Nobre, T. Symbiosis Specificity of the Preceding Host Plant Can Dominate but Not Obliterate the Association between Wheat and Its Arbuscular Mycorrhizal Fungal Partners. Front. Microbiol. 2018, 9, 2029. [Google Scholar] [CrossRef]
- Torrecillas, E.; Alguacil, M.M.; Roldán, A. Host Preferences of Arbuscular Mycorrhizal Fungi Colonizing Annual Herbaceous Plant Species in Semiarid Mediterranean Prairies. Appl. Environ. Microbiol. 2012, 78, 6180–6186. [Google Scholar] [CrossRef] [Green Version]
- Willis, A.; Rodrigues, B.F.; Harris, P.J.C. The Ecology of Arbuscular Mycorrhizal Fungi. CRC Crit. Rev. Plant Sci. 2013, 32, 1–20. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2008; ISBN 9780123705266. [Google Scholar]
- Keymer, A.; Gutjahr, C. Cross-kingdom lipid transfer in arbuscular mycorrhiza symbiosis and beyond. Curr. Opin. Plant Biol. 2018, 44, 137–144. [Google Scholar] [CrossRef]
- Pozo, M.J.; Slezack-Deschaumes, S.; Dumas-Gaudot, E.; Gianinazzi, S.; Azcon-Aguilar, C. Plant defense responses induced by arbuscular mycorrhizal fungi. In Mycorrhizal Technology in Agriculture; Gianinazzi, S., Schüepp, H., Barea, J.M., Hasselwandter, K., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2002; pp. 103–111. [Google Scholar]
- Jacott, C.N.; Murray, J.D.; Ridout, C.J. Trade-Offs in Arbuscular Mycorrhizal Symbiosis: Disease Resistance, Growth Responses and Perspectives for Crop Breeding. Agronomy 2017, 7, 75. [Google Scholar] [CrossRef] [Green Version]
- Oyewole, B.O.; Olawuyi, O.J.; Odebode, A.C.; Abiala, M.A. Influence of Arbuscular mycorrhiza fungi (AMF) on drought tolerance and charcoal rot disease of cowpea. Biotechnol. Rep. 2017, 14, 8–15. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef]
- Shari, Z.; Azadi, N.; Rahimi, S.; Certini, G. Geoderma The response of glomalin-related soil proteins to fi re or tillage. Geoderma 2018, 329, 65–72. [Google Scholar] [CrossRef]
- Morris, E.K.; Morris, D.J.P.; Vogt, S.; Gleber, S.C.; Bigalke, M.; Wilcke, W.; Rillig, M.C. Visualizing the dynamics of soil aggregation as affected by arbuscular mycorrhizal fungi. ISME J. 2019, 13, 1639–1646. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Guo, L.; Chen, B. Arbuscular mycorrhizal symbiosis and active ingredients of medicinal plants: Current research status and prospectives. Mycorrhiza 2013, 23, 253–265. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, E.L.; da Silva, F.A.; da Silva, F.S.B. Arbuscular Mycorrhizal Fungi Increase the Phenolic Compounds Concentration in the Bark of the Stem of Libidibia Ferrea in Field. Open Microbiol. J. 2017, 11, 283–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copetta, A.; Lingua, G.; Berta, G. Effects of three AM fungi on growth, distribution of glandular hairs, and essential oil production in Ocimum basilicum L. var. Genovese. Mycorrhiza 2006, 16, 485–494. [Google Scholar] [CrossRef]
- Brundrett, M.; Bougher, N.; Dell, B.; Grove, T.; Malajczuk, N. Working with Mycorrhizas in Forestry and Agriculture; Australian Centre for International Agricultural Research: Canberra, Australia, 1996; ISBN 1863201815.
- INVAM. International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. West Virginia University. Available online: https://invam.wvu.edu/ (accessed on 31 October 2019).
- Selvakumar, G.; Shagol, C.C.; Kang, Y.; Chung, B.N.; Han, S.G.; Sa, T.M. Arbuscular mycorrhizal fungi spore propagation using single spore as starter inoculum and a plant host. J. Appl. Microbiol. 2018, 124, 1556–1565. [Google Scholar] [CrossRef]
- Leakey, R.R.; Mesen, J.F.; Tchoundjeu, Z.; Longman, K.A.; Dick, J.M.; Newton, A.; Matin, A.; Grace, J.; Munro, R.C.; Muthoka, P.N. Low-Technology techniques for the vegetative propagation of tropical trees. Commonw. For. Rev. 1990, 69, 247–257. [Google Scholar]
- Kabora, T.K. Comparison of Woody Species Regeneration and Soil Fertility in an Indigenous Forest and Neighbouring Exotic Tree Plantations in Muguga Forest. Master’s Thesis, University of Nairobi, Nairobi, Kenya, 2014. [Google Scholar]
- Mugo, E.N. Chuka Participatory Forest Management Plan 2015-2019; Kenya Forest Service: Nairobi, Kenya, 2015; p. 105. [Google Scholar]
- Odee, D.; Indieka, S.; Lesueur, D. Evaluation of inoculation procedures for Calliandra calothyrsus Meisn. grown in tree nurseries. Biol. Fertil. Soils 2002, 36, 124–128. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un systeme radiculaire. Recherche de methodes d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae; INRA: Paris, France, 1986. [Google Scholar]
- Chaudhary, P.; Godara, S.; Cheeran, A.N.; Chaudhari, A.K. Fast and Accurate Method for Leaf Area Measurement. Int. J. Comput. Appl. 2012, 49, 22–25. [Google Scholar] [CrossRef]
- Price, M.L.; Van Scoyoc, S.; Butler, L.G.A. Critical Evaluation of the Vanillin Reaction as an Assay for Tannin in Sorghum Grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Lamaison, J.L.C.; Carnet, A. Teneurs en principaux flavonoids des fleurs de Crataegeus monogyna Jacq et de Crataegeus laevigata (Poiret) DC. en fonction de la vegetation. Pharm. Acta Helv. 1991, 25, 12–16. [Google Scholar]
- Ragazzi, E.; Veronese, G. Quantitative analysis of phenolic compounds after thin-layer chromatographic separation. J. Chromatogr. A 1973, 77, 369–375. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; ISBN 3-900051-07-0. [Google Scholar]
- Nzweundji, G.; Tchiechoua, Y.H.; Tchotet, J.M.T.; Djocgoué, P.F.; Omokolo, D.N.; Niemenak, N.; Ngome, F.A.; Donfagsiteli, N.T.; Claude, J.; Messi, H.M.; et al. Molecular diversity of arbuscular mycorrhizal fungi associated with Prunus afrivana (Hook F.) Kalkman (Rosaceae) In Humid forest zones of Cameroon. Int. J. Dev. Res. 2015, 5, 5289–5297. [Google Scholar]
- Dierks, J.; Denef, K.; van Diepen, L.T.A.; de Graaff, M.-A. Cheatgrass-associated AMF community negatively affects sagebrush root production but not C transfer to the soil. Plant Soil 2019, 436, 381–396. [Google Scholar] [CrossRef]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial Services of Arbuscular Mycorrhizal Fungi—From Ecology to Application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef]
- Cheng, H.; Fan, Q.; Wu, Q. Effects of Indigenous and Exotic Rhizoglomus intraradices Strains on Trifoliate Orange Seedlings. Biotechnology 2019, 18, 42–48. [Google Scholar]
- Ishani, A.; MacDonald, R.; Nelson, D.; Rutks, I.; Wilt, T.J. Pygeum africanum for the treatment of patients with benign prostatic hyperplasia: A systematic review and quantitative meta-analysis. Am. J. Med. 2000, 109, 654–664. [Google Scholar] [CrossRef]
- Bii, C.; Korir, K.R.; Rugutt, J.; Mutai, C. The potential use of Prunus africana for the control, treatment and management of common fungal and bacterial infections. J. Med. Plants Res. 2010, 4, 995–998. [Google Scholar]
- Madivoli, E.S.; Maina, E.G.; Kairigo, P.K.; Murigi, M.K.; Ogilo, J.K.; Nyangau, J.O.; Kimani, P.K.; Kipyegon, C. In vitro antioxidant and antimicrobial activity of Prunus africana (Hook. f.) Kalkman (bark extracts) and Harrisonia abyssinica Oliv. extracts (bark extracts): A comparative study. J. Med. Plants Econ. Dev. 2018, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Komakech, R.; Kang, Y. Ethnopharmacological potential of African cherry [Prunus africana]. J. Herb. Med. 2019, 100283. [Google Scholar] [CrossRef]
- Katz, A.E. Flavonoid and Botanical Approaches to Prostate Health. J. Altern. Complement. Med. 2002, 8, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Sak Current epidemiological knowledge about the role of flavonoids in prostate carcinogenesis. Exp. Oncol. 2017, 39, 98–105. [CrossRef]
- Reale, G.; Russo, G.I.; Di Mauro, M.; Regis, F.; Campisi, D.; Giudice, A.L.; Marranzano, M.; Ragusa, R.; Castelli, T.; Cimino, S.; et al. Association between dietary flavonoids intake and prostate cancer risk: A case-control study in Sicily. Complement. Ther. Med. 2018, 39, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Geneva, M.P.; Stancheva, I.V.; Boychinova, M.M.; Mincheva, N.H.; Yonova, P.A. Effects of foliar fertilization and arbuscular mycorrhizal colonization on Salvia officinalis L. growth, antioxidant capacity, and essential oil composition. J. Sci. Food Agric. 2010, 90, 696–702. [Google Scholar] [PubMed]
- De Oliveira, P.T.F.; Alves, G.D.; da Silva, F.A.; da Silva, F.S.B. Foliar bioactive compounds in Amburana cearensis (Allemao) A.C. Smith seedlings: Increase of biosynthesis using mycorrhizal technology. J. Med. Plants Res. 2015, 9, 712–718. [Google Scholar]
- Zubek, S.; Rola, K.; Szewczyk, A.; Majewska, M.L.; Turnau, K.; Welling, M.T.; Liu, L.; Rose, T.J.; Waters, D.L.E.; Benkendorff, K. Enhanced concentrations of elements and secondary metabolites in Viola tricolor L. induced by arbuscular mycorrhizal fungi. Plant Soil 2015, 390, 129–142. [Google Scholar] [CrossRef] [Green Version]
- Oszmiański, J.; Lachowicz, S.; Gorzelany, J.; Matłok, N. The effect of different maturity stages on phytochemical composition and antioxidant capacity of cranberry cultivars. Eur. Food Res. Technol. 2018, 244, 705–719. [Google Scholar] [CrossRef] [Green Version]
- Kirui, G.K.; Dossaji, S.F.; Amugune, N.O. Changes in Phytochemical Content during Different Growth Stages in Tubers of Five Varieties of Potato (Solanum Tuberosum L.). Curr. Res. Nutr. Food Sci. J. 2018, 6, 12–22. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Provenance | Characteristic | |||
|---|---|---|---|---|
| Location | Altitude | Annual Rainfall and Modality | Soil Parameters | |
| Muguga forest [34] | Long: 36°38′ E, 36°37′ E Lat: 01°13′ S, 01°12′ S | 2070 masl | 990 mm; Bimodal rainfall (March to May and October to December) | Dominated by clay-loam (6.2–6.5) |
| Chuka forest [35] | Long: 37°19′ E, 37°36′ E Lat: 0°11′ S,0°19′30″ S | 2600–4000 masl | 1500–2500 mm; Bimodal rainfall (March to June and October to December) | Red clay soil, Nitisols, Cambisols, Andosols |
| pH (H2O) | EC mS/cm | C% | N% | P ppm | K ppm | Mg ppm | Ca ppm | Mn ppm | Zn ppm | Cu ppm | Fe ppm |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 7.1 | 0.079 | 1.82 | 0.26 | 45 | 475 | 395 | 4633 | 45 | 6.7 | 1.2 | 189 |
| Provenance | Growth Parameters | Mean | Std. Error |
|---|---|---|---|
| Muguga forest | Log10 Number of leaves | 2.572 a | 0.075 |
| Height (cm) | 4.367 a | 0.260 | |
| Leaf surface area (cm2) | 5.604 a | 0.558 | |
| Shoot weight (g) | 0.195 a | 0.025 | |
| Total weight (g) | 0.375 a | 0.037 | |
| Chuka forest | Log10 Number of leaves | 1.939 b | 0.075 |
| Height (cm) | 2.683 b | 0.260 | |
| Leaf surface area (cm2) | 3.502 b | 0.558 | |
| Shoot weight (g) | 0.129 b | 0.025 | |
| Total weight (g) | 0.244 b | 0.037 |
| Provenance | Phytochemical Parameters | Mean | Std. Error |
|---|---|---|---|
| Muguga forest | Tannin (mg.100 g) | 188.892 b | 5.789 |
| Flavonoids (mg.100 g) | 193.355 b | 2.248 | |
| Total phenol (mg.100 g) | 62.229 b | 0.834 | |
| Chuka forest | Tannin (mg.100 g) | 482.905 a | 5.789 |
| Flavonoids (mg.100 g) | 386.385 a | 2.548 | |
| Total phenol (mg.100 g) | 116.606 a | 0.834 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tchiechoua, Y.H.; Kinyua, J.; Ngumi, V.W.; Odee, D.W. Effect of Indigenous and Introduced Arbuscular Mycorrhizal Fungi on Growth and Phytochemical Content of Vegetatively Propagated Prunus africana (Hook. f.) Kalkman Provenances. Plants 2020, 9, 37. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9010037
Tchiechoua YH, Kinyua J, Ngumi VW, Odee DW. Effect of Indigenous and Introduced Arbuscular Mycorrhizal Fungi on Growth and Phytochemical Content of Vegetatively Propagated Prunus africana (Hook. f.) Kalkman Provenances. Plants. 2020; 9(1):37. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9010037
Chicago/Turabian StyleTchiechoua, Yves H., Johnson Kinyua, Victoria Wambui Ngumi, and David Warambo Odee. 2020. "Effect of Indigenous and Introduced Arbuscular Mycorrhizal Fungi on Growth and Phytochemical Content of Vegetatively Propagated Prunus africana (Hook. f.) Kalkman Provenances" Plants 9, no. 1: 37. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9010037