Photo-Protective Mechanisms and the Role of Poly (ADP-Ribose) Polymerase Activity in a Facultative CAM Plant Exposed to Long-Term Water Deprivation



Abstract
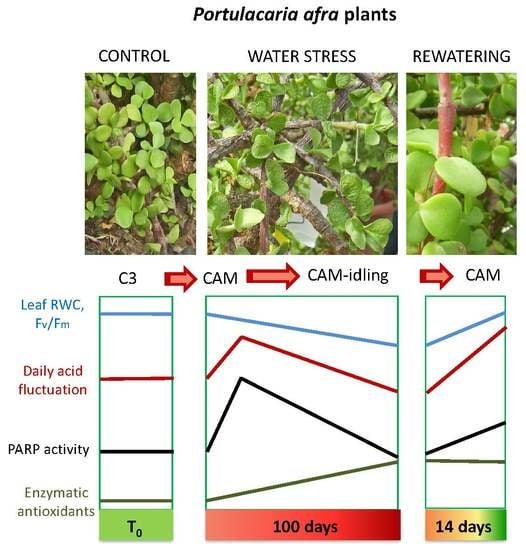
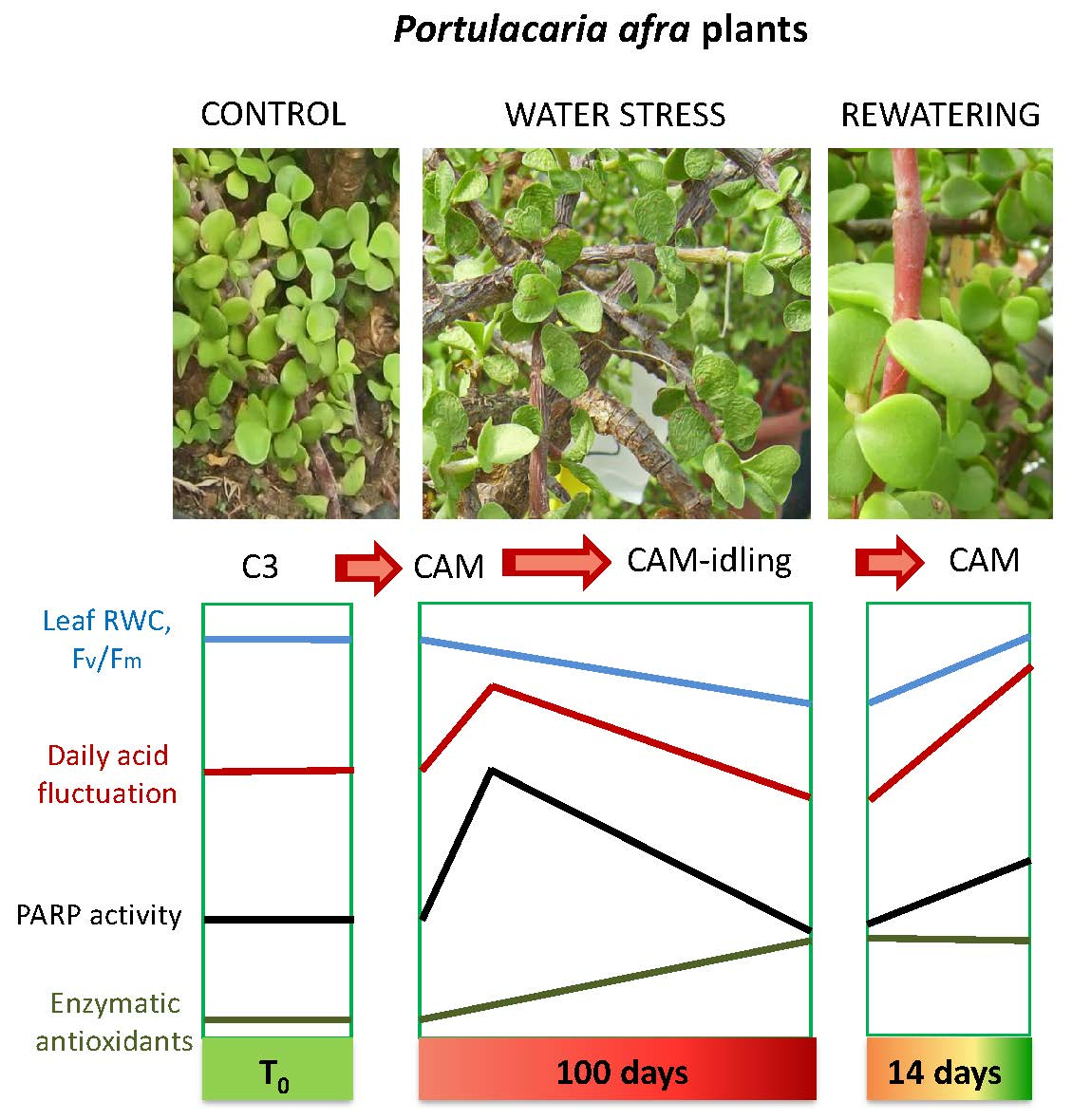
:
1. Introduction
2. Results
2.1. Leaf Water Status and Photosynthetic Pigment Content
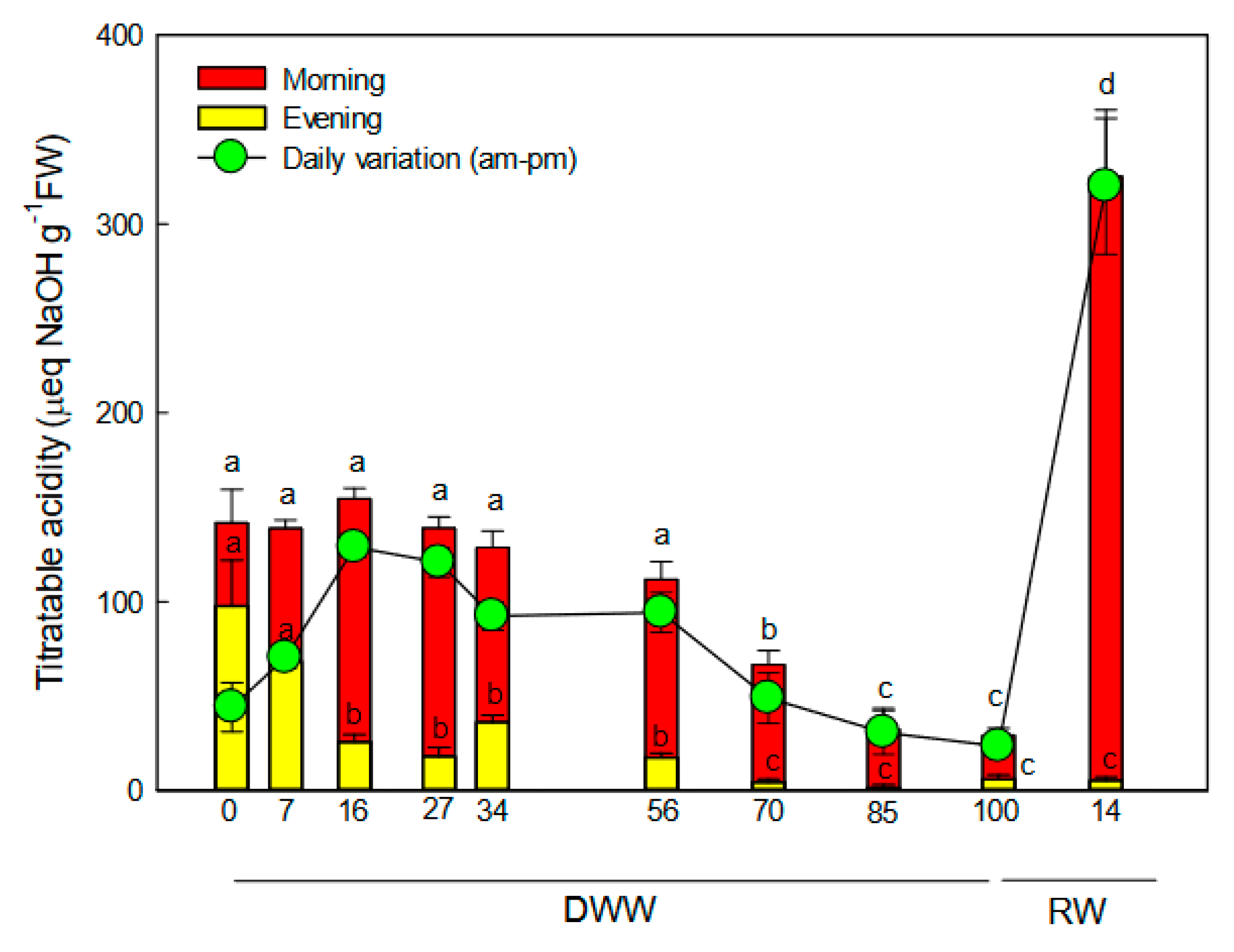
2.2. Titratable Acidity Fluctuation
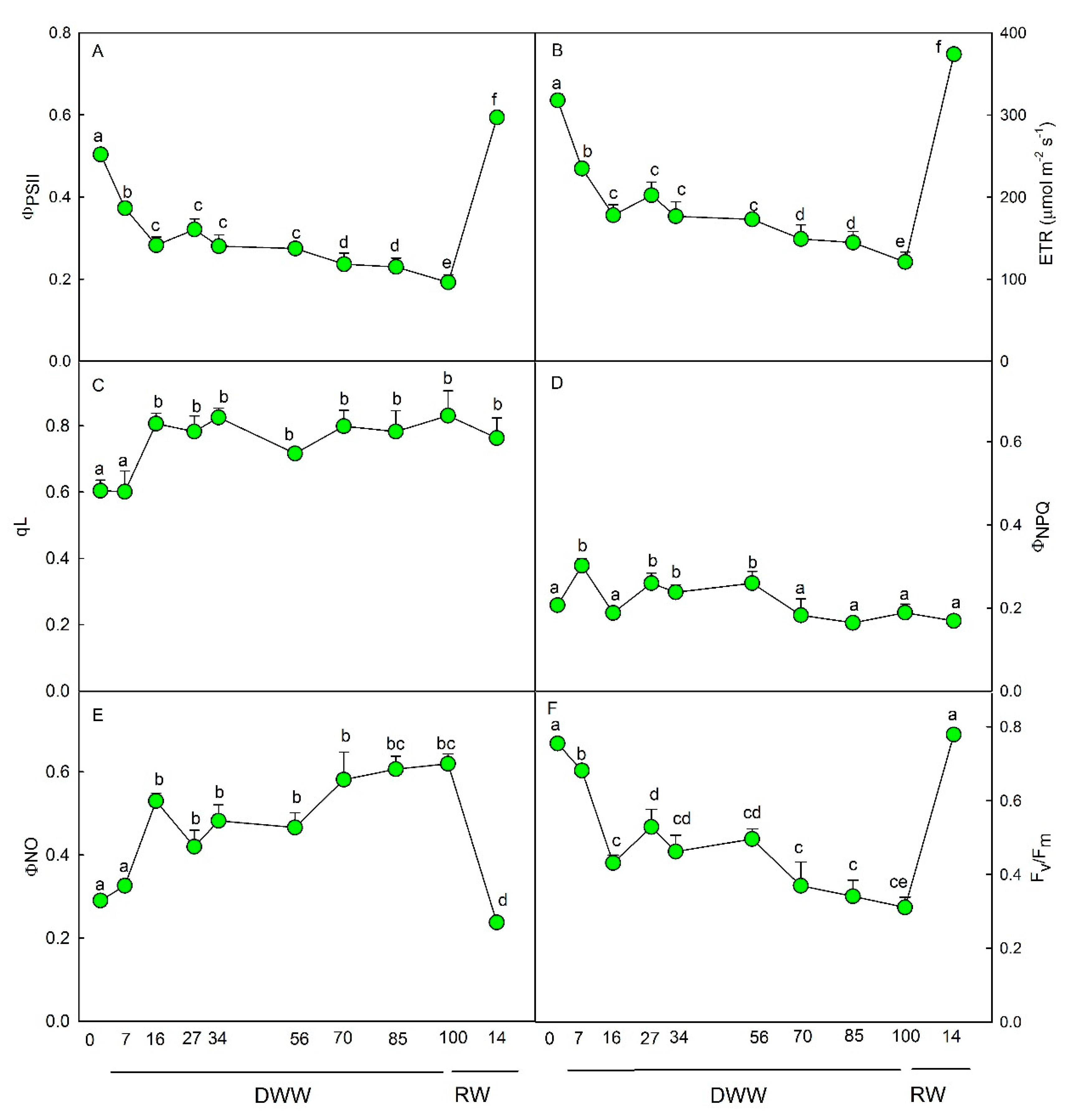
2.3. Fluorescence Measurements
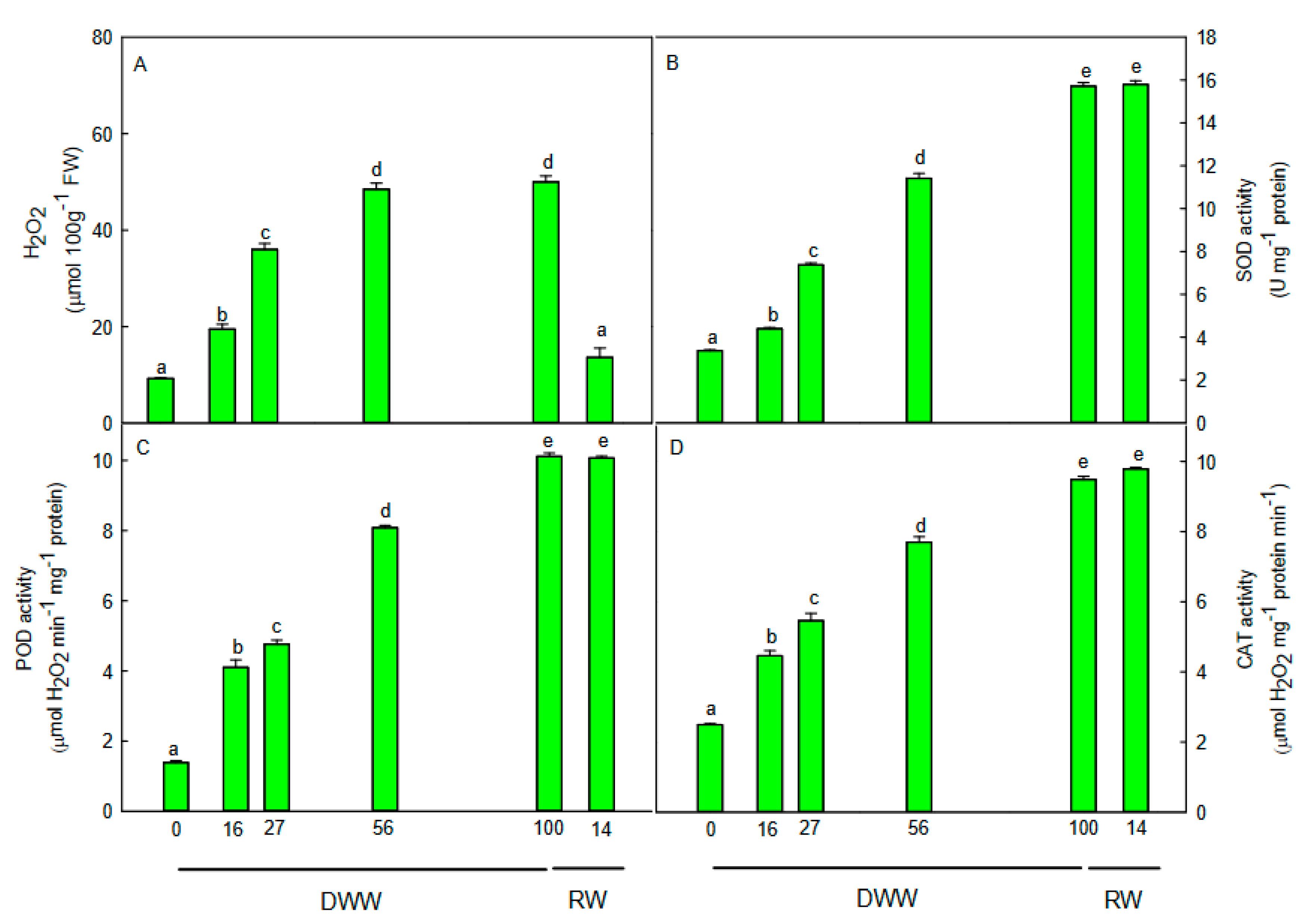
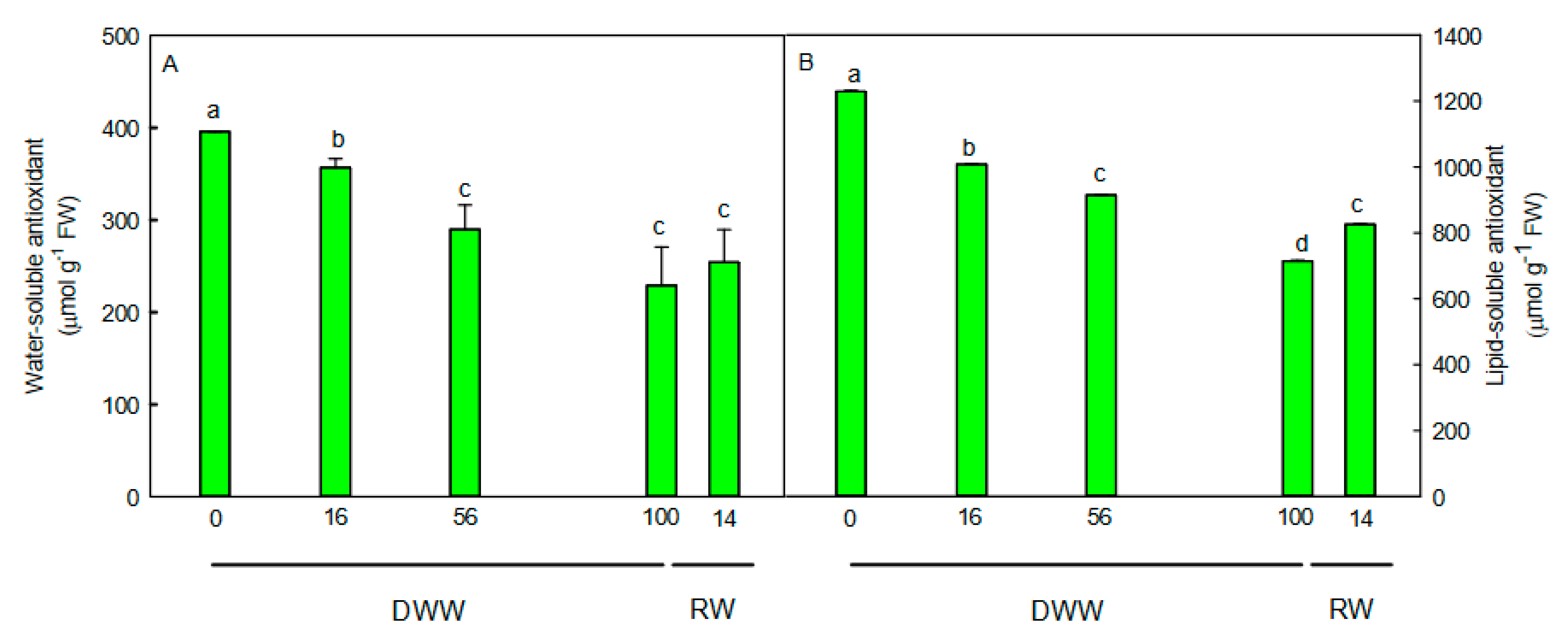
2.4. H2O2 Content, Non-Enzymatic and Enzymatic Antioxidants
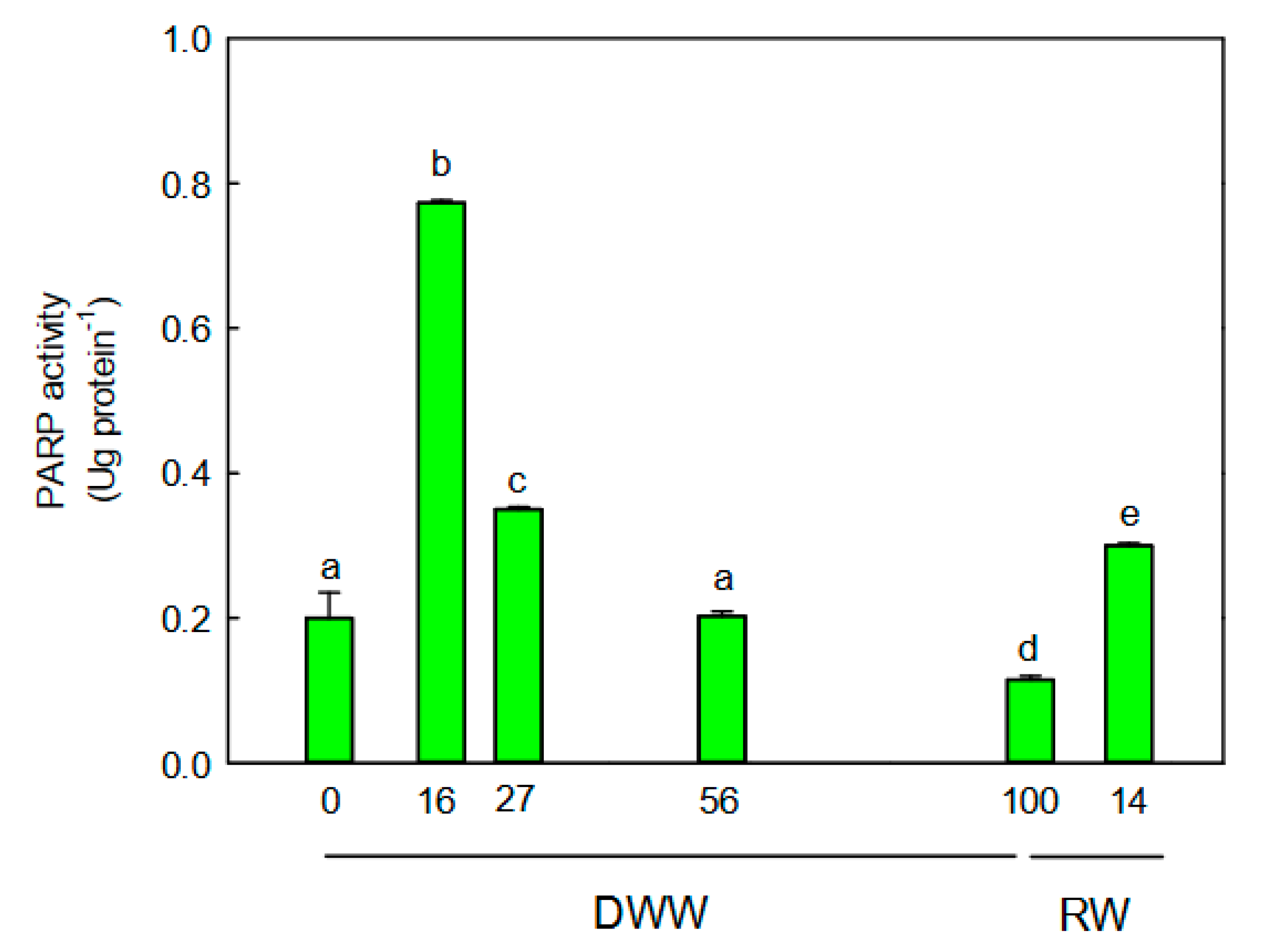
2.5. PARP Activity
2.6. Correlations among Investigated Parameters
3. Discussion
4. Materials and Methods
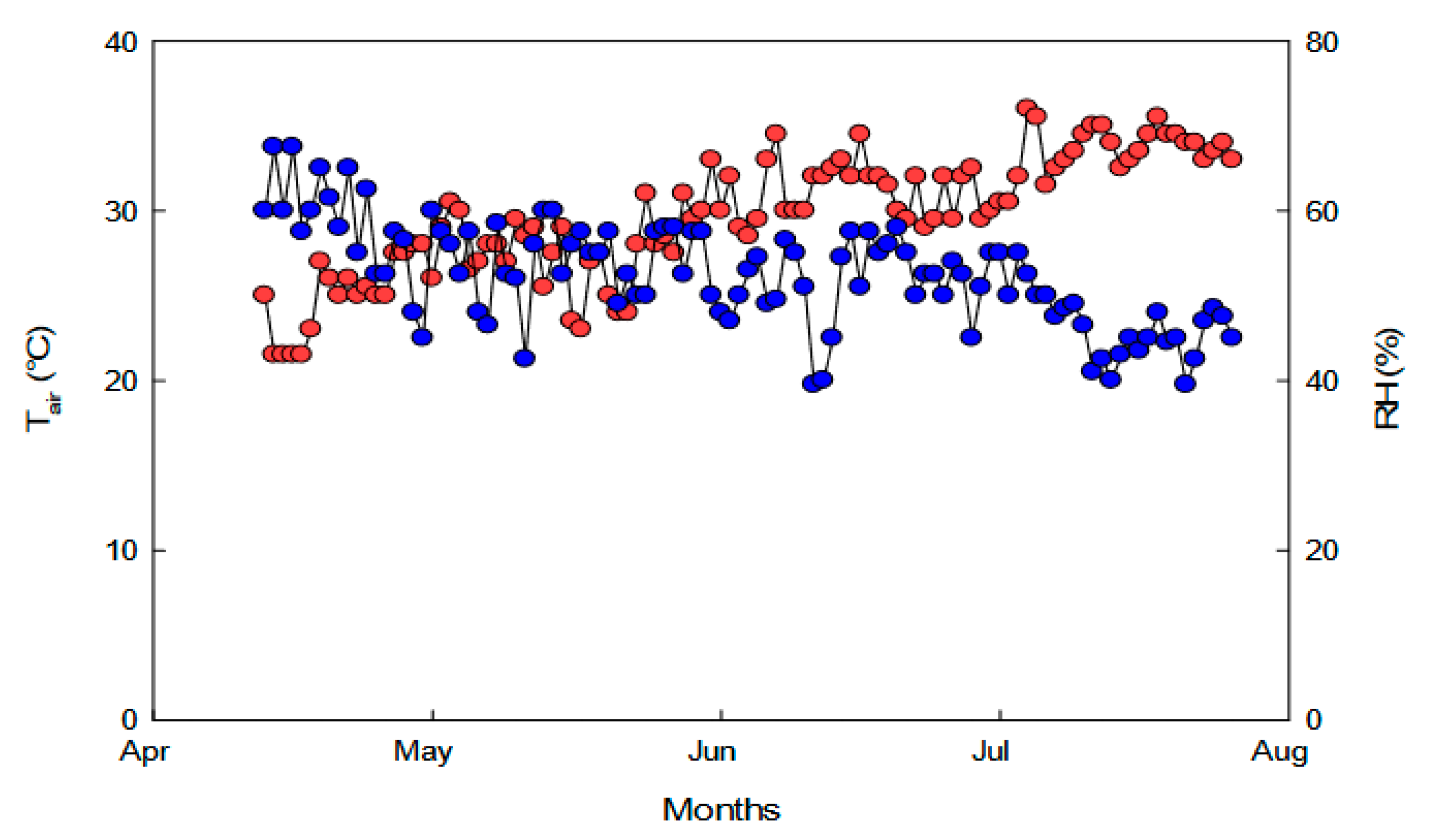
4.1. Plants Growth Condition and Experimental Design
4.2. Leaf Water Status and Photosynthetic Pigment Content Measurement
4.3. Titratable Acidity Determination
4.4. Chlorophyll a Fluorescence Emission Measurements
4.5. Hydrogen Peroxide Determination
4.6. Non-Enzymatic and Enzymatic Antioxidant Measurements
4.7. Poly (ADP-Ribose) Polymerase (PARP) Assay Activity
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sunagawa, H.; Cushman, J.C.; Agarie, S. Crassulacean acid metabolism may alleviate production of reactive oxygen species in a facultative CAM plant, the common ice plant Mesembryanthemum crystallinum L. Plant Prod. Sci. 2010, 13, 256–260. [Google Scholar] [CrossRef]
- Lüttge, U. Ability of crassulacean acid metabolism plants to overcome interacting stresses in tropical environments. AoB Plants 2010, 2010, plq005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osmond, C.B.; Popp, M.; Robinson, S.A. Stoichiometric nightmares: Studies of photosynthetic O2 and CO2 exchange in CAM plants. In Crassulacean Acid Metabolism. Biochemistry, Ecophysiology and Evolution. Ecological Studies, 114; Winter, K., Smith, J.A.C., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1996; pp. 19–30. [Google Scholar]
- Abreu, M.E.; Carvalho, V.; Mercier, H. Antioxidant capacity along the leaf blade of the C3-CAM facultative bromeliad Guzmania monostachia under water deficit conditions. Funct. Plant Biol. 2018, 45, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Qui, N.; Lu, Q.; Wang, B.; Kuang, T. PSII photochemistry, thermal energy dissipation, and the xanthophyll cycle in Kalanchoë daigremontiana exposed to a combination of water stress and high light. Physiol. Plant. 2003, 118, 173–182. [Google Scholar] [CrossRef]
- Pieters, A.J.; Tezara, W.; Herrera, A. Operating of the xanthophyll cycle and degradation of D1 protein in the inducible CAM plant, Talinum triangulare, under water deficit. Ann. Bot. 2003, 92, 393–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, F.J. Antioxidative protection in the inducible CAM plant Sedum album L. following the imposition of severe water stress and recovery. Oecologia 1996, 107, 469–477. [Google Scholar] [CrossRef]
- Broetto, F.; Lüttge, U.; Ratajczak, R. Influence of light intensity and salt-treatment on mode of photosynthesis and enzymes of the antioxidative response system of Mesembryanthemum crystallium L. Funct. Plant Biol. 2002, 29, 13–23. [Google Scholar] [CrossRef]
- Nosek, M.; Gawrońska, K.; Rozpądek, P.; Szechyńska-Hebd, M.; Kornaś, A.; Miszalskic, Z. Withdrawal from functional Crassulacean acid metabolism (CAM) is accompanied by changes in both gene expression and activity of antioxidative enzymes. J. Plant Physiol. 2018, 229, 151–157. [Google Scholar] [CrossRef]
- Cela, J.; Arrom, L.; Munne´-Bosch, S. Diurnal changes in photosystem II photochemistry, photoprotective compounds and stress-related phytohormones in the CAM plant, Aptenia cordifolia. Plant Sci. 2009, 177, 404–410. [Google Scholar] [CrossRef]
- Maxwell, K.; von Caemmerer, S.; Evans, J.R. Is a low internal conductance to CO2 diffusion a consequence of succulence in plants with crassulacean acid metabolism? Funct. Plant Biol. 1997, 24, 777–786. [Google Scholar] [CrossRef]
- Borland, A.M.; Técsi, L.I.; Leegood, R.C.; Walker, R.P. Inducibility of crassulacean acid metabolism (CAM) in Clusia species; physiological/biochemical characterization and intercellular localization of carboxylation and decarboxylation processes in three species which exhibit different degrees of CAM. Planta 1998, 205, 342–351. [Google Scholar] [CrossRef]
- Arena, C.; Mistretta, C.; Natale, E.; Mennella, M.R.F.; De Maio, A. Characterization and role of poly(ADP-ribosyl)ation in the Mediterranean species Cistus incanus L. under different temperature conditions. Plant Physiol. Biochem. 2011, 49, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; de Micco, V.; de Maio, A.; Mistretta, C.; Aronne, G.; Vitale, L. Winter and summer leaves of Cistus incanus: Differences in leaf morphofunctional traits, photosynthetic energy partitioning, and poly(ADP-ribose) polymerase (PARP) activity. Botany 2013, 91, 805–813. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, L.; Bianchi, A.R.; Mistretta, C.; Vitale, E.; Parisi, C.; Guerriero, G.; Magliulo, V.; de Maio, A. The Ageing Process Affects the Antioxidant Defences and the Poly (ADPribosyl) ation Activity in Cistus Incanus, L. Leaves. Antioxidants 2019, 8, 528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Z.; Pan, W.; Chen, W.; Lian, Q.; Wu, Q.; Lv, Z.; Cheng, X.; Ge, X. New perspectives on the plant PARP family: Arabidopsis PARP3 is inactive, and PARP1 exhibits predominant poly (ADP-ribose) polymerase activity in response to DNA damage. BMC Plant Biol. 2019, 19, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miszalski, Z.; Niewiadomska, E.; Slesak, I.; Lüttge, U.; Kluge, M.; Ratajczak, R. The Effect of irradiance on carboxylating/decarboxylating enzymes and fumarase activities in Mesembryanthemum crystallinum L. exposed to salinity stress. Plant Biol. 2001, 3, 17–23. [Google Scholar] [CrossRef]
- Guralnick, L.J.; Gladsky, K. Crassulacean acid metabolism as a continuous trait: Variability in the contribution of Crassulacean acid metabolism (CAM) in populations of Portulacaria afra. Heliyon 2017, 3, e00293. [Google Scholar] [CrossRef] [Green Version]
- Guralnick, L.J.; Rorabaugh, P.A.; Hanscom, Z. Seasonal shift of photosynthesis in Portulacaria afra (L.) Jacq. Plant Physiol. 1984, 76, 643–646. [Google Scholar] [CrossRef] [Green Version]
- Guralnick, L.J.; Ting, I.P. Seasonal response to drought and re-watering in Portulacaria afra (L.) Jacq. Oecologia 1987, 70, 85–91. [Google Scholar] [CrossRef]
- Niewiadomska, E.; Borland, A.M. Crassulacean Acid Metabolism: A Cause or Consequence of Oxidative Stress in Planta? In Progress in Botany, 69; Lüttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Guralnick, L.J.; Gilbert, K.E.; Denio, D.; Antico, N. The Development of Crassulacean Acid Metabolism (CAM) Photosynthesis in Cotyledons of the C4 Species, Portulaca grandiflora (Portulacaceae). Plants 2020, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Ślesak, I.; Libik, M.M.; Miszalski, Z. The foliar concentration of hydrogen peroxide during salt-induced C3-CAM transition in Mesembryanthemum crystallinum L. Plant. Sci. 2008, 174, 221–226. [Google Scholar] [CrossRef]
- Habibi, G.; Hajiboland, R. Comparison of photosynthesis and antioxidative protection in Sedum album and Sedum stoloniferum (Crassulaceae) under water stress. Photosynthetica 2012, 50, 508–518. [Google Scholar] [CrossRef]
- Noctor, G.; Veljovic-Jovanovic, S.; Foyer, C.H. Peroxide processing in photosynthesis: Antioxidant coupling and redox signalling. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2000, 355, 1465–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racchi, M.L. Antioxidant defenses in plants with attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef]
- Azevedo, R.A.; Alas, R.M.; Smith, R.J.; Lea, P.J. Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild-type and a catalase-deficient mutant of barley. Physiol. Plant. 1998, 104, 280–292. [Google Scholar] [CrossRef]
- Herppich, W.B.; Peckmann, K. Influence of drought on mitochondrial activity, photosynthesis, nocturnal acid accumulation and water relations in the CAM plants Prenia sladeniana (ME-type) and Crassula lycopodioides (PEPCK-type). Ann. Bot. 2000, 86, 611–620. [Google Scholar] [CrossRef]
- Peckmann, K.; Martin, C.E.; von Willert, D.J.; Herppich, W.B. Biochemical changes in intact leaf mitochondria throughout the NaCl-induced transition from C3 to CAM in the facultatively halophytic CAM plant Mesembryanthemum crystallinum. In Phyton-Annales Rei Botanicae; Ferdinand Berger Soehne: Horn, Austria, 2019; Volume 59, pp. 109–118. [Google Scholar]
- Pikart, F.C.; Matiza, A.; Alvesa, F.R.R.; Merciera, H. Diurnal modulation of PEPCK decarboxylation activity impacts photosystem II light-energy use in a drought-induced CAM species. Environ. Exp. Bot. 2020, 173, 104003. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustakas, M. A better energy allocation of absorbed light in photosystem II and less photooxidative damage contribute to acclimation of Arabidopsis thaliana young leaves to water deficit. J. Plant. Physiol. 2004, 171, 587–593. [Google Scholar] [CrossRef]
- Arena, C.; de Micco, V.; de Maio, A. Growth alteration and leaf biochemical responses in P. vulgaris plants exposed to different doses of ionizing radiation. Plant Biol. 2014, 16, 194–202. [Google Scholar] [CrossRef]
- Giorio, P.; Guida, G.; Mistretta, C.; Sellami, M.H.; Oliva, M.; Punzo, P.; Iovieno, P.; Arena, C.; De Maio, A.; Grillo, S.; et al. Physiological, biochemical and molecular responses to water stress and rehydration in Mediterranean adapted tomato landraces. Plant Biol. 2018, 20, 995–1004. [Google Scholar] [CrossRef]
- de Block, M.; Verduyn, C.; de Brouwer, D.; Cornelissen, M. Poly (ADPribose) polymerase in plants affects energy homeostasis, cell death and stress tolerance. Plant J. 2005, 41, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Vanderauwera, S.; de Block, M.; van de Steene, N.; van de Cotte, B.; Metzlaff, M.; van Breusegem, F. Silencing of poly(ADP-ribose) polymerase in plants alters abiotic stress signal transduction. Proc. Natl. Acad. Sci. USA 2007, 104, 15150–15155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K. Chlotophylls and Carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Krall, J.P.; Edwards, G.E. Relationship between photosystem II activity and CO2 fixation in leaves. Physiol. Plant. 1992, 86, 180–187. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, J.; Guo, Z.; Tan, H.; Zhu, X. A simple colorimetric method for determination of hydrogen peroxide in plant tissues. Plant Growth Regul. 2006, 49, 113–118. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days Without Water (DWW) | Relative Water Content (%) |
|---|---|
| 0 | 85.5 ± 2.0 a |
| 60 | 66.3 ± 4.3 b |
| 79 | 47.3 ± 3.16 c |
| 94 | 47.9 ± 4.1 c |
| Days of Rewatering (RW) | RWC |
| 14 | 88.9 ± 1.4 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitale, L.; Vitale, E.; Costanzo, G.; De Maio, A.; Arena, C. Photo-Protective Mechanisms and the Role of Poly (ADP-Ribose) Polymerase Activity in a Facultative CAM Plant Exposed to Long-Term Water Deprivation. Plants 2020, 9, 1192. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9091192
Vitale L, Vitale E, Costanzo G, De Maio A, Arena C. Photo-Protective Mechanisms and the Role of Poly (ADP-Ribose) Polymerase Activity in a Facultative CAM Plant Exposed to Long-Term Water Deprivation. Plants. 2020; 9(9):1192. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9091192
Chicago/Turabian StyleVitale, Luca, Ermenegilda Vitale, Giulia Costanzo, Anna De Maio, and Carmen Arena. 2020. "Photo-Protective Mechanisms and the Role of Poly (ADP-Ribose) Polymerase Activity in a Facultative CAM Plant Exposed to Long-Term Water Deprivation" Plants 9, no. 9: 1192. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9091192