Effect of Microalgal Extracts from Chlorella vulgaris and Scenedesmus quadricauda on Germination of Beta vulgaris Seeds

 ,
,  ,
,
Abstract
:1. Introduction
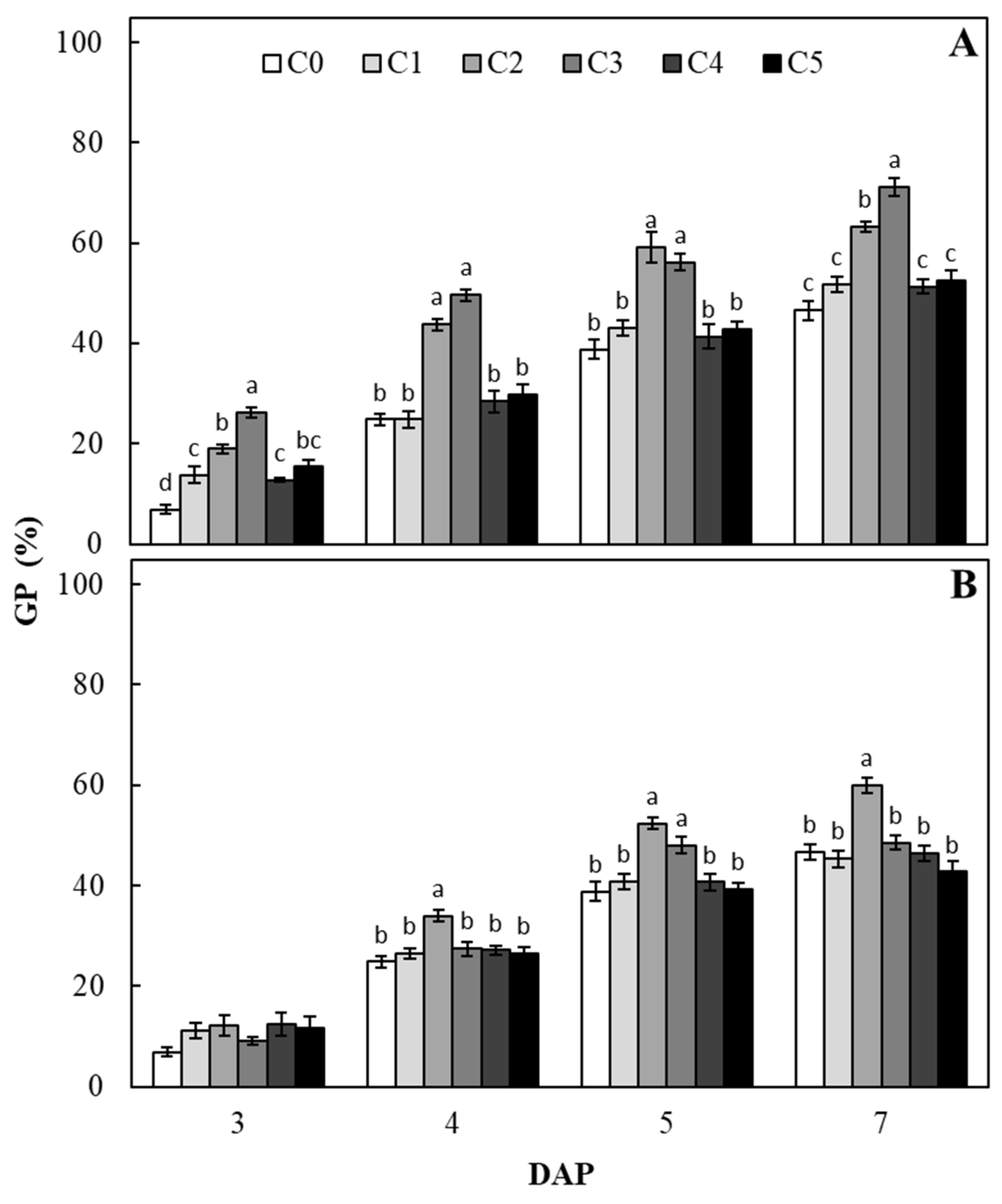
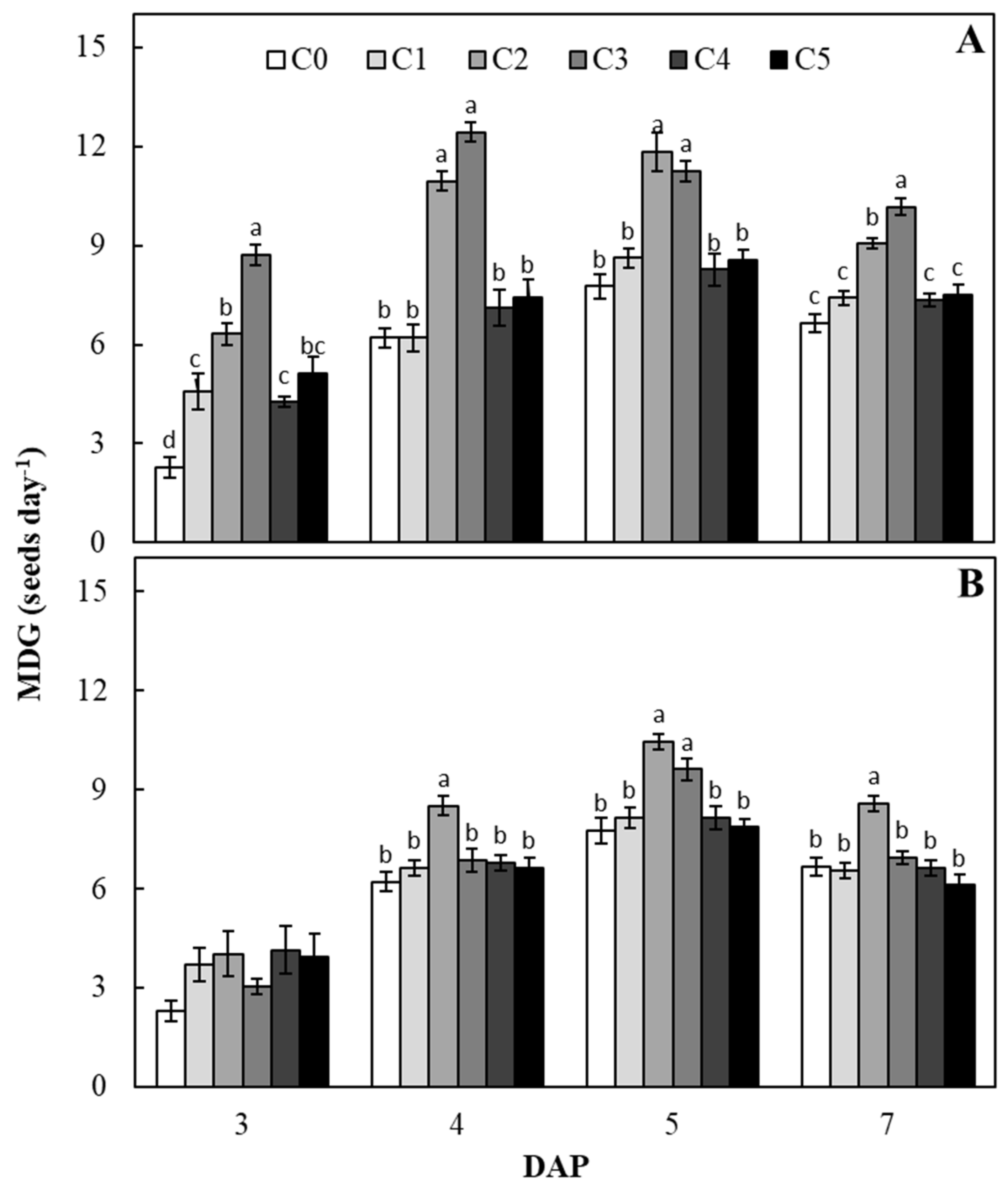
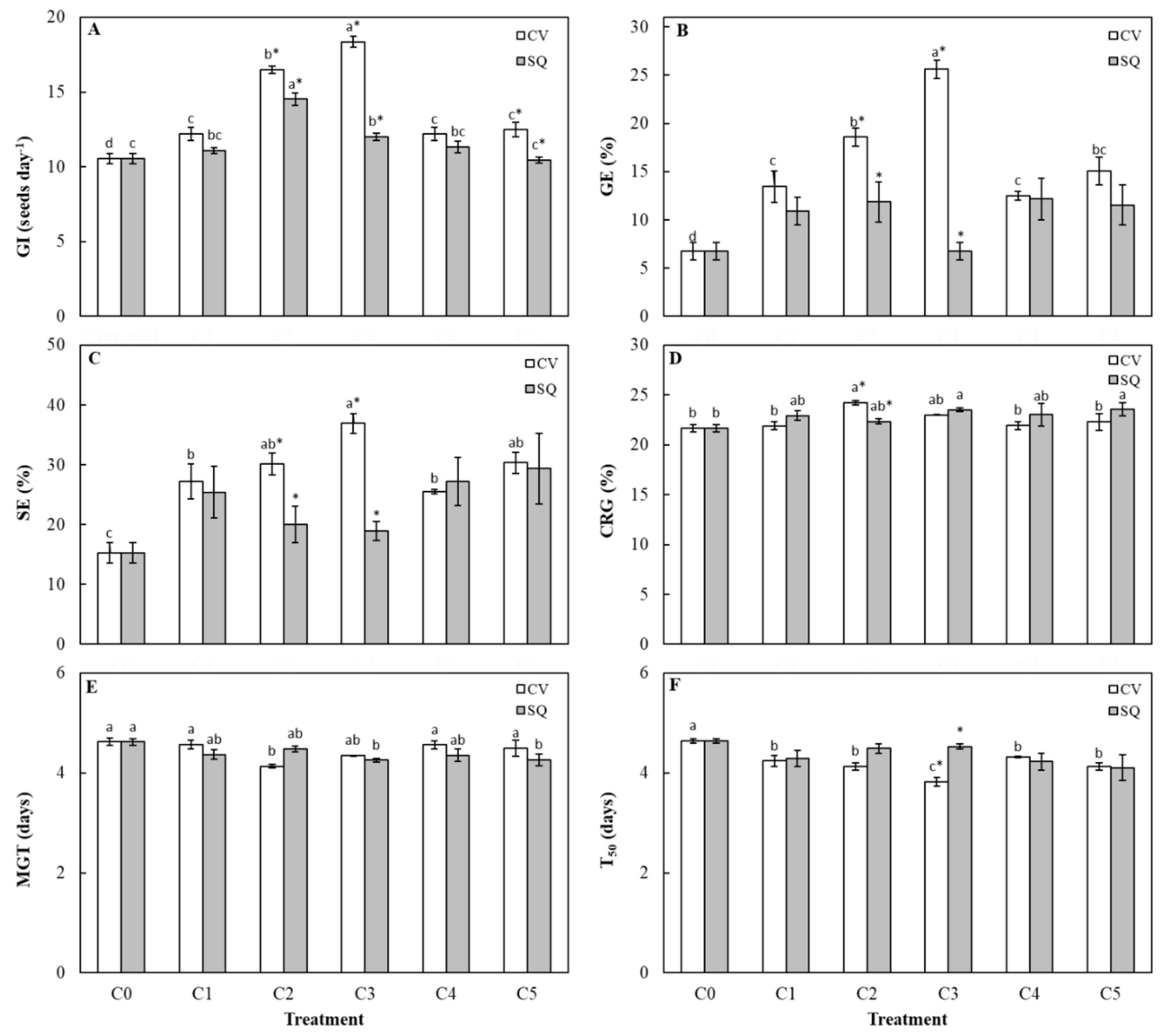
2. Results and Discussions
3. Materials and Methods
3.1. Microalgae Cultivation and Extract Preparation
3.2. Plant Material and Experimental Conditions
3.3. Germination Indices
3.4. Root Morphological Analysis
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mayer, A.M.; Shain, Y. Control of seed germination. Ann. Rev. Plant Physiol. 1974, 25, 167–193. [Google Scholar] [CrossRef]
- Martin, R.C.; Pluskota, W.E.; Nonogaki, H. “Seed Germination” in Plant Developmental Biology—Biotechnological Perspectives; Springer: Berlin/Heidelberg, Germany, 2009; pp. 383–404. [Google Scholar]
- Abdolrahmani, B.; Ghasemi Golezani, K.; Valizadeh, M.; Feizi Asl, V.; Tavakoli, A. Effect of seed priming on the growth trend and grain yield of barley (Hordeum vulgare L.) cv. Abidar under rainfed conditions. Seed Plant Prod. J. 2011, 27, 111–128. [Google Scholar]
- Najafi, G.; Khomari, S.; Javadi, A. Germination response of canola seeds to seed vigor changes and hydro-priming. Seed Res. 2015, 45, 5–70. [Google Scholar]
- Rehman, H.; Iqbal, H.; Basra, S.M.A.; Afzal, I.; Farooq, M.; Wakeel, A.; Ning, W. Seed priming improves early seedling vigor, growth and productivity of spring maize. J. Int. Agric. 2015, 14, 1745–1754. [Google Scholar] [CrossRef]
- Millerd, A.; Thomson, J. Storage proteins of legume seeds: Proteins of legume seeds: Potential for change. CSIRO Div. Plant Ind. Genet. Rep. 1975, 3, 58–68. [Google Scholar]
- Dohm, J.C.; Minoche, A.E.; Holtgräwe, D.; Capella-Gutiérrez, S.; Zakrzewski, F.; Tafer, H.; Rupp, O.; Sörensen, T.R.; Stracke, R.; Reinhardt, R.; et al. The genome of the recently domesticated crop plant sugar beet (Beta vulgaris). Nature 2014, 505, 546–549. [Google Scholar] [CrossRef] [Green Version]
- Zahoor, A.; Faridullah, S.P.; Kakar, K.M.; Sanaullah, B.; El-Sharkawi, H.; Honna, T.; Yamamoto, S. Sugar beet (Beta vulgaris L.) response to different planting methods and row geometries I. Effect on plant growth and yield. Arch. Agron. Soil Sci. 2007, 53, 49–61. [Google Scholar] [CrossRef]
- Vanni, A.; Anfossi, L.; Cignetti, A.; Baglieri, A.; Gennari, M. Degradation of pyrimethanil in soil: Influence of light, oxygen, and microbial activity. J. Environ. Sci. Health B 2006, 41, 67–80. [Google Scholar] [CrossRef]
- Dotto, L.; Silva, V.N. Beet seed priming with growth regulators. Semina: Ciências Agrárias 2017, 38, 1785–1798. [Google Scholar] [CrossRef]
- Kockelmann, A.; Meyer, U. Seed production and quality. In Sugar Beet; Draycott, A.P., Ed.; Blackwell: Oxford, UK, Chapter 5; 2006; pp. 89–112. [Google Scholar]
- Biancardi, E.; McGrath, J.M.; Panella, L.W.; Lewellen, R.T.; Stevanato, P. Sugar Beet. In Handbook of Plant Breeding, Tuberand Root Crops; Bradshaw, J., Ed.; Springer: New York, NY, USA, 2010; Volume 4, pp. 173–219. [Google Scholar]
- Norouzi, P.; Stevanato, P.; Mahmoudi, S.B.; Fasahat, P.; Biancardi, E. Molecular progress in sugar beet breeding for resistance to biotic stresses in sub-arid conditions-current status and perspectives. J. Crop Sci. Biotech. 2017, 20, 99–105. [Google Scholar] [CrossRef]
- Szajsner, H.; Prośba-Białczyk, U.; Sacała, E.; Koszelnik-Leszek, A.; Szubzda, B. The effect of pre-sowing seed stimulation on the germination and pigment content in sugar beet (Beta vulgaris L.) seedlings leaves. Pol. J. Natur. Sci. 2017, 32, 207–222. [Google Scholar]
- Garcia-Gonzalez, J.; Sommerfeld, M. Biofertilizer and biostimulant properties of the microalga Acutodesmus dimorphus. J. Appl. Phycol. 2016, 28, 1051–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, G.; Sahoo, D. Effect of seaweed liquid extract on growth and yield of Triticum aestivum var. Pusa Gold. J. Appl. Phycol. 2011, 23, 251–255. [Google Scholar] [CrossRef]
- Kumari, R.; Kaur, I.; Bhatnagar, A.K. Effect of aqueous extract of Sargassum johnstonii Setchell & Gardner on growth, yield and quality of Lycopersicon esculentum Mill. J. Appl. Phycol. 2011, 23, 623–633. [Google Scholar]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2013, 26, 619–628. [Google Scholar] [CrossRef]
- Barone, V.; Baglieri, A.; Stevanato, P.; Broccanello, C.; Bertoldo, G.; Bertaggia, M.; Cagnin, M.; Pizzeghello, D.; Moliterni, V.M.C.; Mandolino, G.; et al. Root morphological and molecular responses induced by microalgae extracts in sugar beet (Beta vulgaris L.). J. Appl. Phycol. 2018, 30, 1061–1071. [Google Scholar] [CrossRef]
- Barone, V.; Puglisi, I.; Fragalà, F.; Stevanato, P.; Baglieri, A. Effect of living cells of microalgae or their extracts on soil enzyme activities. Arch. Agron. Soil Sci. 2019, 65, 712–726. [Google Scholar] [CrossRef]
- Puglisi, I.; La Bella, E.; Rovetto, E.I.; Lo Piero, A.R.; Baglieri, A. Biostimulant effect and biochemical response in lettuce seedlings treated with a Scenedesmus quadricauda extract. Plants 2020, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Santos, P.L.F.; Zabotto, A.R.; Jordão, H.W.C.; Boas, R.L.V.; Broetto, F.; Tavares, A.R. Use of seaweed-based biostimulant (Ascophyllum nodosum) on ornamental sunflower seed germination and seedling growth. Ornam. Hortic. 2019, 25, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Parađiković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević. M. Biostimulants research in some horticultural plant species—A review. Food Energy Secur. 2019, 8, e00162. [Google Scholar] [CrossRef]
- Lisjak, M.; Tomić, O.; Špoljarević, M.; Teklić, T.; Stanisavljević, A.; Balas, J. Garden cress germinability and seedling vigour after treatment with plant extracts. Agriculture 2015, 21, 41–46. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal biostimulants and biofertilisers in crop productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Baglieri, A.; Cadili, V.; Monterumici, C.M.; Gennari, M.; Tabasso, S.; Montoneri, E.; Nardi, S.; Negre, M. Fertilization of bean plants with tomato plants hydrolysates. Effect on biomass production, chlorophyll content and N assimilation. Sci. Hortic. 2014, 176, 194–199. [Google Scholar] [CrossRef]
- Piccolo, A.; Conte, P.; Spaccini, R.; Mbagwu, J.S.C. Influence of land use on the characteristics of humic substances in some tropical soils of Nigeria. Eur. J. Soil Sci. 2005, 56, 343–352. [Google Scholar] [CrossRef]
- Islam, A.K.M.M.; Kato-Noguchi, H. Phytotoxic activity of Ocimum tenuiflorum extracts on germination and seedling growth of different plant species. Sci. World J. 2014, 2014, 676242. [Google Scholar] [CrossRef] [Green Version]
- Dürr, C.; Boiffin, J.; Fleury, A.; Coulomb, I. Analysis of the variability of sugar beet (Beta vulgaris) growth during the early stages. Agronomy 1992, 12, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Stibbe, C.; Märländer, B. Field emergence dynamics significance to intraspecific competition and growth efficiency in sugar beet (Beta vulgaris L.). Euro. J. Agron. 2002, 3, 161–171. [Google Scholar] [CrossRef]
- Romano, A.; Stevanato, P. Germination Data Analysis by Time-to-Event Approaches. Plants 2020, 9, 617. [Google Scholar] [CrossRef]
- Podlaski, S.; Chomontowski, C. Various methods of assessing sugar beet seed vigour and its impact on the germination process, field emergence and sugar yield. Sugar Tech. 2020, 22, 130–136. [Google Scholar] [CrossRef] [Green Version]
- García, A.C.; de Souza, L.G.A.; Pereira, M.G.; Castro, R.N.; García-Mina, J.M.; Zonta, E.; Lisboa, F.J.G.; Berbara, R.L.L. Structure-property-function relationship in humic substances to explain the biological activity in plants. Sci. Rep. 2016, 6, 20798. [Google Scholar] [CrossRef] [Green Version]
- Spaccini, R.; Piccolo, A.; Contea, P.; Haberhauer, G.; Gerzabek, M.H. Increased soil organic carbon sequestration through hydrophobic protection by humic substances. Soil Biol. Biochem. 2002, 34, 1839–1851. [Google Scholar] [CrossRef]
- Ugena, L.; Hýlová, A.; Podlešáková, K.; Humplík, J.F.; Doležal, K.; De Diego, N.; Spíchal, L. Characterization of biostimulant mode of action using novel multi-trait High-Throughput screening of Arabidopsis germination and Rosette growth. Front. Plant Sci. 2018, 9, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baglieri, A.; Sidella, S.; Barone, V.; Fragalà, F.; Silkina, A.; Nègre, M.; Gennari, M. Cultivating Chlorella vulgaris and Scenedesmus quadricauda microalgae to degrade inorganic compounds and pesticides in water. Environ. Sci. Pollut. Res. 2016, 23, 18165–18174. [Google Scholar] [CrossRef] [PubMed]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barone, V.; Puglisi, I.; Fragalà, F.; Lo Piero, A.R.; Giuffrida, F.; Baglieri, A. Novel bioprocess for the cultivation of microalgae in hydroponic growing system of tomato plants. J. Appl. Phycol. 2019, 31, 465–470. [Google Scholar] [CrossRef]
- Puglisi, I.; Barone, V.; Sidella, S.; Coppa, M.; Broccanello, C.; Gennari, M.; Baglieri, A. Biostimulant activity of humic-like substances from agro-industrial waste on Chlorella vulgaris and Scenedesmus quadricauda. Eur. J. Phycol. 2018, 53, 433–442. [Google Scholar] [CrossRef]
- Hajizadeh, H.S.; Heidari, B.; Bertoldo, G.; Della Lucia, M.C.; Magro, F.; Broccanello, C.; Baglieri, A.; Puglisi, I.; Squartini, A.; Campagna, G.; et al. Expression profiling of candidate genes in sugar beet leaves treated with Leonardite-based biostimulant. High-Throughput 2019, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- The International Seed Testing Association. ISTA International Rules for Seed Testing. ISTA 2020 Rules; The International Seed Testing Association: Zürichstr, Bassersdorf, Switzerland, 2020. [Google Scholar]
- Abbate, C.; Borzì, D.; Caboni, P.; Baglieri, A.; Gennari, M. Behavior of fenhexamid in soil and water. J. Environ. Sci. Health B 2007, 42, 843–849. [Google Scholar] [CrossRef]
- Noorhosseini, S.A.; Jokar, N.K.; Damalas, C.A. Improving seed germination and early growth of garden cress (Lepidium sativum) and Basil (Ocimum basilicum) with hydropriming. J. Plant Growth Regul. 2018, 37, 323–334. [Google Scholar] [CrossRef]
- Maguire, J.D. Speed of germination—Aid in selection and evaluation for seedling emergence and vigor. Crop Sci. 1962, 2, 176–177. [Google Scholar] [CrossRef]
- Soltani, E.; Ghaderi-Far, F.; Baskin, C.C.; Baskin, J.M. Problems with using mean germination time to calculate rate of seed germination. Aust. J. Bot. 2015, 63. [Google Scholar] [CrossRef]
- Ruan, S.; Xue, Q.; Tylkowska, K. The influence of priming on germination of rice (Oryza sativa L.) seeds and seedling emergence and performance in flooded soil. Seed Sci. Technol. 2002, 30, 61–67. [Google Scholar]
- Chiapusio, G.; S´anchez, A.M.; Reigosa, M.J.; Gonzalez, L.; Pellissier, F. Do germination indices adequately reflect allelochemical effects on the germination process? J. Chem. Ecol. 1997, 23, 2445–2453. [Google Scholar] [CrossRef]
- Coolbear, P.; Francis, A.; Grierson, D. The effect of low temperature pre-sowing treatment on the germination performance and membrane integrity of artificially aged tomato seeds. J. Exp. Bot. 1984, 35, 1609–1617. [Google Scholar] [CrossRef]
- Barone, V.; Bertoldo, G.; Magro, F.; Broccanello, C.; Puglisi, I.; Baglieri, A.; Cagnin, M.; Concheri, G.; Squartini, A.; Pizzeghello, D.; et al. Molecular and morphological changes induced by Leonardite-based biostimulant in Beta vulgaris L. Plants 2019, 8, 181. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Length (cm) | Surface Area (cm2) | Mean Root Diameter (mm) | Root Volume (cm3) | Tips Number | Root Number (0.000 < L < 0.500) | Root Number (0.500 < L < 1.000) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | |
| C0 | 0.8 ± 0.1 d | 3.0 ± 0.2 c | 0.2 ± 0.01 e | 0.3 ± 0.01 c | 0.6 ± 0.01 ab | 0.6 ± 0.02 a | 0.003 ± 0.0 d | 0.01 ± 0.0 ab | 1.5 ± 0.07 e | 2.9 ± 0.1 e | 0.2 ± 0.04 b | 1.4 ± 0.1 d | 0.6 ± 0.02 c | 1.4 ± 0.1 a |
| C1 | 2.8 ± 0.2 c | 4.5 ± 0.2 b | 0.5 ± 0.05 d | 0.6 ± 0.05 b | 0.6 ± 0.02 b | 0.4 ± 0.04 bc | 0.01 ± 0.0 c | 0.01 ± 0.0 bc | 3.0 ± 0.14 d | 5.04 ± 0.2 c | 3.0 ± 1.1 a | 3.2 ± 0.2 bc | 1.2 ± 0.2 b | 1.2 ± 0.2 a–c |
| C2 | 4.7 ± 0.1 a | 6.7 ± 0.1 a | 0.8 ± 0.01 b | 0.9 ± 0.03 a | 0.6 ± 0.01 b | 0.4 ± 0.02 c | 0.01 ± 0.0 ab | 0.01 ± 0.0 a | 3.7 ± 0.22 c | 8.0 ± 0.2 b | 2.3 ± 0.6 a | 5.3 ± 0.04 a | 2.1 ± 0.2 a | 1.3 ± 0.06 a–c |
| C3 | 5.1 ± 0.1 a | 6.4 ± 0.2 a | 0.9 ± 0.02 a | 0.9 ± 0.01 a | 0.6 ± 0.02 b | 0.4 ± 0.01 c | 0.01 ± 0.0 a | 0.01 ± 0.0 a–c | 4.6 ± 0.18 a | 11.2 ± 0.2 a | 2.7 ± 0.4 a | 5.3 ± 0.2 a | 2.3 ± 0.2 a | 1.05 ± 0.1 bc |
| C4 | 3.2 ± 0.2 bc | 4.2 ± 0.1 b | 0.6 ± 0.02 c | 0.6 ± 0.01 b | 0.6 ± 0.02 a | 0.5 ± 0.01 b | 0.01 ± 0.0 b | 0.01 ± 0.0 a–c | 4.3 ± 0.14 ab | 4.4 ± 0.1 d | 1.7 ± 0.3 ab | 2.7 ± 0.2 c | 1.2 ± 0.1 b | 1.3 ± 0.1 ab |
| C5 | 3.5 ± 0.1 b | 4.5 ± 0.2 b | 0.6 ± 0.02 c | 0.6 ± 0.01 b | 0.6 ± 0.02 ab | 0.4 ± 0.01 c | 0.01 ± 0.0 b | 0.01 ± 0.0 c | 4.1 ± 0.11 bc | 5.2 ± 0.2 c | 2.5 ± 0.7 a | 3.4 ± 0.2 b | 1.2 ± 0.1 b | 1.0 ± 0.1 c |
| Length (cm) | Surface Area (cm2) | Mean Root Diameter (mm) | Root Volume (cm3) | Tips Number | Root Number (0.000 < L < 0.500) | Root Number (0.500 < L < 1.000) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | 5 days | 7 days | |
| C0 | 0.8 ± 0.1 d | 3.0 ± 0.2 d | 0.2 ± 0.01 c | 0.3 ± 0.01 d | 0.6 ± 0.01 b | 0.6 ± 0.02 a | 0.003 ± 0.0 c | 0.01 ± 0.0 ab | 1.5 ± 0.07 d | 2.9 ± 0.1 b | 0.2 ± 0.04 c | 1.4 ± 0.1 e | 0.6 ± 0.02c | 1.4 ± 0.1 bc |
| C1 | 2.7 ± 0.1 c | 3.9 ± 0.1 c | 0.5 ± 0.01 b | 0.6 ± 0.03 c | 0.6 ± 0.01 b | 0.5 ± 0.01 c | 0.01 ± 0.0 ab | 0.01 ± 0.0 b | 2.5 ± 0.11 c | 3.1 ± 0.1 b | 1.1 ± 0.1 a | 2.8 ± 0.1 bc | 1.9 ± 0.1a | 1.3 ± 0.1 c |
| C2 | 5.2 ± 0.3 a | 6.2 ± 0.2 a | 0.7 ± 0.03 a | 0.9 ± 0.03 a | 0.7 ± 0.01 ab | 0.5 ± 0.01 c | 0.01 ± 0.0 bc | 0.01 ± 0.0 a | 2.8 ± 0.13 c | 5.3 ± 0.2 a | 0.5 ± 0.2 bc | 4.1 ± 0.2 a | 1.2 ± 0.4b | 1.9 ± 0.1 a |
| C3 | 3.5 ± 0.2 b | 5.4 ± 0.3 b | 0.7 ± 0.04 a | 0.9 ± 0.01 a | 0.6 ± 0.04 b | 0.5 ± 0.02 c | 0.01 ± 0.0 a | 0.01 ± 0.0 a | 3.4 ± 0.14 b | 5.0 ± 0.2 a | 1.2 ± 0.3 a | 3.3 ± 0.2 b | 2.1 ± 0.1a | 1.8 ± 0.1 a |
| C4 | 2.9 ± 0.04 c | 3.6 ± 0.2 c | 0.5 ± 0.02 b | 0.7 ± 0.02 b | 0.7 ± 0.02 b | 0.6 ± 0.04 ab | 0.01 ± 0.0 a | 0.01 ± 0.0 ab | 4.0 ± 0.12 a | 4.7 ± 0.2 a | 1.0 ± 0.2 ab | 1.9 ± 0.2 de | 2.1 ± 0.1a | 1.6 ± 0.04 a–c |
| C5 | 2.6 ± 0.1 c | 3.9 ± 0.1 c | 0.5 ± 0.01 b | 0.7 ± 0.01 b | 0.8 ± 0.06 a | 0.5 ± 0.3 bc | 0.01 ± 0.0 a | 0.01 ± 0.0 ab | 2.6 ± 0.3 c | 5.1 ± 0.4 a | 0.6 ± 0.1 bc | 2.5 ± 0.2 cd | 1.8 ± 0.1 a | 1.8 ± 0.3 ab |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puglisi, I.; Barone, V.; Fragalà, F.; Stevanato, P.; Baglieri, A.; Vitale, A. Effect of Microalgal Extracts from Chlorella vulgaris and Scenedesmus quadricauda on Germination of Beta vulgaris Seeds. Plants 2020, 9, 675. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9060675
Puglisi I, Barone V, Fragalà F, Stevanato P, Baglieri A, Vitale A. Effect of Microalgal Extracts from Chlorella vulgaris and Scenedesmus quadricauda on Germination of Beta vulgaris Seeds. Plants. 2020; 9(6):675. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9060675
Chicago/Turabian StylePuglisi, Ivana, Valeria Barone, Ferdinando Fragalà, Piergiorgio Stevanato, Andrea Baglieri, and Alessandro Vitale. 2020. "Effect of Microalgal Extracts from Chlorella vulgaris and Scenedesmus quadricauda on Germination of Beta vulgaris Seeds" Plants 9, no. 6: 675. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9060675