Sequential Application of Antioxidants Rectifies Ion Imbalance and Strengthens Antioxidant Systems in Salt-Stressed Cucumber

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Measurements
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rady, M.O.A.; Semida, W.M.; El-Mageed, T.A.A.; Hemida, K.A.; Rady, M.M. Up-regulation of antioxidative defense systems by glycine betaine foliar application in onion plants confer tolerance to salinity stress. Sci. Hortic. 2018, 240, 614–622. [Google Scholar] [CrossRef]
- Kumar, N.; Pal, M.; Singh, A.; SaiRam, R.K.; Srivastava, G.C. Exogenous proline alleviates oxidative stress and increase vase life in rose (Rosa hybrida L. ‘Grand Gala’). Sci. Hortic. 2010, 127, 79–85. [Google Scholar] [CrossRef]
- Jones, R.W., Jr.; Pike, L.M.; Yourman, L.F. Salinty influences cucumber growth and yield. J. Am. Soc. Hortic. Sci. 1989, 114, 547–551. [Google Scholar]
- Abu-zinada, I.A. Effect of salinity levels and application stage on cucumber and soil under greenhouse condition. Int. J. Agric. Crop Sci. 2015, 8, 73–80. [Google Scholar]
- Furtana, G.B.; Tipirdamaz, R. Physiological and antioxidant response of three cultivars of cucumber (Cucumis sativus L.) to salinity. Turk. J. Biol. 2010, 34, 287–296. [Google Scholar] [CrossRef]
- Peykanpour, E.; AM, G.; Fallahzade, J.; Najarian, M. Interactive effects of salinity and ozonated water on yield components of cucumber. Plant Soil Environ. 2016, 62, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Wan, S.; Kang, Y.; Wang, D.; Liu, S. Effect of saline water on cucumber (Cucumis sativus L.) yield and water use under drip irrigation in North China. Agric. Water Manag. 2010, 98, 105–113. [Google Scholar] [CrossRef]
- Wu, Y.; Jin, X.; Liao, W.; Hu, L.; Dawuda, M.M.; Zhao, X. 5-Aminolevulinic acid (ALA) alleviated salinity stress in cucumber seedlings by enhancing chlorophyll synthesis pathway. Front. Plant Sci. 2018, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, B.; Yasar, F.; Özpay, T.; Türközü, D.; Terziodlu, Ö.; Tamkoç, A. Variations in response to salt stress among field pea genotypes (Pisum sativum sp.arvense L.). J. Anim. Vet. Adv. 2008, 7, 907–910. [Google Scholar]
- Seleiman, M.F.; Kheir, A.S. Saline soil properties, quality and productivity of wheat grown with bagasse ash and thiourea in different climatic zones. Chemosphere 2018, 193, 538–546. [Google Scholar] [CrossRef]
- Seleiman, M.F. Use of plant nutrients in improving abiotic stress tolerance in wheat. In Wheat Production in Changing Environments: Management, Adaptation and Tolerance; Hasanuzzaman, M., Nahar, K., Hossain, A., Eds.; Springer Nature: Singapore, 2019; pp. 481–495. [Google Scholar]
- Seleiman, M.F.; Kheir, A.M.S.; Al-Dhumri, S.; Alghamdi, A.G.; Omar, E.H.; Aboelsoud, H.M.; Abdella, K.A.; AbouElHassan, W.H. Exploring optimal tillage improved soil characteristics and productivity of wheat irrigated with different water qualities. Agronomy 2019, 9, 233. [Google Scholar] [CrossRef] [Green Version]
- Al-Ashkar, I.; Alderfasi, A.; Ben Romdhane, W.; Seleiman, M.F.; El-Said, R.A.; Al-Doss, A. Morphological and genetic diversity within salt tolerance detection in eighteen wheat genotypes. Plants 2020, 9, 287. [Google Scholar] [CrossRef] [Green Version]
- Taha, R.S.; Seleiman, M.F.; Alotaibi, M.; Alhammad, B.A.; Rady, M.M.; Mahdi, A.H.A. Exogenous potassium treatments elevate salt tolerance and performances of glycine max L. by boosting antioxidant defense system under actual saline field conditions. Agronomy 2020, 10, 1741. [Google Scholar] [CrossRef]
- Ding, Z.; Kheir, A.S.; Ali, O.A.; Hafez, E.; Elshamey, E.A.; Zhou, Z.; Wang, B.; Lin, X.; Ge, Y.; Fahmy, A.F.; et al. Vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manag. 2020, 277, 111388. [Google Scholar] [CrossRef]
- Bargaz, A.; Nassar, R.M.A.; Rady, M.M.; Gaballah, M.S.; Thompson, S.M.; Brestic, M.; Schmidhalter, U.; Abdelhamid, M.T. Improved salinity tolerance by phosphorus fertilizer in two phaseolus vulgaris recombinant inbred lines contrasting in their P-efficiency. J. Agron. Crop Sci. 2016. [Google Scholar] [CrossRef]
- Semida, W.M.; Abd El-Mageed, T.A.; Hemida, K.; Rady, M.M. Natural bee-honey based biostimulants confer salt tolerance in onion via modulation of the antioxidant defence system. J. Hortic. Sci. Biotechnol. 2019, 94, 632–642. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Semida, W.M.; Mohamed, G.F.; Rady, M.M. Combined effect of foliar-applied salicylic acid and deficit irrigation on physiological—Anatomical responses, and yield of squash plants under saline soil. S. Afr. J. Bot. 2016, 106, 8–16. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 113, 59–89. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.M.; Taha, S.S.; Kusvuran, S. Integrative application of cyanobacteria and antioxidants improves common bean performance under saline conditions. Sci. Hortic. 2018, 62, 361–366. [Google Scholar] [CrossRef]
- Zhu, J.; Bie, Z.; Li, Y. Physiological and growth responses of two different salt-sensitive cucumber cultivars to NaCl stress. Soil Sci. Plant Nutr. 2008, 54, 400–407. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Semida, W.M.; Hemida, K.A.; Rady, M.M. Sequenced ascorbate-proline-glutathione seed treatment elevates cadmium tolerance in cucumber transplants. Ecotoxicol. Environ. Saf. 2018, 154, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Desoky, E.M.; El-maghraby, L.M.M.; Awad, A.E.; Abdo, A.I.; Rady, M.M.; Semida, W.M. Fennel and ammi seed extracts modulate antioxidant defence system and alleviate salinity stress in cowpea (Vigna unguiculata). Sci. Hortic. 2020, 272, 109576. [Google Scholar] [CrossRef]
- Ejaz, B.; Sajid, Z.A.; Aftab, F. Effect of exogenous application of ascorbic acid on antioxidant enzyme activities, proline contents, and growth parameters of Saccharum spp. hybrid cv. HSF-240 under salt stress. Turk. J. Biol. 2012, 36, 630–640. [Google Scholar] [CrossRef]
- Rady, M.M.; Taha, R.S.; Semida, W.M.; Alharby, H.F. Modulation of salt stress effects on vicia faba l. plants grown on a reclaimed-saline soil by salicylic acid application. Rom. Agric. Res. 2017, 34, 175–185. [Google Scholar]
- Rady, M.M.; Hemida, K.A. Sequenced application of ascorbate-proline-glutathione improves salt tolerance in maize seedlings. Ecotoxicol. Environ. Saf. 2016, 133, 252–259. [Google Scholar] [CrossRef]
- Rady, M.M.; Hemida, K.A. Modulation of cadmium toxicity and enhancing cadmium-tolerance in wheat seedlings by exogenous application of polyamines. Ecotoxicol. Environ. Saf. 2015, 119, 178–185. [Google Scholar] [CrossRef]
- Semida, W.M.; El-Mageed, T.A.A.; Mohamed, S.E.; El-Sawah, N.A. Combined effect of deficit irrigation and foliar-applied salicylic acid on physiological responses, yield, and water-use efficiency of onion plants in saline calcareous soil. Arch. Agron. Soil Sci. 2017, 63, 1227–1239. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Rady, M.M. Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldeen, R.P.; Teare, I.D. Rapid determination of free proline for water—Stress studies. Plant Soil 1973, 207, 205–207. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Vanmontagu, M.; Inze, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Rao, M.V.; Paliyath, G.; Ormrod, P. Ultraviolet-9- and ozone-lnduced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Williams, V.; Twine, S. Flame photometric method for sodium, potassium and calcium. In Modern Methods of Plant Analysis; Springer: Berlin/Heidelberg, Germany, 1960; pp. 3–5. [Google Scholar]
- Chapman, H.D.; Pratt, P.F. Methods of Analysis for Soil, Plants and Water; University of California, Division of Agricultural Science: Berkeley, CA, USA, 1961. [Google Scholar]
- Mousavi, S.A.R.; Chauvin, A.; Pascaud, F.; Kellenberger, S.; Farmer, E.E. GLUTAMATE RECEPTOR-LIKE genes mediate leaf-to-leaf wound signalling. Nature 2013, 500, 422. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Aliniaeifard, S.; Hajilou, J.; Tabatabaei, S.J.; Sifi-Kalhor, M. Effects of ascorbic acid and reduced glutathione on the alleviation of salinity stress in olive plants. Int. J. Fruit Sci. 2016, 16, 395–409. [Google Scholar] [CrossRef]
- Rady, M.M.; Semida, W.M.; Hemida, K.A.; Abdelhamid, M.T. The effect of compost on growth and yield of Phaseolus vulgaris plants grown under saline soil. Int. J. Recycl. Org. Waste Agric. 2016, 5, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Shalata, A.; Neumann, P.M. Exogenous ascorbic acid (vitamin C) increases resistance to salt stress and reduces lipid peroxidation. J. Exp. Bot. 2001, 52, 2207–2211. [Google Scholar] [CrossRef] [PubMed]
- Wutipraditkul, N.; Wongwean, P.; Buaboocha, T. Alleviation of salt-induced oxidative stress in rice seedlings by proline and/or glycinebetaine. Biol. Plant. 2015, 59, 547–553. [Google Scholar] [CrossRef]
- Conklin, P.L. Recent advances in the role and biosynthesis of ascorbic acid in plants. Plant Cell Environ. 2001, 24, 383–394. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between Stress perception and physiological responses. Plant Cell Online 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Scandalios, J.G.; Guan, L.; Polidoros, A.N. Catalases in plants: Gene structure, properties, regulation, and expression. Cold Spring Harb. Monogr. Ser. 1997, 34, 343–406. [Google Scholar]
- Reddy, K.R.; Kakani, V.G.; Zhao, D.; Koti, S.; Gao, W. Interactive effects of ultraviolet-B radiation and temperature on cotton physiology, growth, development and hyperspectral reflectance. Photochem. Photobiol. 2004, 79, 416–427. [Google Scholar] [CrossRef]
- Kanokwan, S.; Tanatorn, S.; Aphichart, K. Effect of salinity stress on antioxidative enzyme activities in tomato cultured in vitro. Pak. J. Bot. 2015, 47, 1–10. [Google Scholar]
- Rezende, R.A.L.S.; Rodrigues, F.A.; Soares, J.D.R.; Silveira, H.R.O.; Pasqual, M.; Dias, G.M.G. Salt stress and exogenous silicon influence physiological and anatomical features of in vitro-grown cape gooseberry. Cienc. Rural 2018, 48. [Google Scholar] [CrossRef] [Green Version]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of arbuscular mycorrhizae on photosynthesis and water status of maize plants under salt stress. Mycorrhiza 2008, 18, 287–296. [Google Scholar] [CrossRef]
- Ranjbarfordoei, A.; Samson, R.; Van Damme, P. Chlorophyll fluorescence performance of sweet almond [Prunus dulcis (Miller) D. Webb] in response to salinity stress induced by NaCl. Photosynthetica 2006, 44, 513–522. [Google Scholar] [CrossRef]
- Younis, M.E.; Mohamed, S.; Tourky, N. Influence of salinity and adaptive compounds on growth parameters, carbohydrates, amino EISSN: Acids and nucleic acids in two cultivars of Vicia faba contrasting in salt tolerance. Plant Knowl. J. 2014, 3, 47–56. [Google Scholar]
- Dolatabadian, A.; Jouneghani, R.S. Impact of exogenous ascorbic acid on antioxidant activity and some physiological traits of common bean subjected to salinity stress. Not. Bot. Horti Agrobot. Cluj-Napoca 2009, 37, 165–172. [Google Scholar] [CrossRef]
- Valizadeh, M.; Asgari, A.; Shiri, M. Proline, glycine betaine, total phenolics and pigment contents in response to osmotic stress in maize seedlings. J. BioSci. Biotechnol. 2015, 4, 313–319. [Google Scholar]
- Darvishan, M.; Tohidi-moghadam, H.R.; Zahedi, H. The effects of foliar application of ascorbic acid (vitamin C) on physiological and biochemical changes of corn (Zea mays L) under irrigation withholding in different growth stages. Maydica 2013, 58, 195–200. [Google Scholar]
- Slabbert, M.M.; Krüger, G.H.J. Antioxidant enzyme activity, proline accumulation, leaf area and cell membrane stability in water stressed Amaranthus leaves. S. Afr. J. Bot. 2014, 95, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.M.; Abd El-Mageed, T.A.; Abdelhamid, M.T.; Abd El-Azeem, M.M.M. Integrative potassium humate and biochar application reduces salinity effects and contaminants and improves growth and yield of eggplant grown under saline conditions. Int. J. Empir. Educ. Res. 2018, 1, 37–56. [Google Scholar] [CrossRef]
- Valero, E.; Macià, H.; De la Fuente, I.M.; Hernández, J.A.; González-Sánchez, M.I.; García-Carmona, F. Modeling the ascorbate-glutathione cycle in chloroplasts under light/dark conditions. BMC Syst. Biol. 2016, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Sairam, R.K.; Tyagi, A. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci. 2004, 86, 407–421. [Google Scholar]
- Rady, M.M.; Kuşvuran, A.; Alharby, H.F.; Alzahrani, Y.; Kuşvuran, S. Pretreatment with proline or an organic bio-stimulant induces salt tolerance in wheat plants by improving antioxidant redox state and enzymatic activities and reducing the oxidative stress. J. Plant Growth Regul. 2019, 38, 449–462. [Google Scholar] [CrossRef]
- Miller, G.; Honig, A.; Stein, H.; Suzuki, N.; Mittler, R.; Zilberstein, A. Unraveling Δ1-pyrroline-5-carboxylate-proline cycle in plants by uncoupled expression of proline oxidation enzymes. J. Biol. Chem. 2009, 84, 19–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semida, W.M.; Rady, M.M. Pre-soaking in 24-epibrassinolide or salicylic acid improves seed germination, seedling growth, and anti-oxidant capacity in Phaseolus vulgaris L. grown under NaCl stress. J. Hortic. Sci. Biotechnol. 2014, 89, 338–344. [Google Scholar] [CrossRef]
- Zhou, Y.; Wen, Z.; Zhang, J.; Chen, X.; Cui, J.; Xu, W.; Liu, H.-y. Exogenous glutathione alleviates salt-induced oxidative stress in tomato seedlings by regulating glutathione metabolism, redox status, and the antioxidant system. Sci. Hortic. 2017, 220, 90–101. [Google Scholar] [CrossRef]
- Desoky, E.S.M.; Elrys, A.S.; Rady, M.M. Integrative moringa and licorice extracts application improves Capsicum annuum fruit yield and declines its contaminant contents on a heavy metals-contaminated saline soil. Ecotoxicol. Environ. Saf. 2019, 169, 50–60. [Google Scholar] [CrossRef]
- Abogadallah, G.M. Antioxidative defense under salt stress. Plant Signal. Behav. 2010, 5, 369–374. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Lim, J.-H.; Park, M.-R.; Kim, Y.-J.; Park, T.-I.; Seo, Y.-W.; Choi, K.-G.; Yun, S.-J. Enhanced antioxidant enzymes are associated with reduced hydrogen peroxide in barley roots under saline stress. BMB Rep. 2005, 38, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Desoky, E.M.; Merwad, A.M.; Semida, W.M.; Ibrahim, S.A.; El-saadony, M.T.; Rady, M.M. Heavy metals-resistant bacteria (HM-RB): Potential bioremediators of heavy metals-stressed Spinacia oleracea plant. Ecotoxicol. Environ. Saf. 2020, 198, 110685. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The Role of Na+ and K+ Transporters in Salt Stress Adaptation in Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Abd El-mageed, T.A.; Abd El-mageed, S.A.; Semida, W.M.; Rady, M.O.A. Silicon Defensive Role in Maize (Zea mays L.) against Drought Stress and Metals-Contaminated Irrigation Water. Silicon 2020. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Gaxiola, R.A.; Rao, R.; Sherman, A.; Grisafi, P.; Alper, S.L.; Fink, G.R. The Arabidopsis thaliana proton transporters, AtNhx1 and Avp1, can function in cation detoxification in yeast. Proc. Natl. Acad. Sci. USA 1999, 96, 1480–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Yan, J.; Shen, G.; Fu, L.; Holaday, A.S.; Auld, D.; Blumwald, E.; Zhang, H. Expression of an Arabidopsis vacuolar sodium/proton antiporter gene in cotton improves photosynthetic performance under salt conditions and increases fiber yield in the field. Plant Cell Physiol. 2005, 46, 1848–1854. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Mao, B.; Ou, S.; Wang, W.; Liu, L.; Wu, Y.; Chu, C.; Wang, X. OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant Mol. Biol. 2014, 84, 19–36. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatments | Shoot Length (cm) | Leaf Area (cm2) | Stem Diameter (mm) | Shoot FW (g) | Shoot DW (g) |
|---|---|---|---|---|---|
| Control | 15.6 ± 0.4 cd | 64.0 ± 2.2 cd | 3.6 ± 0.1 b | 1.71 ± 0.07 b | 0.13 ± 0.02 d |
| AsA | 17.0 ± 0.6 abc | 70.6 ± 5.8 ab | 3.8 ± 0.1 ab | 2.00 ± 0.16 a | 0.15 ± 0.03 bc |
| Pro | 16.6 ± 0.7 bcd | 69.3 ± 3.6 bc | 3.7 ± 0.1 b | 2.03 ± 0.08 a | 0.15 ± 0.04 bc |
| GSH | 17.9 ± 0.7 ab | 70.8 ± 3.3 ab | 3.8 ± 0.1 ab | 2.02 ± 0.08 a | 0.15 ± 0.02 bc |
| AsA-Pro-GSH | 18.3 ± 0.6 a | 75.5 ± 5.0 a | 4.0 ± 0.1 a | 2.03 ± 0.14 a | 0.17 ± 0.04 a |
| NaCl | 11.8 ± 0.6 e | 47.2 ± 2.0 e | 3.0 ± 0.1 c | 1.41 ± 0.05 c | 0.10 ± 0.02 e |
| NaCl + AsA | 16.8 ± 0.3 abc | 63.4 ± 4.2 d | 3.6 ± 0.1 b | 1.95 ± 0.12 a | 0.14 ± 0.02 cd |
| NaCl + Pro | 15.4 ± 0.6 cd | 61.1 ± 2.3 d | 3.6 ± 0.1 b | 1.98 ± 0.06 a | 0.13 ± 0.02 d |
| NaCl + GSH | 15.1 ± 0.5 d | 64.1 ± 3.6 cd | 3.6 ± 0.1 b | 1.98 ± 0.09 a | 0.14 ± 0.03 cd |
| NaCl + AsA-Pro-GSH | 17.9 ± 0.4 ab | 70.7 ± 2.5 ab | 3.7 ± 0.1 b | 2.00 ± 0.07 a | 0.16 ± 0.04 ab |
| Treatments | Stomatal Conductance (mmol−2 S−1) | SPAD Chlorophyll | Fv/Fm | Performance Index (%) |
|---|---|---|---|---|
| Control | 193 ± 5 d | 42.1 ± 2.2 ab | 0.83 ± 0.06 a | 3.64 ± 0.24 ab |
| AsA | 241 ± 7 abc | 42.4 ± 1.0 ab | 0.83 ± 0.05 a | 3.75 ± 0.10 ab |
| Pro | 247 ± 5 abc | 42.8 ± 0.6 ab | 0.83 ± 0.05 a | 3.73 ± 0.12 ab |
| GSH | 254 ± 6 a | 42.9 ± 0.9 ab | 0.83 ± 0.04 a | 3.78 ± 0.14 a |
| AsA-Pro-GSH | 257 ± 8 a | 44.7 ± 1.4 a | 0.84 ± 0.05 a | 3.78 ± 0.12 a |
| NaCl | 142 ± 4 e | 33.8 ± 2.0 e | 0.76 ± 0.04 b | 3.03 ± 0.35 c |
| NaCl + AsA | 232 ± 8 c | 38.9 ± 1.8 cd | 0.82 ± 0.05 a | 3.46 ± 0.30 b |
| NaCl + Pro | 234 ± 5 bc | 38.6 ± 1.9 d | 0.82 ± 0.04 a | 3.46 ± 0.29 b |
| NaCl + GSH | 234 ± 7 bc | 39.0 ± 0.8 cd | 0.82 ± 0.04 a | 3.48 ± 0.31 b |
| NaCl + AsA-Pro-GSH | 252 ± 6 ab | 41.4 ± 1.5 bc | 0.84 ± 0.05 a | 3.72 ± 0.19 ab |
| Treatments | RWC (%) | MSI (%) | Free Proline (mg g−1 DW) | AsA (μmol Ascorbate g−1 DW) | GSH (nmol GSH g-1 DW) |
|---|---|---|---|---|---|
| Control | 87.6 ± 0.7 abc | 76.3 ± 1.1 a | 0.27 ± 0.00 f | 1.48 ± 0.04 g | 0.14 ± 0.01 g |
| AsA | 88.3 ± 1.0 abc | 78.5 ± 0.4 a | 0.31 ± 0.01 ef | 3.54 ± 0.08 d | 0.19 ± 0.01 f |
| Pro | 89.2 ± 0.8 a | 78.3 ± 0.5 a | 0.65 ± 0.04 c | 1.85 ± 0.02 f | 0.20 ± 0.00 f |
| GSH | 88.7 ± 2.1 ab | 79.0 ± 1.1 a | 0.32 ± 0.02 e | 1.97 ± 0.04 f | 0.35 ± 0.01 d |
| AsA-Pro-GSH | 91.3 ± 2.6 a | 79.7 ± 0.8 a | 0.76 ± 0.00 b | 4.27 ± 0.08 c | 0.40 ± 0.00 c |
| NaCl | 74.5 ± 0.4 d | 63.6 ± 2.1 c | 0.33 ± 0.00 e | 2.89 ± 0.08 e | 0.18 ± 0.01 fg |
| NaCl + AsA | 82.7 ± 0.9 c | 69.8 ± 0.7 b | 0.40 ± 0.02 d | 6.32 ± 0.05 b | 0.25 ± 0.00 e |
| NaCl + Pro | 83.2 ± 2.7 bc | 69.9 ± 1.5 b | 0.80 ± 0.01 b | 3.34 ± 0.03 d | 0.28 ± 0.01 e |
| NaCl + GSH | 82.8 ± 1.2 bc | 70.4 ± 0.8 b | 0.41 ± 0.01 d | 3.40 ± 0.04 d | 0.49 ± 0.01 b |
| NaCl + AsA-Pro-GSH | 87.8 ± 1.4 abc | 76.8 ± 1.0 a | 0.87 ± 0.01 a | 7.58 ± 0.13 a | 0.59 ± 0.01 a |
| Treatments | Superoxide Dismutase | Catalase | Glutathione Reductase | Ascorbate Peroxidase |
|---|---|---|---|---|
| μmol mg‒1 Protein min‒1 | ||||
| Control | 0.23 ± 0.01 e | 0.23 ± 0.00 d | 0.27 ± 0.01 d | 0.30 ± 0.01 e |
| AsA | 0.29 ± 0.01 cd | 0.28 ± 0.01 c | 0.32 ± 0.01 c | 0.43 ± 0.02 c |
| Pro | 0.28 ± 0.01 d | 0.30 ± 0.02 c | 0.34 ± 0.02 c | 0.42 ± 0.01 c |
| GSH | 0.28 ± 0.01 d | 0.28 ± 0.01 c | 0.34 ± 0.02 c | 0.42 ± 0.02 c |
| AsA-Pro-GSH | 0.31 ± 0.02 bcd | 0.35 ± 0.00 b | 0.41 ± 0.02 b | 0.52 ± 0.02 b |
| NaCl | 0.25 ± 0.01 e | 0.29 ± 0.00 c | 0.32 ± 0.01 c | 0.36 ± 0.01 d |
| NaCl + AsA | 0.31 ± 0.01 bcd | 0.35 ± 0.01 b | 0.39 ± 0.02 b | 0.49 ± 0.02 b |
| NaCl + Pro | 0.31 ± 0.02 bcd | 0.37 ± 0.00 b | 0.39 ± 0.01 b | 0.50 ± 0.02 b |
| NaCl + GSH | 0.33 ± 0.02 b | 0.37 ± 0.01 b | 0.40 ± 0.02 b | 0.50 ± 0.02 b |
| NaCl + AsA-Pro-GSH | 0.38 ± 0.02 a | 0.41 ± 0.02 a | 0.47 ± 0.02 a | 0.55 ± 0.02 a |
| Parameters | Control (N) | Treatments | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N+ AsA | N+ Pro | N+ GSH | N+ A.P.G | NaCl (S) | S+ AsA | S+ Pro | S+ GSH | S+ A.P.G | ||
| Shoot length | 15.6 | 9.0 | 6.4 | 15 | 17 | −24 | 7.7 | −1.3 | −3.2 | 15 |
| Leaf area | 64.0 | 10 | 8.3 | 11 | 18 | −26 | −0.9 | −4.5 | 0.2 | 10 |
| Stem diameter | 3.6 | 5.6 | 2.8 | 5.6 | 11 | −17 | 0.0 | 0.0 | 0.0 | 2.8 |
| Shoot FW | 1.71 | 17 | 19 | 18 | 19 | −18 | 14 | 16 | 16 | 19 |
| Shoot DW | 0.13 | 15 | 15 | 15 | 31 | −23 | 7.7 | 0.0 | 7.7 | 23 |
| gs | 193 | 25 | 28 | 32 | 33 | −26 | 20 | 21 | 21 | 31 |
| SPAD value | 42.1 | 0.7 | 1.7 | 1.9 | 6.2 | −20 | −7.6 | −8.3 | -7.4 | -1.7 |
| Fv/Fm | 0.83 | 0.0 | 0.0 | 0.0 | 1.2 | −8.4 | −1.2 | −1.2 | −1.2 | 1.2 |
| PI | 3.64 | 3.0 | 2.5 | 3.8 | 3.8 | −17 | −4.9 | -4.9 | −4.4 | 2.2 |
| RWC | 87.6 | 0.8 | 1.8 | 1.3 | 4.2 | −15 | −5.6 | −5.0 | −5.5 | 0.2 |
| MSI | 76.3 | 2.9 | 2.6 | 3.5 | 4.5 | −17 | −8.5 | −8.4 | −7.7 | 0.7 |
| Free proline | 0.27 | 15 | 141 | 19 | 181 | 22 | 48 | 196 | 52 | 222 |
| Ascorbate | 1.48 | 139 | 25 | 33 | 189 | 95 | 327 | 126 | 130 | 412 |
| Glutathione | 0.14 | 36 | 43 | 150 | 186 | 29 | 79 | 100 | 250 | 321 |
| SOD activity | 0.23 | 26 | 22 | 22 | 35 | 8.7 | 35 | 35 | 43 | 65 |
| CAT activity | 0.23 | 22 | 30 | 22 | 52 | 26 | 52 | 61 | 61 | 78 |
| GR activity | 0.27 | 19 | 26 | 26 | 52 | 19 | 44 | 44 | 48 | 74 |
| APX activity | 0.30 | 43 | 40 | 40 | 73 | 20 | 63 | 67 | 67 | 83 |
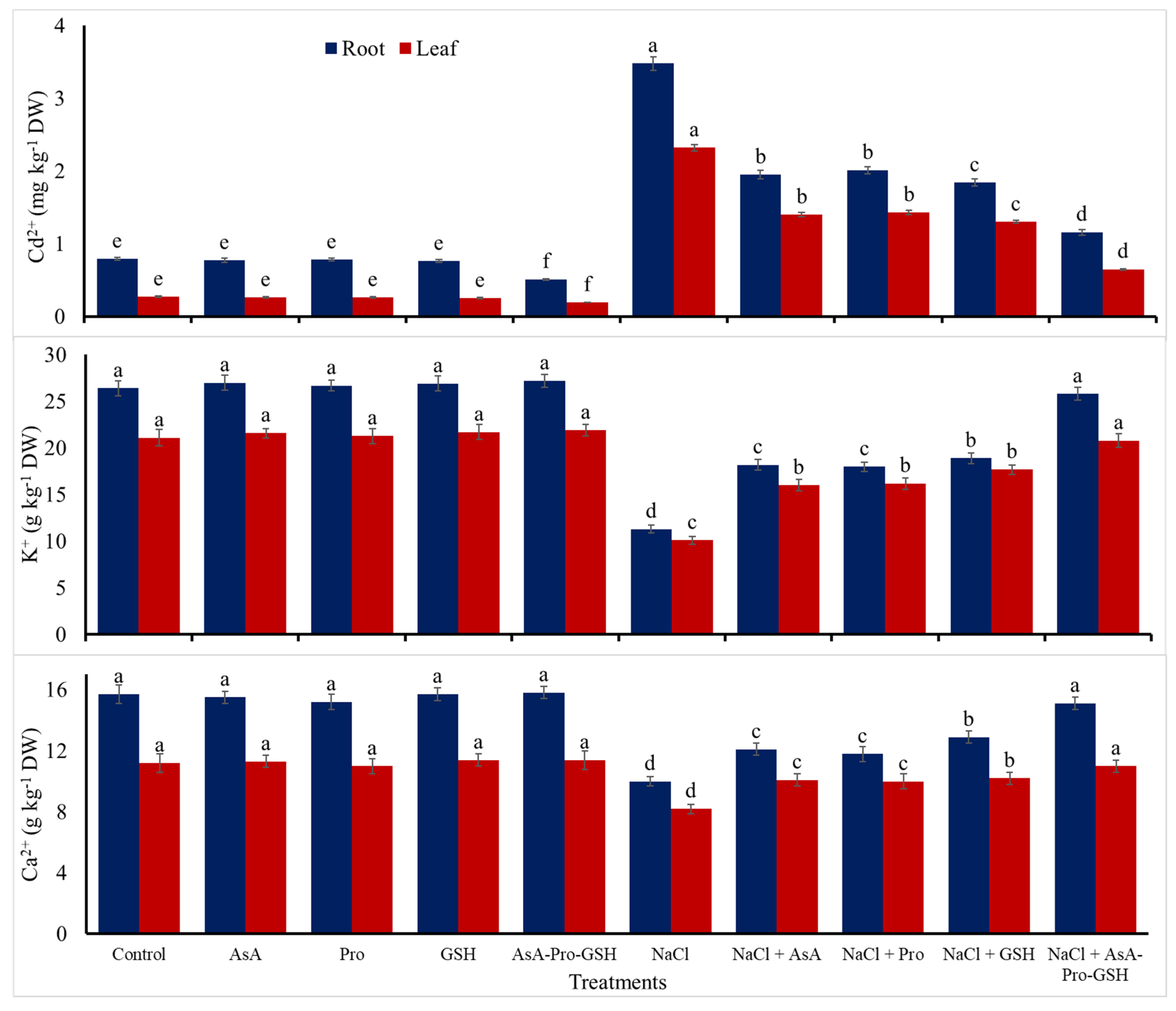
| Cd2+ content | 0.53 | −31 | −2.5 | −5.6 | -33 | 550 | 283 | 292 | 257 | 94 |
| K+ content | 23.8 | 2.3 | 1.0 | 2.4 | 3.4 | −55 | −28 | −28 | −22 | −1.9 |
| Ca2+ content | 13.5 | −0.4 | -2.5 | 0.9 | 1.2 | −31 | -16 | −18 | −13 | −2.8 |
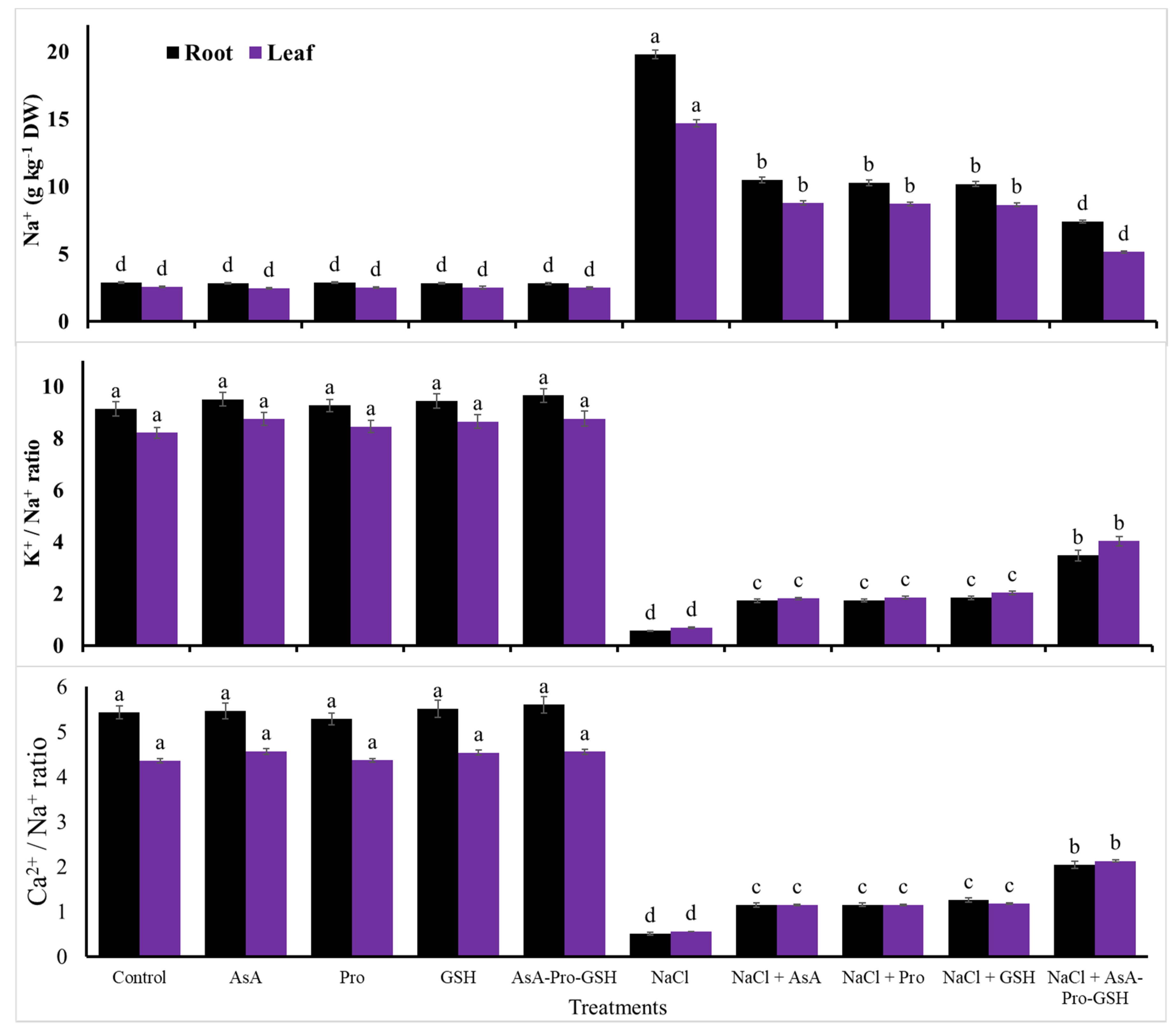
| Na+ content | 2.73 | −2.8 | −1.1 | −1.9 | -2.5 | 529 | 253 | 248 | 245 | 129 |
| K+/Na+ ratio | 8.67 | 5.4 | 2.2 | 4.4 | 6.2 | −93 | −80 | −79 | −77 | −57 |
| Ca2+/Na+ ratio | 4.90 | 2.7 | −2.6 | 2.8 | 3.9 | −89 | −77 | −76 | −75 | −56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seleiman, M.F.; Semida, W.M.; Rady, M.M.; Mohamed, G.F.; Hemida, K.A.; Alhammad, B.A.; Hassan, M.M.; Shami, A. Sequential Application of Antioxidants Rectifies Ion Imbalance and Strengthens Antioxidant Systems in Salt-Stressed Cucumber. Plants 2020, 9, 1783. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9121783
Seleiman MF, Semida WM, Rady MM, Mohamed GF, Hemida KA, Alhammad BA, Hassan MM, Shami A. Sequential Application of Antioxidants Rectifies Ion Imbalance and Strengthens Antioxidant Systems in Salt-Stressed Cucumber. Plants. 2020; 9(12):1783. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9121783
Chicago/Turabian StyleSeleiman, Mahmoud F., Wael M. Semida, Mostafa M. Rady, Gamal F. Mohamed, Khaulood A. Hemida, Bushra Ahmed Alhammad, Mohamed M. Hassan, and Ashwag Shami. 2020. "Sequential Application of Antioxidants Rectifies Ion Imbalance and Strengthens Antioxidant Systems in Salt-Stressed Cucumber" Plants 9, no. 12: 1783. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9121783