Genetic Diversity, Population Structure and Marker-Trait Association for 100-Seed Weight in International Safflower Panel Using SilicoDArT Marker Information


 , , ,
, , ,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
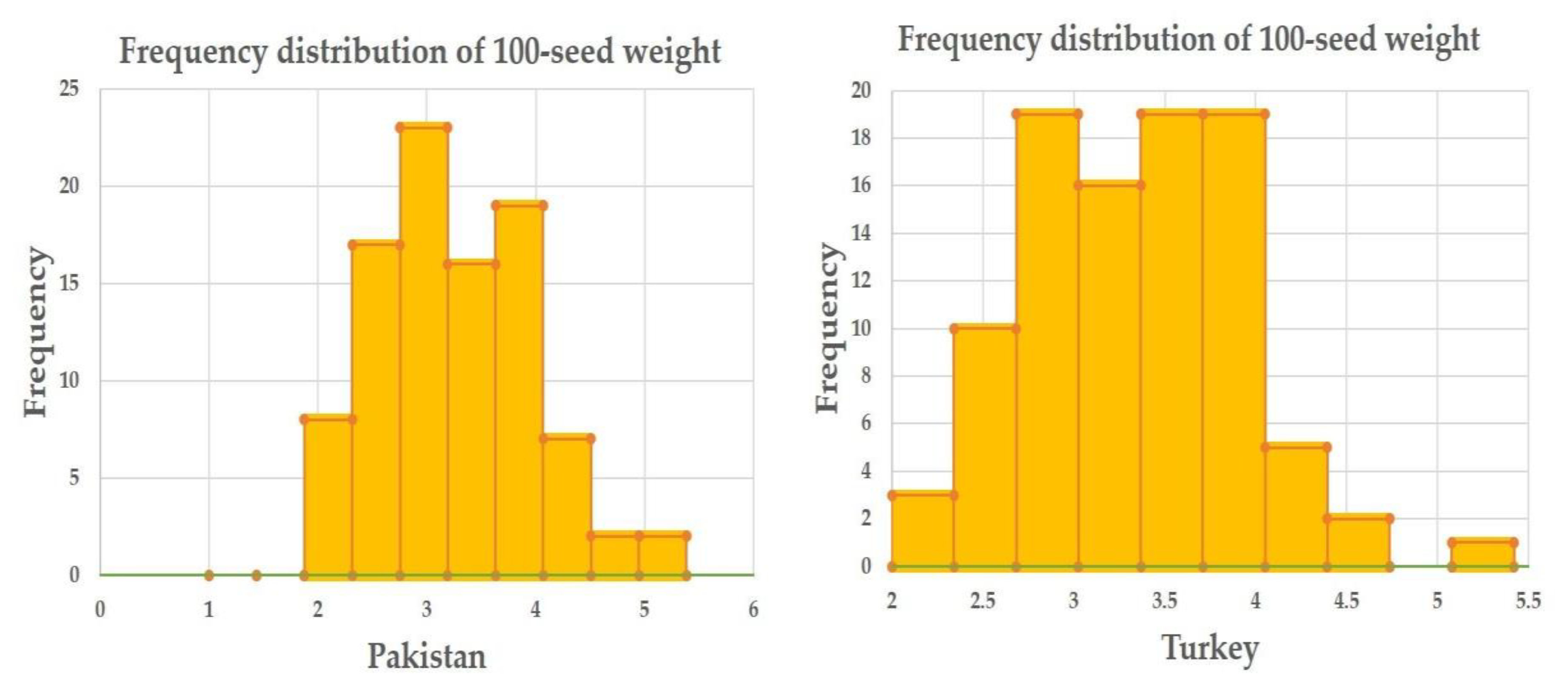
2.1. Phenotypic Data Evaluation
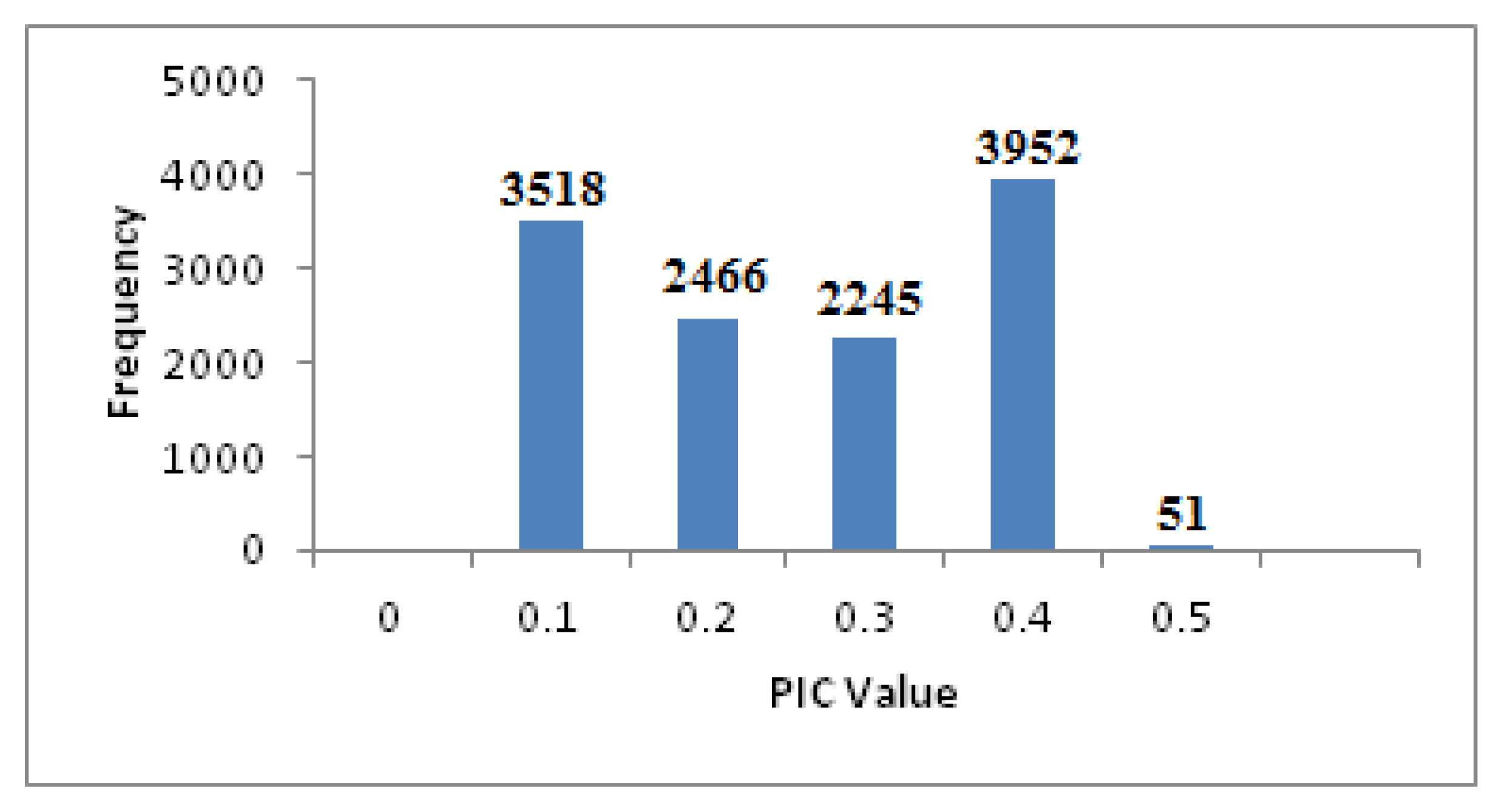
2.2. DArTseq Profiling by GBS
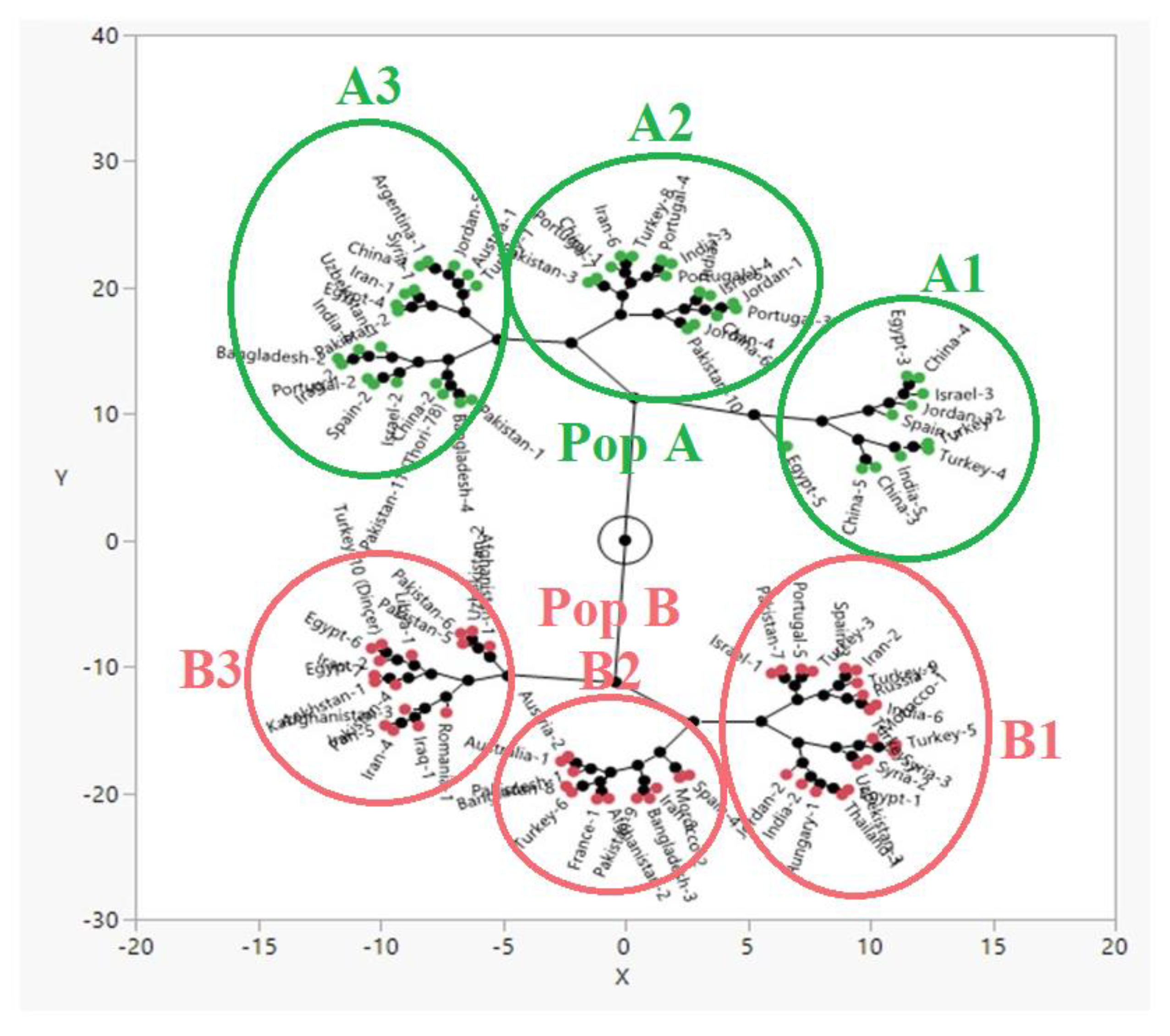
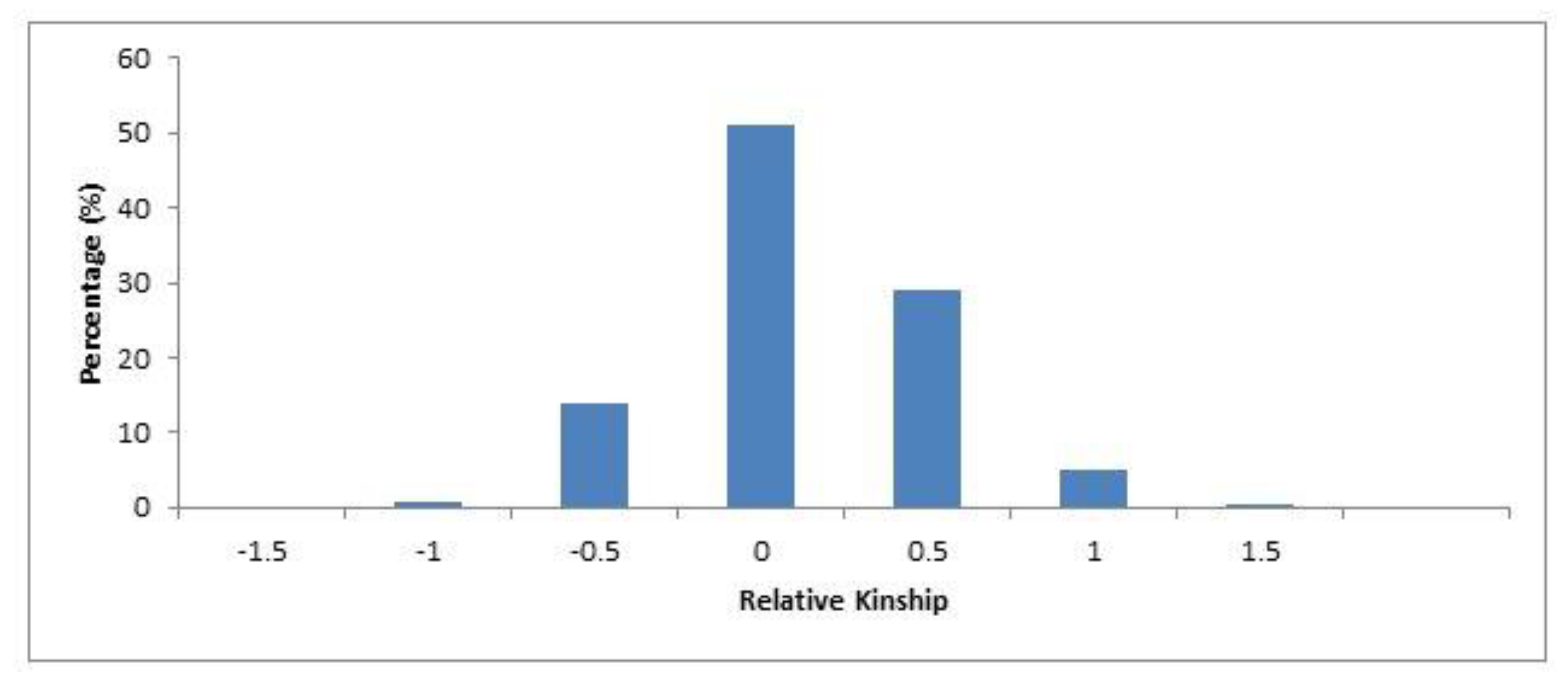
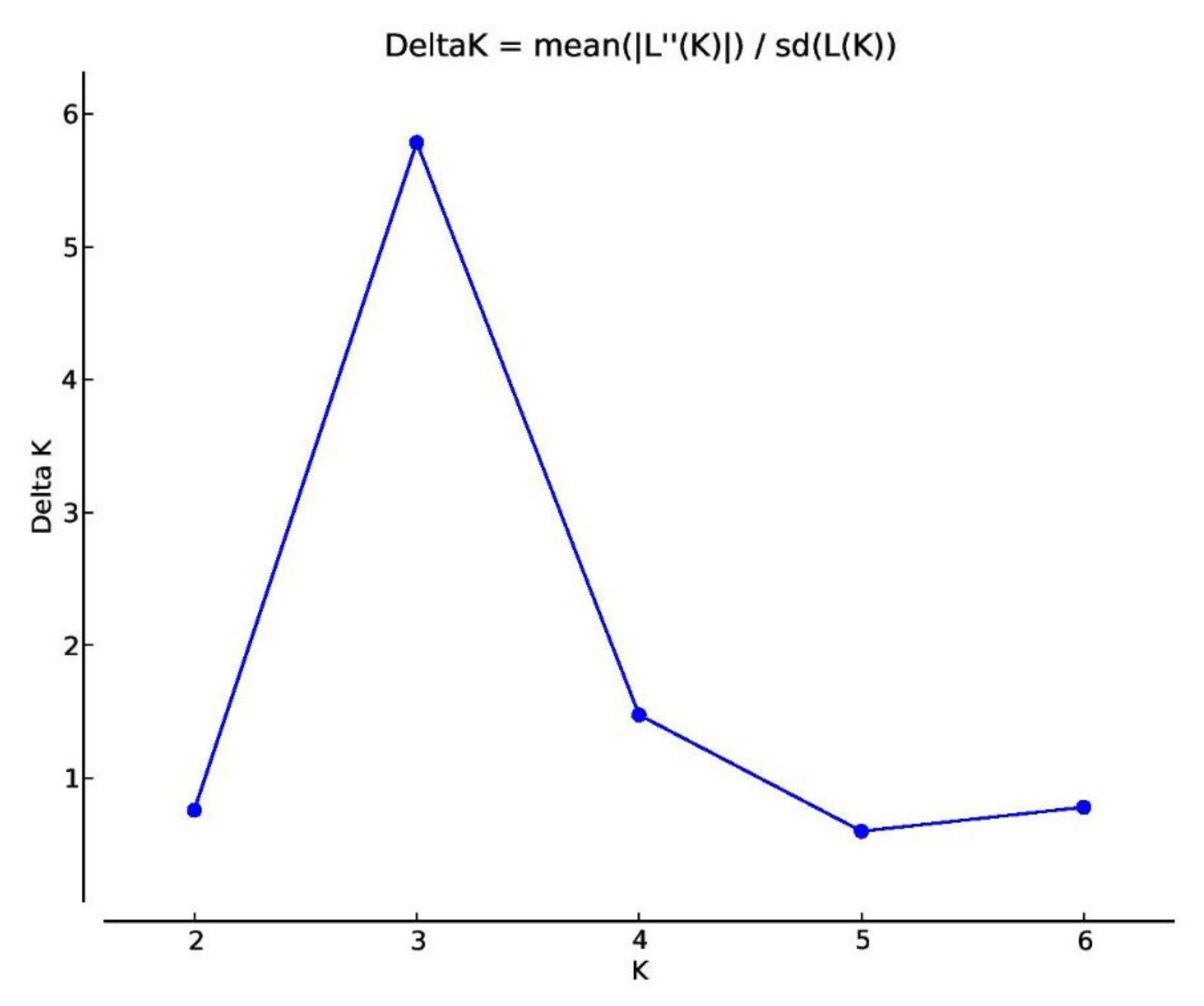
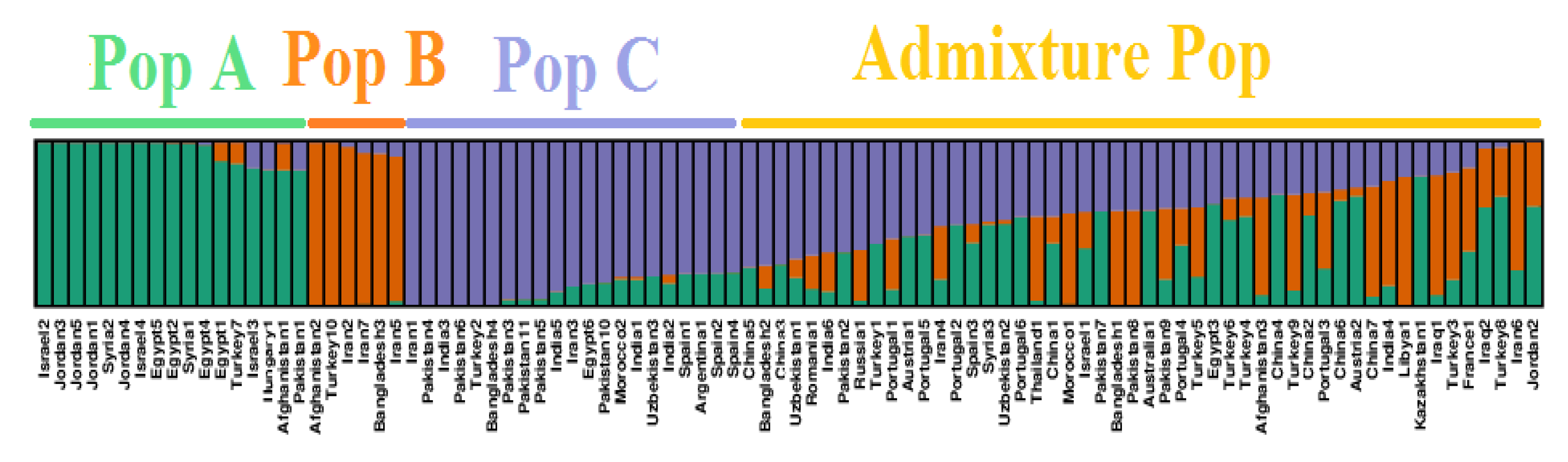
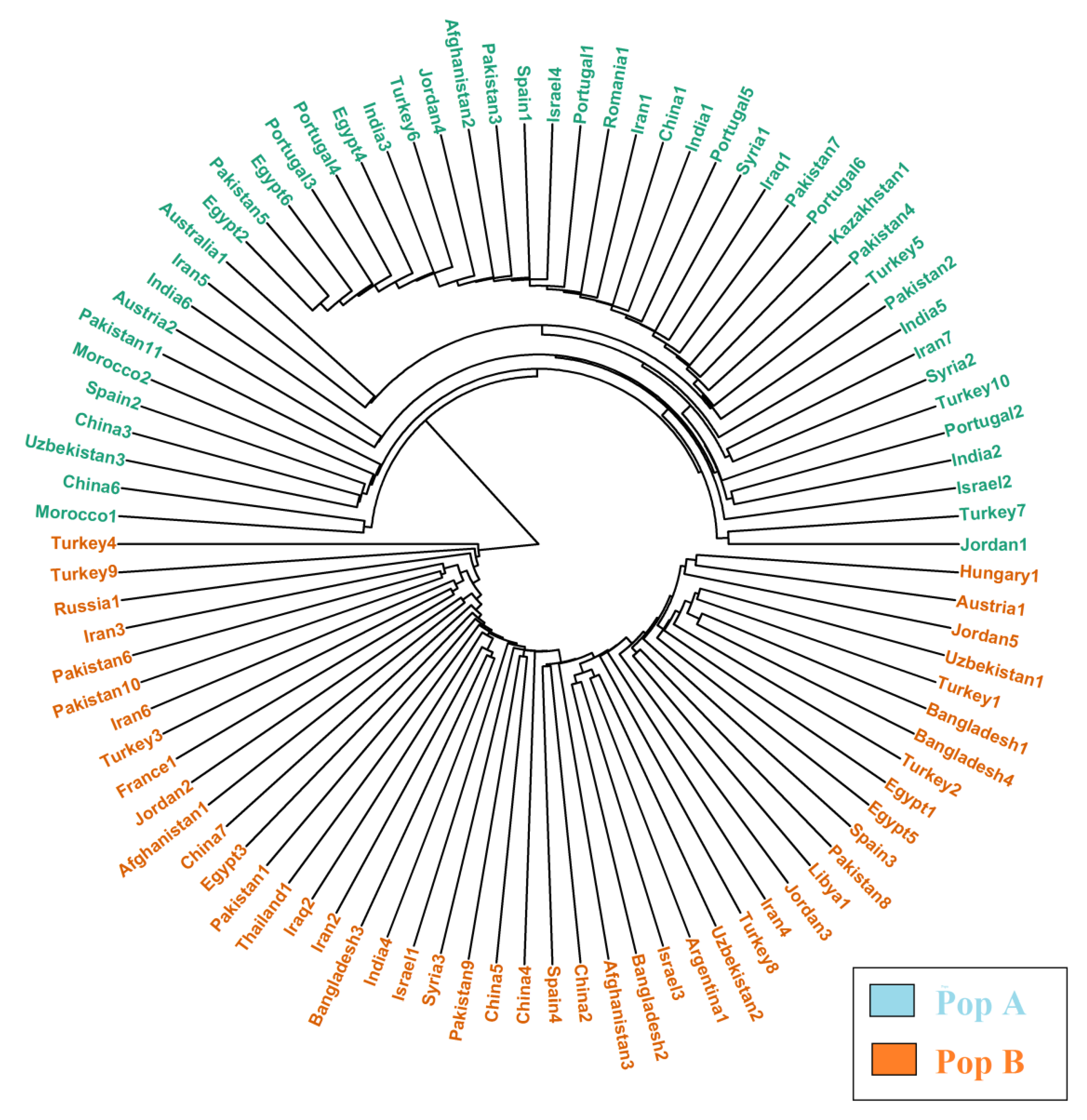
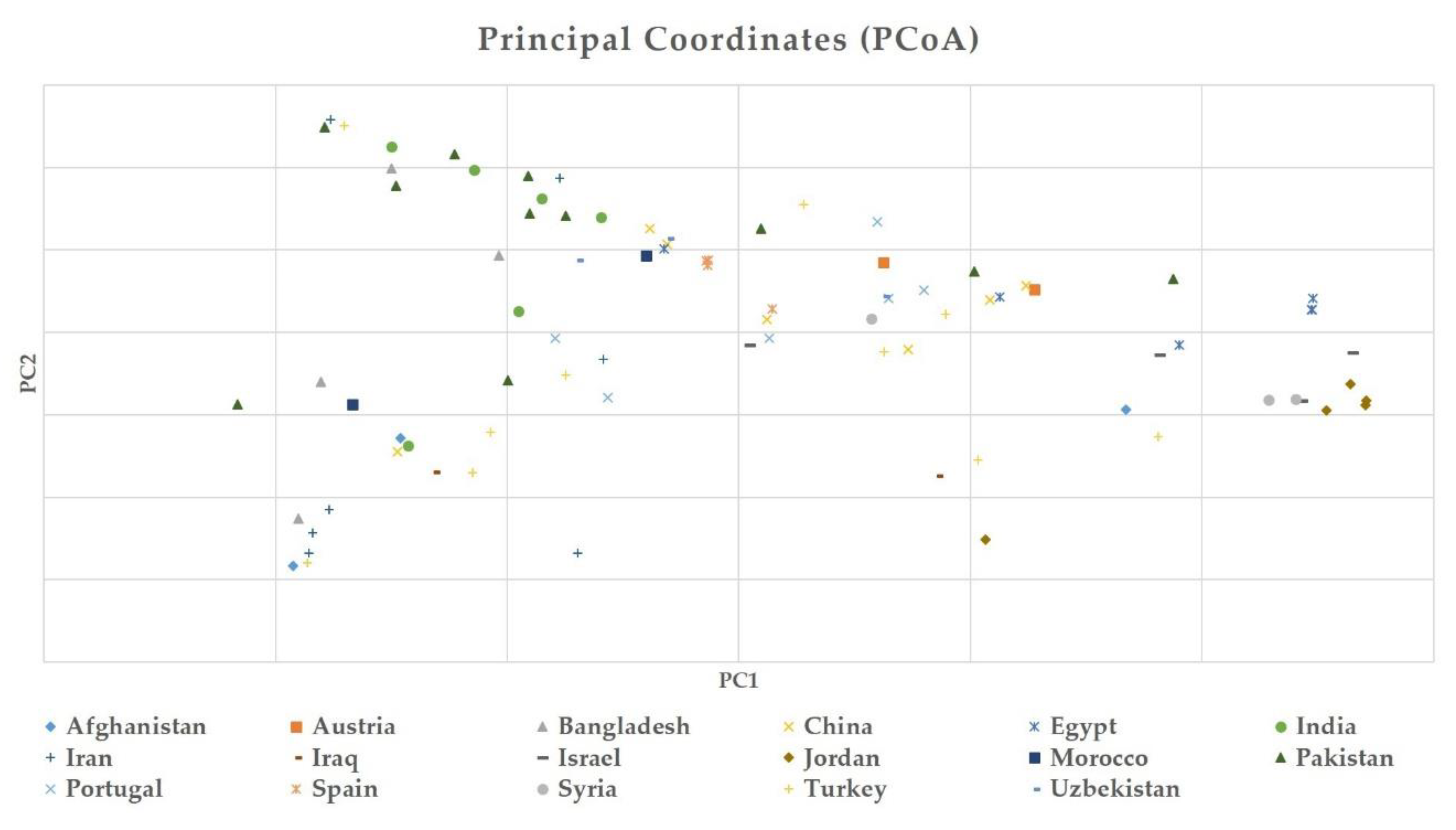
2.3. Genetic Diversity and Population Structure Analysis in Safflower Panel
2.4. Marker-Trait Associations and Putative Candiadate Genes Identification for 100-Seed Weight
3. Discussion
3.1. Genetic Structure and Diversity in Safflower Panel
3.2. Marker-Trait Associations for 100-Seed Weight
4. Materials and Methods
4.1. Plant Materials and Phenotypic Evaluation
4.2. Genomic DNA Isolation
4.3. DArTseq-Generated SilicoDArT Marker Analysis
4.4. Statistical Analysis
4.4.1. Phenotypic Data Analysis
4.4.2. DArTseq Markers Analysis
4.4.3. Genetic Diversity Analyses
4.4.4. Functional Analysis for Putative Candidate Gene Identification
4.4.5. Genome-Wide Association Mapping for 100-Seed Weight
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kumari, S.; Choudhary, R.C.; Kumara, S.R.; Saharan, V.; Joshi, A.; Munot, J. Assessment of genetic diversity in safflower (Carthamus tinctorius L.) genotypes through morphological and SSR marker. J. Pharmacogn. Phytochem. 2017, 6, 2723–2731. [Google Scholar]
- Ali, F.; Yılmaz, A.; Nadeem, M.A.; Habyarimana, E.; Subaşı, I.; Nawaz, M.A.; Chaudhary, H.J.; Shahid, M.Q.; Ercişli, S.; Zia, M.A.B.; et al. Mobile genomic element diversity in world collection of safflower (Carthamus tinctorius L.) panel using iPBS-retrotransposon markers. PLoS ONE 2019, 14, e0211985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, F.; Yilmaz, A.; Chaudhary, H.J.; Nadeem, M.A.; Rabbani, M.A.; Arslan, Y.; Nawaz, M.A.; Habyarimana, E.; Baloch, F.S. Investigation of Morpho-Agronomic Performance and Selection Indices in the International Safflower Panel for Breeding Perspectives. Turk. J. Agric. For. 2020, 44, 103–120. [Google Scholar] [CrossRef]
- Ali, F.; Nadeem, M.A.; Habyarimana, E.; Yilmaz, A.; Nawaz, M.A.; Khalil, I.H.; Ercisli, S.; Chung, G.; Chaudhary, H.J.; Baloch, F.S. Molecular characterization of genetic diversity and similarity centers of safflower accessions with ISSR markers. Braz. J. Bot. 2020, 43, 109–121. [Google Scholar] [CrossRef]
- Dajue, L.; Mündel, H.H. Safflower. Carthamus Tinctorius L. Promoting the Conservation and Use of Underutilized and Neglected Crops, 7th ed.; Institute of Plant Genetics and Crop Plant Research: Rome, Italy, 1996; pp. 1–83. [Google Scholar]
- Ferwerda, J.; Oilseed Crops. Weiss, E.A., Ed.; Elsevier: Harlow, Essex, UK, 1983; ISBN 0-582-46338-6.
- Dordas, C.A.; Sioulas, C. Dry matter and nitrogen accumulation, partitioning, and retranslocation in safflower (Carthamus tinctorius L.) as affected by nitrogen fertilization. Field Crops Res. 2009, 110, 35–43. [Google Scholar] [CrossRef]
- Marinova, E.; Riehl, S. Carthamus species in the ancient Near East and south-eastern Europe: Archaeobotanical evidence for their distribution and use as a source of oil. Veg. Hist. Archaeobot. 2009, 18, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Zeinali, E. Safflower (Characteristics, Production and Utilization); Gorgan University of Agricultural Science and Natural Resources: Gorgan, Iran, 1999; p. 137. [Google Scholar]
- Yang, Y.X.; Wu, W.; Zheng, Y.L.; Chen, L.; Liu, R.J.; Huang, C.Y. Genetic diversity and relationships among safflower (Carthamus tinctorius L.) analyzed by inter-simple sequence repeats (ISSRs). Genet. Resour. Crop. Evol. 2007, 54, 1043–1051. [Google Scholar] [CrossRef]
- Mayerhofer, M.; Mayerhofer, R.; Topinka, D.; Christianson, J.; Good, A.G. Introgression potential between safflower (Carthamus tinctorius) and wild relatives of the genus Carthamus. BMC Plant Biol. 2011, 11, 47. [Google Scholar] [CrossRef] [Green Version]
- Baloch, F.S.; Alsaleh, A.; Shahid, M.Q.; Çiftçi, V.; de Miera, L.E.S.; Aasim, M.; Nadeem, M.A.; Aktaş, H.; Özkan, H.; Hatipoğlu, R. A whole genome DArTseq and SNP analysis for genetic diversity assessment in durum wheat from central fertile crescent. PLoS ONE 2017, 12, e0167821. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, M.A.; Habyarimana, E.; Çiftçi, V.; Nawaz, M.A.; Karaköy, T.; Comertpay, G.; Shahid, M.Q.; Hatipoğlu, R.; Yeken, M.Z.; Ali, F.; et al. Characterization of genetic diversity in Turkish common bean gene pool using phenotypic and whole-genome DArTseq-generated silicoDArT marker information. PLoS ONE 2018, 13, e0205363. [Google Scholar] [CrossRef]
- Mary, S.; Gopalan, A. Dissection of genetic attributes yield traits of fodder cowpea in F3 and F4. J. Appl. Sci. Res. 2006, 2, 805–808. [Google Scholar]
- Arystanbekkyzy, M.; Nadeem, M.A.; Aktas, H.; Yeken, M.Z.; Zencirci, N.; Nawaz, M.A.; Ali, F.; Haider, M.S.; Tunc, K.; Chung, G.; et al. Phylogenetic and taxonomic relationship of turkish wild and cultivated emmer (triticum turgidum ssp. dicoccoides) revealed by iPBSretrotransposons markers. Int. J. Agric. Biol. 2018, 10. [Google Scholar] [CrossRef]
- Yaldiz, G.; Camlica, M.; Nadeem, M.A.; Nawaz, M.A.; Baloch, F.S. Genetic diversity assessment in Nicotiana tabacum L. with iPBS-retrotransposons. Turk. J. Agric. For. 2018, 42, 154–164. [Google Scholar] [CrossRef]
- Castañeda-Álvarez, N.P.; Khoury, C.K.; Achicanoy, H.A.; Bernau, V.; Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Harker, R.H.; Jarvis, A.; Maxted, N. Global conservation priorities for crop wild relatives. Nat. Plants 2016, 2, 16022. [Google Scholar] [CrossRef]
- Johnson, R.C.; Kisha, T.; Evans, M. Characterizing safflower germplasm with AFLP molecular markers. Crop Sci. 2007, 47, 1728–1736. [Google Scholar] [CrossRef] [Green Version]
- Amiri, R.; Azdi, S.; Ghanadha, M.; Abd, M. Detection of DNA polymorphism in landrace populations of safflower in Iran using RAPD-PCR technique. Iran. J. Agric. Sci. 2001, 32, 737–745. [Google Scholar]
- Sehgal, D.; Raina, S.N.; Devarumath, R.M.; Sasanuma, T.; Sasakuma, T. Nuclear DNA assay in solving issues related to ancestry of the domesticated diploid safflower (Carthamus tinctorius L.) and the polyploid (Carthamus) taxa, and phylogenetic and genomic relationships in the genus Carthamus L. (Asteraceae). Mol. Phylogenet. Evol. 2009, 53, 631–644. [Google Scholar] [CrossRef]
- Ambreen, H.; Kumar, S.; Kumar, A.; Agarwal, M.; Jagannath, A.; Goel, S. Association mapping for important agronomic traits in safflower (Carthamus tinctorius L.) core collection using microsatellite markers. Front. Plant Sci. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Ambreen, H.; Kumar, S.; Variath, M.T.; Joshi, G.; Bali, S.; Agarwal, M.; Kumar, A.; Jagannath, A.; Goel, S. Development of genomic microsatellite markers in Carthamus tinctorius L.(safflower) using next generation sequencing and assessment of their cross-species transferability and utility for diversity analysis. PLoS ONE 2015, 10, e0135443. [Google Scholar] [CrossRef] [Green Version]
- Amini, F.; Saeidi, G.; Arzani, A. Study of genetic diversity in safflower genotypes using agro-morphological traits and RAPD markers. Euphytica 2008, 163, 21–30. [Google Scholar] [CrossRef]
- Khan, M.A.; von Witzke-Ehbrecht, S.; Maass, B.L.; Becker, H.C. Relationships among different geographical groups, agro-morphology, fatty acid composition and RAPD marker diversity in safflower (Carthamus tinctorius). Genet. Resour. Crop Evol. 2009, 56, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Ambreen, H.; Murali, T.; Bali, S.; Agarwal, M.; Kumar, A.; Goel, S.; Jagannath, A. Assessment of genetic diversity and population structure in a global reference collection of 531 accessions of Carthamus tinctorius L.(Safflower) using AFLP markers. Plant Mol. Biol. Rep. 2015, 33, 1299–1313. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.A.; Sung, J.S.; Lee, S.Y.; Chung, J.W.; Yi, J.Y.; Kim, Y.G.; Lee, M.C. Genetic assessment of safflower (Carthamus tinctorius L.) collection with microsatellite markers acquired via pyrosequencing method. Mol. Ecol. Resour. 2014, 14, 69–78. [Google Scholar] [CrossRef]
- Pearl, S.A.; Burke, J.M. Genetic diversity in Carthamus tinctorius (Asteraceae; safflower), an underutilized oilseed crop. Am. J. Bot. 2014, 101, 1640–1650. [Google Scholar] [CrossRef] [Green Version]
- Raman, H.; Raman, R.; Nelson, M.N.; Aslam, M.; Rajasekaran, R.; Wratten, N.; Cowling, W.A.; Kilian, A.; Sharpe, A.G.; Schondelmaier, J. Diversity array technology markers: Genetic diversity analyses and linkage map construction in rapeseed (Brassica napus L.). DNA Res. 2011, 19, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Hassani, S.M.R.; Talebi, R.; Pourdad, S.S.; Naji, A.M.; Fayaz, F. In-depth genome diversity, population structure and linkage disequilibrium analysis of worldwide diverse safflower (Carthamus tinctorius L.) accessions using NGS data generated by DArTseq technology. Mol. Biol. Rep. 2020, 47, 2123–2135. [Google Scholar] [CrossRef] [PubMed]
- Flint-Garcia, S.A.; Thuillet, A.C.; Yu, J.; Pressoir, G.; Romero, S.M.; Mitchell, S.E.; Doebley, J.; Kresovich, S.; Goodman, M.M.; Buckler, E.S. Maize association population: A high-resolution platform for quantitative trait locus dissection. Plant J. 2005, 44, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Buckler, E.S. Genetic association mapping and genome organization of maize. Curr. Opin. Biotech. 2006, 17, 155–160. [Google Scholar] [CrossRef]
- Abdurakhmonov, I.Y.; Abdukarimov, A. Application of association mapping to understanding the genetic diversity of plant germplasm resources. Int. J. Plant Genom. 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimi, F.; Majidi, M.M.; Arzani, A.; Mohammadi-Nejad, G. Association analysis of molecular markers with traits under drought stress in safflower. Crop Pasture Sci. 2017, 68, 167–175. [Google Scholar] [CrossRef]
- Mahmood, T.; Rahman, M.H.; Stringam, G.R.; Yeh, F.; Good, A. Molecular markers for yield components in Brassica juncea—Do these assist in breeding for high seed yield? Euphytica 2005, 144, 157–167. [Google Scholar] [CrossRef]
- El-Lattief, E.A. Evaluation of 25 safflower genotypes for seed and oil yields under arid environment in upper Egypt. Asian J. Crop Sci. 2012, 4, 72–79. [Google Scholar] [CrossRef]
- Rahim, A.D.A. Dimension, geometric, agricultural and quality characteristics of safflower seeds. Turk. J. Field Crops 2014, 19, 7–12. [Google Scholar]
- Köse, A.; Koşar, F.; Bilir, Ö. Agricultural performances of some safflower (Carthamus tinctorius L.) lines developed by single plant selection method. J. Agric. Sci. 2018, 24, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Tantasawat, P.A.; Wang, W.; Gao, X.; Zhang, L. Population structure of Chinese southwest wheat germplasms resistant to stripe rust and powdery mildew using the DArT-seq technique. Ciênc. Rural. 2018, 48, 1–9. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef] [Green Version]
- Panahi, B.; Neghab, M.G. Genetic characterization of Iranian safflower (Carthamus tinctorius) using inter simple sequence repeats (ISSR) markers. Physiol. Mol. Biol. Plants 2013, 19, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Sung, J.S.; Cho, G.T.; Lee, G.A.; Baek, H.J.; Huh, M.K. Phylogenetic Relationships and Genetic Diversity in Collected Resources of Carthamus tinctorius by Random Amplified Polymorphic DNA Markers. J. Life Sci. 2010, 20, 1764–1771. [Google Scholar] [CrossRef]
- Hirano, R.; Kikuchi, A.; Kawase, M.; Watanabe, K.N. Evaluation of genetic diversity of bread wheat landraces from Pakistan by AFLP and implications for a future collection strategy. Genet. Resour. Crop Evol. 2008, 55, 1007–1015. [Google Scholar] [CrossRef]
- Bouchet, S.; Pot, D.; Deu, M.; Rami, J.F.; Billot, C.; Perrier, X.; Rivallan, R.; Gardes, L.; Xia, L.; Wenzl, P. Genetic structure, linkage disequilibrium and signature of selection in sorghum: Lessons from physically anchored DArT markers. PLoS ONE 2012, 7, e33470. [Google Scholar] [CrossRef] [Green Version]
- Newell, M.A.; Cook, D.; Hofmann, H.; Jannink, J.L. An algorithm for deciding the number of clusters and validation using simulated data with application to exploring crop population structure. Ann. Appl. Stat. 2013, 7, 1898–1916. [Google Scholar] [CrossRef] [Green Version]
- Chapman, M.A.; Hvala, J.; Strever, J.; Burke, J.M. Population genetic analysis of safflower (Carthamus tinctorius; Asteraceae) reveals a Near Eastern origin and five centers of diversity. Am. J. Bot. 2010, 97, 831–840. [Google Scholar] [CrossRef]
- Pearl, S.A.; Bowers, J.E.; Reyes-Chin-Wo, S.; Michelmore, R.W.; Burke, J.M. Genetic analysis of safflower domestication. BMC Plant Biol. 2014, 14, 43. [Google Scholar] [CrossRef] [Green Version]
- Yol, E.; Uzun, B. Geographical patterns of sesame accessions grown under Mediterranean environmental conditions, and establishment of a core collection. Crop Sci. 2012, 52, 2206–2214. [Google Scholar] [CrossRef]
- Chaudhary, S.K. Path analysis for seed yield in safflower (Carthamus tinctorius L.) in acid soil under mid altitude conditions. Int. J. Trop. Agric. 1990, 8, 129–132. [Google Scholar]
- Pascual-Villalobos, M.J.; Alburquerque, N. Genetic variation of a safflower germplasm collection grown as a winter crop in southern Spain. Euphytica 1996, 92, 327–332. [Google Scholar] [CrossRef]
- Omidi, T.A.H. Correlation between traits and path analysis for grain and oil yield in spring safflower. Sesame Safflower Newsl. 2000, 15, 78–82. [Google Scholar]
- Mirzahashemi, M.; Mohammadi-Nejad, G.; Golkar, P. A qtl linkage map of safflower for yield under drought stress at reproductive stage. Iran. J. Genet. Plant Breed. 2015, 4, 20–27. [Google Scholar]
- Zumajo-Cardona, C.; Ambrose, B.A. Phylogenetic analyses of key developmental genes provide insight into the complex evolution of seeds. Mol. Phylogenet. Evol. 2020, 147, 106778. [Google Scholar] [CrossRef]
- Téllez-Robledo, B.; Manzano, C.; Saez, A.; Navarro-Neila, S.; Silva-Navas, J.; de Lorenzo, L.; González-García, M.P.; Toribio, R.; Hunt, A.G.; Baigorri, R.; et al. The polyadenylation factor FIP1 is important for plant development and root responses to abiotic stresses. Plant J. 2019, 99, 1203–1219. [Google Scholar] [CrossRef]
- Paez-Garcia, A.; Motes, C.M.; Scheible, W.R.; Chen, R.; Blancaflor, E.B.; Monteros, M.J. Root traits and phenotyping strategies for plant improvement. Plants 2015, 4, 334–355. [Google Scholar] [CrossRef]
- Yildiz, M.; Kocak, M.; Nadeem, M.A.; Cavagnaro, P.; Barboza, K.; Baloch, F.S.; Argün, D.; Keleş, D. Genetic diversity analysis in the Turkish pepper germplasm using iPBS retrotransposonbased markers. Turk. J. Agric. For. 2020, 44, 1–14. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Karaköy, T.; Yeken, M.Z.; Habyarimana, E.; Hatipoğlu, R.; Çiftçi, V.; Nawaz, M.A.; Sönmez, F.; Shahid, M.Q.; Yang, S.H.; et al. Phenotypic characterization of 183 Turkish common bean accessions for agronomic, trading, and consumer-preferred plant characteristics for breeding purposes. Agronomy 2020, 10, 272. [Google Scholar] [CrossRef] [Green Version]
- Barut, M.; Nadeem, M.A.; Karaköy, T.; Baloch, F.S. DNA fingerprinting and genetic diversity analysis of world quinoa germplasm using iPBS-retrotransposon marker system. Turk. J. Agric. For. 2020, 44. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 39–40. [Google Scholar]
- Available online: https://www.diversityarrays.com/orderinstructions/plant-dna-extraction protocol for-dart/ (accessed on 20 December 2019).
- Available online: http://www.diversityarrays.com/ (accessed on 20 December 2019).
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Raman, H.; Raman, R.; Kilian, A.; Detering, F.; Carling, J.; Coombes, N.; Diffey, S.; Kadkol, G.; Edwards, D.; McCully, M. Genome-wide delineation of natural variation for pod shatter resistance in Brassica napus. PLoS ONE 2014, 9, e101673. [Google Scholar] [CrossRef] [Green Version]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C. Diversity arrays technology: A generic genome profiling technology on open platforms. In Data Production and Analysis in Population Genomics; François, P., Aurélie, B., Eds.; Springer: Totowa, NJ, USA, 2012; pp. 67–89. [Google Scholar]
- Li, H.; Vikram, P.; Singh, R.P.; Kilian, A.; Carling, J.; Song, J.; Burgueno-Ferreira, J.A.; Bhavani, S.; Huerta-Espino, J.; Payne, T. A high density GBS map of bread wheat and its application for dissecting complex disease resistance traits. BMC Genomics 2015, 16, 216. [Google Scholar] [CrossRef] [Green Version]
- Rathore, A.; Parsad, R.; Gupta, V. Computer aided construction and analysis of augmented designs. J. Indian Soc. Agric. Stat 2004, 57, 320–344. [Google Scholar]
- Statistical Analysis Software/Stat Software, Version 9.3, 2nd ed.; SAS Institute: Cary, NC, USA, 2011.
- Available online: www.xlstat.com (accessed on 3 April 2020).
- Cömertpay, G.; Baloch, F.S.; Kilian, B.; Ülger, A.C.; Özkan, H. Diversity assessment of Turkish maize landraces based on fluorescent labelled SSR markers. Plant Mol. Biol. Rep. 2012, 30, 261–274. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/ (accessed on 4 May 2020).
- Available online: http://phytozome.jgi.doe.gov/pz/portal.html (accessed on 4 May 2020).
- Available online: http://www.arabidopsis.org (accessed on 4 May 2020).
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. Tassel: Software for association mapping of complex traits in diverse samples. Bioinform 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Gündogdu, M.; Ercisli, S.; Karaköy, T.; Saracoglu, O.; Habyarimana, E.; Lin, X.; Hatipoglu, R.; Nawaz, M.A.; Sameeullah, M.; et al. Uncovering phenotypic diversity and DArTseq marker loci associated with antioxidant activity in common bean. Genes 2020, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Zia, M.A.B.; Demirel, U.; Nadeem, M.A.; Caliskan, M.E. Genome-wide association study identifies various loci underlying agronomic and morphological traits in diversified potato panel. Physiol. Mol. Biol. Plants. 2020, 26, 1003–1020. [Google Scholar] [CrossRef]
- Jin, F.X.; Ji, S.D.; Xie, X.B.; Kang, J.W.; Ju, H.G.; Ahn, S.N. Detection of epistatic interaction of two qtls, gw8. 1 and gw9. 1, underlying grain weight using nearly isogenic lines in rice. Breed. Sci. 2011, 61, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.D. qqman: An R package for visualizing GWAS results using QQ and Manhattan plots. Biorxiv 2014, 005165, 1–2. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pakistan | |||||
| Source | DF | SS | Mean Square | F Value | Pr > F |
| Block | 5 | 0.66 | 0.13 | 3.25 | 0.1108 |
| Check | 1 | 4.58 | 4.58 | 112.09 | 0.0001 ** |
| Accessions | 86 | 42.38 | 0.49 | 12.04 | 0.0053 ** |
| Error | 5 | 0.20 | 0.04 | ||
| Turkey | |||||
| Source | DF | SS | Mean Square | F Value | Pr > F |
| Block | 5 | 0.82 | 0.16 | 6.99 | 0.0261 |
| Check | 1 | 2.13 | 2.13 | 90.86 | 0.0002 ** |
| Accessions | 86 | 30.14 | 0.35 | 14.93 | 0.0032 ** |
| Error | 5 | 0.11 | 0.02 | ||
| 100-Seed Weight (g) | Minimum | Maximum | Mean | Std. Deviation |
|---|---|---|---|---|
| Pakistan | 1.88 | 5.29 | 3.26 | 0.74 |
| Turkey | 2.16 | 5.32 | 3.33 | 0.59 |
| Overall | 2.17 | 5.31 | 3.29 | 0.59 |
| Population/Country | Polymorphic Loci (%) | Na | Ne | I | He | uHe | Mean GD | GD Range |
|---|---|---|---|---|---|---|---|---|
| Overall population | - | 1.99 | 1.54 | 0.48 | 0.32 | 0.32 | 0.50 | 0.14–0.76 |
| Afghanistan | 74.97 | 1.53 | 1.45 | 0.41 | 0.28 | 0.34 | 0.46 | - |
| Austria | 49.96 | 1.25 | 1.35 | 0.30 | 0.21 | 0.28 | 0.48 | - |
| Bangladesh | 87.37 | 1.74 | 1.57 | 0.48 | 0.33 | 0.37 | 0.44 | - |
| China | 98.44 | 1.98 | 1.66 | 0.56 | 0.38 | 0.41 | 0.46 | - |
| Egypt | 96.73 | 1.94 | 1.63 | 0.54 | 0.36 | 0.40 | 0.41 | - |
| India | 96.73 | 1.95 | 1.65 | 0.55 | 0.37 | 0.41 | 0.48 | - |
| Iran | 98.44 | 1.96 | 1.65 | 0.55 | 0.37 | 0.40 | 0.45 | - |
| Iraq | 49.90 | 1.24 | 1.35 | 0.30 | 0.21 | 0.28 | 0.42 | - |
| Israel | 87.37 | 1.73 | 1.57 | 0.48 | 0.33 | 0.38 | 0.44 | - |
| Jordan | 93.53 | 1.90 | 1.63 | 0.53 | 0.36 | 0.40 | 0.27 | - |
| Morocco | 49.90 | 1.23 | 1.34 | 0.30 | 0.20 | 0.26 | 0.42 | - |
| Pakistan | 99.81 | 1.98 | 1.69 | 0.58 | 0.39 | 0.42 | 0.44 | - |
| Portugal | 96.73 | 1.93 | 1.63 | 0.51 | 0.36 | 0.40 | 0.42 | - |
| Spain | 87.37 | 1.81 | 1.59 | 0.49 | 0.34 | 0.39 | 0.38 | - |
| Syria | 74.97 | 1.61 | 1.50 | 0.42 | 0.28 | 0.34 | 0.38 | - |
| Turkey | 99.82 | 1.99 | 1.68 | 0.58 | 0.39 | 0.42 | 0.53 | - |
| Uzbekistan | 74.97 | 1.62 | 1.52 | 0.43 | 0.29 | 0.35 | 0.48 | - |
| Source of Variation | Df | SS | MS | Est. Var. | % Variations |
|---|---|---|---|---|---|
| Among Countries | 25 | 56719.33 | 3336.43 | 225.00 | 9 |
| Within Country | 68 | 165646.96 | 2179.56 | 2179.56 | 91 |
| Total | 93 | 222366.29 | - | 2404.57 | 100 |
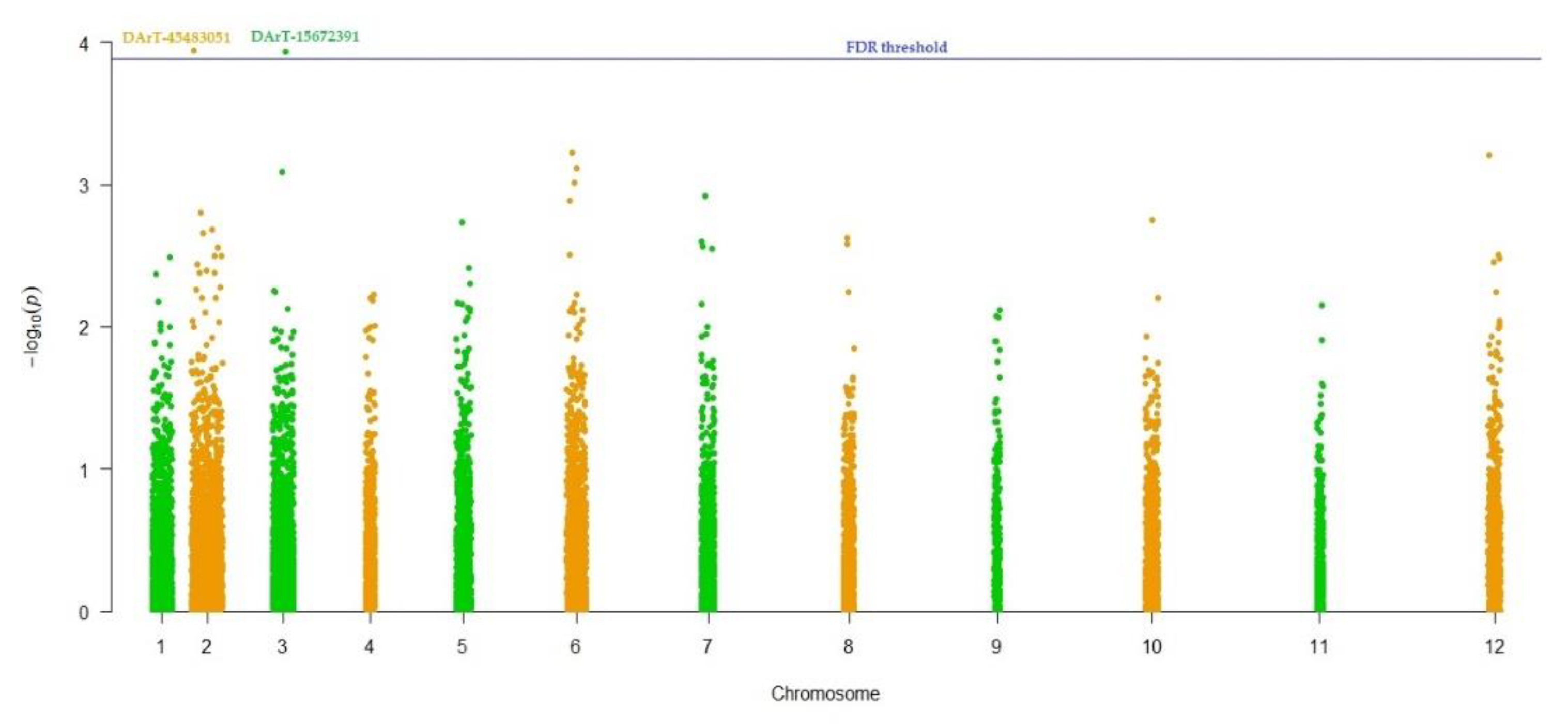
| Trait | Markers | p-Value | R2 | Putative Gene |
|---|---|---|---|---|
| 100-seed weight | DArT-45483051 | 1.17E-04 | 17.40% | AT1G01040 |
| DArT-15672391 | 1.15E-04 | 18.60% | AT5G58040 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, F.; Nadeem, M.A.; Barut, M.; Habyarimana, E.; Chaudhary, H.J.; Khalil, I.H.; Alsaleh, A.; Hatipoğlu, R.; Karaköy, T.; Kurt, C.; et al. Genetic Diversity, Population Structure and Marker-Trait Association for 100-Seed Weight in International Safflower Panel Using SilicoDArT Marker Information. Plants 2020, 9, 652. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9050652
Ali F, Nadeem MA, Barut M, Habyarimana E, Chaudhary HJ, Khalil IH, Alsaleh A, Hatipoğlu R, Karaköy T, Kurt C, et al. Genetic Diversity, Population Structure and Marker-Trait Association for 100-Seed Weight in International Safflower Panel Using SilicoDArT Marker Information. Plants. 2020; 9(5):652. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9050652
Chicago/Turabian StyleAli, Fawad, Muhammad Azhar Nadeem, Muzaffer Barut, Ephrem Habyarimana, Hassan Javed Chaudhary, Iftikhar Hussain Khalil, Ahmad Alsaleh, Rüştü Hatipoğlu, Tolga Karaköy, Cemal Kurt, and et al. 2020. "Genetic Diversity, Population Structure and Marker-Trait Association for 100-Seed Weight in International Safflower Panel Using SilicoDArT Marker Information" Plants 9, no. 5: 652. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9050652