Structure of the Chemical and Genetic Diversity of the True Lavender over Its Natural Range

 , and
, and
Abstract
:1. Introduction
2. Results
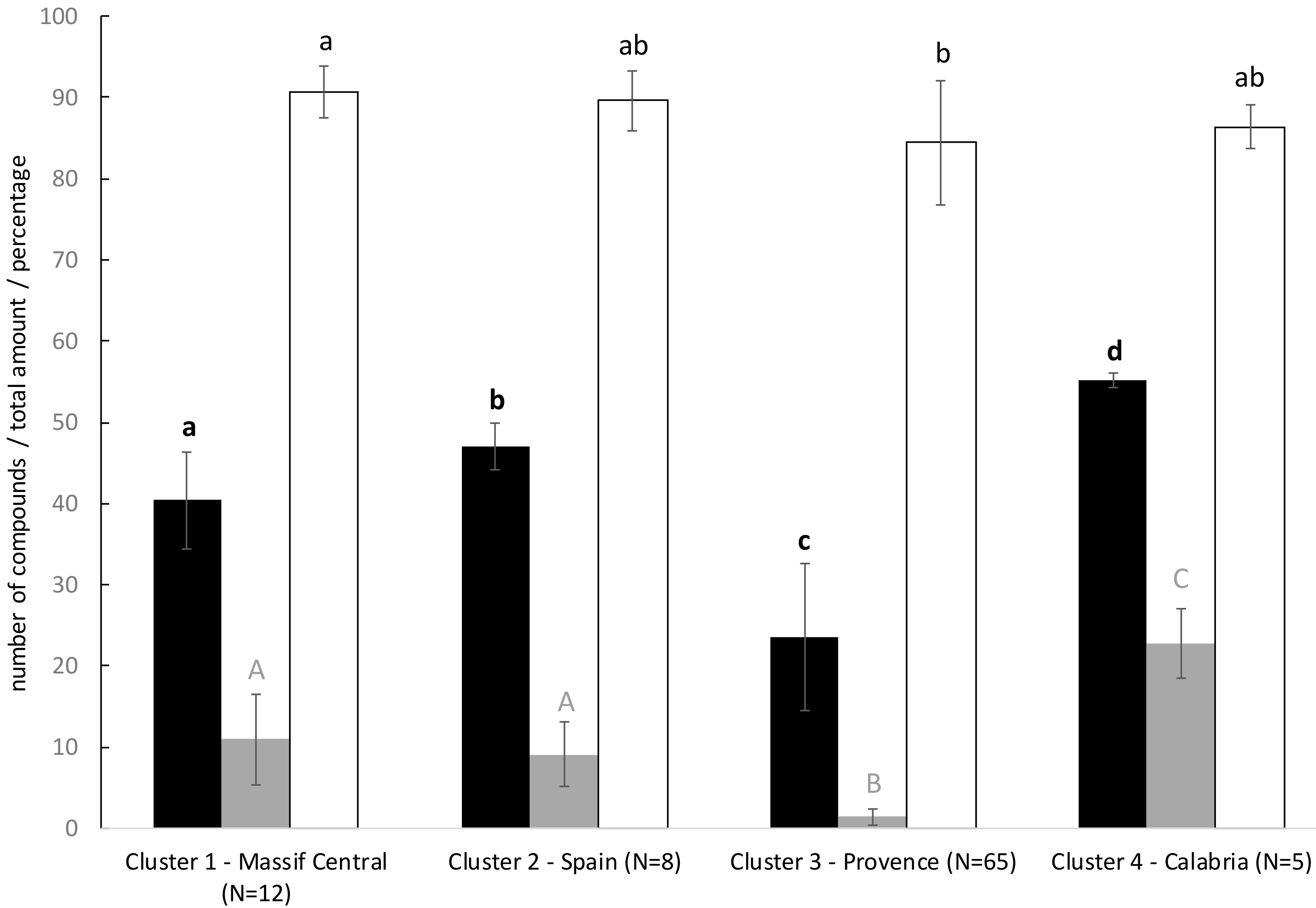
2.1. Global Composition and Amount of VOCs
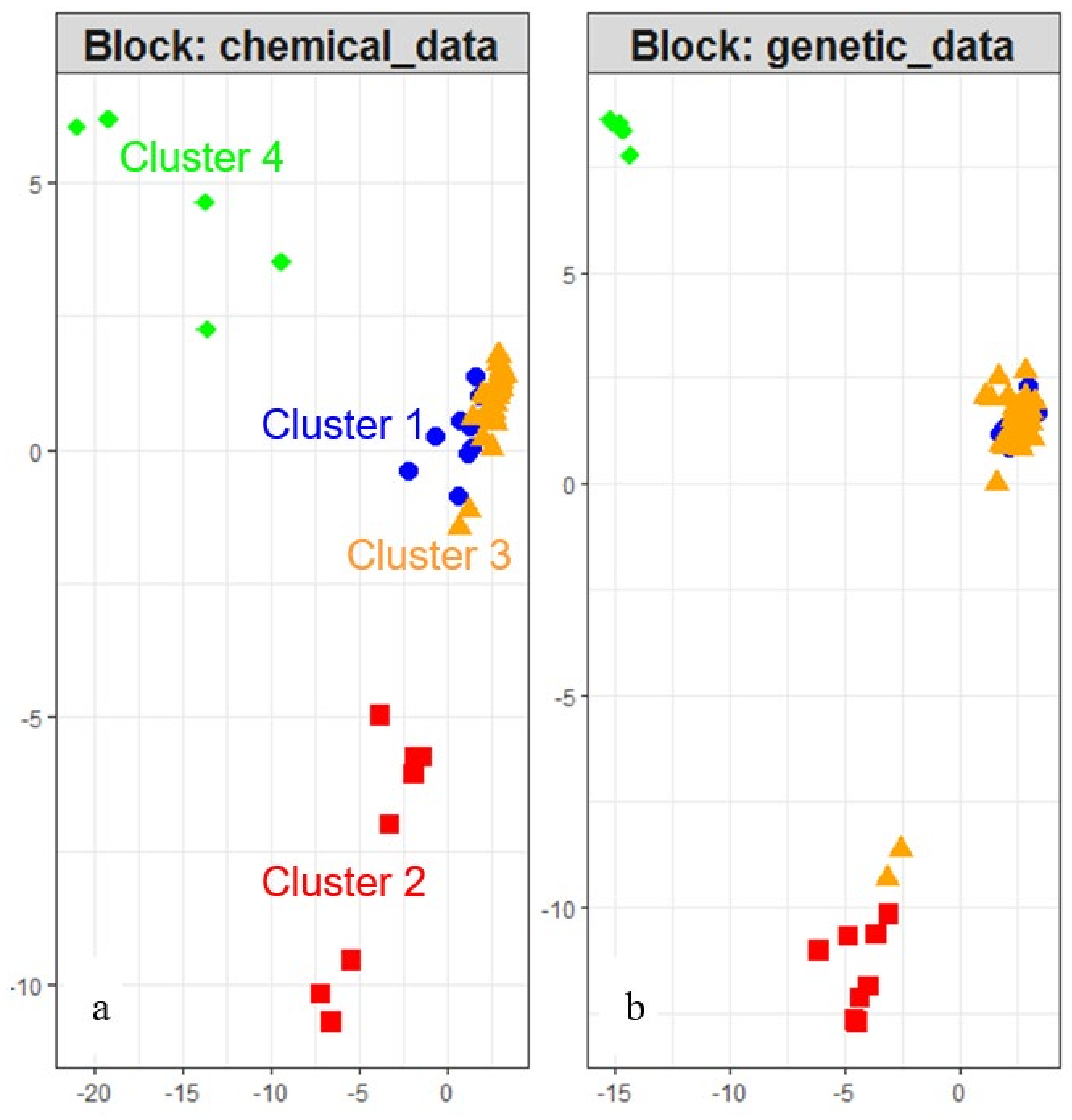
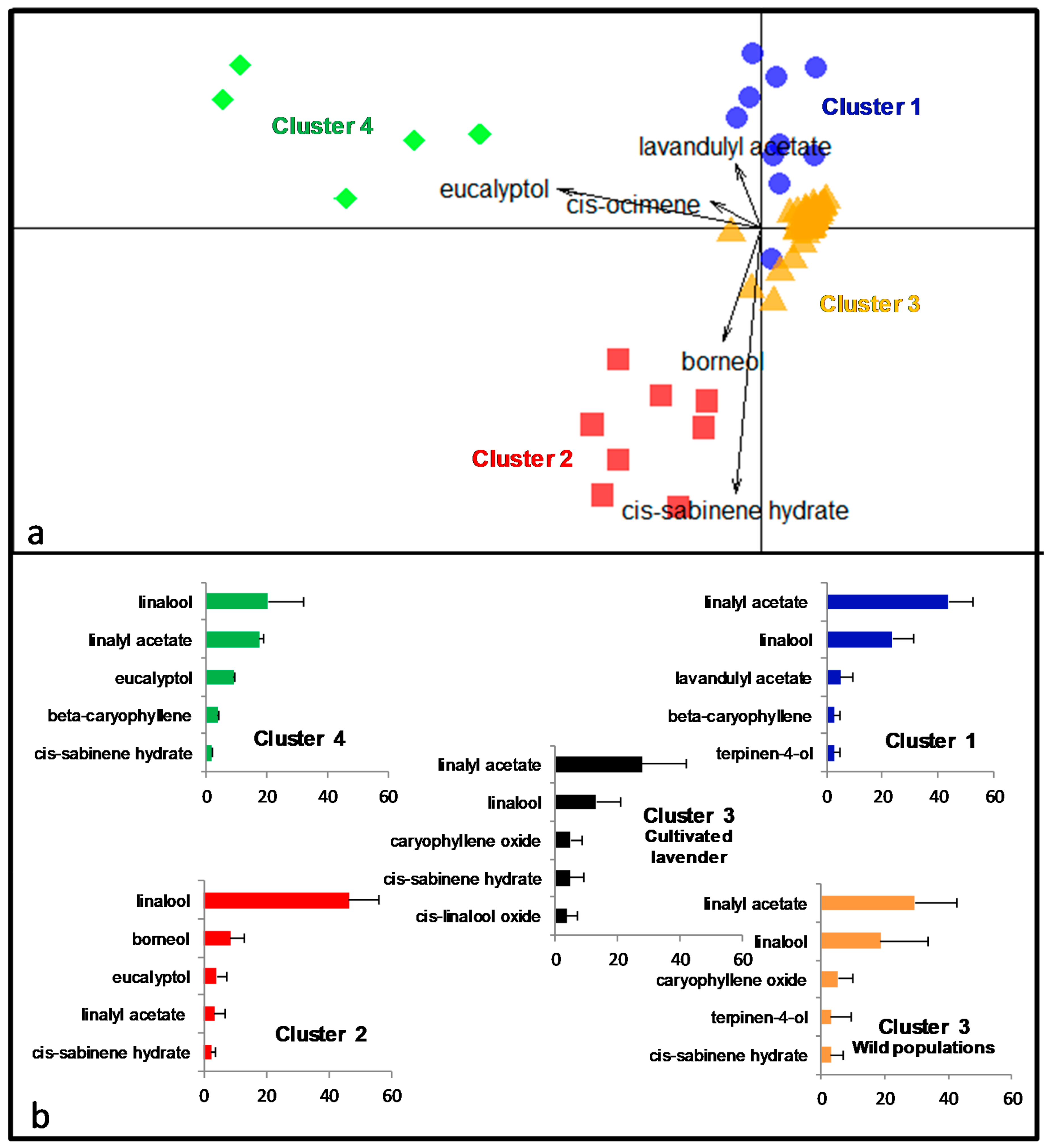
2.2. Unsupervised Identification of the Structure of Individuals
2.3. Composition of the Four Lavender Chemotypes
3. Discussion
4. Materials and Methods
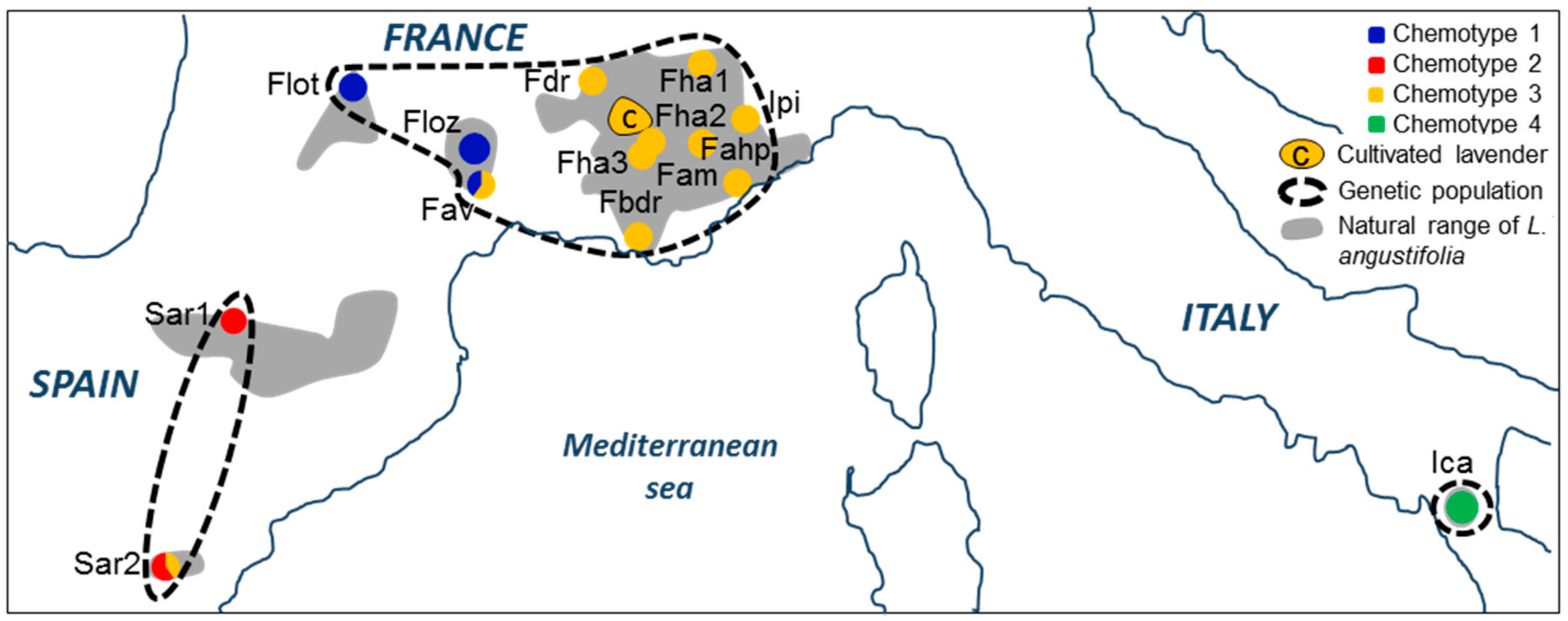
4.1. Plant Sampling
4.2. Genetic Fingerprinting
4.3. VOCs Extraction and Analysis
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Upson, T.; Andrews, S. The Genus Lavandula (A Botanical Magazine Monograph); Royal Botanic Gardens, Kew: Richmond, UK, 2004; ISBN 0-88192-642-6. [Google Scholar]
- Passalacqua, N.G.; Tundis, R.; Upson, T.M. A new species of Lavandula sect. Lavandula (Lamiaceae) and review of species boundaries in Lavandula angustifolia. Phytotaxa 2017, 292, 161–170. [Google Scholar] [CrossRef]
- Moja, S.; Guitton, Y.; Nicolè, F.; Legendre, L.; Pasquier, B.; Upson, T.; Jullien, F. Genome size and plastid trnK-matK markers give new insights into the evolutionary history of the genus Lavandula L. Plant Biosyst. 2016, 150, 1216–1224. [Google Scholar] [CrossRef]
- Cavanagh, H.M.A.; Wilkinson, J.M. Biological activities of Lavender essential oil. Phyther. Res. 2002, 16, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Stanev, S.; Zagorcheva, T.; Atanassov, I. Lavender cultivation in Bulgaria—21st century developments, breeding challenges and opportunities. Bulg. J. Agric. Sci. 2016, 22, 584–590. [Google Scholar]
- Guitton, Y.; Nicolè, F.; Moja, S.; Valot, N.; Legrand, S.; Jullien, F.; Legendre, L. Differential accumulation of volatile terpene and terpene synthase mRNAs during lavender (Lavandula angustifolia and L. x intermedia) inflorescence development. Physiol. Plant. 2010, 138, 150–163. [Google Scholar] [CrossRef]
- International Organization for Standardization. ISO 3515:2002/Cor 1:2004: Oil of French lavender (Lavandula angustifolia Miller). 2002. Available online: https://www.iso.org (accessed on 1 February 2004).
- Wells, R.; Truong, F.; Adal, A.M.; Sarker, L.S.; Mahmoud, S.S. Lavandula essential oils: A current review of applications in medicinal, food, and cosmetic industries of lavender. Nat. Prod. Commun. 2018, 13, 1403–1417. [Google Scholar] [CrossRef] [Green Version]
- Gaudin, J.; Semetey, O.; Foissac, X.; Eveillard, S. Phytoplasma titer in diseased lavender is not correlated to lavender tolerance to stolbur phytoplasma. Bull. Insectology 2011, 64, 179–180. [Google Scholar]
- Skoula, M.; Abidi, C.; Kokkalou, E. Essential oil variation of Lavandula stoechas L. ssp. stoechas growing wild in Crete (Greece). Biochem. Syst. Ecol. 1996, 24, 255–260. [Google Scholar] [CrossRef]
- Benabdelkader, T.; Zitouni, A.; Guitton, Y.; Jullien, F.; Maitre, D.; Casabianca, H.; Legendre, L.; Kameli, A. Essential oils from wild populations of algerian Lavandula stoechas L.: Composition, chemical variability, and in vitro biological properties. Chem. Biodivers. 2011. [Google Scholar] [CrossRef]
- Chograni, H.; Zaouali, Y.; Rajeb, C.; Boussaid, M. Essential oil variation among natural populations of Lavandula multifida L. (Lamiaceae). Chem. Biodivers. 2010, 7, 933–942. [Google Scholar] [CrossRef]
- Muñoz-Bertomeu, J.; Arrillaga, I.; Segura, J. Essential oil variation within and among natural populations of Lavandula latifolia and its relation to their ecological areas. Biochem. Syst. Ecol. 2007, 35, 479–488. [Google Scholar] [CrossRef]
- Behnke, K.; Ehlting, B.; Teuber, M.; Bauerfeind, M.; Louis, S.; Hänsch, R.; Polle, A.; Bohlmann, J.; Schnitzler, J.-P. Transgenic, non-isoprene emitting poplars don’t like it hot. Plant J. 2007, 51, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Vickers, C.E.; Possell, M.; Cojocariu, C.I.; Velikova, V.B.; Laothawornkitkul, J.; Ryan, A.; Mullineaux, P.M.; Nicholas Hewitt, C. Isoprene synthesis protects transgenic tobacco plants from oxidative stress. Plant Cell Environ. 2009, 32, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Laothawornkitkul, J.; Paul, N.D.; Vickers, C.E.; Possell, M.; Taylor, J.E.; Mullineaux, P.M.; Hewitt, C.N. Isoprene emissions influence herbivore feeding decisions. Plant Cell Environ. 2008, 31, 1410–1415. [Google Scholar] [CrossRef] [PubMed]
- Vernet, P.; Gouyon, R.H.; Valdeyron, G. Genetic control of the oil content in Thymus vulgaris L.: A case of polymorphism in a biosynthetic chain. Genetica 1986, 69, 227–231. [Google Scholar] [CrossRef]
- Thompson, J.D.; Chalchat, J.C.; Michet, A.; Linhart, Y.B.; Ehlers, B. Qualitative and quantitative variation in monoterpene co-occurrence and composition in the essential oil of Thymus vulgaris chemotypes. J. Chem. Ecol. 2003, 29, 859–880. [Google Scholar] [CrossRef]
- Moore, B.D.; Andrew, R.L.; Külheim, C.; Foley, W.J. Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 2014, 201, 733–750. [Google Scholar] [CrossRef]
- Jombart, T.; Collins, C. A tutorial for Discriminant Analysis of Principal Components (DAPC) using adegenet. Rvignette 2015. [Google Scholar] [CrossRef]
- Singh, A.; Shannon, C.; Gautier, B.; Rohart, F.; Vacher, M.; Tebbutt, S.; Lê Cao, K.-A. DIABLO: From multi-omics assays to biomarker discovery, an integrative approach. bioRxiv 2016, 067611. [Google Scholar] [CrossRef]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Hervé, M.R.; Nicolè, F.; Lê Cao, K.A. Multivariate analysis of multiple datasets: A practical guide for chemical ecology. J. Chem. Ecol. 2018, 44, 212–234. [Google Scholar]
- Herraiz-peñalver, D.; Ángeles, M.; Varela, F.; Navarrete, P.; Sánchez-vioque, R.; Usano-alemany, J. Chemical characterization of Lavandula latifolia Medik. essential oil from Spanish wild populations. Biochem. Syst. Ecol. 2013, 46, 59–68. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Conord, C.; Despinasse, Y.; Moja, S.; Rioux, D.; Till-Bottraud, I.; Jullien, F.; Nicolè, F. Menaces sur l’or bleu—Etude de la diversité génétique des lavandes fines sauvages et cultivées. In Proceedings of the Actes des Secondes Rencontres Végétales du Massif Central, Limoges, France, 10–14 October 2012; pp. 147–154. [Google Scholar]
- Moulin, N. Les cueilleuses “dans le sauvage”, femmes du Verdon et parfum de lavande. Garance Voyag. 2003, 61, 19–23. [Google Scholar]
- An, M.; Haig, T.; Hatfield, P. On-site field sampling and analysis of fragrance from living Lavender (Lavandula angustifolia L.) flowers by solid-phase microextraction coupled to gas chromatography and ion-trap mass spectrometry. J. Chromatogr. A 2001, 917, 245–250. [Google Scholar] [CrossRef]
- Shellie, R.; Mondello, L.; Marriott, P.; Dugo, G. Characterisation of lavender essential oils by using gas chromatography–mass spectrometry with correlation of linear retention indices and comparison with comprehensive two-dimensional gas chromatography. J. Chromatogr. A 2002, 970, 225–234. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Amiot, J.; Borron, C.; Linhart, Y.B.; Keeefover-Ring, K.; Gauthier, P. Spatial heterogeneity of gall formation in relation to chemotype distribution in Thymus vulgaris. Plant Ecol. 2019, 220, 777–788. [Google Scholar] [CrossRef]
- Vickers, C.E.; Gershenzon, J.; Lerdau, M.T.; Loreto, F. A unified mechanism of action for volatile isoprenoids in plant abiotic stress. Nat. Chem. Biol. 2009, 5, 283–291. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef]
- Baser, K.H.C.; Özek, T.; Konakchiev, A. Enantiomeric distribution of linalool, linalyl acetate and camphor in bulgarian lavender oil. J. Essent. Oil Res. 2005. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gauthier, P.; Amiot, J.; Ehlers, B.K.; Collin, C.; Fossat, J.; Barrios, V.; Arnaud-Miramont, F.; Keefover-Ring, K.E.N.; Linhart, Y.B. Ongoing adaptation to mediterranean climate extremes in a chemically polymorphic plant. Ecol. Monogr. 2007, 77, 421–439. [Google Scholar] [CrossRef] [Green Version]
- Mitra, J. Genetics and genetic improvement of drought resistance in crop plants. Curr. Sci. 2001, 80, 758–763. [Google Scholar]
- Rosati-Marzetti, C. Fiche d’Inventaire du Patrimoine Culturel Immatériel: Les pratiques de Culture des Lavandes dans les Alpes-de-Haute-Provence. Ministère Français de la culture. 2018. Available online: https://www.culture.gouv.fr/Media/Thematiques/Patrimoine-culturel-immateriel/Files/Fiches-inventaire-du-PCI/Les-pratiques-de-culture-des-lavandes-en-Haute-Provence (accessed on 12 December 2004).
- Pistelli, L.; Najar, B.; Giovanelli, S.; Lorenzini, L.; Tavarini, S.; Angelini, L.G. Agronomic and phytochemical evaluation of lavandin and lavender cultivars cultivated in the Tyrrhenian area of Tuscany (Italy). Ind. Crops Prod. 2017, 109, 37–44. [Google Scholar] [CrossRef]
- Nicolè, F.; Tellier, F.; Vivat, A.; Till-Bottraud, I. Conservation unit status inferred for plants by combining interspecific crosses and {AFLP}. Conserv. Genet. 2007, 8, 1273–1285. [Google Scholar] [CrossRef]
- Herrmann, D.; Poncet, B.N.; Manel, S.; Rioux, D.; Gielly, L.; Taberlet, P.; Gugerli, F. Selection criteria for scoring amplified fragment length polymorphisms (AFLPs) positively affect the reliability of population genetic parameter estimates. Genome 2010, 53, 302–310. [Google Scholar] [CrossRef]
- Nicolè, F.; Guitton, Y.; Courtois, E.A.; Moja, S.; Legendre, L.; Hossaert-Mckey, M. MSeasy: Unsupervised and untargeted GC-MS data processing. Bioinformatics 2012, 28, 2278–2280. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography–Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2017; ISBN 1-932633-21-9/978-1-932633-21-4. [Google Scholar]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Despinasse, Y.; Fiorucci, S.; Antonczak, S.; Moja, S.; Bony, A.; Nicolè, F.; Baudino, S.; Magnard, J.-L.; Jullien, F.; Despinasse, Y. Bornyl-diphosphate synthase from Lavandula angustifolia: A major monoterpene. Phytochemistry 2017, 137, 24–33. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J. Vegan: An Introduction to Ordination. Available online: http://cran.r-project.org/web/packages/vegan/vignettes/introvegan.pdf (accessed on 31 August 2004).
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NF ISO3515:2004 | |
|---|---|
| β-phellandrene | Max 0.5 |
| 3-octanone | Max 2 |
| camphor | Max 0.5 |
| limonene | Max 0.5 |
| eucalyptol | Max 1.0 |
| α-terpineol | Max 1.0 |
| terpinene-4-ol | 2–6 |
| Cis-ocimene | 4–10 |
| Trans-ocimene | 1.5–6.0 |
| linalool | 25–38 |
| linalyl acetate | 25–45 |
| lavandulol | Min 0.3 |
| lavandulyl acetate | Min 2.0 |
| Site | N | Long (DD) | Lat (DD) | Altitude (m) | % Total Area | % Mono | % Sesqui | Total Amount | Number of Compounds |
|---|---|---|---|---|---|---|---|---|---|
| Fahp | 5 | 6.6 | 44.17 | 1400 | 83.4 ± 10.9 | 73.2 ± 14.1 | 6.9 ± 3 | 1.4 ± 0.7 | 29.6 ± 5.2 |
| Fam | 3 | 7.08 | 43.76 | 772–800 | 59.0 ± 18.5 | 48.4 ± 17 | 9.0 ± 3.1 | 0.8 ± 0.3 | 10.7 ± 2.3 |
| Fav | 5 | 5.45 | 43.9 | 450–600 | 92.0 ± 3.8 | 78.4 ± 8.5 | 12.1 ± 6 | 4.5 ± 4.7 | 32.0 ± 9.8 |
| Fbdr | 3 | 5.66 | 43.32 | 725–800 | 71.0 ± 4.1 | 53.3 ± 9.6 | 17.0 ± 13.2 | 2.4 ± 1.1 | 22.3 ± 3.1 |
| Fdr | 5 | 5.09 | 44.85 | 590–600 | 72.4 ± 9.9 | 65.6 ± 9.4 | 5.0 ± 2.8 | 1.3 ± 1 | 17.6 ± 5.2 |
| Fha1 | 4 | 6.69 | 44.93 | 1450–1600 | 77.8 ± 16.4 | 70.3 ± 18.9 | 6.0 ± 3 | 2.3 ± 1.9 | 21.8 ± 6 |
| Fha2 | 5 | 5.94 | 44.06 | 755–800 | 92.7 ± 3.1 | 78.5 ± 7.1 | 11.9 ± 3.5 | 2.3 ± 1.3 | 29.0 ± 7.7 |
| Fha3 | 5 | 5.85 | 44.12 | 1635–1800 | 80.9 ± 7.1 | 67.7 ± 9.8 | 10.8 ± 3.6 | 1.2 ± 0.6 | 27.2 ± 7.8 |
| Flot | 5 | 1.53 | 44.95 | 250–400 | 88.7 ± 7.3 | 77.6 ± 9.1 | 6.8 ± 2.4 | 7.4 ± 3.5 | 41.0 ± 7.6 |
| Floz | 5 | 3.37 | 44.22 | 980–1000 | 92.0 ± 7.6 | 85.8 ± 9 | 4.1 ± 1 | 16.0 ± 8.1 | 39.0 ± 7.4 |
| Ica | 5 | 16.05 | 39.9 | 1150–1200 | 66.7 ± 12.3 | 58.0 ± 11.9 | 8.2 ± 1.1 | 35.2 ± 9.2 | 55.2 ± 0.8 |
| Ipi | 5 | 7.23 | 44.34 | 1275–1400 | 74.5 ± 10.1 | 66.3 ± 8.6 | 4.8 ± 1.6 | 1.1 ± 0.4 | 13.4 ± 1.5 |
| Sar1 | 5 | 0.21 | 42.57 | 1000 | 88.5 ± 2.4 | 79.7 ± 4.3 | 7.5 ± 3 | 12.8 ± 3.9 | 48.4 ± 2.9 |
| Sar2 | 5 | −0.55 | 40.51 | 1450–1600 | 84.4 ± 2.7 | 76.8 ± 3.9 | 5.1 ± 1.4 | 5.4 ± 2 | 43.0 ± 2.5 |
| Cult | 25 | - | - | - | 83.0 ± 9.1 | 70.0 ± 12.1 | 10.7 ± 4.8 | 1.1 ± 0.6 | 29.1 ± 7.9 |
| Source of Variation | df | Sum of Squares | Estimated Variance | Percentage of Variance | PHI-Statistics | p |
|---|---|---|---|---|---|---|
| Between genetic clusters | 2 | 440.3 | 14.83 | 53 | PHIRT = 0.535 | <0.001 |
| Among pop within clusters | 16 | 422.3 | 4.18 | 15 | PHIPR = 0.324 | <0.001 |
| Within populations | 63 | 549.9 | 8.73 | 31 | PHIPT = 0.685 | <0.001 |
| French Genetic Cluster | Calabrese Genetic Cluster | Spanish Genetic Cluster | |
|---|---|---|---|
| French genetic cluster | - | 0.001 | 0.001 |
| Calabrese genetic cluster | 0.557 | - | 0.001 |
| Spanish genetic cluster | 0.552 | 0.752 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Despinasse, Y.; Moja, S.; Soler, C.; Jullien, F.; Pasquier, B.; Bessière, J.-M.; Baudino, S.; Nicolè, F. Structure of the Chemical and Genetic Diversity of the True Lavender over Its Natural Range. Plants 2020, 9, 1640. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9121640
Despinasse Y, Moja S, Soler C, Jullien F, Pasquier B, Bessière J-M, Baudino S, Nicolè F. Structure of the Chemical and Genetic Diversity of the True Lavender over Its Natural Range. Plants. 2020; 9(12):1640. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9121640
Chicago/Turabian StyleDespinasse, Yolande, Sandrine Moja, Catherine Soler, Frédéric Jullien, Bernard Pasquier, Jean-Marie Bessière, Sylvie Baudino, and Florence Nicolè. 2020. "Structure of the Chemical and Genetic Diversity of the True Lavender over Its Natural Range" Plants 9, no. 12: 1640. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9121640