Evaluation of Cross-Species Transferability of SSR Markers in Foeniculum vulgare

 ,
,  and
and
Abstract
:1. Introduction
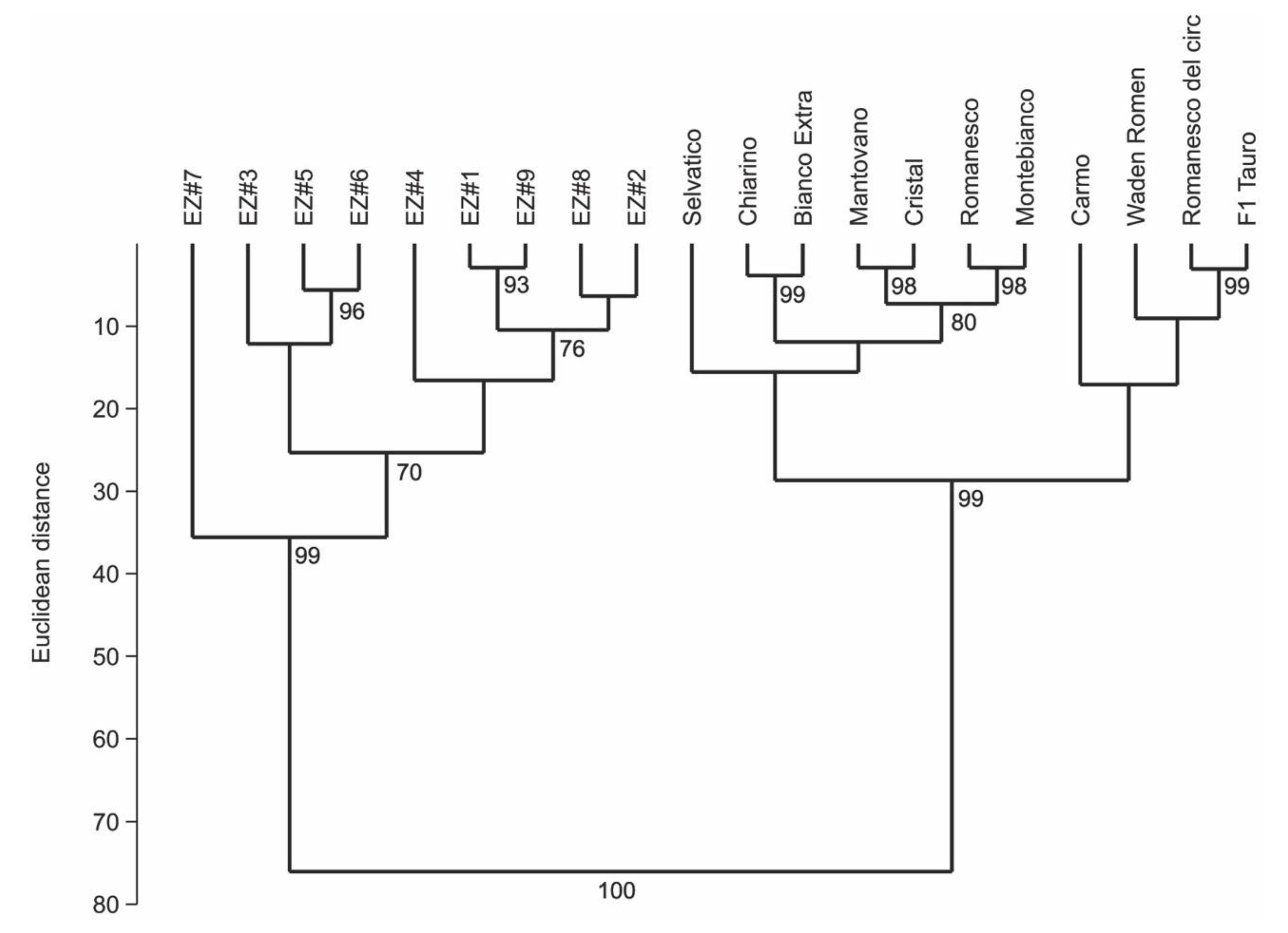
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material and Genomic DNA Extraction
4.2. Primer Design and SSR Amplification by PCR
4.3. Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Krishnamurthy, K.H. Medicinal plants, Madhurikā, saunf or fennel (Foeniculum vulgare, Gaertn). NAMAH 2011, 19, 1–4. [Google Scholar]
- Rather, M.A.; Dar, B.A.; Sofi, S.N.; Bhat, B.A.; Qurishi, M.A. Foeniculum vulgare: A comprehensive review of its traditional use, phytochemistry, pharmacology, and safety. Arabian J. Chem. 2012, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Badgujar, S.B.; Patel, V.V.; Bandivdekar, A.H. Foeniculum vulgare Mill: A review of its botany, phytochemistry, pharmacology, contemporary application, and toxicology. Biomed. Res. Int. 2014, 2014, 842674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betts, T.J. Anethole and fenchone in the developing fruits of Foeniculum vulgare Mill. J. Pharm. Pharmacol. 1998, 20, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Statti, G.; Uzunov, D.; Menichini, F. Comparative chemical composition and antioxidant activities of wild and cultivated Laurus nobilis L. leaves and Foeniculum vulgare subsp. piperitum (Ucria) coutinho seeds. Biol. Pharm. Bull. 2006, 29, 2056–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Maroto, M.C.; Pearez-Coello, M.S. Comparison of the volatile composition of wild fennel samples (Foeniculum vulgare Mill.) from Central. J. Agric. Food Chem. 2006, 54, 6814–6818. [Google Scholar] [CrossRef]
- He, W.; Huang, B. A review of chemistry and bioactivities of a medicinal spice: Foeniculum vulgare. J. Med. Plant Res. 2011, 5, 3595–3600. [Google Scholar]
- Malhotra, S.K. Fennel and fennel seed. In Handbook of Herbs and Spices, 2nd ed.; Peter, K.V., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2012; pp. 275–302. [Google Scholar]
- Syed, F.Q.; Mirza, M.B.; Elkady, A.I.; Hakeem, K.R.; Alkarim, S. Plant and Human Health; Springer: Basel, Switzerland, 2019. [Google Scholar]
- Pieroni, A.; Giusti, M.E. Ritual botanicals against the Evil-Eye in Tuscany, Italy. Econ. Bot. 2002, 56, 201–203. [Google Scholar] [CrossRef] [Green Version]
- Gori, L.; Gallo, E.; Mascherini, V.; Mugelli, A.; Vannacci, A.; Firenzuoli, F. Can estragole in fennel seed decoctions really be considered a danger for human health? A fennel safety update. Evid. Based Complement. Alternat. Med. 2012, 2012, 860542. [Google Scholar] [CrossRef] [Green Version]
- Lucotte, G.; Thomasset, T.; Salmon, A. Fennel (Foeniculum vulgare) Rests on the Holy Maria-Magdalena’s Hairs, Studied by Scanning Electron Microscopy and Elemental Analysis. Archaeol. Discov. 2018, 6, 216–270. [Google Scholar] [CrossRef] [Green Version]
- Kuijper, W.J.; Turner, H. Diet of a Roman centurion at Alphen aan den Rijn, The Netherlands, in the first century AD. Rev. Palaeobot. Palynol. 1992, 73, 187–204. [Google Scholar] [CrossRef]
- Hanlidou, E.; Karousou, R.; Kleftoyanni, V.; Kokkini, S. The herbal market of Thessaloniki (N Greece) and its relation to the ethnobotanical tradition. J. Ethnopharmacol. 2004, 91, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Aboelsoud, N.H. Herbal medicine in ancient Egypt. J. Med. Plant Res. 2010, 4, 82–86. [Google Scholar]
- Choi, E.M.; Hwang, J.K. Anti-inflammatory, analgesic and antioxidant activities of the fruit of Foeniculum vulgare. Fitoterapia 2004, 75, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Koppula, S.; Kumar, H. Foeniculum vulgare Mill (Umbelliferae) Attenuates Stress and Improves Memory in Wister Rats. Trop. J. Pharm. Res. 2013, 12, 553–558. [Google Scholar] [CrossRef] [Green Version]
- Kooti, W.; Moradi, M.; Ali-Akbari, S.; Sharafi-Ahvazi, N.; Asadi-Samani, M.; Ashtary-Larky, D. Therapeutic and pharmacological potential of Foeniculum vulgare Mill: A review. J. HerbMed. Pharmacol. 2015, 4, 1–9. [Google Scholar]
- El-Soud, N.A.; El-Laithy, N.; El-Saeed, G.; Wahby, M.S.; Khalil, M.; Morsy, F.; Shaffie, N. Antidiabetic Activities of Foeniculum Vulgare Mill. Essential Oil in Streptozotocin-Induced Diabetic Rats. Maced. J. Med. Sci. 2011, 4, 139–146. [Google Scholar]
- Xue, G.Q.; Liu, Q.; Han, Y.Q.; Wei, H.G.; Dong, T. Determination of thirteen metal elements in the plant Foeniculum vulgare Mill. by flame atomic absorption spectrophotometry. Guang Pu 2006, 26, 1935–1938. [Google Scholar]
- Salama, Z.A.; El Baz, F.K.; Gaafar, A.A.; Zaki, M.F. Antioxidant activities of phenolics, flavonoids and vitamin C in two cultivars of fennel (Foeniculum vulgare Mill.) in responses to organic and bio-organic fertilizers. J. Saudi Soc. Agric. Sci. 2015, 14, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Khammassi, M.; Loupassaki, S.; Tazarki, H.; Mezni, F.; Slama, A.; Tlili, N.; Zaouali, Y.; Mighri, H.; Jamoussi, B.; Khaldi, A. Variation in essential oil composition and biological activities of Foeniculum vulgare Mill. populations growing widely in Tunisia. J. Food Biochem. 2018, 42, 12532. [Google Scholar] [CrossRef]
- Rezayat, S.M.; Dehpour, A.R.; Motamed, S.M.; Yazdanparast, M.; Chamanara, M.; Sahebgharani, M.; Rashidian, A. Foeniculum vulgare essential oil ameliorates acetic acid-induced colitis in rats through the inhibition of NF-kB pathway. Inflammopharmacology 2018, 26, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Bennici, A.; Anzide, M.; Vendramin, G.G. Genetic stability and uniformity of Foeniculum vulgare Mill. Regenerated plants through organogenesis and somatic embryogenesis. Plant Sci. 2004, 166, 221–227. [Google Scholar] [CrossRef]
- Grover, S.; Jakhar, M.L.; Malik, C.P. Genetic diversity of different varieties Foeniculum vulgare Miller by RAPD markers. Arch. Appl. Sci. Res. 2011, 3, 17–25. [Google Scholar]
- Torabi, S.; Hasani, M.H.; Omidi, M.; Etminan, A.; Dasmalchi, T.; Gharakhanlou, H. Evaluation of genetic diversity in Fennel accessions using AFLP markers. Adv. Environ. Biol. 2012, 6, 2821–2828. [Google Scholar]
- Abou El-Nasr, T.H.S.; Ibrahim, M.M.; Aboud, K.A.; Al-Kordy, M.A.A. Genetic variation among three fennel (Foeniculum vulgare Mill.) varieties on the basis of morphological characters, essential oil composition and ISSR markers. J. Appl. Sci. Res. 2013, 9, 1594–1603. [Google Scholar]
- Godwin, I.D.; Aitken, E.A.B.; Smith, L.W. Application of inter-simple sequence repeats (ISSR) markers to plant genetics. Electrophoresis 1997, 18, 1524–1528. [Google Scholar] [CrossRef]
- Maghsoudi Kelardashti, H.; Rahimmalek, M.; Talebi, M. Genetic diversity in Iranian Fennel (Foeniculum vulgare Mill.) populations based on sequence related amplified polymorphism (SRAP) markers. J. Agric. Sci. Tech. 2015, 17, 1789–1803. [Google Scholar]
- Schlötterer, C.; Vogl, C.; Tautz, D. Polymorphism and locus-specific effects on polymorphism at microsatellite loci in natural in Drosophila melanogaster populations. Genetics 1997, 146, 309–320. [Google Scholar]
- Yu, K.; Park, S.J.; Poysa, V.; Gepts, P. Integration of Simple Sequence Repeat (SSR) markers into a molecular linkage map of common bean (Phaseolus vulgaris L.). J. Hered. 2000, 91, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Collard, B.C.; Mackill, D.J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2008, 363, 557–572. [Google Scholar] [CrossRef] [Green Version]
- Guichoux, E.; Lagache, L.; Wagner, S.; Chaumeil, P.; Léger, P.; Lepais, O.; Lepoittevin, C.; Malausa, T.; Revardel, E.; Salin, F.; et al. Current trends in microsatellite genotyping. Mol. Ecol. Resour. 2011, 11, 591–611. [Google Scholar] [CrossRef] [PubMed]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Islam, K.N.; Latif, M.A. A review of microsatellite markers and their applications in rice breeding programs to improve blast disease resistance. Int. J. Mol. Sci. 2013, 14, 22499–22528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squirrell, J.; Hollingsworth, P.M.; Woodhead, M.; Russell, J.; Lowe, A.J.; Gibby, M.; Powell, W. How much effort is required to isolate nuclear microsatellites from plants? Mol. Ecol. 2003, 12, 1339–1348. [Google Scholar] [CrossRef]
- Peakall, R.; Gilmore, S.; Keys, W.; Morgante, M.; Rafalski, A. Cross-species amplification of soybean (Glycine max) simple sequence repeats (SSRs) within the genus and other legume genera: Implications for the transferability of SSRs in plants. Mol. Biol. Evol. 1998, 15, 1275–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbará, T.; Palma-Silva, C.; Paggi, G.M.; Bered, F.; Fay, M.F.; Lexer, C. Cross-species transfer of nuclear microsatellite markers: Potential and limitations. Mol. Ecol. 2007, 16, 3759–3767. [Google Scholar] [CrossRef]
- Castillo, A.; Budak, H.; Varshney, R.K.; Dorado, G.; Graner, A.; Hernandez, P. Transferability and polymorphism of barley EST-SSR markers used for phylogenetic analysis in Hordeum chilense. BMC Plant Biol. 2008, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Zhang, Y.; Jin, M.; Li, H.; Song, Z.; Wang, Y.; Chen, J. Characterization and high cross-species transferability of microsatellite markers from the floral transcriptome of Aspidistra saxicola (Asparagaceae). Mol. Ecol. Resour. 2014, 14, 569–577. [Google Scholar] [CrossRef]
- Roa, A.C.; Chavarriaga-Aguirre, P.; Duque, M.C.; Maya, M.M.; Bonierbale, M.W.; Iglesias, C.; Tohme, J. Cross-species amplification of cassava (Manihot esculenta) (Euphorbiaceae) microsatellites: Allelic polymorphism and degree of relationship. Am. J. Bot. 2000, 87, 1647–1655. [Google Scholar] [CrossRef]
- Palumbo, F.; Vannozzi, A.; Vitulo, N.; Margherita Lucchini, M.; Barcaccia, G. The leaf transcriptome of fennel (Foeniculum vulgare Mill.) enables characterization of the t-anethole pathway and the discovery of microsatellites and single nucleotide variants. Sci. Rep. 2018, 8, 10459. [Google Scholar] [CrossRef]
- Palumbo, F.; Galla, G.; Nicola Vitulo, N.; Barcaccia, G. First draft genome sequencing of fennel (Foeniculum vulgar Mill.): Identification of simple sequence repeats and their application in marker-assisted breeding. Mol. Breed. 2018, 38, 122. [Google Scholar] [CrossRef]
- Cavagnaro, P.F.; Chung, S.M.; Manin, S.; Yildiz, M.; Ali, A.; Alessandro, M.S.; Iorizzo, M.; Senalik, D.A.; Simon, P.W. Microsatellite isolation and marker development in carrot -genomic distribution, linkage mapping, genetic diversity analysis and markertransferability across Apiaceae. BMC Genom. 2011, 12, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, P.; Tavaré, S. The distribution of rare alleles. J. Math. Biol. 1995, 33, 602–618. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef]
- Gupta, P.K.; Rustgi, S.; Sharma, S.; Singh, R.; Kumar, N.; Balyan, H.S. Transferable EST-SSR markers for the study of polymorphism and genetic diversity in bread wheat. Mol. Genet. Genom. 2003, 270, 315–323. [Google Scholar] [CrossRef]
- Saha, M.C.; Mian, M.A.R.; Eujayl, I.; Zwonitzer, J.C.; Wang, L.; May, G.D. Tall fescue EST-SSR markers with transferability across several grass species. Theor. Appl. Genet. 2004, 109, 783–791. [Google Scholar] [CrossRef]
- Feng, S.P.; Li, W.G.; Huang, H.S.; Wang, J.Y.; Wu, Y.T. Development, characterization and cross-species/genera transferability of EST-SSR markers for rubber tree (Hevea brasiliensis). Mol. Breed. 2009, 23, 85. [Google Scholar] [CrossRef]
- Simko, I. Development of EST-SSR markers for the study of population structure in lettuce (Lactuca sativa L.). J. Hered. 2009, 100, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Zhang, Q.; Tan, C.; Zhang, X.Q.; Li, C. Development of genome-wide InDel markers and their integration with SSR, DArT and SNP markers in single barley map. BMC Genom. 2015, 16, 804. [Google Scholar] [CrossRef] [Green Version]
- Sourdille, P.; Tavaud, M.; Charmet, G.; Bernard, M. Transferability of wheat microsatellites to diploid Triticeae species carrying the A., B and D genomes. Theor. Appl. Genet. 2001, 103, 346. [Google Scholar] [CrossRef]
- Liewlaksaneeyanawin, C.; Ritland, C.E.; El-Kassaby, Y.A.; Ritland, K. Single-copy, species-transferable microsatellite markers developed from loblolly pine ESTs. Theor. Appl. Genet. 2004, 109, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Cholin, S.S.; Poleshi, C.A.; Manikanta, D.S.; Christopher, C. Exploring the genomic resources of carrot for cross-genera transferability and phylogenetic assessment among orphan spices and vegetables of Apiaceae family. Hortic. Environ. Biotechnol. 2019, 60, 81–93. [Google Scholar] [CrossRef]
- Rossetto, M. Sourcing of SSR markers from related plant species. In Plant genotyping-the DNA fingerprinting of plant; Henry, R.J., Ed.; CABI Publishing: Wallingford, OX, USA, 2001; pp. 211–224. [Google Scholar]
- Katzir, N.; Danin-Poleg, Y.; Tzuri, G.; Karchi, Z.; Lavi, U.; Cregan, P.B. Length polymorphism and homologies of microsatellites in several Cucurbitaceae species. Theor. Appl. Genet. 1996, 93, 1282–1290. [Google Scholar] [CrossRef]
- Steinkellner, H.; Fluch, S.; Turetschek, E.; Lexer, C.; Streiff, R.; Kremer, A.; Burg, K.; Glössl, J. Identification and characterization of (GA/CT)n-microsatellite loci from Quercus petraea. Plant Mol. Biol. 1997, 33, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Whitton, J.; Rieseberg, L.H.; Ungerer, M.C. Microsatellite loci are not conserved across the Asteraceae. Mol. Biol. Evol. 1997, 14, 204–209. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.D.; Eggler, P.; Seaton, G.; Rossetto, M.; Ablett, E.M.; Lee, L.S.; Henry, R.J. Analysis of SSRs derived from grape ESTs. Theore. Appl. Gen. 2000, 100, 723–726. [Google Scholar] [CrossRef]
- FitzSimmons, N.N.; Moritz, C.; Moore, S. Conservation and dynamics of microsatellite loci over 300 million years of marine turtle evolution. Mol. Biol. Evol. 1995, 12, 432–440. [Google Scholar]
- Dirlewanger, E.; Cosson, P.; Tavaud, M.; Aranzana, J.; Poizat, C.; Zanetto, A.; Arús, P.; Laigret, F. Development of microsatellite markers in peach [Prunus persica (L.) Batsch] and their use in genetic diversity analysis in peach and sweet cherry (Prunus avium L.). Theor. Appl. Genet. 2002, 105, 127–138. [Google Scholar] [CrossRef]
- Holton, T.A.; Christopher, J.T.; McClure, L.K.; Harker, N.; Henry, R.J. Identification and mapping of polymorphic SSR markers from expressed gene sequences of barley and wheat. Mol. Breed. 2002, 9, 63–71. [Google Scholar] [CrossRef]
- Callen, D.F.; Thompson, A.D.; Shen, Y.; Phillips, H.A.; Richards, R.I.; Mulley, J.C.; Sutherland, G.R. Incidence and origin of null alleles in the (AC)n microsatellite markers. Am. J. Hum. Genet. 1993, 52, 922–927. [Google Scholar]
- Chapuis, M.P.; Estoup, A. Microsatellite null alleles and estimation of population differentiation. Mol. Biol. Evol. 2007, 24, 621–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico, C.; Cuesta, J.A.; Drake, P.; Macpherson, E.; Bernatchez, L.; Marie, A.D. Null alleles are ubiquitous at microsatellite loci in the Wedge Clam (Donax trunculus). Peer J. 2017, 18, e3188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimaldi, M.C.; Crouau-Roy, B. Microsatellite allelic homoplasy due to variable flanking sequences. J. Mol. Evol. 1997, 44, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Meglécz, E.; Péténian, F.; Danchin, E.; Coeur D’acier, A.; Rasplus, J.Y.; Faure, E. High similarity between flanking regions of different microsatellites detected within each of two species of Lepidoptera: Parnassius apollo and Euphydryas aurinia. Mol. Ecol. 2004, 13, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Rallo, P.; Dorado, G.; Martín, A. Development of simple sequence repeats (SSRs) in olive tree (Olea europaea L.). Theor. Appl. Genet. 2000, 101, 984–989. [Google Scholar] [CrossRef]
- Lia, V.V.; Confalonieri, V.A.; Poggio, L. B chromosome polymorphism in maize landraces: Adaptive vs. demographic hypothesis of clinal variation. Genetics 2007, 177, 895–904. [Google Scholar] [CrossRef]
- Britten, R.J.; Rowen, L.; Williams, J.; Cameron, R.A. Majority of divergence between closely related DNA samples is due to indels. Proc. Natl. Acad. Sci. USA 2003, 100, 4661–4665. [Google Scholar] [CrossRef] [Green Version]
- Burstin, J.; Deniot, G.; Potier, J.; Weinachter, C.; Aubert, G.; Barranger, A. Microsatellite polymorphism in Pisum sativum. Plant Breed. 2001, 120, 311–317. [Google Scholar] [CrossRef]
- Moretzsohn, M.C.; Leoi, L.; Proite, K.; Guimarães, P.M.; Leal-Bertioli, S.C.M.; Gimenes, M.A.; Martins, W.S.; Valls, J.F.M.; Grattapaglia, D.; Bertioli, D.J. A microsatellite-based, gene-rich linkage map for the AA genome of Arachis (Fabaceae). Theor. Appl. Genet. 2005, 111, 1060–1071. [Google Scholar] [CrossRef]
- Mun, J.H.; Kim, D.J.; Choi, H.K.; Gish, J.; Debellé, F.; Mudge, J.; Denny, R.; Endré, G.; Saurat, O.; Dudez, A.M.; et al. Distribution of microsatellites in the genome of Medicago truncatula: A resource of genetic markers that integrate genetic and physical maps. Genetics. 2006, 172, 2541–2555. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wang, X.; He, Q.; Liu, X.; Xu, W.; Li, L.; Gao, J.; Wang, F. Transcriptome analysis of the roots at early and late seedling stages using Illumina paired-end sequencing and development of EST-SSR markers in radish. Plant Cell Rep. 2012, 31, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Brohele, J.; Ellegren, H. Microsatellite evolution: Polarity of substitution within repeats and neutrality of flanking sequences. Proc. Royal Soc. Lond B Biol. Sci. 1999, 266, 825–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellegren, H. Heterogeneous mutation processes in human microsatellite DNA sequences. Nat. Genet. 2000, 24, 400–402. [Google Scholar] [CrossRef] [PubMed]
- Jakupciak, J.P.; Wells, R.D. Gene conversion (recombination) mediates expansions of CTG CAG repeats. J. Biol. Chem. 2000, 275, 40003–40013. [Google Scholar] [CrossRef] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Rozen, S.; Skaletsky, H.J. Bioinformatics Methods and Protocols: Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2000; pp. 365–386. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Waits, L.P.; Luikart, G.; Taberlet, P. Estimating the probability of identity among genotypes in natural populations: Cautions and guidelines. Mol. Ecol. 2001, 10, 249–256. [Google Scholar] [CrossRef]
- Evett, I.; Weir, B. Interpreting DNA Evidence: Statistical Genetics for Forensic Scientists; Sinauer Associates Inc.: Sunderland, UK, 1998; p. 278. [Google Scholar]
- Kloosterman, A.D.; Budowle, B.; Daselaar, P. PCR-amplification and detection of the human D1S80 VNTR locus. Int. J. Legal Med. 1993, 105, 257–264. [Google Scholar] [CrossRef]
- Ward, J.H., Jr. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]



{kind=link}
{kind=link}
| Locus | Accession No. | SSR Motifs | |
|---|---|---|---|
| D. carota | F. vulgare | ||
| GSSR-16 | Fj816126 | (TG)9 tacgc (ATGT)3 | (AT)3 |
| GSSR-35 | Fj816145 | (GA)13 | (GA)9 |
| GSSR-97 | Fj816206 | (GA)8(AG)7 aagtattcca(AG)6(GA)7 | (GA)4; (AG)11 |
| GSSR-138 | Fj816246 | (GT)5 ata (GT)7(AG)21 | (GA)3 |
| GSSR-154 | Fj816262 | (TC)11 | (TC)5 (TC)13 |
| BSSR-14 | FJ816268 | (TTA)4 | (TTA)3 |
| BSSR-53 | Fj148355 | (AT)8 | (TG)3 (TA)4 |
| BSSR-59 | Fj148394 | (TA)7 | (TA)5 |
| BSSR-75 | Fj148202 | (TAC)5 | (GCA)3; (TAT)3; (CTA)4 |
| BSSR-91 | Fj148825 | (GGT)4 gc(TTG)4(TTG)4 | (TTG)3(TTG)4 |
| SSR ID | Primer ID | Primer Sequences (5′-3′) | SSR Motif (5′-3′) | Annealing T (°C) |
|---|---|---|---|---|
| GSSR-16 * | P080 P081 | Fwd: ACTTTTGTTCCTGCATTACACAGT Rev: TGTGATGTTTGCAGGACATGG | (AT)3 | 59 |
| GSSR-35 * | P082 P083 | Fwd: TGCGCTCAGTCAATTGATTTACT Rev: TCAGACACCCCTTTGTTGTTTTC | (GA)5; (GA)3; (GA)9; (GA)3 | 61 |
| GSSR-97 | P084 P043 | Fwd: GGCAAAGAAACAGATTTGGAGA Rev: CTGCCCTAGCATCAAAACAAAC | (GA)4; (AG)11 | 61 |
| GSSR-138 * | P085 P086 | Fwd: CCTCTTGCTGTTGTTGGTGA Rev: CCGTGGAAAGTCAGAATCATC | (GA)3 | 60 |
| GSSR-154 | P064 P065 | Fwd: CTTATATGTGATGGCGTCGAAA Rev: GACTGCACCGCTCCTAACTC | (TC)5 (TC)13 | 59 |
| BSSR-14 | P08 9P067 | Fwd: TACCCATAACTCAAGTTGGATAATTC Rev: AATGTCTAAACCCACTGATTTAAAAG | (TTA)3 | 58 |
| BSSR-59 | P070 P090 | Fwd: GATGAACTTAGATCATGTGGGACT Rev: GTACAGCTGGTCAATCCGATG | (TA)5 | 58 |
| BSSR-75 | P091 P073 | Fwd: ATGAAAGCAGGGATAAAAGTATCCAG Rev: AGAAGAAGGATTCAAGAAATGGCACA | (GCA)3; (TAT)3; (CTA)4 | 62 |
| BSSR-91 | P092 P075 | Fwd: AGCTTCAACAGGGTCTTGAGTTC Rev: CTTGGCTCAACTTATGCAACTTCT | (TTG)3; (TTG)4 | 61 |
| Locus | Range of Allele Size (bp) | Na | Ne | Ho | He | F | PIC | NAF | PID Unrelated | PIDsib |
|---|---|---|---|---|---|---|---|---|---|---|
| GSSR-16 | 293–305 | 3 | 2.12 | 0.050 | 0.529 | 0.906 | 0.406 | 0.8298 | 0.3439 | 0.5778 |
| GSSR-35 | 249–253 | 3 | 1.17 | 0.150 | 0.145 | 0.035 | 0.136 | 0.0300 | 0.7428 | 0.8651 |
| GSSR-97 | 256–268 | 5 | 3.65 | 0.650 | 0.726 | 0.105 | 0.656 | 0.0194 | 0.1372 | 0.4305 |
| GSSR-138 | 390 | 1 | 1.00 | 0.000 | 0.000 | - | 0.000 | ND | 1.0000 | 1.0000 |
| GSSR-154 | 300–370 | 12 | 6.49 | 0.400 | 0.846 | 0.527 | 0.810 | 0.3365 | 0.0456 | 0.3489 |
| BSSR-14 | 222 | 1 | 1.00 | 0.000 | 0.000 | - | 0.000 | ND | 1.0000 | 1.0000 |
| BSSR-59 | 222–230 | 2 | 1.63 | 0.300 | 0.385 | 0.221 | 0.305 | 0.1111 | 0.4609 | 0.6777 |
| BSSR-75 | 174–242 | 2 | 1.11 | 0.000 | 0.097 | 1.000 | 0.090 | 0.7350 | 0.8235 | 0.9084 |
| BSSR-91 | 174 | 1 | 1.00 | 0.000 | 0.000 | - | 0.000 | ND | 1.0000 | 1.0000 |
| Mean | - | 3.33 | 2.13 | 0.172 | 0.303 | 0.454 | 0.267 | - | - | - |
| Total | - | 30 | - | - | - | - | - | - | 6.06×10−4 | 4.62×10−2 |
| Varieties | Company | Varieties | Company |
|---|---|---|---|
| Bianco Extra | Fratelli Ingegnoli | Wadenromen | FOUR - Blumen Group |
| Carmo | Fratelli Ingegnoli | EZ#1 | Enza Zaden |
| Chiarino | Fratelli Ingegnoli | EZ#2 | Enza Zaden |
| Cristal | Fratelli Ingegnoli | EZ#3 | Enza Zaden |
| Mantovano | FOUR - Blumen Group | EZ#4 | Enza Zaden |
| Montebianco | Fratelli Ingegnoli | EZ#5 | Enza Zaden |
| Romanesco | Dom Sementi - SDD | EZ#6 | Enza Zaden |
| Romanesco sel. Circeo | FOUR - Blumen Group | EZ#7 | Enza Zaden |
| Selvatico | Fratelli Ingegnoli | EZ#8 | Enza Zaden |
| Tauro | Fratelli Ingegnoli | EZ#9 | Enza Zaden |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aiello, D.; Ferradini, N.; Torelli, L.; Volpi, C.; Lambalk, J.; Russi, L.; Albertini, E. Evaluation of Cross-Species Transferability of SSR Markers in Foeniculum vulgare. Plants 2020, 9, 175. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9020175
Aiello D, Ferradini N, Torelli L, Volpi C, Lambalk J, Russi L, Albertini E. Evaluation of Cross-Species Transferability of SSR Markers in Foeniculum vulgare. Plants. 2020; 9(2):175. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9020175
Chicago/Turabian StyleAiello, Domenico, Nicoletta Ferradini, Lorenzo Torelli, Chiara Volpi, Joep Lambalk, Luigi Russi, and Emidio Albertini. 2020. "Evaluation of Cross-Species Transferability of SSR Markers in Foeniculum vulgare" Plants 9, no. 2: 175. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9020175