Foliar Application of an Amino Acid-Enriched Urea Fertilizer on ‘Greco’ Grapevines at Full Veraison Increases Berry Yeast-Assimilable Nitrogen Content

 , and
, and
Abstract
:1. Introduction
2. Results
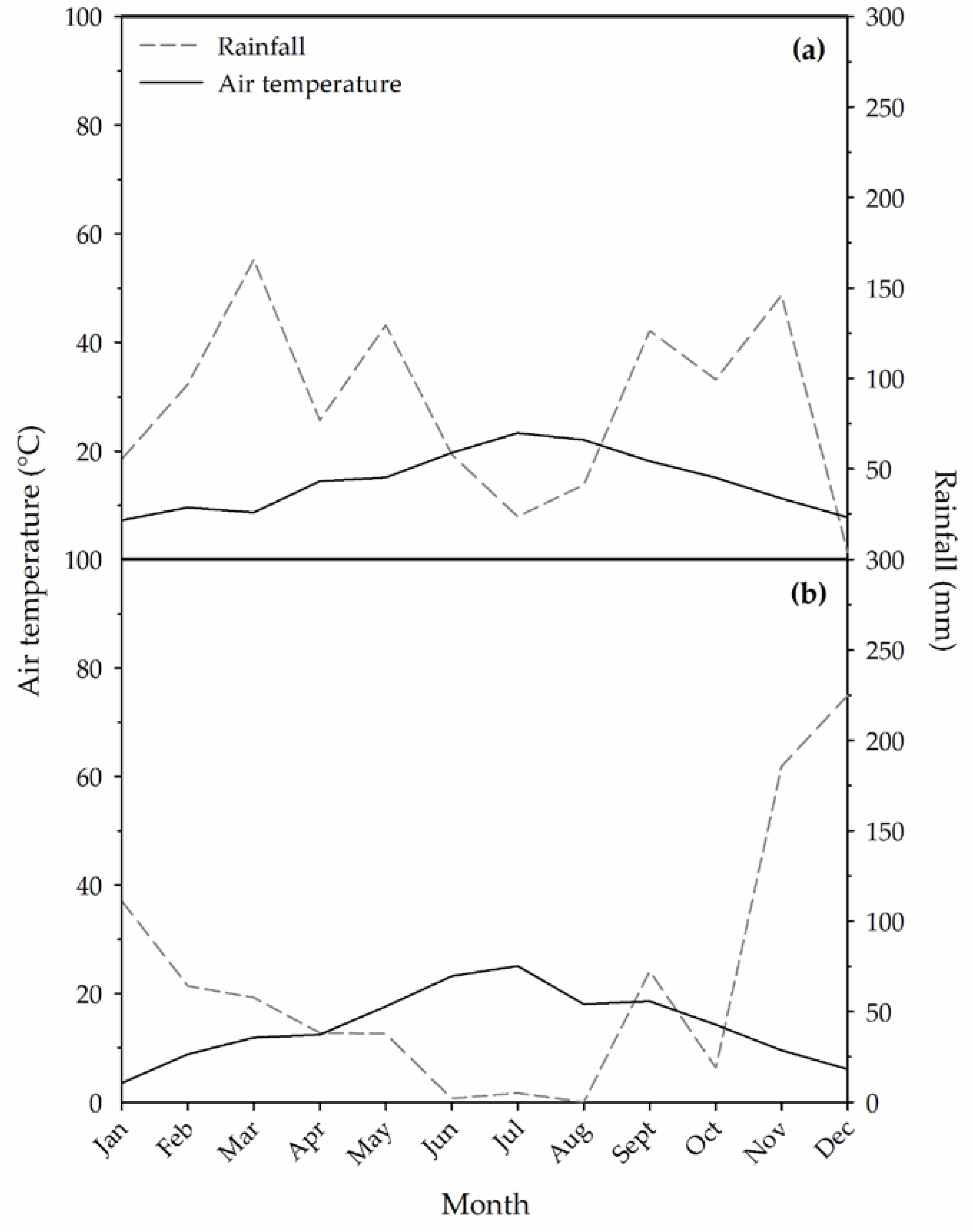
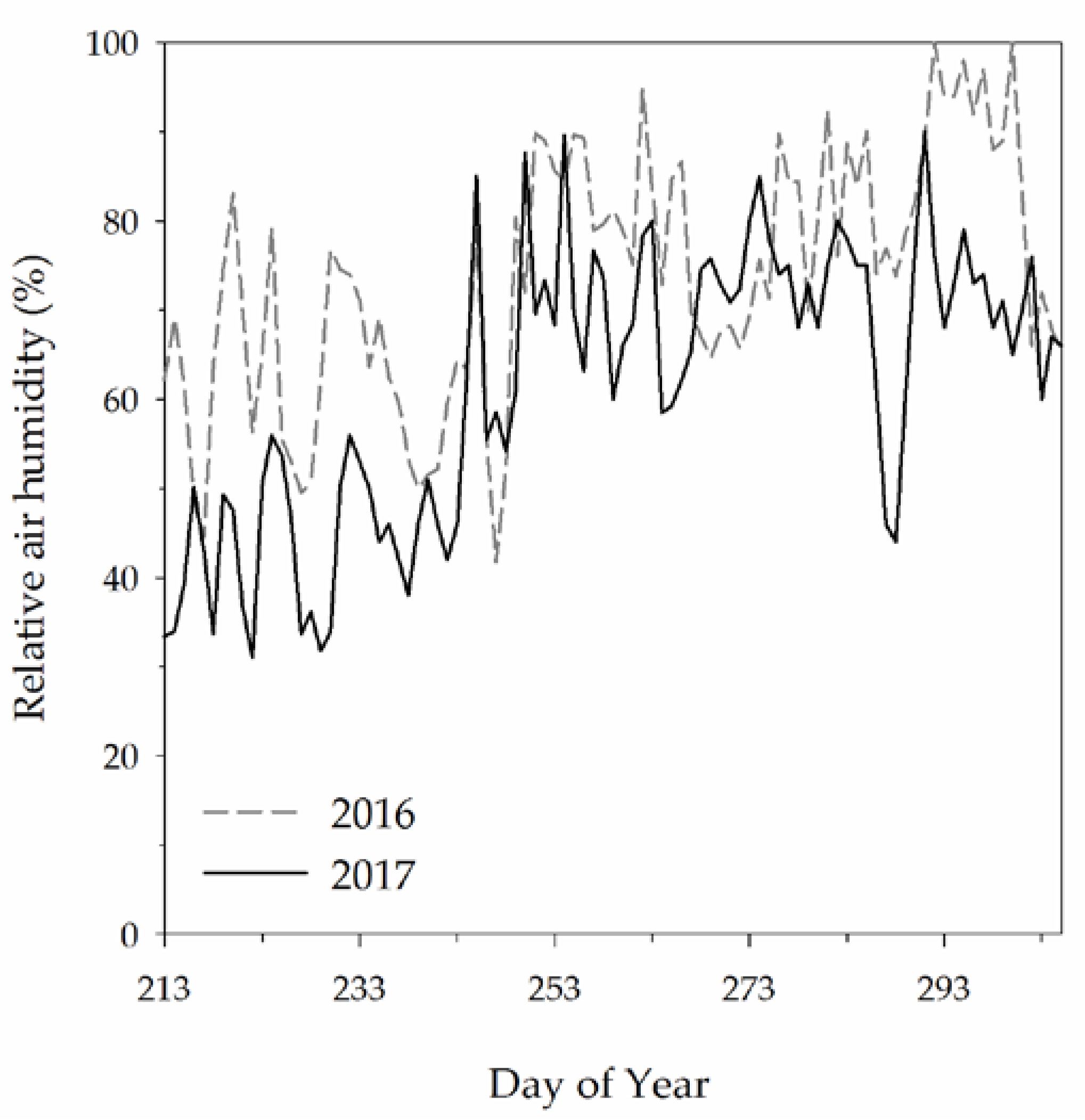
2.1. Climatic Data
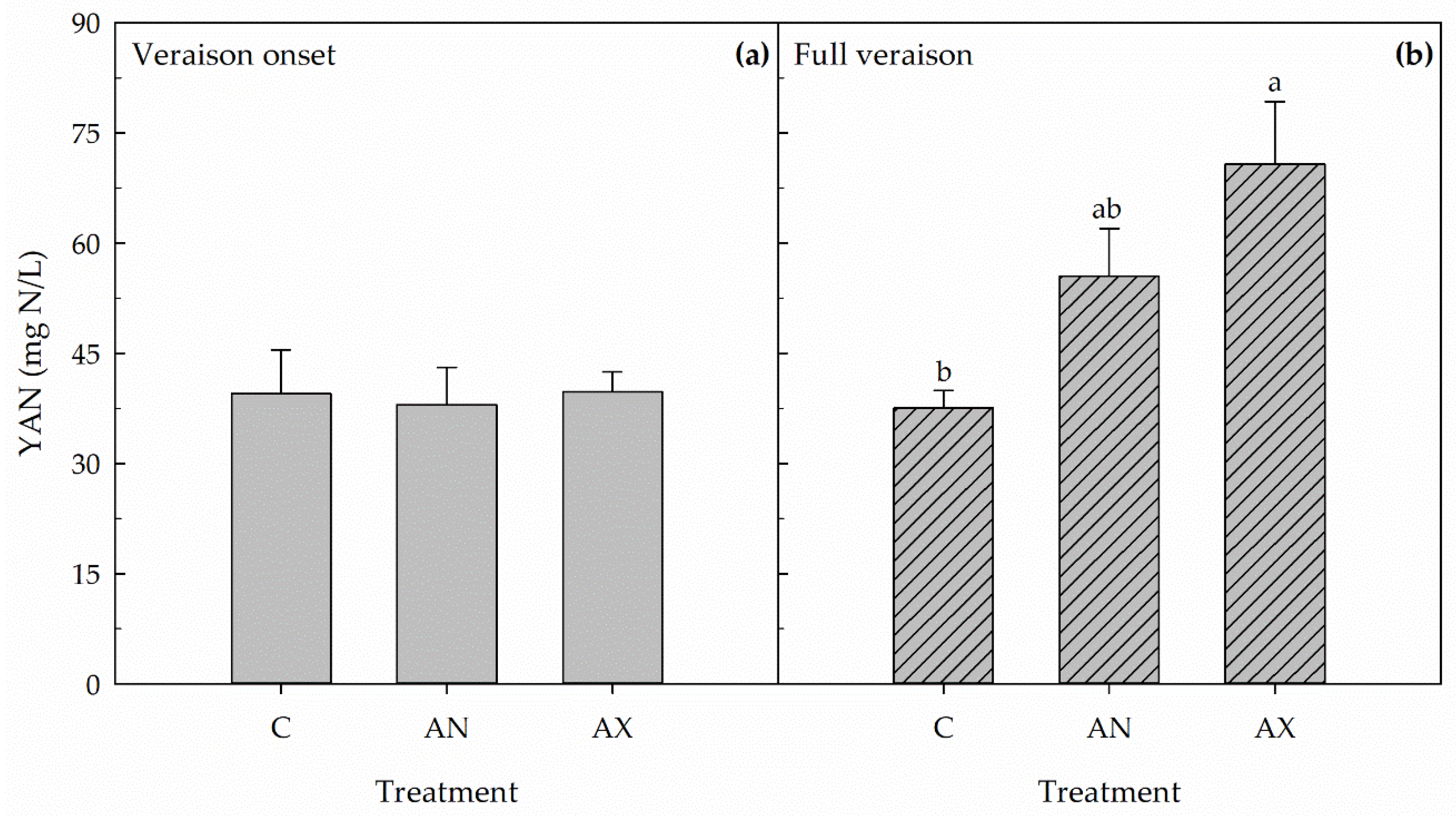
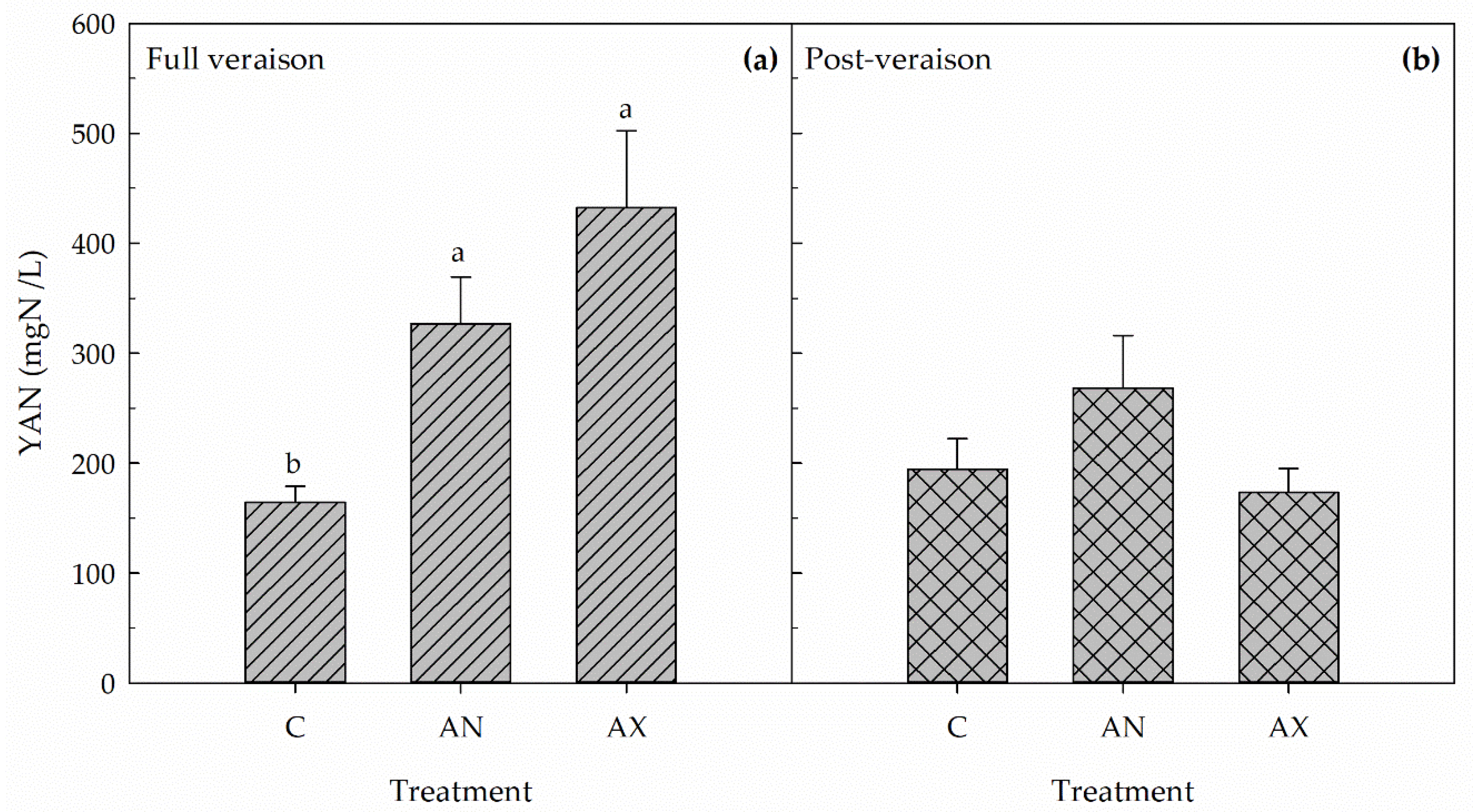
2.2. Yeast Assimilable Nitrogen (YAN) Concentration
2.3. Aminic and Ammonium N
2.4. Total Soluble Solids (TSS), pH, Malic Acid, and Total Acidity
3. Discussion
4. Materials and Methods
4.1. Experimental Site and Plant Material
4.2. Experimental Design and Treatments
4.3. Grape Composition at Harvest
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Poni, S.; Gatti, M.; Palliotti, A.; Dai, Z.; Duchene, E.; Truong, T.T.; Ferrara, G.; Matarrese, A.M.S.; Gallotta, A.; Bellincontro, A.; et al. Grapevine quality: A multiple choice issue. Sci. Hortic. 2018, 234, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Swiegers, J.H.; Pretorius, I.S. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Gutiérrez-Gamboa, G.; Fernández-Novales, J.; Pérez-Álvarez, E.P.; Diago, M.P. Towards the definition of optimal grape harvest time in Grenache grapevines: Nitrogenous maturity. Sci. Hortic. 2018, 239, 9–16. [Google Scholar] [CrossRef]
- Stines, A.P.; Grubb, J.; Gockowiak, H.; Henschke, P.A.; Høj, P.B.; van Heeswijck, R. Proline and arginine accumulation in developing berries of Vitis vinifera L. in Australian vineyards: Influence of vine cultivar, berry maturity and tissue type. Aust. J. Grape Wine Res. 2000, 6, 150–158. [Google Scholar] [CrossRef]
- Ough, C.S.; Cook, J.A.; Lider, L.A. Rootstock-scion interactions concerning wine making. II. Wine compositional and sensory changes attributed to rootstock and fertilizer level differences. Am. J. Enol. Vitic. 1968, 19, 254–265. [Google Scholar]
- Reynard, J.S.; Zufferey, V.; Nicol, G.C.; Murisier, F. Soil parameters impact the vine-fruit-wine continuum by altering vine nitrogen status. OENO One 2011, 45, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Nicolini, G.; Larcher, R.; Versini, G. Status of yeast assimilable nitrogen in Italian grape musts and effects of variety, ripening and vintage. Vitis 2004, 43, 89–96. [Google Scholar]
- Pérez-Álvarez, E.P.; Garde-Cerdán, T.; Santamaría, P.; García-Escudero, E.; Peregrina, F. Influence of two different cover crops on soil N availability, N nutritional status, and grape yeast-assimilable N (YAN) in a cv. Tempranillo vineyard. Plant Soil 2015, 390, 143–156. [Google Scholar] [CrossRef]
- Bell, S.J. The Effect of Nitrogen Fertilisation on the Growth, Yield and Juice Composition of Vitis vinifera cv. Cabernet Sauvignon Grapevines. Ph.D. Thesis, The University of Western Australia, Perth, Australia, 1994. [Google Scholar]
- Bell, A.A.; Ough, C.S.; Kliewer, W.M. Effects on must and wine composition, rates of fermentation, and wine quality of nitrogen fertilisation of Vitis vinifera var. Thompson Seedless grapevines. Am. J. Enol. Vitic. 1979, 30, 124–129. [Google Scholar]
- Strebel, O.W.H.M.; Duynisveld, W.H.M.; Böttcher, J. Nitrate pollution of groundwater in western Europe. Agric. Ecosyst. Environ. 1989, 26, 189–214. [Google Scholar] [CrossRef]
- Zhou, Z. A global assessment of nitrate contamination in groundwater. In International Groundwater Resources Assessment Center; Internship report; Wageningen University: Wageningen, The Netherlands, 2015. [Google Scholar]
- Kaye-Blake, W.; Schilling, C.; Monaghan, R.; Vibart, R.; Dennis, S.; Post, E. Quantification of environmental-economic trade-offs in nutrient management policies. Agric. Syst. 2019, 173, 458–468. [Google Scholar] [CrossRef]
- EC. EU Agricultural Outlook for Markets and Income, 2018–2030; European Commission, DG Agriculture and Rural Development: Brussels, Belgium, 2018. [Google Scholar]
- Epa.gov. United States Environmental Protection Agency. The Sources and Solutions: Agriculture. Available online: https://www.epa.gov/nutrientpollution/sources-and-solutions-agriculture (accessed on 1 February 2020).
- Fernández, V.; Eichert, T. Uptake of hydrophilic solutes through plant leaves: Current state of knowledge and perspectives of foliar fertilization. Crit. Rev. Plant Sci. 2009, 28, 36–68. [Google Scholar] [CrossRef] [Green Version]
- Haytova, D. A review of foliar fertilization of some vegetables crops. Annu. Res. 2013, 3, 455–465. [Google Scholar]
- Viik, E.; Maend, M.; Karise, R.; Laeaeniste, P.; Williams, I.H.; Luik, A. The impact of foliar fertilization on the number of bees (Apoidea) on spring oilseed rape. Zemdirbyste 2012, 99, 41–46. [Google Scholar]
- Dong, S.; Neilsen, D.; Neilsen, G.H.; Fuchigami, L.H. Foliar N application reduces soil NO 3−N leaching loss in apple orchards. Plant Soil 2005, 268, 357–366. [Google Scholar] [CrossRef]
- Canoura, C.; Kelly, M.T.; Ojeda, H. Effect of irrigation and timing and type of nitrogen application on the biochemical composition of Vitis vinifera L. cv. Chardonnay and Syrah grapeberries. Food Chem. 2018, 241, 171–181. [Google Scholar] [CrossRef]
- Lasa, B.; Menendez, S.; Sagastizabal, K.; Cervantes, M.E.C.; Irigoyen, I.; Muro, J.; Aparicio-Tejo, P.M.; Ariz, I. Foliar application of urea to “Sauvignon Blanc” and “Merlot” vines: Doses and time of application. Plant Growth Regul. 2012, 67, 73–81. [Google Scholar] [CrossRef]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Portu, J.; López-Alfaro, I.; Gómez-Alonso, S.; López, R.; Garde-Cerdán, T. Changes on grape phenolic composition induced by grapevine foliar applications of phenylalanine and urea. Food Chem. 2015, 180, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Garde-Cerdán, T.; López, R.; Portu, J.; González-Arenzana, L.; López-Alfaro, I.; Santamaría, P. Study of the effects of proline, phenylalanine, and urea foliar application to Tempranillo vineyards on grape amino acid content. Comparison with commercial nitrogen fertilisers. Food Chem. 2014, 163, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Garde-Cerdán, T.; Portu, J.; López, R.; Santamaría, P. Effect of foliar applications of proline, phenylalanine, urea, and commercial nitrogen fertilizers on stilbene concentrations in Tempranillo musts and wines. Am. J. Enol. Vitic. 2015, 66, 542–547. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Gutiérrez-Gamboa, G.; López, R.; Rubio-Bretón, P.; Pérez-Álvarez, E.P. Influence of foliar application of phenylalanine and urea at two doses to vineyards on grape volatile composition and amino acids content. Vitis 2018, 57, 137–141. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Garde-Cerdán, T.; Gonzalo-Diago, A.; Moreno-Simunovic, Y.; Martínez-Gil, A.M. Effect of different foliar nitrogen applications on the must amino acids and glutathione composition in Cabernet Sauvignon vineyard. LWT Food Sci. Technol. 2017, 75, 147–154. [Google Scholar] [CrossRef]
- Biasi, R.; Brunori, E.; Ferrara, C.; Salvati, L. Assessing Impacts of Climate Change on Phenology and Quality Traits of Vitis vinifera L.: The Contribution of Local Knowledge. Plants 2019, 8, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, V.; Guzmán-Delgado, P.; Graça, J.; Santos, S.; Gil, L. Cuticle structure in relation to chemical composition: Re-assessing the prevailing model. Front. Plant Sci. 2016, 7, 427. [Google Scholar] [CrossRef] [Green Version]
- Bondada, B.R.; Oosterhuis, D.M.; Norman, R.J. Cotton leaf age, epicuticular wax, and nitrogen-15 absorption. Crop Sci. 1997, 37, 807–811. [Google Scholar] [CrossRef]
- Bondada, B.R.; Syvertsen, J.P.; Albrigo, L.G. Urea nitrogen uptake by citrus leaves. HortScience 2001, 36, 1061–1065. [Google Scholar] [CrossRef] [Green Version]
- Baker, E.A.; Hunt, G.M. Developmental changes in leaf epicuticular waxes in relation to foliar penetration. New Phytol. 1981, 88, 731–747. [Google Scholar] [CrossRef]
- Scognamiglio, P. Effetto di Diverse Intensità di Defogliazione e Ombreggiamento Precoci Sulla Compattezza del Grappolo, le Componenti della Produzione, la Composizione dell’acino e le Caratteristiche Sensoriali del Vino nel Vitigno ‘Greco’. Ph.D. Thesis, University of Naples Federico II, Portici, Italy, 2017. [Google Scholar] [CrossRef]
- Bondada, R.B.; Petracek, D.P.; Syvertsen, P.J.; Albrigo, L.G. Cuticular penetration characteristics of urea in citrus leaves. J. Hortic. Sci. Biotechnol. 2006, 81, 219–224. [Google Scholar] [CrossRef]
- Toselli, M.; Thalheimer, M.; Tagliavini, M. Leaf uptake and subsequent partitioning of urea-N as affected by the concentration and volume of spray solution and by the shoot leaf position in apple (Malus domestica) trees. J. Hortic. Sci. Biotechnol. 2004, 79, 97–100. [Google Scholar] [CrossRef]
- Jiménez-Moreno, N.; Moler, J.A.; Palacios, M.B.; Esparza, I.; Nieto-Rojo, R.; Ancín-Azpilicueta, C. Foliar application of urea to Tempranillo vines increased the amino acid concentration of the must. Food Addit. Contam. A 2019, 37, 216–227. [Google Scholar] [CrossRef]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein hydrolysate stimulates growth in tomato coupled with N-dependent gene expression involved in N assimilation. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant application with a tropical plant extract enhances Corchorus olitorius adaptation to sub-optimal nutrient regimens by improving physiological parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activity of two protein hydrolyzates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 64, 145–158. [Google Scholar] [CrossRef]
- Ertani, A.; Nardi, S.; Altissimo, A. Review: Long-term research activity on the biostimulant properties of natural origin compounds. Acta Hortic. 2012, 1009, 181–188. [Google Scholar] [CrossRef]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an alfalfa protein hydrolysate on the gene expression and activity of enzymes of the tricarboxylic acid (TCA) cycle and nitrogen metabolism in Zea mays L. J. Agricult. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef]
- Lawlor, D.W. Carbon and nitrogen assimilation in relation to yield: Mechanisms are the key to understanding production systems. J. Exp. Bot. 2002, 53, 773–787. [Google Scholar] [CrossRef] [PubMed]
- Keller, M. Partitioning of Assimilates. In The Science of Grapevines: Anatomy and Physiology, 2nd ed.; Keller, M., Ed.; Elsevier: Alpharetta, GA, USA, 2015; pp. 145–192. [Google Scholar] [CrossRef]
- Kliewer, W.M.; Schultz, H.B. Influence of environment on metabolism of organic acids and carbohydrates in Vitis vinifera. I. Temperature. Am. J. Enol. Vitic. 1964, 15, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Gerés, H. Biochemical changes throughout grape berry development and fruit and wine quality. Food 2007, 1, 1–22. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 10th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2010.
- Greco. Registro Nazionale delle Varietà di Vite. Ministero delle Politiche Agricole Alimentari e Forestali. Available online: http://catalogoviti.politicheagricole.it/result.php?codice=097 (accessed on 4 May 2020).
- Greco di Tufo Guidelines 2014 Agricultural Department Website. (Campania Region). Available online: http://www.agricoltura.regione.campania.it/viticoltura/disciplinari/DOCG_Greco.pdf (accessed on 20 February 2020).
- González-Santamaría, R.; Ruiz-González, R.; Nonell, S.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. Influence of foliar riboflavin applications to vineyard on grape amino acid content. Food Chem. 2018, 240, 601–606. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YAN (mg N/L) | NH4+-N (mg N/L) | Aminic N (mg N/L) | TSS (°Brix) | pH | TA (g/L Tartaric Acid) | Malic Acid (g/L) | |
|---|---|---|---|---|---|---|---|
| First year | |||||||
| Fertilizer (F) | |||||||
| C | 38.5b | 18.0 | 20.5b | 23.3 | 2.70 | 10.1 | 2.38 |
| AX | 55.2a | 19.5 | 35.7a | 23.1 | 2.75 | 9.4 | 2.29 |
| AN | 46.7ab | 17.0 | 29.7a | 22.8 | 2.74 | 9.5 | 2.30 |
| Phenological stage (PS) | |||||||
| Veraison onset | 39.1b | 15.3 | 23.7b | 23.2 | 2.73 | 9.7 | 2.42 |
| Full veraison | 54.6a | 21.0 | 33.6a | 23.0 | 2.73 | 9.7 | 2.22 |
| Probability Z | |||||||
| F | * | ns | ** | ns | ns | ns | ns |
| PS | ** | ns | ** | ns | ns | ns | ns |
| F × PS | * | ns | * | ns | ns | ns | ns |
| Second Year | |||||||
| Fertilizer (F) | |||||||
| C | 179.5b | 112.2b | 67.2 | 22.9 | 3.07 | 7.3 | 0.54 |
| AX | 302.7a | 230.4a | 72.4 | 22.6 | 3.12 | 6.8 | 0.56 |
| AN | 297.6a | 224.0a | 73.6 | 22.5 | 3.08 | 7.3 | 0.58 |
| Phenological stage (PS) | |||||||
| Full veraison | 307.8a | 307.8a | 234.7 | 73.1 | 3.09 | 7.1 | 0.54 |
| Post-veraison | 212.1b | 212.1b | 143.0 | 69.1 | 3.09 | 7.1 | 0.59 |
| Probability | |||||||
| F | * | * | ns | ns | ns | ns | ns |
| PS | * | * | ns | ns | ns | ns | ns |
| F × PS | ** | * | ns | ns | ns | ns | ns |
| Year | Treatment | Veraison Onset | Full Veraison | Post-Veraison | |||
|---|---|---|---|---|---|---|---|
| NH4+-N (mg N/L) | Aminic N (mg N/L) | NH4+-N (mg N/L) | Aminic N (mg N/L) | NH4+-N (mg N/L) | Aminic N (mg N/L) | ||
| 2016 | C | 19.5 ± 4.4 | 20.0 ± 1.7 | 16.5 ± 2.2 | 21.0 ± 1.8b | - | - |
| AX | 13.7 ± 2.4 | 26.0 ± 0.9 | 25.2 ± 3.9 | 45.5 ± 6.5a | - | - | |
| AN | 12.7 ± 2.7 | 25.2 ± 2.5 | 21.2 ± 5.9 | 34.2 ± 2.9ab | - | - | |
| 2017 | C | - | - | 104.2 ± 7.8b | 60.2 ± 7.0 | 120.2 ± 11.3 | 74.2 ± 16.6 |
| AX | - | - | 351.0 ± 71.7a | 81.2 ± 5.9 | 109.7 ± 12.5 | 63.5 ± 9.5 | |
| AN | - | - | 249.0 ± 41.9ab | 77.7 ± 7.4 | 199.0 ± 42.5 | 69.5 ± 8.9 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mataffo, A.; Scognamiglio, P.; Dente, A.; Strollo, D.; Colla, G.; Rouphael, Y.; Basile, B. Foliar Application of an Amino Acid-Enriched Urea Fertilizer on ‘Greco’ Grapevines at Full Veraison Increases Berry Yeast-Assimilable Nitrogen Content. Plants 2020, 9, 619. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9050619
Mataffo A, Scognamiglio P, Dente A, Strollo D, Colla G, Rouphael Y, Basile B. Foliar Application of an Amino Acid-Enriched Urea Fertilizer on ‘Greco’ Grapevines at Full Veraison Increases Berry Yeast-Assimilable Nitrogen Content. Plants. 2020; 9(5):619. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9050619
Chicago/Turabian StyleMataffo, Alessandro, Pasquale Scognamiglio, Antonio Dente, Daniela Strollo, Giuseppe Colla, Youssef Rouphael, and Boris Basile. 2020. "Foliar Application of an Amino Acid-Enriched Urea Fertilizer on ‘Greco’ Grapevines at Full Veraison Increases Berry Yeast-Assimilable Nitrogen Content" Plants 9, no. 5: 619. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9050619