Naringenin Regulates Doxorubicin-Induced Liver Dysfunction: Impact on Oxidative Stress and Inflammation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals and Experimental Design
2.3. Post Mitochondrial Supernatant Preparation
2.4. Estimation of Serum Enzyme Activities
2.4.1. Estimation of Serum Aspartate Aminotransferase (AST) and Alanine Aminotransferases (ALT)
2.4.2. Assay for Lactate Dehydrogenase Activity
2.4.3. Estimation of Protein
2.4.4. Estimation of Alkaline Phosphatase (ALP)
2.5. Measurement of ROS
2.6. Estimation of Lipid Peroxidation
2.7. Estimation of Antioxidant Enzyme Armory
2.7.1. Assay for Superoxide Dismutase Activity (SOD)
2.7.2. Catalase Activity
2.7.3. Estimation of Glutathione (GSH)
2.7.4. Glutathione Reductase (GR) Activity
2.7.5. Glutathione Peroxidase Activity
2.8. Measurement of Nitric Oxide (NO)
2.9. Assay for Hydrogen Peroxide (H2O2)
2.10. Histopathological Examination
2.11. Immunostaining
2.12. Inflammatory Mediators
2.13. Ethical Statement
2.14. Statistical Analysis
3. Results
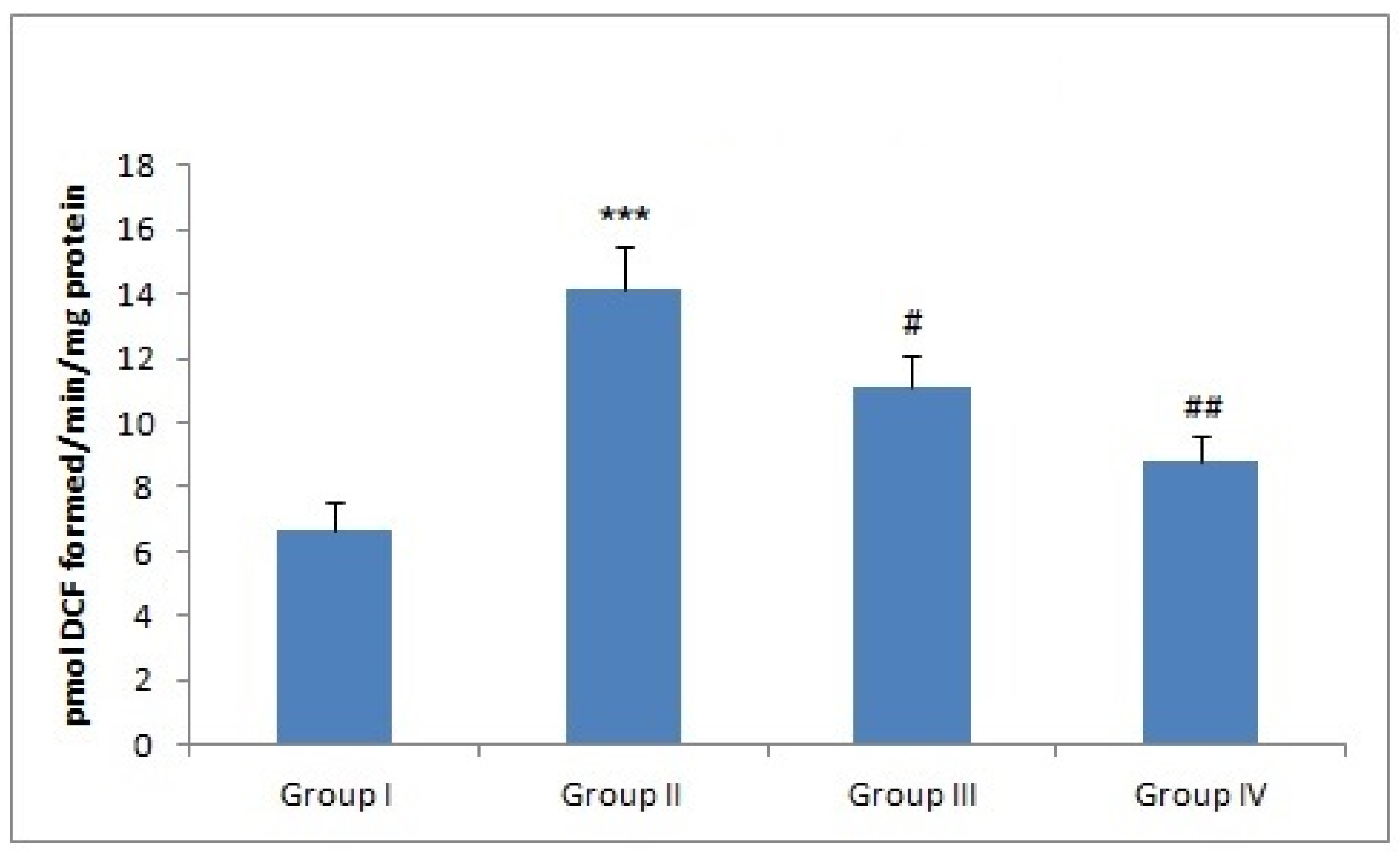
3.1. Effect of Naringenin Treatment on the ROS Levels
3.2. Effect of Naringenin Treatment on the Antioxidant Enzyme System
3.3. Effect of Naringenin Treatment on H2O2 Levels
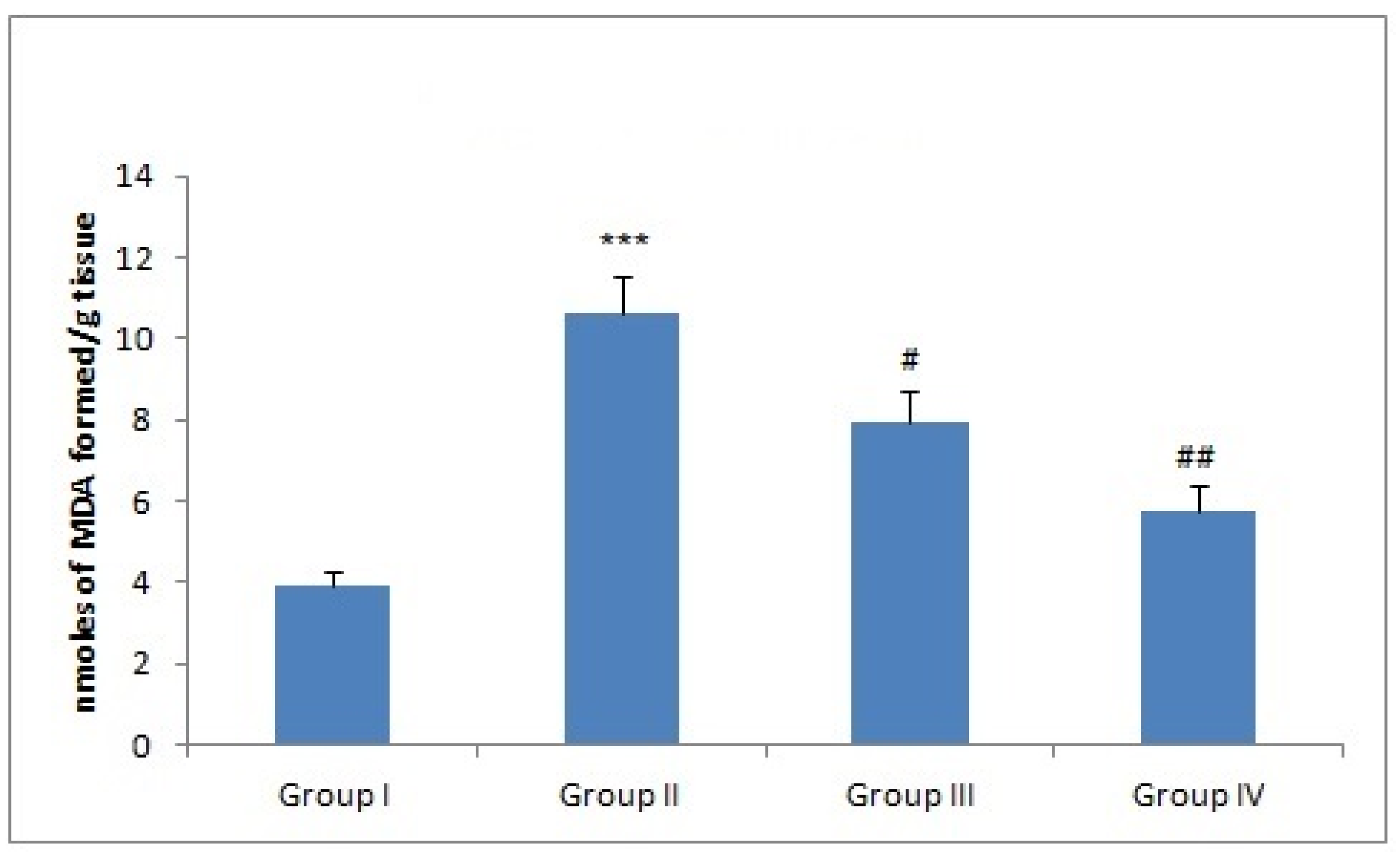
3.4. Effect of Naringenin Treatment on MDA Levels
3.5. Naringenin Treatment Alleviates Serum Toxicity Parameters in Dox-Induced Liver Toxicity
3.6. Effect of Naringenin and Dox Treatment on NF-κB and Other Inflammatory Mediators
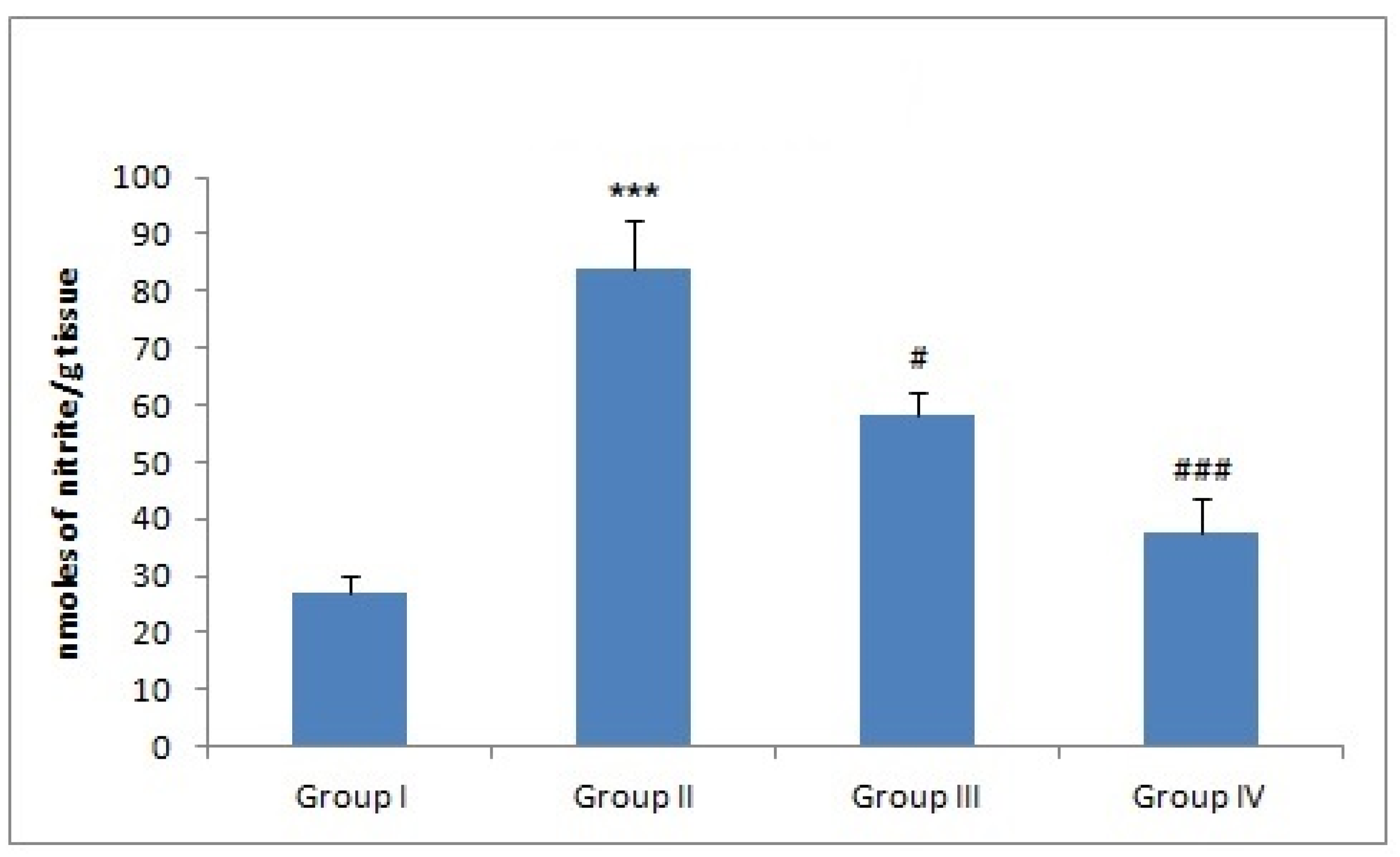
3.7. Effect of Naringenin on NO Production
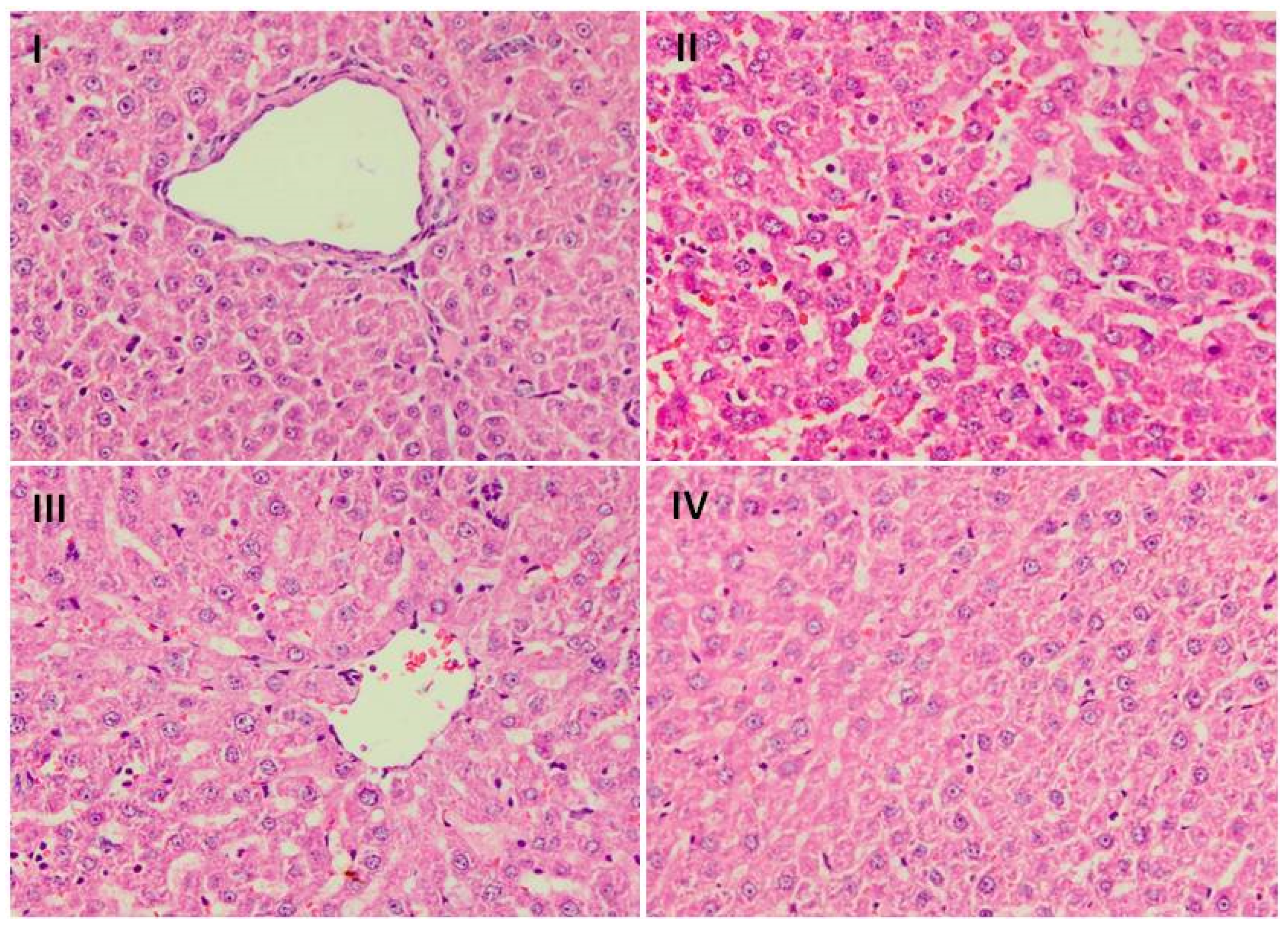
3.8. Effect of Naringenin on Liver Histology
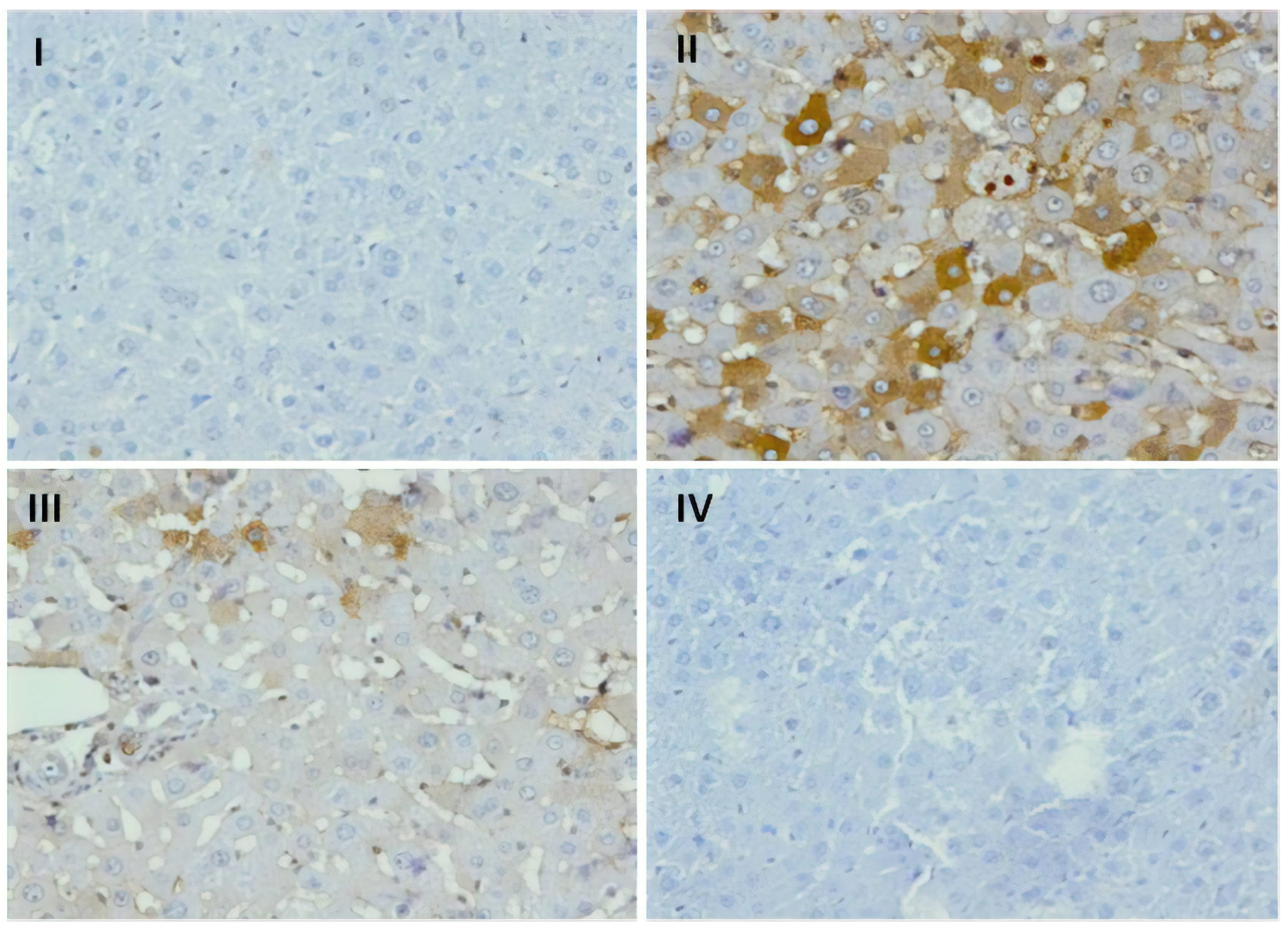
3.9. Effect of Naringenin on Expression of COX-2
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, Y.; Mei, X.; Yuan, J.; Lu, W.; Li, B.; Xu, D. Taurine zinc solid dispersions attenuate Doxorubicin-induced hepatotoxicity and cardiotoxicity in rats. Toxicol. Appl. Pharm. 2015, 289, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jacevic, V.; Djordjevic, A.; Srdjenovic, B.; Milic-Tores, V.; Segrt, Z.; Dragojevic-Simic, V.; Kuca, K. Fullerenol nanoparticles prevents Doxorubicin-induced acute hepatotoxicity in rats. Exp. Mol. Pathol. 2017, 102, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Damodar, G.; Smitha, T.; Gopinath, S.; Vijayakumar, S.; Rao, Y. An evaluation of hepatotoxicity in breast cancer patients receiving injection Doxorubicin. Ann. Med. Health Sci. Res. 2014, 4, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.G.; Attia, N.; Hashem, F.Z.A.; El-Heneidy, M.A.R. Cerium oxide nanoparticles: In pursuit of liver protection against Doxorubicin-induced injury in rats. Biomed. Pharm. 2018, 103, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Camaggi, C.M.; Comparsi, R.; Strocchi, E.; Testoni, F.; Angelelli, B.; Pannuti, F. Epirubicin and Doxorubicin comparative metabolism and pharmacokinetics. A cross-over study. Cancer Chemother. Pharm. 1988, 21, 221–228. [Google Scholar] [CrossRef]
- Rehman, M.U.; Tahir, M.; Khan, A.Q. D-limonene suppresses Doxorubicin-induced oxidative stress and inflammation via repression of COX-2, iNOS, and NFκB in kidneys of Wistar rats. Exp. Biol. Med. 2014, 239, 465–476. [Google Scholar] [CrossRef]
- Omobowale, T.O.; Oyagbemi, A.A.; Ajufo, U.E.; Adejumobi, O.A.; Ola-Davies, O.E.; Adedapo, A.A.; Yakubu, M.A. Ameliorative effect of gallic acid in Doxorubicin-induced hepatotoxicity in wistar rats through antioxidant defence system. J. Diet. Suppl. 2018, 15, 183–196. [Google Scholar] [CrossRef]
- Gutteridge, J.M. Lipid peroxidation and possible hydroxyl radical formation stimulated by the self-reduction of a Doxorubicin–iron (III) complex. Biochem. Pharm. 1984, 33, 1725–1728. [Google Scholar] [CrossRef]
- Saad, S.Y.; Najjar, T.A.; Al-Rykaby, A.C. The preventive role of deferoxamine against acute Doxorubicin-induced cardiac, renal and hepatic toxicity in rats. Pharm. Res. 2011, 43, 211–218. [Google Scholar] [CrossRef]
- Hajra, S.; Patra, A.R.; Basu, A.; Bhattacharya, S. Prevention of Doxorubicin (DOX)-induced genotoxicity and cardiotoxicity: Effect of plant derived small molecule indole-3-carbinol (I3C) on oxidative stress and inflammation. Biomed. Pharm. 2018, 101, 228–243. [Google Scholar] [CrossRef]
- Rashid, S.; Ali, N.; Nafees, S.; Ahmad, S.T.; Arjumand, W.; Hasan, S.K.; Sultana, S. Alleviation of Doxorubicin-induced nephrotoxicity and hepatotoxicity by chrysin in Wistar rats. Toxicol. Mech. Methods 2013, 23, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, J.; Jesudoss, V.A.S.; Menon, V.P.; Namasivayam, N. Anti-inflammatory role of naringenin in rats with ethanol induced liver injury. Toxicol. Mech. Methods 2012, 22, 568–576. [Google Scholar]
- Aluise, C.D.; Miriyala, S.; Noel, T. Butterfield DA 2-Mercaptoethane sulfonate prevents Doxorubicin-induced plasma protein oxidation and TNF-a release: Implications for the reactive oxygen species-mediated mechanisms of chemobrain. Free Radic. Biol. Med. 2011, 50, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.; Nafees, S.; Vafa, A.; Afzal, S.M.; Ali, N.; Rehman, M.U.; Hasan, S.K.; Siddiqi, A.; Barnwal, P.; Majed, F.; et al. Inhibition of precancerous lesions development in kidneys by chrysin via regulating hyperproliferation, inflammation and apoptosis at pre clinical stage. Arch. Biochem. Biophys. 2016, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Zhang, X. Interleukin-10: New perspectives on an old cytokine. Immunol. Rev. 2008, 226, 205–218. [Google Scholar] [CrossRef]
- Song, S.; Chu, L.; Liang, H.; Chen, J.; Liang, J.; Huang, Z.; Zhang, B.; Chen, X. Protective Effects of Dioscin Against Doxorubicin-Induced Hepatotoxicity Via Regulation of Sirt1/ FOXO1/NF-κb Signal. Front. Pharm. 2019, 10, 1030. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Rajesh, M.; Bátkai, S. Role of superoxide, nitric oxide, and peroxynitrite in Doxorubicin-induced cell death in vivo and in vitro. Am. J. Physiol. 2019, 296, 1466–1483. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Wang, C.; Kong, Q.; Wu, X.; Lu, J.J.; Chen, X. Recent progress in Doxorubicin induced cardiotoxicity and protective potential of natural products. Phytomedicine 2018, 1, 125–139. [Google Scholar] [CrossRef]
- Kocahan, S.; Dogan, Z.; Erdemli, E.; Taskin, E. Protective Effect of Quercetin Against Oxidative Stressinduced Toxicity Associated with Doxorubicin and Cyclophosphamide in Rat Kidney and Liver Tissue. Iran J. Kidney Dis. 2017, 11, 124–131. [Google Scholar]
- Li, Y.; Yang, D.; Wang, Y.; Li, Z.; Zhu, C. Co-delivery Doxorubicin and silybin for antihepatoma via enhanced oral hepatic-targeted efficiency. Int. J. Nanomed. 2019, 14, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Patel, N.; Joseph, C.; Corcoran, G.B.; Ray, S.D. Silymarin modulates Doxorubicin-induced oxidative stress, Bcl-xL and p53 expression while pre-venting apoptotic and necrotic cell death in the liver. Toxicol. Appl. Pharm. 2010, 245, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Oktem, G.; Uysal, A.; Oral, O. Resveratrol attenuates Doxorubicin-induced cellular damage by modulating nitric oxide and apoptosis. Exp. Toxicol. Pathol. 2012, 64, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Holden, J.M.; Bhagwat, S.A.; Patterson, K.Y. Development of a multinutrient data quality evaluation system. J. Food Comp. Anal. 2002, 15, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Lou, C.; Zhang, F.; Yang, M. Naringenin Decreases Invasiveness and Metastasis by Inhibiting TGF-β-Induced Epithelial to Mesenchymal Transition in Pancreatic Cancer Cells. PLoS ONE 2012, 7, e50956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, T.; Wang, Y.; Ma, Y.; Su, W.; Bai, Y.; Zhao, P. A rapid LC/ MS/MS quantitation assay for naringin and its two metabolites in rat’s plasma. J. Pharm. Biomed Anal. 2006, 40, 454–459. [Google Scholar] [CrossRef]
- Raso, G.M.; Meli, R.; Di Carlo, G.; Pacilio, M.; Di Carlo, R. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 expression by flavonoids in macrophage J774A.1. Life Sci. 2001, 68, 921–931. [Google Scholar] [CrossRef]
- Rehman, M.U.; Mir, M.U.; Farooq, A. Naringenin (4,5,7-trihydroxyflavanone) suppresses the development of precancerous lesions via controlling hyperproliferation and inflammation in the colon of Wistar rats. Environ. Toxicol. 2018, 33, 422–435. [Google Scholar] [CrossRef]
- Rehman, M.U.; Tahir, M.; Khan, A.Q.; Khan, R.; Lateef, A.; Hamiza, O.O.; Ali, F.; Sultana, S. Diosmin protects against trichloroethylene-induced renal injury in Wistar rats: plausible role of p53, Bax and caspases. Br. J. Nutr. 2013, 110, 699–710. [Google Scholar] [CrossRef] [Green Version]
- Rehman, M.U.; Ali, N.; Rashid, S.; Jain, T.; Nafees, S.; Tahir, M.; Khan, A.Q.; Lateef, A.; Khan, R.; Hamiza, O.O.; et al. Alleviation of hepatic injury by chrysin in cisplatin administered rats: probable role of oxidative and inflammatory markers. Pharm. Rep. 2014, 66, 1050–1059. [Google Scholar] [CrossRef]
- Rashid, S.; Nafees, S.; Siddiqi, A.; Vafa, A.; Afzal, S.M.; Parveen, R.; Ali, N.; Hasan, S.K.; Barnwal, P.; Shahid, A.; et al. Partial protection by 18β Glycrrhetinic acid against Cisplatin induced oxidative intestinal damage in wistar rats: Possible role of NFkB and caspases. Pharm. Rep. 2017, 69, 1007–1013. [Google Scholar] [CrossRef]
- Rashid, S.; Ali, N.; Nafees, S.; Hasan, S.K.; Sultana, S. Mitigation of 5-Fluorouracil induced renal toxicity by chrysin via targeting oxidative stress and apoptosis in wistar rats. Food Chem. Toxicol. 2014, 66, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Reitman, S.; Frankel, S.A. Colorimetric method for determination of serum glutamic oxaloacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, A.; In-Colowick, A.P.; Kaplan, N.O. Methods of Enzymology; Academic Press: New York, NY, USA, 1963; Volume VI. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin Phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Liua, C.M.; Mab, J.Q.; Suna, J.Q. Quercetin protects the rat kidney against oxidative stress-mediated DNA damage and apoptosis induced by lead. Environ. Toxicol. Pharm. 2010, 30, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.R.; Colby, H.D.; Miles, P.R. Cytosolic factors which affect microsomal lipid peroxidation in lung and liver. Arch. Biochem. Biophys. 1981, 206, 296–304. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Claiborne, A. Catalase activity. In CRC Handbook of Methods in Oxygen Radical Research; Greenwald, R.A., Ed.; CRC: Boca Raton, RA, USA, 1985; pp. 283–284. [Google Scholar]
- Mohandas, M.; Marshall, J.J.; Duggin, G.G.; Horvath, J.S.; Tiller, D. Differential distribution of glutathione and glutathione related enzymes in rabbit kidney. Cancer Res. 1984, 44, 5086–5091. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15 N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Pick, E.; Mizel, D. Rapid microassays for the measurement of superoxide and hydrogen peroxide production by macrophages in culture using an automatic enzyme immunoassay reader. J. Immunol. Methods 1981, 46, 211–226. [Google Scholar] [CrossRef]
- Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A. Naringenin ameliorates daunorubicin induced nephrotoxicity by mitigating AT1R, ERK1/2-NFkappaB p65 mediated inflammation. Int. Immunopharmacol 2015, 28, 154–159. [Google Scholar] [CrossRef]
- Prsa, P.; Karademir, B.; Biçim, G. The potential use of natural products to negate hepatic, renal and neuronal toxicity induced by cancer therapeutics. Biochem. Pharm. 2019, 19, 30226-6. [Google Scholar]
- Khan, R.; Rehman, M.U.; Khan, A.Q.; Tahir, M.; Sultana, S. Glycyrrhizic acid suppresses 1,2-dimethylhydrazine-induced colon tumorigenesis in Wistar rats: Alleviation of inflammatory, proliferation, angiogenic, and apoptotic markers. Environ. Toxicol. 2018, 12, 1272–1283. [Google Scholar] [CrossRef] [PubMed]
- Faghihzadeh, F.; Hekmatdoost, A.; Adibi, P. Resveratrol and liver: A systematic review. J. Res. Med. Sci. 2015, 20, 797–810. [Google Scholar]
- Ezzat, S.M.; Salama, M.M.; Seif El-Din, S.H.; Saleh, S.; El-Lakkany, N.M.; Hammam, O.A.; Salem, M.B.; Botros, S.S. Metabolic profile and hepatoprotective activity of the anthocyanin-rich extract of Hibiscus sabdariffa calyces. Pharm. Biol. 2016, 54, 3172–3181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lila, M.A. Anthocyanins and Human Health: An In Vitro Investigative Approach. J. Biomed. Biotechnol. 2004, 5, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Koyuncu, I.; Kocyigit, A.; Gonel, A.; Arslan, E.; Durgun, M. The Protective Effect of Naringenin-Oxime on Cisplatin-Induced Toxicity in Rats. Biochem. Res. Int. 2017, 2017, 9478958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Kuzu, M.; Yıldırım, S.; Kandemir, F.M.; Küçükler, S.; Çağlayan, C.; Türk, E.; Dörtbudak, M.B. Protective effect of morin on Doxorubicin-induced hepatorenal toxicity in rats. Chem. Biol. Interact 2019, 1, 89–100. [Google Scholar] [CrossRef]
- Subburaman, S.; Ganesan, K.; Ramachandran, M. Protective role of naringenin against Doxorubicin-induced cardiotoxicity in a rat model: histopathology and mRNA expression profile studies. J. Environ. Pathol. Toxicol. Oncol. 2014, 33, 363–376. [Google Scholar] [CrossRef]
- Hernández-Aquino, E.; Muriel, P. Beneficial effects of naringenin in liver diseases: Molecular mechanisms. World J. Gastroenterol. 2018, 24, 1679–1707. [Google Scholar] [CrossRef]
- Edwardson, D.W.; Narendrula, R.; Chewchuk, S.; Mispel-Beyer, K.; Mapletoft, J.P.; Parissenti, A.M. Role of Drug Metabolism in the Cytotoxicity and Clinical Efficacy of Anthracyclines. Curr. Drug Metab. 2015, 16, 412–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afsar, T.; Razak, S.; Almajwal, A. Effect of Acacia hydaspica R. Parker extract on lipid peroxidation, antioxidant status, liver function test and histopathology in Doxorubicin treated rats. Lipids Health Dis. 2019, 18, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Zhang, J.; Tong, N.; Chen, Y.; Luo, Y. Protective effects of berberine on Doxorubicin-induced hepatotoxicity in mice. Biol. Pharm. Bull. 2012, 35, 796–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henninger, C.; Huelsenbeck, J.; Huelsenbeck, S.; Grösch, S.; Schad, A.; Lackner, K.J.; Kaina, B.; Fritz, G. The lipid lowering drug lovastatin protects against Doxorubicin-induced hepatotoxicity. Toxicol. Appl. Pharm. 2012, 261, 66–73. [Google Scholar] [CrossRef]
- Osama, M.A.; Hanaa, I.F.; Heba, Y.A. The Preventive Effects and the Mechanisms of Action of Navel Orange Peel Hydroethanolic Extract, Naringin, and Naringenin in N-Acetyl-p-aminophenol-Induced Liver Injury in Wistar Rats. Oxid. Med. cell longev. 2019, 2745352, 1–19. [Google Scholar]
- Liu, Z.; Huang, P.; Law, S.; Tian, H.; Leung, W.; Xu, C. Preventive Effect of Curcumin Against Chemotherapy-Induced Side-Effects. Front. Pharm. 2018, 9, 1374. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; SoonSand Moon, J.O. The flavonoid naringenin inhibits dimethylnitrosamine induced liver damage in rats. Biol. Pharm. Bull 2004, 27, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Kwatra, M.; Kumar, V.; Jangra, A.; Mishra, M.; Ahmed, S.; Ghosh, P.; Vohora, D.; Khanam, R. Ameliorative effect of naringin against Doxorubicin-induced acute cardiac toxicity in rats. Pharm. Biol. 2016, 54, 637–647. [Google Scholar] [CrossRef]
- Doroshow, J.H. Effect of anthracycline antibiotics on oxygen radical formation in rat heart. Cancer Res. 1983, 43, 460–472. [Google Scholar]
- Han, X.; Ren, D.; Fan, P.; Shen, T.; Lou, H. Protective effects of naringenin-7-O-glucoside on Doxorubicin-induced apoptosis in H9C2 cells. Eur. J. Pharm. 2008, 581, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renugadevi, J.; Prabu, S.M. Naringenin protects against cadmium-induced oxidative renal dysfunction in rats. Toxicology 2009, 4, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Rashmi, R.; Bojan Magesh, S.; Mohanram Ramkumar, K.; Suryanarayanan, S.; Venkata SubbaRao, M. Antioxidant Potential of Naringenin Helps to Protect Liver Tissue from Streptozotocin-Induced Damage. Rep. Biochem Mol. Biol. 2018, 7, 76–84. [Google Scholar] [PubMed]
- Lee, C.T.; Yu, L.E.; Wang, J.Y. Nitric Oxide. antioxidant as a potential strategy to attenuate the oxidative/nitrosative stress induced by hydrogen peroxide plus nitric oxide in cultured neurons. Nitr. Oxide 2016, 1, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Mokni, M.; Hamlaoui, S.; Kadri, S. Efficacy of grape seed and skin extract against Doxorubicin-induced oxidative stress in rat liver. Pak. J. Pharm. Sci. 2015, 28, 1971–1978. [Google Scholar] [PubMed]
- Szwed, M.; Wrona, D.; Kania, K.D.; Koceva-Chyla, A.; Marczak, A. Doxorubicin-transferrin conjugate triggers pro-oxidative disorders in solid tumor cells. Toxicol. In Vitro 2016, 31, 60–71. [Google Scholar] [CrossRef]
- Chen, Y.; Jungsuwadee, P.; Vore, M.; Butterfield, D.A.; Clair, D.K. Collateral damage in cancer chemotherapy: oxidative stress in non-targeted tissues. Mol. Interv. 2007, 7, 147–156. [Google Scholar] [CrossRef]
- Kabel, A.M. Zinc/alogliptin combination attenuates testicular toxicity induced by Doxorubicin in rats: role of oxidative stress, apoptosis and TGF-b1/NF-kB signaling. Biomed Pharm. 2018, 97, 439e49. [Google Scholar] [CrossRef]
- Sun, Z.; Schriewer, J.; Tang, M. The TGF-b pathway mediates Doxorubicin effects on cardiac endothelial cells. J. Mol. Cell Cardiol. 2016, 90, 129e38. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Treatment |
|---|---|
| Group I | Animals received normal diet for 20 consecutive days. |
| Group II | A single intraperitoneal injection of Dox at the dose of 20 mg/kg body weight on 20th day along with normal diet for 20 consecutive days. |
| Group III | Animals were first treated with normal diet and naringenin (50 mg/kg body weight) for 20 days before the Dox treatment (as in group II). |
| Group IV | Animals were first treated with normal diet and naringenin (100 mg/kg body weight) for 20 days before the Dox treatment (as in group II). |
| Group I | Group II | Group III | Group IV | |
|---|---|---|---|---|
| Reduced Glutathione (GSH; nmol mg protein) | 333.10 ± 15.1 | 146.12 ± 16.2 *** | 192.32 ± 13.4 # | 307.42 ± 29.7 ### |
| Oxidized Glutathione (GSSG; nmol mg protein) | 43.42 ± 3.02 | 89.32 ± 7.71 *** | 64.34 ± 5.32 # | 46.03 ± 3.38 ### |
| GPx (nmol/min/mg protein) | 190.39 ± 19.3 | 79.15 ± 7.07 *** | 147.33 ± 12.1 ## | 159.65 ± 11.2 ## |
| GR (nmol min/min/mg protein) | 227.61 ± 19.6 | 101.31 ± 10.5 *** | 165.09 ± 13.5 ## | 210.20 ± 22.8 ### |
| SOD (units/min/mg protein) | 14.93 ± 2.87 | 4.83 ± 0.71 *** | 8.53 ± 0.82 # | 10.14 ± 1.32 ### |
| Catalase (nmol H2O2 consumed/min/mg protein) | 9.41 ± 0.81 | 2.01 ± 0.11 *** | 5.82 ± 0.51 ### | 7.32 ± 0.64###b |
| H2O2 (nmol of H2O2 /g tissue) | 192.1 ± 18.7 | 407.3 ± 19.2 *** | 333.2 ± 26.1 # | 212.1 ± 19.5 ### |
| Treatment Regimen | ALT (IU/L) | AST (IU/L) | ALP (IU/L) | Total Protein (IU/L) | LDH (nmol NADH oxidized/min/mg Protein) |
|---|---|---|---|---|---|
| Group I | 67.73 ± 4.12 | 108.3 ± 9.9 | 103. 4± 9.11 | 6.01 ± 0.49 | 112.91 ± 15.3 |
| Group II | 119.4 ± 6.42 *** | 300.6 ± 14.9 *** | 201.3 ± 8.20 *** | 3.13 ± 0.19 *** | 302.23 ± 19.7 *** |
| Group III | 99.21 ± 7.92 # | 210.2 ± 16.5 # | 172.2 ± 9.32 # | 3.87 ± 0.22 ns | 198.76 ± 25.9 # |
| Group IV | 80.31 ± 4.64 ## | 151.1 ± 7.83 ### | 115.9 ± 10.8 ### | 5.01 ± 0.49 ## | 151.03 ± 15.9 ## |
| Group I | Group II | Group III | Group IV | |
|---|---|---|---|---|
| NFκ-B (pg/mL) | 901.62 ± 71.6 | 2341.10 ± 61.5 *** | 1952.32 ± 38.2 # | 1102.32 ± 31.7 ### |
| TNF-α (pg/mL) | 247.30 ± 17.8 | 552.18 ± 27.2 *** | 321.01 ± 14.0 ## | 269.11 ± 19.2 ### |
| IL-1β (pg/mL) | 892.90 ± 41.4 | 1746.18 ± 61.0 *** | 1485.44 ± 70.1 # | 901.14 ± 38.3 ### |
| IL-6 (pg/mL) | 975.15 ± 80.2 | 2127.83 ± 57.1 *** | 1763.20 ± 44.1 # | 1001.62 ± 67.7 ### |
| TGF-β (pg/mL) | 259.18 ± 13.8 | 722.95 ± 42.1 *** | 431.38 ± 21.4 ## | 271.83 ± 31.8 ### |
| PGE-2 (pg/mL) | 139.97 ± 14.4 | 527.36 ± 32.0 *** | 154.11 ± 13.2 ### | 146.02 ± 13.0 ### |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wali, A.F.; Rashid, S.; Rashid, S.M.; Ansari, M.A.; Khan, M.R.; Haq, N.; Alhareth, D.Y.; Ahmad, A.; Rehman, M.U. Naringenin Regulates Doxorubicin-Induced Liver Dysfunction: Impact on Oxidative Stress and Inflammation. Plants 2020, 9, 550. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9040550
Wali AF, Rashid S, Rashid SM, Ansari MA, Khan MR, Haq N, Alhareth DY, Ahmad A, Rehman MU. Naringenin Regulates Doxorubicin-Induced Liver Dysfunction: Impact on Oxidative Stress and Inflammation. Plants. 2020; 9(4):550. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9040550
Chicago/Turabian StyleWali, Adil Farooq, Summya Rashid, Shahzada Mudasir Rashid, Mushtaq Ahmad Ansari, Mohammad Rashid Khan, Nazrul Haq, Dhafer Yahya Alhareth, Ajaz Ahmad, and Muneeb U. Rehman. 2020. "Naringenin Regulates Doxorubicin-Induced Liver Dysfunction: Impact on Oxidative Stress and Inflammation" Plants 9, no. 4: 550. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9040550